Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Neuroregeneration wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Adult neurogenesis wikipedia , lookup
Metastability in the brain wikipedia , lookup
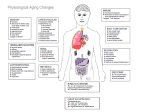
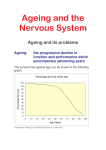
Aging brain wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Development of the nervous system wikipedia , lookup
Optogenetics wikipedia , lookup
Haemodynamic response wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Subventricular zone wikipedia , lookup
1 Boyko O. G. 2007. Differentiation of radial glia cells into astrocytes is a possible ageing mechanism in mammals \\ Zhurnal Obshchei Biologii (Journal of General Biology). V. 68 № 1 P. 35-51 Differentiation of radial glia cells into astrocytes is a possible ageing mechanism in mammals O. G. Boyko National Agrarian University ul. Geroyev oborony 15, Kiev 03041, Ukraine e-mail: [email protected] Several obscure facts of gerontology are briefly reviewed. The attempt is made to shape new notions of the phenomenon based on the astrocyte hypothesis of ageing in mammals. This hypothesis interprets mammal ageing as a genetic disease with fatal outcome. The disease is caused by single character acquired by the theriomorph lineage of the vertebrates in the course of evolution: the transformation of radial glia (RG) cells into star-shaped astrocytes during the postnatal development, i.e. the disappearance of the fetal radial ways of nerve cell migration from proliferative zones to the sites of their ultimate localization in the brain of adult individuals. This process is the cause for the mammal brain being postmitotic. The disappearance of RG cell induces a cascade of system processes termed age-dependent mechanism of self-destruction of mammals (AMSM). The disappearance of RG cell inhibits the replacement of the nerve cells that have exhausted their living resources. Nerve cells are rigidly specialized and have restricted lifetime and ability of reparation. After some period, the level of homeostasis in nerve cells starts changing steadily for the worse due to irreversible pathological changes in the cells (especially in the neurosecretory cells). This brings damage to lifesupport systems of the mammal organism thus causing its death. The species-specific maximum life span is thus determined by the rate of metabolism in the organism. AMSM probably displays a general evolutionary principle: outer factors causing death (in non-ageing organisms) are replaced by inner factors. 2 Till now the discussion started still in days of A. Weismann about an acceptability of two concepts of ageing is not stopped: death as result of an accumulation of casual breakages or starting of the program of "suicide". In a number of reviews (Khalyavkin, Yashin, 2003) a conclusion is made that even such standard concepts as free-radical and telomere – telomerase hypotheses are not quite suitable for an explanation of an original cause of ageing. Williams (Williams, 1957) summarized arguments against the idea of programmed death: in the nature practically all animals die at rather young age from the casual reasons. In other words - if there is a mechanism of programmed death it cannot define life-span at overwhelming number of individuals of a population; searches of the mechanism of programmed death were not successful; it is difficult to imagine how such attribute (the program of death) could arise during natural selection. Therefore today a number of known gerontologists, for example spouse L.А. and N.S. Gavrilov (Gavrilov, Gavrilova, 2002), Weismann’s ideas (Weismann, 1889) that ageing and death, is «something secondary arisen during adaptation» (the theory of programmed death - PDT), call precisely "deceased". V.P. Skulachev (1999, 2001), trying to update PDT, has offered the phenoptosis, concept. The programmed death of an organism, or phenoptosis, is a self-liquidation of an individual in interests of a population, and also its last boundary of protection against epidemics and monsters with the broken genetic program. In view of these ideas the death from an old age is considered as one of many displays of phenoptosis, caused by starting of the program of self-liquidation at the individuals which have appeared useless or harmful for population. As if to concrete mechanisms of ageing the analysis of the literature allows to draw a conclusion, that the majority of authors, for example Johnson (Johnson et al., 1999), are sure, that it is possible to reduce a phenomenon of ageing to a small number of cellular and all organism processes, such as programmed cellular death or apoptosis, accumulation of damages by molecules mitochondrial DNA (mtDNA), shortening of telomeres at mitosis, defects of a cellular cycle, etc. V.P. Skulachev allocates a key role during ageing to apoptosis. His arguments seem convincing, however they have not made due impression on other researchers. Objections cause representations about a parity and interdependence of phenoptosis, organoptosis and apoptosis, the chain of events is more exact than the programmed destruction of separate bodies: The programmed destruction of mitochondria or mitoptosis, - apoptosis, - organoptosis - 3 phenoptosis, (Skulachev, 1999) as it is obvious, that relations between these processes is much more complex and are poorly predicted (Vanyushin, 2001; Kaluev, 2003). The impression is created, that the facts mentioned by V.P. Skulachev have individual character, frequently they are discussible enough and can be treated differently depending on belief of the researcher (Kaluev, 2003). THE REASON OF DISCREPANCY OF KNOWN THEORIES OF AGEING Discussing this or that theory of ageing, authors, as a rule, miss existence of species of multicellular organisms which actually are not subject to ageing and possess potential immortality with that do not interfere generation of reactive oxygen species, telomeres shortening, apoptosis and other processes with which ageing usually explain. Moreover, these species have long evolutionary history, and their evolutionary progenitors in most cases also were potentially immortal. Ignoring of these fundamental facts of gerontology, probably, has very simple reason - researchers cannot explain them. Unfortunately, uniform classification of processes of ageing does not exist yet, and potential immortality is proved only for a hydra (Cnidaria vulgaris) (Mart´inez, 1998). Caleb Finch (Finch, 1990) proposes an original typology of senescence organisms from rapid to gradual to negligible. He introduced the term "negligible senescence" to mean "senescence too slight to be statistically distinguishable from zero with the sample sizes at our disposal". That includes non-senescence, of course -- truly zero correlation between age and risk of death. The minimal criteria are offered (Finch, Austad, 2001) for reference of a concrete species in this category: (1) no observable increase in age-specific mortality rate; (2) no decrease in reproduction rate after sexual maturity; and (3) no observable age-related decline in physiological capacity or (4) disease resistance. Such subjects should grow old so slowly that to fix age changes would be practically impossible. Except for a fresh-water hydra to them carry, for example: a lot of species of a genus Rockfish (Sebastes) (Leaman, Beamish, 1984; Cailliet et al., 2001; Munk, 2001); fresh-water Blanding's Turtle (Emydoidea blandingii); a painted turtle (Chrysemys picta); a significant part of wood forms of vascular plants (Finch, Austad, 2001); and also sea-urchin sea who can live vaguely long, being reproduce at any age, and the is more senior, the more actively (Ebert, Southon, 2003). Patnaik (Patnaik, 1994) includes into species with negligible senescence to the crocodiles and (Kara, 1994) - tailed amphibians rather plausibly, that 4 species with negligible senescence are more numerous. It is possible that in the near future in this category will carry many species of bivalve, lobsters (Homarus) and birds. SPECIFIC ATTRIBUTES OF "IMMORTAL" ORGANISMS The hydra (Cnidaria vulgaris) concerns to one of most close situated at the root basis of a phylogenetic tree of group of multicellular animals: - coelenterates. Brien (Brien, 1953) informed for the first time about potential immortality of this organism and the fact of potential immortality has proved by (Mart´inez, 1998). The hydra has a simple structure: - a tube generated from approximately 20 cellular types, being products of differentiation of three stem cell (SC) lineage with unlimited ability to self-updating: two epithelial and one interstitial (Campbell, 1965, 1967a). Amazing feature of a hydra - dynamics of a tissue. Epithelial cells of a wall of a body continuously stay in mitotic cycle and as though flow from the middle of a body by his ends, constantly moving or upwards to a head, or basal to vegetative buds, or to a bottom. Cells of a hydra have very short life span. Not dividing differentiated cells are replaced in a body of a hydra within 20 days (Campbell, 1967b). Thus, the specialized cells carry out strictly certain function, and have the limited viability. Age-related changes in these cells, capable to affect functioning of an organism, are prevented by their fast change with newly generated cells. Probably, this ability also is the reason of potential immortality of a hydra. Nevertheless, varying external conditions, it is possible to achieve ageing and from a hydra (Comfort, 1979; Lamb, 1980). At adverse change of temperature or structure of water cell fission is slowed down, the hydra grows old and perishes, i.e. depending on conditions of environment potentially not growing old organism can be in growing old or in not growing old condition (Khalyavkin, 1998), that depends exclusively on the external reasons. Species of bony fishes. At fishes, mainly fast or gradual ageing reminding ageing of mammal is observed. Nevertheless, a number of species, for example genus Sebastes shows not ageing behavior (Guerin, 2004). At old individuals Danio rerio it is not revealed congestions of granules of lipofuscin in cells of a muscular tissue, the high level of activity of telomerase in cells of somatic tissue from embryo conditions at least till 93 years is observed. Authors explain these phenomena as at this fish growth and cellular proliferation occurs for life (Klapper et al., 1998; Kishi et al., 2003). 