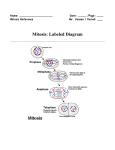
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cell growth wikipedia , lookup
Cell culture wikipedia , lookup
Endomembrane system wikipedia , lookup
Cellular differentiation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cytokinesis wikipedia , lookup
Arch Microbiol (1998) 170 : 227–235 © Springer-Verlag 1998 O R I G I N A L PA P E R Carl Robinow · Esther R. Angert Nucleoids and coated vesicles of “Epulopiscium” spp. Received: 3 December 1997 / Accepted: 11 June 1998 Abstract We describe here aspects of the anatomy of two “Epulopiscium” morphotypes, unusually large bacteria that are not yet cultured and that reproduce by the internal generation of two or more vegetative daughter cells. Two morphotypes, A and B, which are enteric symbionts of several species of herbivorous surgeonfish (Acanthuridae), were collected around the Great Barrier Reef of Australia, preserved there, and later stained for light microscopy. Some samples were examined by electron microscopy. In both morphotypes, countless discrete nucleoplasms or nucleoids were found to occupy a single shallow layer just beneath the surface all around these organisms. At each end of the morphotype B cells, a membranebound compartment containing dense cords of chromatin was observed. When these were found at each end of growing daughter cells, no polar compartments were then found in their mother organism. Electron micrographs of sections of morphotype A symbionts show that their outermost region is composed of tightly packed coated vesicles, each surrounded by a thin, dense, spacious capsule. Near the surface of type A organisms the remains of broken vesicles, broken capsules, and a finely fibrous matrix fuse to form a fabric that serves as the cell wall. Morphotype B organisms, however, were observed to have a distinct, morphologically continuous outer wall. Key words ”Epulopiscium” · Nucleoids · Polar chromatin · Coated vesicles · Unusual composite wall · Daughter cells C. Robinow (Y) Department of Microbiology and Immunology, Health Sciences Centre, University of Western Ontario, London, Ontario, Canada N6A 5C1 e-mail:[email protected] Tel. +1-519-661-3427; Fax+1-519-661-3499 E. R. Angert Department of Molecular and Cellular Biology, Harvard University, Cambridge, MA 02138, USA e-mail: [email protected] Tel.+1-617-495-0532; Fax+1-617-496-4642 Abbreviations DAPI 4′,6-Diamidino-2-phenylindole · PBS Phosphate buffered saline Introduction ”Epulos” has been used as the name of members of a large and varied group of uncommonly large bacteria (Clements et al. 1989). The first such organism was reported from the gut of the herbivorous surgeonfish Acanthurus nigrofuscus from the Red Sea. It was given the name “Epulopiscium fishelsoni” and was placed in the kingdom Protista (Fishelson et al. 1985; Montgomery and Pollak 1988). Various organisms sharing some of its properties were later encountered in surgeonfish from Australia’s Great Barrier Reef (Clements et al. 1989). Electron microscopy of sections of the large organisms showed that they contain typically prokaryotic strands of chromatin and have bacterial-type flagella. Their large size apart (some of them attain a length of more than 500 µm), these bacteria are unusual in that they multiply (or merely rejuvenate themselves) in a viviparous mode of internal formation of one, two, or more daughter cells that escape to the outside through a tear in the wall of the mother organism (Montgomery and Pollak 1988). Sequence comparisons of the 16S rRNA gene (Angert et al. 1993) have placed “Epulopiscium” among the anaerobic, gram-positive, spore-bearing clostridia. More recently, Angert et al. (1996) have reported that the unusually large Metabacterium polyspora (Chatton and Pérard 1913), which is a symbiont of the cecum of guinea pigs and other rodents and which forms two or more refractile endospores per cell, is also located in this branch of the genealogical tree. Spore-bearing cells of Metabacterium spp. superficially resemble “Epulopiscium” morphotypes engaged in the formation of vegetative daughter cells. Here we describe observations made with the light microscope on nucleoids and other forms of chromatin in “Epulopiscium” morphotypes. A brief account is also provided of elements of the complex outermost layer or “cortex” of type A organisms as seen in electron micrographs of sections. 228 Materials and methods tail found to have been passably well-preserved, and this only in the outermost regions of these symbionts. Sample collection Numerous samples of gut contents of surgeonfish were collected on several occasions on Lizard Island (a part of the Great Barrier Reef, Australia) by K.D. Clements. Samples were preserved there in several different ways appropriate for 4′,6-diamidino-2-phenylindole(DAPI)-enabled fluorescence of DNA, the staining of bacterial chromatin for light microscopy and electron microscopy (see below). DAPI fluorescence Samples of gut contents were fixed and stored in 80% ethanol in 1993 and were recently prepared for the detection of DNA in the following manner. Fixed samples were applied to poly-L-lysinecoated coverslips and air-dried. The coverslips were then immersed in a solution of 0.5 µg/ml DAPI in phosphate-buffered saline (PBS) for approximately 2 min; subsequently, they were rinsed with PBS. The coverslips were next mounted over a PBS/ 50% glycerol solution. Slides were viewed and photographed using an Olympus BX 60 epifluorescence microscope as previously described by Harry et al. (1995). Cells were visualized using an excitation cube unit (U-MNU) appropriate for viewing DAPI fluorescence with a narrow band-pass (360–370 nm) excitation filter and a long band-pass (420 nm) barrier filter. Photographs of fluorescent preparations were obtained on Kodak T-MAX film, ASA 400. Transmission light microscopy The fixative for light microscopy of the large bacteria from surgeonfish was