Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Proteolysis wikipedia , lookup
Mitochondrion wikipedia , lookup
Photosynthetic reaction centre wikipedia , lookup
Electron transport chain wikipedia , lookup
Nicotinamide adenine dinucleotide wikipedia , lookup
Photosynthesis wikipedia , lookup
NADH:ubiquinone oxidoreductase (H+-translocating) wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Butyric acid wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Microbial metabolism wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Metalloprotein wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Biosynthesis wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Biochemistry wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
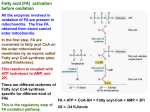
The process of beta oxidation is named after the carbon atom in the beta position of the fatty acyl-CoA which becomes the most oxidized during the cyclic redox reactions that remove C2 units in form of acetyl-CoA from the fatty acyl chain. The beta carbon becomes the new carboxyl end of the shortened (n-2) fatty acyl-CoA. The oxidation steps are strictly analogous to the reaction steps in the citric acid cycle converting succinyl-CoA to oxaloacetate involving an initial oxidation by acyl-CoA dehydrogenase (EC 1.3.99.3; driven by FAD reduction), an hydration by enoyl-CoA hydratase (EC 4.2.1.17), and a second oxidation by hydroxyacyl-CoA dehydrogenase (EC 1.1.1.35 driven by NAD+ reduction). A C2 unit is released by beta-ketothiolase (EC 2.3.1.16) to produce acetyl-CoA and a shortened acyl(n-2)-CoA. The latter is recycled until the acyl chain is shortened to its acetyl-CoA end product and oxidized by the citric acid cycle enzymes. The acyl-CoA dehydrogenase is specific for the length of the acyl chain being oxidized. Three types of the dehydrogenase exist in mitochondria; type I (EC 1.3.99.12; long chain) which oxidizes C12-C18 fatty acids, type II (EC 1.3.99.3) which oxidizes C4-C14 fatty acids, and type III (EC 1.3.99.2; butyryl dehydrogenase) which oxidizes C4 and C6 acyl-CoA substrates. The energy yield per cycle is 5 mols of ATP for each round, 2 mols per FADH2 (goes into complex II) and 3 mols per NADH/H+ (goes into complex I). The energy balance for palmitic acid (sixteen carbon atoms) is: CH3(CH2)14CO-S-CoA + 7H2O +7CoA + 7FAD + 7NAD+ 8CH3-CO-S-CoA + 7FADH2 + 7NADH + 7H+ The completion of the degradation process (coenzyme oxidation) requires the citric acid cycle which yields an additional 96 mols of ATP for all 8 acetyl-CoA units oxidized in the process. The total energy yield of palmitic acid oxidation results in some 130 mols of ATP, 34 units from the beta-oxidation cycle and 96 form the citric acid cycle. Beta oxidation of odd numbered fatty acids yields a (C2) acetyl-CoA and (C3) propionyl-CoA end product during the very last cycle. Propionyl-CoA cannot be further oxidized as such and is converted to methyl-malonyl-CoA by propionyl carboxylase (EC 4.1.1.41). This is an energy consuming step using 1 mol of ATP per mol of propionyl-CoA. The net reaction is: propionyl-CoA + CO2 + ATP = (D/L)-methylmalonyl-CoA + ADP + Pi The methylmalonyl is formed as racemic mixture containing equal amounts of the Dand L-enantiomer. The (D)methylmalonyl-CoA is isomerized to succinyl-CoA by methylmalonyl CoA mutase: (D)-methylmalonyl-CoA = succinyl-CoA Note that this reaction is also used for the degradation of hydrocarbon side chains of the amino acids valine, leucine, and isoleucine; (S)-methylmalonyl is the same as Lmethylmalonyl; from KEGG pathway MAP00280) Similarly, unsaturated fatty acids need special enzymes to provide the beta oxidation intermediate trans-D2-enoyl-CoA, the substrate of enoyl-CoA hydratase. Desaturation can result in two unusual degradation intermediates: the cis-3- and the cis-4-enoyl-CoA. The beta oxidation intermediate, however is a trans-2 isomer. The cis-3-enoyl-CoA intermediate is isomerized to the trans-2 form by isomerase (EC 5.1.2.3). The cis-4-enoyl-CoA form is converted in two steps to cis-3-enoylCoA intermediate. The first step is catalyzed by acyl-CoA dehydrogenase (the first enzyme in the beta oxidation cycle) to form 2,4-dienoyl-CoA which in turn is isomerized by 2,4-dienoyl-CoA reductase to cis-3-enoyl-CoA.