5 FROM IMMORTALITY TO SELF-DESTRUCTION. THE VECTOR OF EVOLUTION OF HERBACEOUS PLANT. At Gymnospermae and dicotyledons among metasperms the adult tree consists from secondary thickened stalk (trunk) with a secondary branching, a root, leaves, and reproductive organs, thus a trunk and a root - constant organs and exist for life. Other organs are short life span. The trunk and a root of a tree are formed primary and secondary meristems – tissues in which division of poorly differentiated cells generates the new specialized cells. They form leaves, lateral buds, flowers and other bodies, and also provide increase in the sizes of a tree. Activity of meristems is shown during all life of a plant. Therefore wood plants continuously produce and lose a vascular tissue, leaves and flowers and do not show attributes of ageing though rate of growth can be reduced with the years. The death of a tree occurs as result of achievement of the limiting sizes incompatible with viability or the external reasons: wreckers, shading, etc. Wood plants - the most long-living organisms. Maximal life span (MLS) of the majority of them does not exceed 700 years, but there are also such species, with MLS estimated in millenia: for example, a bristlecone pine (Pinus longaeva) - more than 4 thousand years; giant sequoia (Sequoia gigantea) - more than 2.5 thousand years; Fizroya cupressoides - about 2 thousand years; juniper (Juniperus turkestanica) - about 1.3 thousand years (Vaganov et al., 2000). As well clones as naturally arising, and regrafted (subinoculation) cultural wood plants, within millenia do not lose the juvenile qualities that gives to them features of boundless longevity. The clone of (Lomatia tasmanica), for example, has age of 43 thousand years (Lynch et al., 1998). On A. Takhtajan (1964, 1966, 1970), wood species - the most primitive within the limits of the taxons and all variety metasperms conducts the beginning from primitive wood forms. Early herbaceous plants were trees, which during the further evolution by a somatic reduction have given rise to bushes and to various grasses. Their propensity to neoteny allowed them during evolution to pass easily from primary immortal wood forms to annual or shortly living grassy forms which as a matter of fact are the wood sprouts which have received ability to reproduction. Grassy, especially annual plants, have serious advantages before trees. They have short life cycle, early maturing and as result - fast alternation of generations, that finally allows them faster evolve. On B.F. Vanyushin (2001), the destruction of a grassy plant is controllable by hormonal phenoptosis: at least monocarp plants so finish a life. Thus, the core of evolution of herbaceous plants went from potentially immortal 6 wood forms to short living grasses. This tendency is traced for the majority of families dicotyledons. TRANSFORMATION OF RADIAL GLIA CELLS IN ASTROCYTES – REASON FOR POSTMITOTICY OF THE MAMMAL BRAIN AND IT IS POSSIBLE, THEIR AGEING Among phylogenetic branches of vertebrates not growing old species occur at fishes, tailed amphibians, turtles, and it is possible, at birds. At the same time any not growing old theriomorphic animal species, neither in the paleontologic annals, nor among nowadays living mammal is unknown. At mammal MLS varies from 3 years at the mouse (Mus musculus) till 211 years at the Bowhead whales (Balaena mysticetus) (George, Bada, 1999). Thus ageing with the extremely similar pathophysiology is observed at all species (Lamberts et al., 1997). The arthritis, atherosclerosis, change of the hormonal status, menopause, osteoporosis, vascular abnormality, deleting of a teeth, graying and balding, a cataract, accumulation of intercellular collagen, increase in quantity of lipofuscin in cells occur at all investigated species. In the certain sites of a brain there are similar change of structures and functioning of neurons (Morrison, Hof, 1997). These facts allow to assume, that the reason of ageing has arisen somewhere at the beginning of mammal evolution. Among vertebrates postmitotic brain is the unique phenomenon that occurs only at mammal. Today there is no interpretation of physiological function of this attribute. Researches XVIII - XX centuries have shown an opportunity of reparative and physiological regeneration of a nervous tissue of fishes, amphibians and reptiles (Polezhaev, 1968; Barres, 1999). However the higher vertebrates- birds and mammal, apparently, have lost ability to regenerations of a nervous tissue. But physiological regeneration of a nervous tissue of higher vertebrates - a debatable question, especially concerning mammal. In a brain of adults mammal neurogenesis nevertheless takes place, but in strictly limited zones from which new neurons migrate and are incorporated also in strictly limited parts of a brain. These zones are rudiments of neurogenic zones of embryo brain. The largest of these germinal regions in the adult brain is the subventricular zone (SVZ), which lines the lateral walls of the lateral ventricles. Neural SC produce neuroblasts that migrate from the SVZ along a discrete pathway, the rostral migratory 7 stream, into the olfactory bulb where they form mature neurons involved in the sense of smell. The subgranular layer (SGL) of the hippocampal dentate gyrus is another neurogenic region; new SGL neurons migrate only a short distance and differentiate into hippocampal granule cells (Lennington et al., 2003). Due to these facts the opinion prevails, that except for olfactory bulbs and hippocampus, new neurons normally are not updated in a brain of adult mammal individuals (Eriksson et al., 1998; Barres, 1999; Gould et al., 2001). Long time was considered, that birds have a postmitotic brain, but (Goldman, Nottebohm, 1983; Paton, Nottebohm, 1984) show, that in a brain of birds neurons are produced in ventricular / subventricular zone where persisted neural SC into adulthood, and then migrate along fibers of RG on a long distance, mainly filling up a pool of neurons in a telencephalon (Goldman, 1998). It is shown also a presence of well advanced radial network in a brain of adult birds (Kalman et al., 1993). Therefore, the fact of participation of RG cells has been recognized during migration of newly generated neurons, and that RG is kept in those areas of an adult brain, where proceeds neurogenesis and migration of neurons (Goldman, Nottebohm, 1983; Alvarez-Buylla et al., 1987, 1990; Alvarez-Buylla, Kirn, 1997). These and similar facts became the basis of a hypothesis that RG cells are precursors of neurons both in embryogenesisе, and at adult vertebrates. Really, RG cells can be precursors of a part of neurons of a brain and participate in posttraumatic reparation of a spinal cord of amphibians and salamanders (Margotta et al., 1991), and also in a cortex of a brain of reptiles (Molowny et al., 1995; Font et al., 2001). Therefore is conventional, that at fishes, amphibians, reptiles and birds RG not only is kept, but also produced neurons up to the end of a life (Weissman et al., 2003). Peculiarity of neurogenesis and his relation with apoptosis features are found out from adult bony fishes (Zupanc, 1999). The brain of these fishes possesses enormous potential for the production of new neurons in the adult brain. So, at Apteronotus leptorhynchus during 2 h it is produced about 0.2 % of the total population of cells in the adult brain (Zupanc, Horschke, 1995). Quantitatively of mitotic active cells of a brain are distributed approximately so: 25 % from them are in the telencephalon, diencephalon, mesencephalon and rhombencephalon. The others of 75 % of such cells are located in proliferative zones of a cerebellum which represent remnants of embryonic proliferation zones (Zupanc et al., 8 1996). The conclusion has been made, that the cerebellum of adult bony fishes cerebellum during adulthood most closely resembles the mammalian cerebellum during its development shortly before and after birth. After migration in a place of final localization many of the newborn neurons remain alive for extremely long periods and, most likely, for the rest of the fish’s life (Zupanc et al., 1996, Ott et al., 1997). This long-term survival, together with the continuous production of new cells, leads to a permanent growth of the entire brain. While the body mass of the fish increases from 1 g to 16 g, the total number of brain cells approximately doubles from 5x107 up to 108 (Zupanc, Horschke, 1995). The ability of bony fishes, and probably all fishes, to generate new cells in the brain during adulthood is paralleled by their enormous potential to posttraumatic neuronal regeneration after injuries (Meyer et al., 1985; Waxman, Anderson, 1986; Stuermer et al., 1992; Zupanc, 1999). The path caused by the stab wound becomes gradually reduced over the period following the lesion, until, several weeks after the injury, it finally disappears. Signs of inflammation, cavitation and scarring do not occur (Zupanc, 1999). Restoration of a nervous tissue is reached by the cascade of processes, including a production and a direction of migration of new neurons to a place of damage by RG fibers (Zupanc, Clint, 2001). Presence of extensive zones of neurogenesis at adult individuals, the facts of radial migration and long-term existence (alive) of newly generated neurons are confirmed and for other species of bony fishes (Zupanc et al., 2005). As researchers could not find out pyknotic nuclei and establish the period during which the quantity of cells in CNS reduce, it was considered, that cell death had been thought to be absent from the brain of fish in stages beyond embryogenesis (Fox, Richardson, 1982; Crapon de Caprona, Fritzsch, 1983; Waxman, Anderson, 1985; Fine, 1989). But it have shown by researches (Soutschek, Zupanc, 1995, 1996), that in many brain areas, including the prepacemaker nucleus (Soutschek and Zupanc, 1995) and the cerebellum (Soutschek and Zupanc, 1996), a large number of cells undergo apoptosis. The density of apoptotic cells is significantly higher in places of final migration, than in the corresponding proliferative zones, implying that cell death may mainly be involved in the regulation of the cell number of the newborn cells after they have reached their target areas. On the basis of this fact the conclusion has been made, that, probably, one of functions of apoptosis - to eliminate new neurons which after arrival on a place of 9 final localization have failed to join in function chart of CNS1 (Soutschek, Zupanc, 1996). It is interesting, that with the years at bony fishes the quantity of apoptosis events on a mass unit of a nervous tissue decreases (an age-dependent reduction in apoptotic events). As a whole about half of young cells is exposed to apoptosis, and other part lives up to the end of a life of a fish, and the most part of the survived cells is differentiated in neurons (Zupanc, 2006). As well over a posttraumatic reparation of a nervous tissue at bony fishes dominates apoptosis though at once after damage a part of cells are exposed of necrosis, but in some hours apoptosis becomes prevailing, and it is possible, and unique type of cellular death. Therefore at bony fishes even at traumas of CNS for removal of the damaged cells the mechanism reminding embryo apoptosis of mammals is used. Although in the initial stages immediately following a lesion some cells may undergo necrosis, at longer post-lesioning survival times starting at a few hours, apoptosis is the predominant, and presumably even the exclusive, type of cell death. Thus, bony fish appear to use a mechanism for the removal of damaged cells after injuries in the central nervous system that resembles the apoptotic cell death occurring in mammals during embryonic development (Zupanc, 1999). In mammalian systems, necrosis is usually accompanied by an acute inflammatory response (Kerr et al., 1995). The inflammation triggers further necrotic events, thus gradually transforming the site of injury into large cavities devoid of cells (Zhang et al., 1997). These cavities are typically bordered by scars which act as mechanical and biochemical barriers preventing the ingrowth of nerve fibres and the migration of cells into the site of the lesion. In contrast, these negative side effects are characteristically absent in apoptosis. The use of this ‘clean’ type of cell death by bony fish in the process of regeneration may, therefore, at least partially explain their enormous regenerative capability. There is an opinion, it is likely that the ability of fish to replace neurons damaged through injury or disease is causally linked to their capacity to generate new cells in the uninjured brain. Since a pool of undifferentiated cells is continuously filled through normal proliferative activity, recruitment of these cells enables the brain to replace damaged cells, in the event of an injury, much faster than would be the case by just inducing cell proliferation after injuries. 1 Earlier, similar ideas about function of apoptosis in embryogenesis has offered by (Raff, 1992; Raff et al., 1993). 10 In addition, these facts testify also that the concept of phenoptosis, of V.P. Skulachev - the speculative abstraction which is starting with false preconditions as she assumes, that phenoptosis, is caused by apoptosis, in a hypothetical chain of events: mitoptosis - apoptosis- organoptosis - phenoptosis, (Skulachev, 1999). Against it that fact testifies, that in a brain of adult mammals surprisingly low level of apoptosis is observed and consequently the pool of neurons during their life decreases insignificantly (Morrison, Hof, 1997). In other words, it is obvious, that at mammal harmful age induced displays of apoptosis are not the reason of ageing, but consequence of postmitoticity of a brain. That is why struggle with apoptosis cannot lead to significant break in the field of increase in duration of a human life. At mammal during neurogenesis 80-90 % of neuron precursors in a brain cortex migrate along RG fibers which represent as though clues of a direction of this migration (Gupta et al., 2002). But as against those vertebrates at which neurogenesis at adult individuals it is observed, and RG cells are kept for life, at mammal after the period of neurons migration RG cells are transformed in stellate astrocytes (King, 1966; Zupanc, 1999), except for hippocampal dentate gyrus (Cameron et al., 1993). Here RG cells generate neurons directly or through transitive progenitor daughter cells (Seri et al., 2001). Moreover, in some structures of the mammalian brain, RG cells adapt to the local functional requirements and spatial conditions and transform into specialized astrocytic cell types (e.g., the Bergmann glial cells of the cerebellum, tanycytes of the hypothalamus, or Muller cells of the retina). These specialized cells retain the basic morphological, immunological, and biochemical features of RG cells. Thus, they may be considered as genealogical, morphological, and biochemical descendents of the transient fetal RG cells (Rakic, 2003). From the submitted facts it is easy to draw a conclusion, that presence or absence of RG cells in a brain of adults vertebrates the defines a level of neurogenesis and by that, probably, life duration. Therefore earlier in most general view the astrocytes hypothesis of ageing of mammal (Boyko, 2004), connecting practical absence of RG cells in a brain of adults mammal with the possible reason of their ageing has been put forward. DISCUSSION The essence of astrocytes hypotheses of mammal ageing is reduced to the proposal that in theriomorphic lineages of tetrapods RG cells disappear in a brain of 11 adult individuals, as defines postmitoticity of a mammal brain. In other words: neurons are rigidly specialized cells with limited reparative opportunities and service life (however, as well as all other specialized cells of an organism arising from SC by asymmetrical mitosis ). Disappearance of RG cells in the end of embryogenesis and in the beginning of postnatal period forbids replacement of neurons exhausting a vital resource Therefore after the certain period as the result of irreversible pathological changes in neurons (especially in hormone - synthesizing) a level of a homeostasis starts to change steadily and adversely that damages life-support systems of an organism by means of what and finally a life of an organism is stopped / interrupts (Boyko, 2004). As a matter of fact, in this case the question is not about ageing in classical understanding of this term, but about functioning of programmed mechanism of age depending self-destructions of mammal (AMSM). In a context of astrocyte hypotheses insignificantly, that RG cells though and in insignificant quantity, but nevertheless are kept as in a brain of mammal – the hippocampal dentate gyrus, (Cameron et al., 1993), and birds. Important that at mammal adults the embryonal radial network of neuron migration is destroyed completely, and at adult birds at relative small number of RG cells well advanced radial network is kept (Kalman et al., 1993), providing migration of newly produced neurons (Alvarez-Buylla, Kirn, 1997). The facts submitted in this review confirm V.P. Skulachev's opinion (1997) that the essence of ageing is reduced to dependent on age self-destruction of an organism or phenoptosis, but at the same time sharply contradict one of the main postulates of his concept of phenoptosis, that aging is a specific biological function that promotes the progressive evolution of sexually reproducing species. Differently, - ageing obligatory attribute of existence of multicellular organisms (metaphytes / metazoan). But, from mentioned above facts follows, that ageing is typical only for separate phylogenetic groups. In particular, among vertebrates ageing is observed of a number of exceptions only at mammal. However, absence of this process was not an obstacle for progressive evolution of other groups of vertebrates. The second serious mistake in the concept of V.P. Skulachev is a question on concrete mechanisms of this process. Reduction of principles to mechanisms - the widespread methodological mistake frequently meeting in biology. But, V.P. Skulachev (1997, 1999, 2001), apparently, has committed unique in own way a mistake, – ignoring the strict scientific facts, has erected the mechanism of the programmed death of a cell 12 (apoptosis) in certain «apoptotic credo», recognizing that if apoptosis carries out function of phenoptosis, at procaryotes and unicellular eukaryotes and at multicellular organisms this process, is ostensibly an original cause of their ageing. But, as is known, the facts - a thing obstinate. With occurrence of multicellular state, the first multicellular organisms as though get rid of the program of self-destruction and receive potential immortality. In these organisms the cell from a self-sufficient organism turns to a part of complex system. However, it receives in the inheritance from unicellular progenitors the program of apoptosis. This mechanism in a condition to destroy only this or that cell of an organism, but not the organism as a whole and practically does not limit life expectancy, as follows from the above described example with a fresh-water hydra. (However, the same is observed and at more advanced forms, for example, bony fishes). Thus, at the beginning of evolution the nature created immortal organisms. Therefore, ageing is typical only of the limited quantity of multicellular forms, and is later evolutionary adaptation. According the astrocytes hypotheses ageing among vertebrates arises only at mammal by purchase of such attribute, as almost full disappearance of RG cells in the beginning of postnatal period. It seems, it is possible to offer four possible variants of these evolutionary events: 1. By-product of evolution of the mechanism of restriction of growth of vertebrates (Bidder, 1932) of theriomorphic lineage. 2. By-product of evolution of the mechanism of postnatal developments of mammals (Dilman, 1958; Dilman, 1971, 1981). 3. AMSM has arisen as the mechanism of age-depending suicides under script of Bowles (Bowles, 1998, 2000). 4. Result of the further evolution of the mechanism of restriction growth of theriomorph in the multitask mechanism Finding-out of the reasons of evolutionary disappearance of RG cells in a brain of adults mammal causes interest also because this event in evolutionary prospect, apparently, initiated «physiological revolution». Therefore each of the submitted scripts deserves separate and detailed discussion. The mechanism of age-depending self-destructions of mammals, as by-product of evolution of the mechanism of restriction of growth theriomorph Bidder (Bidder, 1932) assumed, that ageing is a by-product of evolution of the mechanism of restriction 13 of growth nonaqueous vertebrates, i.e. «Ageing is a result of proceeding action of a regulator after the termination of growth» and as fishes have no mechanism of restriction of growth, hence, they should not have also mechanisms of ageing. (In other words, Bidder proposed that cessation of growth leads to the onset of aging). Really, the modern investigations in any measure confirm this conclusion: a number of species of fishes shows negligible ageing. Truly as well that the sizes of a body of tetrapods are limited by mechanical and other factors. Therefore if not all tetrapods then, birds and mammal have the limited growth. Reznick (Reznick et al., 2001) tries to deny the Bidder’s hypothesis offering in exchange idea about prolongated ageing for species with unlimited growth. But even he ascertained, that at short living species of fishes – guppies for which the early maturity and very high rate of death from the external reasons are characteristic, early attributes of decrease in reproductive potential are not observed as that was expected by researchers (Reznick et al., 2004). The submitted above facts show, that at fishes neurogenesis proceeds outside of embryo stages of development, providing continuous growth of a brain and his posttraumatic reparation. Two key factors admitting this phenomenon, - elimination of the damaged or unnecessary cells of a brain with apoptosis and replacement of the lost cells with newly produced cells alongside with constant updating of a pool of nervous cells as a whole. These processes are provided, in particular, with RG presence at adult individuals. As in a brain of vertebrates zones of neuron’s proliferation and their final localization are disconnected, disappearance of radial ways of neuron’s migration can stop growth at least of a brain. To assert, that RG disappearance at mammal deprives brain SC and by that stops the proliferation and growth of a nervous tissue, probably will be not absolutely correct, as SC SVZ at adults mammal are identified as astrocytes (Doetsch et al., 1999; Alvarez-Buylla et al., 2001). But, whether can a phenomenon of RG cells transformation to astrocytes limits growth of a body? I believe, that the following stage in this cascade of processes - a brain with the limited growth limits growth of a body as a whole. If to analyze classical representations about postnatal development, for example for humans, as soon after a birth transformation of RG cells in astrocytes occurs and the termination of processes of neurogenesis and neurons migrations2 . Further there is a 2 Probably, discussion about specification of terms of the termination of these processes as in a number of reviews, for example I.V. Viktorov (2001) affirms already began, that during the period from 15 months till 6 years there is a double increase in number of cortex’s neurons, i.e. postnatal neurogenesis in a brain of humans 14 growth of an organism up to species-specific size. It seems that two reasons can suspend growth in such conditions A number of facts specifies that, probably, at fishes postembryonic neurogenesis is connected to process of unlimited growth of a body. At mammal with their limited growth of a body, during postnatal developments, muscle growth is based on an increase in size, but not in number, of muscle fibres (Rowe, Goldspink, 1969). In contrast, in fish the formation of new muscle fibres continues well into adult life (Koumans, Akster, 1995). Therefore it is possible that the increase in the number of peripheral motor elements prompts a concomitant increase in the number of central neuronal elements (CNE) involved in the physiological control of the associated muscle activity. Opportunities of postmitotic brain do not allow to increase CNE quantity thus, probably, limiting growth of muscles. Moreover, the number of sensory receptor cells, receptor organs or receptor units in the periphery has been shown to increase with age in several species of fish. Such a postembryonic production has been demonstrated, for example, for sensory hair cells in the inner ear of sharks (Corwin, 1981), for retinal cells in the eye of goldfish (Johns, Easter, 1977), etc. Thus, as described for motor systems, the continuous increase in the number of sensory structures in fish may necessitate the generation of additional central neurons to process the associated sensory information (Zupanc, 1997). In mammals, in contrast, the production of sensory cells appears to cease by the end of gestation (Ruben, 1967). At mammal soon after a birth neurogenesis stops and the quantity of neurons also is stabilized including hormone synthesizing, but the quantity of hormoneproducing cells in other part of a body in process of juvenile growth increases for the size measured by degrees. As neuron secretion and hormonal secretion in other part of body are connected in difficultly adjustable cybernetic loop, at the certain quantitative parity of neuron producing cells to the rests hormone-producing cells, probably, such hormonal balance is established at which not only growth stops, but adverse changes can occur3. The mechanism of age-depending self-destructions of mammal, as by-product of evolution of the mechanism of postnatal developments. V.M. Dilman’s script. It proceeds at least till 6 years. But, even if it and so it at all does not change essence of an affair. The brain earlier or later to become practically postmitotic. 3 The idea, that the reason of mammal’s ageing consists in lagging behind of a gain of weight of a brain from a gain of weight of a body in development, belongs to S.S. Schwarz (1976). 15 seems, that consequences of disappearance of RG cells at adults mammal are well described in elevation theories of ageing of V.M. Dilman (Dilman, 1958; Dilman, 1971, 1981). He has supposed that if the stability of internal environment of organism is a sine qua non condition of its existence, a programmed deviation of homeostasis is a condition of its development (law of deviation of homeostasis). In other words, Dilman postulates an age dependent mechanism of postnatal juvenile development of organism based on the changing of hypothalamus sensitivity as a response to feedback signals. However during the reproductive period it enters to pathological phase and its further functioning leads to adverse changes of a homeostasis. Sensitivity of hypothalamus to brake influence of the factors signaling by a principle of a negative feedback about a condition of three basic homeostatic systems - power, reproductive and adaptable is reduced. Therefore with the years levels of cholesterol, insulin and cortisone in blood, for example, increase, tolerance of an organism to glucose is reduced, and also there are other adverse changes. Infringements of a homeostasis lead to development of characteristic diseases for advanced age: obesity, diabetes, atherosclerosis, cancrophilia, depression, hypertensions, metabolic immunodepression, hyperadiposis, autoimmune diseases and climax (V.M. Dilman considered a climax as age disease). In other words, ageing is considered as occurrence dependent on age and development of these diseases, and as a whole all is reduced to age decrease (it agrees other data, changes with the years, Frolkis, et al., 1991) an ability of hypothalamus to catch signals from periphery that is caused by decrease in quantity of hormonal receptors on a surface of hypothalamus’s neurons. But even if a number of conclusions of elevation theories of ageing also appeared false, that, as a whole it does not put under doubt the postulated by V.M. Dilman (Dilman, 1986; Dilman, 1987) ontogenetic model of ageing: а) performance of the program of development of an organism demands the programmed infringement of a homeostasis; б) at mammal change of a threshold of hypothalamus sensitivity to regulating signals causes necessary infringement of a homeostasis, that plays a key role in juvenile development; в) preservation of this mechanism after end of development causes transformation of the program of development in the mechanism of ageing and the main illnesses linked to ageing. But, what is the molecular basis of the process leading to so sad changes in an organism of mammal? There is an opinion, that mechanisms of cellular ageing are independent on external signals and processes of organism and cellular ageing can coexist or serve as the trigger one to another (Johnson et al., 1999). Arguing thus, a 16 number of researchers offer two hypothetical scripts of events. The first is reduced to that ageing of multicellular organisms is determined by mitochondria’s of post mitotic cells and slowly updated tissues because of accumulation of the damaged molecules mtDNA (Litoshenko, Hartwig, 1998; Gracy et al., 1999). It is possible, that a postmitotic mammal’s brain - not only an excellent illustration of this model, but also a place where accumulation of mutant mtDNA is simply a function from time and speeds of a metabolism. Therefore in due course as a result of accumulation of mutant molecules of mtDNA the significant part of this pool (including hormone synthesizing neurons) has defects of oxidative phosphorylation. As the result, the production of ATP decreases and functioning and biogenesis of mitochondria are broken, and also the level of ROS production and accumulation of these damaged structures raises. The process is auto accelerative. Recognizing that change of energetic balance of neurons can change sensitivity of neurosecreting cells of a brain to hormonal stimulus can lead to change of a level of neurosecretion and quantities of hormonal receptors on a surface of these neurons. If including this scheme in ontogenetic model of ageing of V. M. Dilman, the accumulation of the damaged molecules of mtDNA in hormone synthesizing neurons can be considered as the starting trigger and internal clocks of juvenile developments and ageing (Boyko, 2000). The second script has offered by A.M. Olovnikov (2003), trying in a context of modern representations about a role of telomere to explain ageing of postmitotic neurons and, in particular, neuroendocrinal cells. In fact they not exposed to proliferation and consequently the process of underreplication DNA on the ends of telomeres is not applicable to them, but basically underrepair of DNA is possible (Olovnikov, 1971, 1992). Process of incomplete copying of the end of DNA proves both in dividing, and in postmitotic cells owing to, that the DNA polymerase of repair synthesis, as well as DNA-polymerase of replicative synthesis, cannot fill in an end gap on 5´-overhang of a molecule in view of absence the free 3´-overhang which is capable to increase DNA- polymerase of reparative synthesis. Therefore all those factors (for example, free radicals) which conduct to occurrence of corresponding end gaps, also inevitably conduct to shortening of the ends of a double helix of DNA even in postmitotic cells. Excessive telomeres shortening can conduct to alterations in neuroendocrinal cells with similar to above mentioned outcome. It is necessary to pay attention that from the data received in last years, contours 17 of the scheme of hormonal regulation of juvenile growth at mammal start to appear. Ideas testify to it and an extensive material submitted in the review (Daftary, Gore, 2005) in which the known facts that insulin-like growth factor 1 (IGF-1) is the integrated hormone connecting juvenile growth and maturing of reproductive system are persuasively interpreted, for example. But in this review (however, as well as in other known literature) does not have the analysis of quantitative parities (magnitude relation) between producers of neuroendocrine hormones and producers of hormones of other part of a body. In view of this important parameter, (Daftary, Gore, 2005), it is possible to reduce interpretation of the facts stated in the review to more convincing scheme. 1. Juvenile growth - the complex process demanding coordinated action of many hormones, but a key role plays pituitary’s somatotropin (growth hormone, GH). I.e. the certain concentration or quantity of circulating GH is necessary for growth 2. GH is a component of a hormonal axis in which two hypothalamic releasing factors are included: growth hormone-releasing hormone (GHRH) causing secretion of GH by a hypophysis and somatostatin [SS] which inhibits it. Actually GH of hypophysis, inducing the secretion of IGF-1 by tissues of internals, mainly by liver, and also a gastric hormone ghrelin which also stimulates the production of GH. Thus SS is synthesized, except for hypothalamus as well by tissues of internals and can similarly to IGF-1 penetrate through hematoencephalic barrier. GH, GHRH, SS and IGF-1 are connected with homeostatic relations functioning by a principle of a negative feedback. The high level of circulating IGF-1 causes decrease in GH secretion as by inhibition of somatotropic cells of a hypophysis, and by stimulation of SS secretion by hypothalamus. 3. As the quantity of cells of body during the period of juvenile developments steadily grows at constant quantity of neurosecretory cells, accordingly during this period the total answer secretion of IGF-1 on GH increases and, accordingly, the concentration of circulating IGF-1. 4. IGF-1 stimulates a transcription of a gene of gonadotropin-releasing hormone (GnRH), that causes an increase of a production by hypothalamus and as the result the production of follicle-stimulating (FSH) and luteinizing (LH) hormones also increases. At the certain level of circulating IGF-1 the cascade of the events increasing a level of circulating sexual steroids is started. In other words, when the quantity of circulating IGF-1 reaches the certain critical values, pubertate period is initiated and the start of sexual function. Moreover IGF -1 lowers effects of a negative feedback of estrogens on 18 liberation of LH, that also approaches the beginning of a sexual maturity at female. There are data, that estrogens can regulate the IGF -1 expression In a phase of the beginning of a pubescence there are less clear processes as a result of which IGF-1 loses an influence on gene GH expression, sensitivity of tissues of a body to GH and IGF-1 is reduced, the growth stops, secretion of SS causing decrease of GH level (approximately, about 10 mg / dl of blood in 21 years up to 2 mg /dl of blood in 61 years for humans) and IGF-1 with the years increases. It defines in a result many symptoms of ageing. But in a context of the submitted scheme these processes should not cause surprise. Probably, in a phase of the beginning of a pubescence the parity of quantity of hormone producing neurons and others hormone producing cells is those, that such hormonal balance at which sensitivity of hypothalamus is modulated, and of some other tissues to hormones of axis GH/IGF-1 is established, as stops growth. Up to this point, really, such scheme is conformable to V.M. Dilman's ideas and connects juvenile growth and sexual development in uniform process. But with the years CNS, consisting of irreplaceable elements, it is doomed for destruction and, collapsing, will negatively change life supporting functions of an organism as a whole, probably, mainly through neuroendocrinal regulation. Probably there is a number of the reasons and the factors, capable to modulate this process from accumulation of mutant mtDNA to changes of a climate and mood of the favorite mother-in-law. But at the same time, such postmitotic CNS in itself can be the factor (probably, the key factor) of postnatal juvenile developments of an mammal organism, that also is conformable to V.M. Dilman's ideas. The mechanism of age-depending mammal’s self-destructions could arise and as the mechanism of age-depending self-destructions and as the multitask mechanism. On (Bowles, 1998), genetic drift in the finite populations leads to homozygosity, and selection changes a vector of drift in alleles which increase viability of this or that phenotype depending on a direction of the selection pressure. Ageing increases population turnover, which accelerates the benefit of genetic drift. This advantage of aging led to the evolution of aging systems. Periodic predation was the most prevalent episodic selection pressure in evolution. In evolution effective protection against predators allows to develop exceptionally long life span. Without episodic predation, aging provides no advantage and aging systems will be deactivated to 19 increase reproductive potential. The periodic advantage of aging led to the periodic evolution of aging systems. From this point of view it is supposed, that postulate of Medawar: - if in natural conditions the majority of organisms perishes earlier, than has time to grow old, the mechanism of ageing has no practical value for a species and consequently could not be selected by evolution (Medawar, 1952), is inapplicable to many species during their certain periods of phylogenesis (Bowles, 2000). Almost all researchers are agree upon the existence of two independent evolutionary branches of tetrapods - theriomorphic animals, crowned by mammals, and sauromorphic ones, progenitors of dinosaurs and birds, have gone away at a level of amphibians. Both groups have arisen simultaneously, in later Carboniferous. On S.V. Savelyev (2005), environment in which mammal have arisen, was very complex, dynamically changeable and rather aggressive. It is supposed, that their ancestors, having left carboniferously vegetative blockages, went to dwelling on branches of trees. Such pattern of existence, probably, rigidly defined fixed species-specific sizes of a body. As the first 2/3 evolutionary histories mammal stayed in a stage of a small dimensional class, that, probably, both the environment of existence, and the significant period of evolutionary time promoted evolution of the mechanism limiting unlimited growth. Moreover, the small dimensional class in most cases also per se assumes, short life span (Rougier, Novacek, 1998) and fast maturing. Therefore the mechanism limiting growth, but causing ageing, probably, could arise as a by-effect, as ageing in the given script is an neutral attribute. The alternative point of view can be proved by that early mammal in view of the sizes could be object of carnivorism that, on (Bowles, 1998), induces development of mechanisms of ageing. And transformation of RG cells in astrocytes just also could provide this animals with fast development, restriction of growth and life span simultaneously. At all stages of mammal evolution there were "returns" to ground forms of a life. One of the first «have gone down from trees» were monotremes, and then marsupials. Much later them have joined with eutherians (Savelyev, 2005). But as significant differences in display of a phenomenon of ageing of these groups are not present, it, probably, testifies that AМSМ was generated or at the beginning of evolutionary history of mammal, or even in reptile period of their evolution. If to believe, that AМSM is a by-product of occurrence and the further evolution of the mechanism of restriction of growth at mammal, it is not clear, why the similar mechanism generated in a parallel evolutionary of sauromorphic lineage has more 20 "humane" character. These facts allow to assume, that AМSМ could arise as the mechanism of life span restriction or as the derivative mechanism, but not as a byproduct of evolution. The same facts give ground for the assumption, that AМSМ has arisen during evolution as the multitask mechanism for maintenance of the accelerated maturing, restriction of growth and life span. INTERPRETATION OF THE KNOWN AND OBSCURE FACTS IN THE LIGHT OF THE ASTROCYTES HYPOTHESIS My problem as the author of astrocytes hypotheses of ageing of mammals at the given stage consists not only in putting forward one more hypothesis of ageing, but also in its approving; differently, to interpret in its light the obscure facts and phenomena of biology of ageing. On the other hand, such interpretations can be considered as indirect demonstrations (indirect proof) of astrocytes hypotheses. The facts from area of genetics. A number of researchers believe, that ageing at mammal supervises with very insignificant number of genes. For example, in A.I. Potapenko and A.P. Akif’ev's review (2003) it is affirmed that universality of dynamics of death rate and exact preservation of a spectrum of senile changes at mammal, at interspecific distinction in life span up to 50 times, and also growth of the average life span more than in 2 times in evolution from hominids up to humans also with preservation of a spectrum of senile changes, at change no more than 2 % of genes, is possible only if that work of the mechanism of ageing is defined with a small number of genes. According to astrocytes hypotheses, ageing at mammal is caused by transformation of RG cells after end of processing neurogenesis in astrocytes, that, obviously, does not demand significant reorganizations in genome: perhaps, changes in one gene only. The facts from area of comparative histology of astrocytes. Assumption of Ramon y Cajal that RG cells - precursors of astrocytes, has proved to be true (Levitt, Rakic 1980; Benjelloun-Touimi et al., 1985; Voight, 1989). RG - group of cells which is present at all groups of vertebrates: dominates over fishes and amphibians, but in the limited quantity meets at adult reptiles, birds, and mammal. The prevailing form of RG 21 cells at fishes and amphibious, - ependymal astrocytes 4, but already this form of RG can be found out in reptiles only in strictly certain areas of a brain (Suarez et al., 1995). Role of RG as precursor of glia cells, and also a significant part of neurons at high vertebrates (Alvarez-Buylla et al., 2001) is confirmed. The facts stated in the review (Suarez et al., 1995) persuasively testify, that evolution of astrocytes in a brain of vertebrates reminds their ontogenesis at high vertebrates: the first cells of astrocyte lineages – ependymal astrocytes, then arises RG cells - the precursor of astrocytes. Functionally RG at low vertebrates covers all functions attributed to astrocytes at high vertebrates. But if at lower vertebrates regional specialization of radial processes of RG cells is observed (Miller, Liuzzi, 1986) high vertebrates have a specialization of astrocytes. Actually transformation of RG cells in astrocytes at mammal- it is a deepening of specialization of RG with loss of a part of functions inherent to RG, including the guidance of migrating new neurons by radial glial fibers. In evolution of astrocytes among vertebrates reptiles are considered as key group, as it is possible to find out in them genuine astrocytes (Bodega et al., 1990) alongside with ependymal astrocytes and RG cells. Occurrence of astrocytes at reptiles is accompanied by reduction of relative quantity of RG cells that assumes cellular replacement. In a developing of mammal’s brain RG appears during embryogenesis time. RG cells form long radial processes which connect internal and external membranes of a nerve tube, forming ways of migration of neurons. In the end of embryogenesis and in early postnatal ontogenesis large transformation of fetal RG in astrocytes (Schmechel, Rakic, 1979) is observed. According to classical representations it occurs during the moment when RG cells lose touch with membranes of a nerve tube. Only those RG cells which lose touch both with internal, and with external membranes of a nerve tube (Voronova, Klimova, Mendgeritskiy, 2005) develop in astrocytes. In view of absence of the paleontologic data it is difficult to expect, that the comparative analysis of histology of a brain of modern vertebrates will give answers to all intriguing questions. But, probably, the submitted facts testify that the phenomenon of transformation of RG cells in astrocytes started to arise with reptile level of evolution of mammal that coincides with their initial small size and necessity of restriction of 4The term « radial glia » has been repeatedly entered by (Rakic, 1981), and in the literature frequently includes not only RG but, also ependymal astrocytes and isolated radial shoots. 22 growth and, probably, specifies the connection of these processes as that and is assumed with astrocyte hypothesis. The facts from area of transplantology. The studies on inbred and syngenetic mice have shown that skin flaps received from an old host can be transplanted in succession to several young ones. When the second host-mouse grew old, transplant was replaced to younger mouse etc. In such a way the cell viability and proliferative potential were saved during 7 - 8 years - a period similar to bird’s life span (for birds the corresponding sizes) and exceeding significantly the mice MLS. The same was observed and with a transplant of an epithelial tissue of mammary gland which kept viability within 6 years (Daniel, Young, 1971; Daniel et al., 1975). Following the ovary transplantation from old rats to young ovaryotomyzed ones, the estrous function is restored, the young animal ovary having perished in the old rat organism (Kushima et al., 1961; Aschheim, 1976). Exclusive interest is represented with messages that transplantation of the embryonal hypothalamus to old mice and rats restored their fertility, the immune status and a number of other attributes inherent in a young organism, and prolonged a life to experimental rodents (Ata-Muradova, Dontsov, 1987; Huang et al., 1987). [the fetal hypothalamic grafts can survive and develop in the brain of impotent aged male rats and restore neuroendorince and reproductive function in senescent rats (Huang et al., 1987) text editing] In other words, moving of a tissue or an organ from an old organism in homeostatic conditions of a young organism returns to this organ juvenile qualities, and restoration of a level, a homeostasis in a growing old organism up to a level, characteristic for young age, returns juvenile characteristics of an organism as a whole and extends life span. Usually similar facts are interpreted so, that many cells dividing in an organism have not time to exhaust completely for a human life Hayflick limit (Olovnikov, 2003). But in light of astrocyte hypotheses these facts specify that the potential longevity of the majority of somatic tissues of a mammal organism is comparable with those at birds, but his realization is interfered by functioning of AMSM. I.e. the postmitotic brain with the limited viability, collapsing, destroys the internal environment of an organism that finds acknowledgement in effects from the mentioned transplantation of fetal hypothalamus to old animal. Interpretation of experiments of G.M. Butenko. With the purpose to check the idea of programmed ageing blood systems of the old and young mouse were connected 23 to create heterochronic parabiont. Proceeding that if ageing occurs under the program in blood of old animal there should be factors of ageing, supplied with the program, and at unification of blood streams these substances have to proceed from old animal to young and it should grow old quickly. Really, in some weeks the young animal showed attributes of ageing. Senile changes were observed in immune and reproductive systems, a connective tissue, a skin and a liver and at organismic level, reducing a viability of young organism, connected to old one, that results in reduction of the young parabiont's life-span up to the life limit of his old partner. Actually, these researches have proved, that ageing occurs at a system level and in blood of old animal is unknown factor which blocks any genes or, on the contrary, starts it, as causes ageing (Butenko, 1990; Butenko, 1990). In light of astrocyte hypotheses of ageing of mammal there is anything outstanding in the experiments, the old animal as though shared the already achieved level of homeostatic anarchy or a hormonal background with younger, that causes senile changes and shortening the life span of younger animal. In other words, the level of a homeostasis, characteristic for old age, as though interfered in a young organism, making senile destructions before a time / prematurely. The fact of high potential longevity of birds in comparison with mammal of similar sizes and « an energetic law »of Rubner. Researchers agree that birds grow old more slowly, than mammal (Holmes, Austad, 1995; Holmes et al., 2001), but there are no checked up data on birds showing negligible ageing. Nevertheless among birds, species with very long MLS are usual, at insignificant age decrease in reproductive potential or even its increase. As a whole, except for gallinaceous birds (Galliformes) and some other species, birds show insignificant increase of death rate with the years (Ottinger, 2001). These facts are observed on a background of that MLS is much more for birds, than at mammal with comparable weight of a brain and a body, at higher level of a metabolism and a body temperature that is obscure from the point of view of existing theories of ageing. This tendency rather essential. For example, among animals with weight up to 30 g mammal have MLS from 2 up to 12 times smaller, than birds of the same weight of a body (Boyko, 2004). As the weight grows proportionally to linear sizes taken cubed, and the area - in squared, the organism of large warm-blooded, for example a whale, for maintenance of temperature from calculation on a mass unit of a body is required to less energy, than on "warming up" of an organism of the mouse, in view of disproportion of a parity of 24 weight and a surface of a body. Therefore at large mammal the level of a metabolism is much lower in comparison with small sized. Appeared, that large mammal live longer, than smalls. These law is based an "energetic" rule of a surface – hypotheses of ageing of (Rubner, 1908). The most interesting result of these researches appeared, that behind few exceptions representatives of various species of mammal during a life use approximately identical quantity of energy on a mass unit - 200 kcal/g. Therefore the high power consumption at the mouse leads to that she settles the power resources allocated to it faster, than the elephant, and term of its life is much shorter. Astrocyte hypothesis assumes the limited viability of post mitotic mammal’s brain. It has been shown above, that the vital potential of the majority of mammal tissues much more exceeds MLS of their organism as a whole. In other words, dying, the mammal’s brain kills other somatic tissues. Therefore, probably, these 200 kcal /g limit viability only of a mammal’s brain and specify the uniform mechanism of their ageing. It is obvious, that if viability of a brain is limited to volume of the used energy then evolutionary modulation of species-specific MLS is possible only by two ways: decrease in metabolic loading, i.e. increase in weight of a body, or repeated duplication of cells, which irreplaceable, i.e. increase in the sizes of a brain. And it is valid, energetic rule of Rubner has been confirmed in researches of Sacher on 63 species of mammal, but more severe dependence was observed in the event that the cefalization index, – the relation of weight of a brain to weight of a body in a degree 2/3 (Sacher, 1977, 1978) was considered. Comparison mammal with birds finds out surprising paradox. At birds MLS just as at mammal, keeps the tendency to increase with increase in the size. However MLS of birds is much higher (sometimes in the times), than at mammal of the same size, and total expenses of energy at birds during a life on a mass unit much more exceed calculated according a power rule of a surface, even in view of amendments of Sacher. According V.A. Gusev (2000), isolated mitochondria of a brain, a liver and hearts of the pigeon consume in 2 - 3 times more oxygen, than at rats, but thus mitochondria of the pigeon produce only 1/3 or hardly more than that volume of H2O2 that generate mitochondria of rats. Proceeding from this fact, V.A. Gusev (2000) explains big MLS of birds with their more perfect, than at mammal, mitochondrial processes. But on a background of the fact that MLS of the humming-bird (weight from 1.5 г), showing the higher level of a metabolism among warm-blooded vertebrates, reaches at some species till 15 years (Calder, 1989), such interpretation is unpersuasive/inconclusive. 25 Differently, the majority of species of the humming-bird consuming daily energy more, than by the average human (Best, 2006). It is a very simple calculation: in day the humming-bird consumes energy so much, how the human in weight of 70- 90 kg, thus it is produced in 10 times less ROS and H2O2, but, the humming-bird has weight 1.5- 3 г! Such rate of a metabolism can be compared unless to burning. It is obvious, if the humming-bird (however, as well as other birds) had the same rate of ROS and H2O2 production, as mammal they would poison with them. Obviously as well that as a whole the volume of ROS and H2O2 produced on a mass unit at the humming-bird is much more, than at mammal. From here there is a conclusion: there are no bases to assume, that at birds generation of oxidants by mitochondria is connected rigidly and directly with life span. Probably, the perfection of mitochondrial processes - not the reason of longevity of birds, but adaptation to record levels of a metabolism. Therefore this phenomenon submitted in the review of V.A. Gusev (2000), more persuasively is interpreted in light of astrocyte hypotheses: birds have huge potential of longevity because they have an opportunity of replacement of neurons exhausted their resource. CONCLUSIONS It is possible, that astrocyte hypothesis of ageing of mammal, can be essential argument for the benefit of idea of programmed ageing, at least for mammal, as transformation of RG cells in stellate astrocytes - the process certainly programmed. But it is impossible to exclude, that ageing of mammal has arisen as a by-product of evolution of other mechanisms. Programmed self-destruction of mammal, probably, is display of the general evolutionary principle - replacements of external factors of death of not growing old multicellular organisms on internal. From "meteorological" ageing of potentially immortal hydra it is clearly, that by variation of external conditions it is possible to achieve ageing of its organism, breaking process of replacement of the specialized somatic cells which have the finite resource. In other words, loss of this process at the organism consisting from potentially immortal SC and growing old specialized somatic cells, induces ageing and loss of potential immortality. Similarly, loss of the same process induces ageing and at mammal, but the difference is that this loss at mammal is genetically programmed and does not depend on external factors. With a number of essential clauses it is possible to recognize, that at grassy seed plants ageing is realized 26 by the same way - a reduction in phylogenesis an "immortal" part of a trunk which provides negligible ageing of wood plants. СПИСОК ЛИТЕРАТУРЫ Alvarez-Buylla A., Kirn J.R., 1997. Birth, migration, incorporation, and death of vocal control neurons in adult songbirds // J. Neurobiol. V. 33. P. 585-601. Alvarez-Buylla A., Buskirk D.R., Nottebohm F., 1987. Monoclonal antibody reveals radial glia in adult avian brain// J. Comp. Neurol. V. 264. P.159-170. Alvarez-Buylla A., Theelen M., Nottebohm F., 1990. Proliferation ‘hot spots’ in adult avian ventricular zone reveal radial cell division // Neuron. V.5. P. 101–109. Alvarez-Buylla A., Garcia-Verdugo J.M. Tramontin A.D., 2001. A unified hypothesis on the lineage of neural stem cells // Nat. Rev. Neurosci. V. 2. P. 287 - 293. Aschheim P., 1976. Aging in the hypothalamic-hypophyseal- ovarian axis in the rat // Hypothalamus, Pituitary and Aging /Eds Everitt A., Burges J.A. Springfield: Ch. C. Thomas. P. 376-418. Ata-Muradova F.A., Dontsov V.I., 1987. Effect of transplantation of the embryonal hypothalamus on the lymphoid tissue of old mice // Dokl Akad Nauk SSSR. V. 297. № 1. P. 237-40. (in Russian). Barres B.A., 1999. A new role for glia: generation of neurons! // Cell. V. 97. P. 667670. Benjelloun-Touimi S., Jacque C.M., Derer P., De Vitry F., Maunory R., Dupouey P., 1985. Evidence that mouse astrocytes may be derived from the radial glia. An immunohistochemical study of the cerebellum in the normal and reeler mouse // J. Neuroimmunol. V. 9. P. 87-97. Best B., 2006. Mechanisms of Aging / Рукопись монографии. http://www.benbest.com/lifeext/aging.html Bidder G.P., 1932. Senescence // Brit. Med. J. V. 2. P. 583-585. Bodega G., Suirez I., Rubio M., Fernandez B., 1990. Distribution and characteristics of the different astroglial cell types in the adult lizard (Lacerta lepzda) spinal cord // Anat. Embryol. V. 181. P. 567-575. Boyko O.G., 2000. Hypothetical method to overcoming barrier of maximum life span of mammals and humans. In:Proceedings of the 3th National Congress of Gerontologists and Geriatricians of Ukraine.2000 Sept. 26 - 28; Kyiv, Ukraine.P..170. (in Russian). 27 Boyko O.G., 2004. Do mammals die young!? An age-dependent mechanism of mammals self-destruction // Ukr. Bioorg. Acta. V.1-2. P. 3-12. Bowles J.T., 1998. The evolution of aging: a new approach to an old problem of biology // Med. Hypotheses. V. 51. №3. P. 179-221. Bowles J., 2000. Shattered: Medawar's test tubes and their enduring legacy of chaos // Med. Hypotheses. V. 54. № 2. P. 326-339. Brien P., 1953. La pe´rennite´ somatique // Biol. Rev. Cambr. Philos. Soc. V. 28. P. 308–349. Butenko G.M., 1990. Heterochronic chimeras as a model in gerontological research // The Theoretical Basis of Ageing Research / Eds Robert L., Hofecker G. Wien: Universitatsverlag. P. 65-67. Butenko G.M., 1990. Active mechanisms of dysfunction in the process of aging // Vestn Akad Med Nauk SSSR. № 1 P. 20-23. (Russian). [PubMed ] Cailliet G.M., Andrews A.H., Burton E.J., Watters D.L., Kline D.E., Ferry-Graham L.A., 2001. Age determination and validation studies of marine fishes: do deepdwellers live longer? // Exp. Gerontol. V. 36. P. 739-764. Calder W.A.I., 1989. Avian longevity and aging // Genetic Effects on Aging II / Ed. Harrison D.E. Caldwell: Telford Press. P. 187-204. Cameron H.A., Woolley C.S., McEwen B.S., Gould E., 1993. Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat // Neuroscience. V. 56. P. 337–344. Campbell R.D., 1965. Cell proliferation in hydra: An autoradiographic approach // Science. V. 148. P. 1231–1232. Campbell R.D., 1967a. Tissue dynamics of steady state growth in Hydra littoralis. I. Patterns of cell division // Dev. Biol. V. 15. P. 487–502. Campbell R.D., 1967b. Tissue dynamics of steady state growth in Hydra littoralis. II. Patterns of tissue movement // J. Morphol. V. 121. V. 19–28. Comfort A., 1979. The Biology of senescence. Edinburgh; London: Churchill Livins. 414p. Corwin J.T., 1981. Postembryonic production and aging of inner ear hair cells in sharks // J. Comp. Neurol. V. 201. P. 541–553. Crapon de Caprona M.D., Fritzsch B., 1983. The development of the retinopetal nucleus olfacto-retinalis of two cichlid fish as revealed by horseradish peroxidase // Dev. Brain Res. V.11. P. 281–301. 28 Daftary S.S., Gore A.C., 2005. IGF-1 in the Brain as a Regulator of Reproductive Neuroendocrine Function// Exp. Biol. Med. V.230. P.292-306. Daniel C.W., Young J.T., 1971. Influence of cell division on an aging process // Expl. Cell Res. V. 65. P. 27-32. Daniel C.W., Aidells B.D., Medina D., Faulkin L.J.Jr., 1975. Unlimited division potential of precancerous mouse mammary cells after spontaneous or carcinogeninduced transformation // Proc. F.A.S.E.B. V. 34. P. 64-67. Dilman V.M. 1958. On age raising activity of somehypothalamic centres //Proceedings of the Ivan P. Pavlov Institute of Physiology, Academy of Sciences of the USSR.V 7. P. 326 - 336 (in Russian). Dilman V.M., 1971. Age-associated elevation of hypothalamic, threshold to feedback control, and its role in development, ageine, and disease // Lancet. № 1 (7711). P. 1211-1219. Dilman V.M., 1981. The law of deviation of homeostasis and disease of aging. Boston: J. Wright PSG Inc. 380 p. Dilman V.M., 1986. Ontogenetic model of aging and disease formation and mechanisms of natural selection // J. Theor. Biol. V. 118. № 1. P. 73-81. Dilman V.M. 1987. Four models of medicine. - Leningrad:Meditsina.— 288 pp.(in Russian). Doetsch F., Caille I., Lim D.A., Garcia-Verdugo J.M., Alvarez-Buylla A., 1999. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain // Cell. V. 97. P. 703–716. Ebert T.A., Southon J.R., 2003. Red sea urchins (Strongylocentrotus franciscanus) can live over 100 years: confirmation with A-bomb 14carbon // Fish Bull. V. 101 №4. P. 915-922. Eriksson P.S., Perfilieva E., Bjork-Eriksson T., Alborn A.M., Nordborg C., Peterson D.A., Gage F.H., 1998. Neurogenesis in the adult human hippocampus // Nat. Med. V. 4. P. 1313-1317. Finch C.E., 1990. Longevity, Senescence, and the Genome. Chicago: Univ. of Chicago Press. 922 p. Finch C.E., Austad S.N., 2001. History and prospects: symposium on organisms with slow aging // Exp. Gerontol. V. 36. №. 4-6. P. 593-597. Fine M.L., 1989. Embryonic, larval and adult development of the sonic neuromuscular system in the oyster toadfish // Brain Behav. Evol. V.34. P. 13–24. 29 Font E., Desfilis E., Perez-Canellas M.M., Garcia-Verdugo J.M., 2001. Neurogenesis and neuronal regeneration in the adult reptilian brain // Brain Behav. Evol. V. 58. P. 276–295. Fox G.Q., Richardson G.P., 1982. The developmental morphology of Torpedo marmorata: electric lobeelectromotoneuron proliferation and cell death // J. Comp. Neurol. V. 207. P. 183–190. Frolkis V.V., Bezrukov V.V., Bogatskaya L.N., Burchinsky S.G., Verkhratsky N.S., Gorban E.N., Kvitnitskaya-Ryzhova T.Yu., Kuznetsova S.M., Mankovsky N.B., Martynenko O.A., Mezhiborskaya N.A., Polyukhov A.M. Potapenko R.I., Pugach B.V., Rushkevich Yu.E. Stupina A.S., Tanin S.A., 1991. Aging of the brain. Leningrad: Nauka. P. 223-225. (in Russian) Gavrilov L.A., Gavrilova N.S., 2002. Evolutionary Theories of Aging and Longevity // Sci.World J. V. 2. P. 339-356. George J.C., Bada J., 1999. Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization // Can. J. Zool. V. 77. P. 571-580. Goldman S.A., 1998. Adult neurogenesis: from canaries to the clinic / J. Neurobiol. V. 36. № 2. P. 267-286. Goldman S., Nottebohm F., 1983. Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain // Proc. Natl. Acad. Sci. USA. V. 80. P. 2390-2394. Gould E., Vail N., Wagers M., Gross C.G., 2001. Adult-generated hippocampal and neocortical neurons in macaques have a transient existence // Proc. Natl. Acad. Sci. USA. V. 98. P. 10910-10917. Gracy R.W., Talent J.M., Kong Y., Conrad C.C., 1999. Reactive oxygen species: the unavoidable environmental insult? //Mutat. Res. V.428. P.17-22. Guerin J. C., 2004. Emerging Area of Aging Research: Long-lived Animals with "Negligible Senescence" // http://www.peter.unmack.net/archive/rml/rmljun96/0117.htmlAnn. N.Y. Acad. Sci. V. 1019. P. 518 - 520. Gupta A, Tsai L.H., Wynshaw-Boris A., 2002. Life is a journey: a genetic look at neocortical development // Nat. Rev. Genet. V. 3. P. 342 - 355. Gusev V.A., 2000. The free-radical theory of senescence in the paradigm of gerontology // Adv.Gerontol. № 4. P. 271-272. (in Russian). Free 30 Holmes D.J., Austad S.N., 1995. Birds as animal models for the comparative biology of aging: a prospectus // J. Gerontol. A Biol. Sci. Med. Sci. V. 50. №2. P. 59-66. Holmes D.J., Fluckiger R., Austad S.N., 2001. Comparative biology of aging in birds: an update // Exp. Gerontol. V. 36. № 4-6. P. 869-883. Huang H.H., Kissane J.Q., Hawrylewicz E.J., 1987. Restoration of sexual function and fertility by fetal hypothalamus transplant in impotent aged male rats // Neurobiol. Aging. V. 8. P. 465-472. PubMed Johns P.R., Easter S.S.Jr., 1977. Growth of the adult goldfish eye. II. Increase in retinal cell number // J. Comp. Neurol. V. 176. P. 331–342. Johnson F.B., Sinclar, D.A., Guarente L., 1999. Molecular biology of aging // Cell. V. 96. P. 291-302. Kalman M., Szekely A.D., Csillag A., 1993. Distribution of glial fibrillary acidic proteinimmunopositive structures in the brain of the domestic chicken (Gallus domesticus) // J. Comp. Neurol. V. 330. P. 221-237. Kaluev A.V., 2003. Phenoptosis and human (concerning conception of the academician V.P. Skulachev) // Modern neurobiology and neuroscience / Scientific-educational server. URL: http://www.neuroscience.ru/content/view/18/26/ (in Russian). Kara T.C., 1994. Ageing in amphibians //Gerontology. V.40. №2-4. P. 161-173. Kerr J.F.R., Gobé G.C., Winterford C. M., Harmon B.V., 1995. Anatomical methods in cell death // Cell Death / Eds Schwartz L.M., Osborne B.A. San Diego: Acad. Press. P. 1–27. King J.S., 1966. A comparative investigation of neuroglia in representative vertebrates: a silver carbonate study // J. Morphol. V. 119. P. 435–466. Kishi S., Uchiyama J., Baughman A.M., Goto T, Lin M.C., Tsai S.B., 2003. The zebrafish as a vertebrate model of functional aging and very gradual senescence // Exp. Gerontol. V. 38. №. 7. P. 777-786. Khalyavkin A.V. 1998. "Organism-environment" interaction and origin of aging // Adv.Gerontol. №. 2. P. 43-48. (in Russian) Khalyavkin A.V., Yashin A.I. 2003. Environmental and genetic modification of aging pattern. Borders of plasticity // Problemy starenija i dolgoletija. V. 12. № 4. P. 417425. Klapper W, Heidorn K, Kuhne K, Parwaresch R, Krupp G., 1998. Telomerase in “immortal fish” // FEBS Lett. V. 434. P. 409-412. 31 Koumans J.T.M., Akster H.A., 1995. Myogenic cells in development and growth of fish // Comp. Biochem. Physiol. V. 110A. P. 3–20. Kushima K., Kamio K., Okuda V., 1961. Climacterium, climacteric disturbances on rejuvenation of sex center // Tohoku J. Exp. Med. V. 74. P. 113-129. Lamb M.J., 1980. Biology of Ageing. Moscow: Mir. 206 p. (in Russian). Lamberts S.W.J., Beld A.W., Lely A.J., 1997. The endocrinology of aging // Science. V. 278. P. 419-425. Leaman B.M., Beamish R.J., 1984. Ecological and management implications of longevity in some northeast Pacific groundfishes // Bull. Int. North Pacific Commn. V. 42. P. 85-97. Lennington J.B., Yang Z., Conover J.C., 2003. Neural stem cells and the regulation of adult neurogenesis // Reprod. Biol. Endocrinol. V.1. P. 99-106. Levitt P., Rakic P., 1980. Immunoperoxidase localization of glial fibrrllary acidic protein in radial glial cells and astrocytes of the developing Rhesus monkey brain // J. Comp. Neurol. V. 193. P. 815-840. Litoshenko A.Ya., Hartwig M. 1998. Mitochondria and aging // Problemy starenija i dolgoletija. V. 7. N 3. P.241 -250 (in Russian). Lynch A.J.J., Barnes, R.W., Cambecedes J., Vaillancourt R.E., 1998. Genetic Evidence that Lomatia tasmanica (Proteaceae) Is an Ancient Clone // Austr. J. Bot. V. 46 P. 25-33. Margotta V., Fonti R., Palladini G., Filoni S., Lauro G.M., 1991. Transient expression of glial-fibrillary acidic protein (GFAP) in the ependyma of the regenerating spinal cord in adult newts // J. Hirnforsch. V. 32. P. 485–490. Mart´inez D.E.,1998. Mortality patterns suggest lack of senescence in hydra // Exp. Gerontol. V. 33. № 3. P.217-225. Medawar P.B., 1952. An unsolved problem of biology. L.:H.C. Lewis & Co LTD. 24 p. Meyer R.L., Sakurai K., Schauwecker E., 1985. Topography of regenerating optic fibers in goldfish traced with local wheat germ injections into retina: evidence for discontinuous microtopography in the retinotectal projection // J. Comp. Neurol. P. 239. P. 27–43. Miller R.H., Liuzzi F.J., 1986. Regional specialization of the radial glial cells of the adult frog spinal cord // J. Neurocytol. V. 15. P. 187-196. 32 Molowny A., Nacher J., Lopez-Garcia C., 1995. Reactive neurogenesis during regeneration of the lesioned medial cerebral cortex of lizards // Neuroscience V. 68. P. 823–836. Morrison J.H., Hof P.R., 1997. Life and death of Neurons in the Aging brain // Science. V. 278. P. 412-419. Munk K.M., 2001. Maximum ages of groundfishes in waters of Alaska and British Columbia and considerations of age determination // Alaska Fish Res. Bull. V. 8. P. 12-21. Olovnikov AM. 1971. Principle of marginotomy in template synthesis of polynucleotides // Dokl Akad Nauk SSSR. V. 201. № 6. P. 1496-1499. (Russian). Olovnikov AM. 1992. Aging is a result of a shortening of the "differotene" in the telomere due to end under-replication and under-repair of DNA // Izv Akad Nauk SSSR Biol. V.4. P.641-643. (Russian). Olovnikov AM. 2003. Aging as an universal chronic "disease of quantitative traits": cellular senescence and rna-dependent ionic modulation of genes productivity // Medline.RU V. 4. P. 31. Free Ott R., Zupanc G.K.H., Horschke I., 1997. Long-term survival of postembryonically born cells in the cerebellum of gymnotiform fish, Apteronotus leptorhynchus // Neurosci. Lett. V. 221. P. 185–188. Ottinger M.A., 2001. Quail and other short-lived birds // Exp. Gerontol. V. 36. № 4-6. P. 859-868. Patnaik B.K., 1994. Ageing in reptiles // Gerontology. V.40. №2-4. P. 200-220. Paton J.A., Nottebohm F.N., 1984. Neurons generated in the adult brain are recruited into functional circuits // Science. V. 225. P. 1046–1048. Polezhaev L.V. 1968. Loss and repair of regenerative ability among animals. Moscow: Nauka. 248 pp.(in Russian). Potapenko A.I., Akif'ev AP., 2003. On the way of search of the program and the initial substratum of senescence // Medline.RU. V. 4. P. 103-107. Free. (in Russian). Raff M.C., 1992. Social controls on cell survival and cell death // Nature. V. 356. P. 397–400. Raff M.C., Barres B.A., Burne J. F., Coles H.S., Ishizaki Y., Jacobson M.D., 1993. Programmed cell death and the control of cell survival: lessons from the nervous system // Science. V. 262. P. 695–700. 33 Rakic P., 1981. Neuronal-glial interaction during brain development // Trends Neurosci. V. 4. P. 184-187. Rakic P., 2003. Developmental and Evolutionary Adaptations of Cortical Radial Glia // Cereb. Cortex. V. 13. №. 6. P. 541-549. Reznick D., Buckwalter G., Groff J., Elder D., 2001. The evolution of senescence in natural populations of guppies (Poecilia reticulata): a comparative approach // Exp. Gerontol. V. 36. P. 791–812. Reznick D.N., Bryant M.J., Roff D., Ghalambor C.K., Ghalambor D.E., 2004. Effect of extrinsic mortality on the evolution of senescence in guppies // Nature. V. 431. № 7012. P. 1095-1099. Rougier G.W., Novacek M.J., 1998. Early mammals: teeth, jaws and finally a skeleton!// Curr. Biol. V.8. №. 8. P. 284-287. Rowe R.W.D., Goldspink G., 1969. Muscle fibre growth in five different muscles in both sexes of mice. I.Normal mice // J. Anat. V. 104. P. 519–530. Ruben R.J., 1967. Development of the inner ear of the mouse: a radioautographic study of terminal mitoses // Acta Otolaryngol. (Suppl.) V. 220. P. 1–44. Rubner M., 1908. Das problem des Lebensdauer und seine Beziehungen zu Wachstum und Ernährung. Műnich: med. Wochenshr. 260 s. Sacher G.A., 1977. Life table modification and life prolongation // Handbook of the Biology of Aging /Eds Finch C., Hayflick L. N.Y.: Reihold. P. 582-638. Sacher G.A., 1978. Longevity and aging in vertebrate evolution // Bioscience. V. 28. P. 497-501. Savelyev S.V., 2005. Origin of brain. Moskow: Vedy. 368 p. Schmechel D., Rakic P., 1979. A Golgi study of radial glial cells in developing monkey telencephalon: Morphogenesis and transformation into astrocytes // Anat. Embriol. V. 156. № 2. P. 115-152. Schwarz S.S., 1976. Ecological approaches to the analysis of senescence and change of life-span // Gerontology and geriatrics. Annual. 1975. Biological possibilities of increase of life-span. Kirov. P. 19 - 29. Seri B., Garcia-Verdugo J.M., McEwen B.S., Alvarez-Buylla A., 2001. Astrocytes give rise to new neurons in the adult mammalian hippocampus // J. Neurosci. V. 21. P. 7153–7160. Skulachev V.P., 1997. Aging is a Specific Biological Function Rather than the Result of a Disorder in Complex Living Systems: Biochemical Evidence in Support of 34 Weismann's Hypothesis // Biokhimia (Moscow). V. 62. P. 1191-1195. Free (Translated in Inglis) Skulachev V.P., 1999. Mitochondrial physiology and pathology; concepts of programmed death of organelles, cells and organisms // Mol. Aspects Med. V. 20. P. 139-184. Skulachev V.P. 1999. Phenoptosis: programmed death of an organism // Biokhimia (Moscow). V. 64. N12. P. 1418 -1426. Free. (in Russian). Skulachev V.P., 2001. Ageing: particular case of phenoptosis // Soros Educational Journal. V. 7. № 10. P. 7-11. Free Soutschek J., Zupanc G.K.H., 1995. Apoptosis as a regulator of cell proliferation in the central posterior/prepacemaker nucleus of adult gymnotiform fish, Apteronotus leptorhynchus // Neurosci. Lett. V. 202. P. 133–136. Soutschek J., Zupanc G.K.H., 1996. Apoptosis in the cerebellum of adult teleost fish, Apteronotus leptorhynchus // Dev. Brain Res. V. 97. P. 279–286. Suarez I., Bodega G., Rubio M., Fernhuiez B., 1995. Evolution of Astrocytes in the Vertebrate CNS // Neuron-Glia Interrelations During Phylogeny / I. Phylogeny and Ontogeny of Glial Cells. Totowa: Humana Press. P. 41-58 Stuermer C.A.O., Bastmeyer M., Bähr M., Strobel G., Paschke K., 1992. Trying to understand axonal regeneration in the CNS of fish // J. Neurobiol. V. 23. P. 537– 550. Takhtajan A. 1964. Principles of the developmental morphology of metasperms. Moscow, Leningrad: Nauka. 236 p. (in Russian) Takhtajan A. 1966. A system and phylogeny of the flowering plants. Moscow, Leningrad: Nauka. 612 p. (in Russian) Takhtajan A. 1970. Origin and settling of the flowering plants. Leningrad: Nauka. 147 p. (in Russian) Vaganov Е.А., Naurzbaev M.M., Hughes M.K., 2000. Witnesses rise of warming of climate in the Middle Ages // Priroda. № 12. P. 54-57. (Russian) Free Vanyushin B.F., 2001. Apoptosis in plants // Usp. Biol. Khim. V. 41. P. 3-38. (in Russian). Free Viktorov I.V., 2001.Stem Cells of Mammalian Brain: Biology of the Stem Cells in vivo and in vitro // Izv Akad Nauk Ser Biol. № 6. P. 646-655. (in Russian). PubMed Free Voight T., 1989. Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from radial glia into astrocytes // J. Comp. Neurol. V. 289. P. 35 74-88. Voronova N.V., Klimova N.M., Mendgeritskiy A.M., 2005. Anatomy of the central nervous system: The tutorial for the students of institute of higher education. Moskow: Aspect Press. 128 p. (in Russian). Waxman S.G., Anderson M.J., 1985. Generation of electromotor neurons in Sternarchus albifrons: differences between normally growing and regenerating spinal cord // Dev. Biol. V. 112. P. 338–344. Waxman S.G., Anderson M.J., 1986. Regeneration of central nervous system structures: Apteronotus spinal cord as a model system // Electroreception / Eds Bullock T.H., Heiligenberg W. N. Y.: John Wiley & Sons. P. 183–208. Weismann A., 1889. Essays upon heredity and kindred biological problems. V. 1. Oxford: Claderon Press. 455 p. Weissman T., Noctor S.C., Clinton B.K., Honig L.S., Kriegstein A.R., 2003. Neurogenic Radial Glial Cells in Reptile, Rodent and Human: from Mitosis to Migration // Cereb. Cortex. V. 13. №. 6. P. 550-559. Williams G.C., 1957. Pleiotropy, natural selection, and the evolution of senescence // Evolution. V. 11. P. 398-411. species Zhang Z., Krebs C.J., Guth L., 1997. Experimental analysis of progressive necrosis after spinal cord trauma in the rat: etiological role of the inflammatory response // Exp. Neurol. V. 143. P. 141–152. Zupanc G. K. H., 1996. Peptidergic transmission: from morphological correlates to functional implications // Micron. V. 27. P. 35–91. Zupanc G.K.H., 1997. Towards a cellular understanding of motivation // Adv. Ethol. V. 32. P. 19. Zupanc G.K.H., 1999. Neurogenesis, cell death and regeneration in gymnotiform brain // J. Exp. Biol. V. 202. P. 1435–1446. Zupanc G.K., 2006. Neurogenesis and neuronal regeneration in the adult fish brain // J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. / Epub ahead of print. P. 1-22 Zupanc G.K., Clint S.C., 2001. Radial glia-mediated up-regulation of somatostatin in the regenerating adult fish brain // Neurosci. Lett. V. 309. № 3. P. 149 - 152. Zupanc G.K.H., Horschke I., 1995. Proliferation zones in the brain of adult gymnotiform fish: a quantitative mapping study // J. Comp. Neurol. V. 353. P. 213– 233. 36 Zupanc G.K.H., Horschke I., Ott R., Rascher G.B., 1996. Postembryonic development of the cerebellum in gymnotiform fish // J. Comp. Neurol. V. 370. P. 443–464. Zupanc G.K., Hinsch K., Gage F.H., 2005. Proliferation, migration, neuronal differentiation, and long-term survival of new cells in the adult zebrafish brain // J. Comp. Neurol. V. 488. №3. P. 290-319.