a mixture of two parts of a saturated aqueous solution of mercuric chloride with one part of absolute alcohol and three parts water with acetic acid added to the mixture to make it 2% of the total volume. Fixation time was 10–15 min, after which the sample was washed with and stored in 70% ethanol. Nucleoid staining was achieved by both the Feulgen procedure and the HClGiemsa method (Piekarski 1937). Hydrolysis for 8–10 min with 1 M HCl at 60°C was followed by staining with the Giemsa mixture of dyes. We used Gurr’s Giemsa “Improved 66” (British Drug Houses, Pool, England) at one drop of stain per ml of distilled water that had been given a pH of 6.8 by the use of Gurr’s buffer tablets (British Drug Houses). The progress of staining was checked with a × 40 water immersion lens. Photographs were taken of wet, stained preparations mounted over buffer of pH 6.8. Electron microscopy Samples intended for electron microscopy were prepared at the site of collection in 2.5% glutaraldehyde/0.2 M cacodylate buffer (pH 7.2). Samples were fixed for 30–45 min, then washed with and stored in 0.2 M cacodylate (pH 7.2) at 4°C. On arrival at the London laboratory, the deposit of centrifuged sample was gelled in Noble agar and post-fixed for 1 h in 1% OsO4 in 0.2 M cacodylate (pH 7.2), washed with water, and dehydrated via a series of ethanol solutions of a concentration gradually increasing from 30 to 100%. Dehydrated samples in agar were next placed into Spurr resin that was polymerized overnight at 60°C. Sections were cut on a hand-operated Porter Blum microtome, were stained for 5 min with uranyl acetate (2%) followed by 1 min in 1.4% lead citrate/1.8% sodium citrate, and were then examined in a Philips 300 electron microscope. The fixation procedures just described have routinely provided satisfactory images of the membranes, nucleoids and ribosomes of bacteria studied in our laboratory, but the interval between primary fixation of the giant symbionts in Australia and secondary fixation with osmium 5 weeks later proved to have been too long. Only in sections of the A morphotype was de- Results The A morphotypes in our samples measured 200–250 µm in length, were slender, and tapered towards their tips. Some of them bore one or two daughter cells, mostly at the very early stages of development. B morphotypes were only 100–185 µm long; they were straight cylinders with rounded ends or were of slender cigar shape. Many of these organisms bore one or two daughter cells. Nucleoids of morphotype B As Fig. 1 (A–E) and Fig. 2 (A,B) clearly show, B morphotypes have innumerable, closely-packed, small nucleoids spaced all over their periphery in a shallow layer just beneath their surface. In DAPI preparations, brightly fluorescing polar compartments were invariably found at the tips of free, mature individuals or in the tips of internally arising offspring. These will be discussed below. At the higher magnification of the Giemsa-stained preparation in Fig. 3, we found that the nucleoid layer is composed of large numbers of thin, flexible entities, nucleoids or nucleoplasms, packed together in single-layered, flat patches of widely differing size and shape. Neighboring patches are connected with strands or narrow ribbons also composed of nucleoids. Overall, the arrangement here is that of a reticulate sheet – to borrow a term used by Smith (1956) to describe the structure of the chloroplast of the green alga Oedogonium. The nucleoids of B symbionts vary in length and on the whole appear to be somewhat smaller than those of common Bacillus species. After Feulgen-style acid hydrolysis, the nucleoids of B morphotypes displayed an affinity for purple components of the Giemsa stain, as in the case of other bacteria, but they did so rather weakly. Oddly enough, they also proved to be only weakly, if quite distinctly, Feulgen-positive in preparations in which the nuclei of protozoa, which were often scattered among the giant symbionts, gave the expected strongly positive response to the Feulgen procedure. In retrospect, it seems possible that a shorter time spent in acid hydrolysis would have evoked a stronger positive response of the B-type nucleoids to the Feulgen treatment, particularly since DAPI scans showed the nucleoids of B symbionts fluorescing as brightly as the nucleoids of other bacteria contained in the same samples. Densely chromatinic cords in membrane-enclosed space at the tips of B-type symbionts The weakly positive response of B morphotypes to the Feulgen procedure is all the more remarkable because in our samples, mature singles or pairs of daughter cells still enclosed in their mother organism displayed, in the ma- 229 Fig. 1A–E Epifluorescence of B morphotypes stained with 4′,6diamidino-2-phenylindole. A and B show median- and upper-surface optical sections, respectively, of the same cell. A The layer of nucleoids just under the cell cortex is seen as a bright broken line. DNA associated with putative daughter cell primordia is seen at both poles. B A surface view of the network of islands of nucleoids (A,B bars 50 µm). C and D show examples of B morphotypes with mature daughter cells. Note the fine broken line of mother cell nucleoids surrounding the brighter daughter cell nucleoid layer. C Two daughter cells in this image show coalesced DNA at the poles and at regions along the side of each daughter cell. These may represent “granddaughter” primordia. D The unusual instance of a single daughter cell. Coalesced DNA is seen at the poles of the daughter cell but not in the mother cell. E A surface view of a lateral chromatin cluster of cords of DNA (C–E bars 20 µm) jority of instances, at both poles small lens-shaped compartments containing several cords (which may be part of a single knot) that were strongly Feulgen-positive and also stained deeply after HCl-Giemsa (Fig. 4, A–C). We shall refer to these structures as terminal or polar chromatin compartments. The cords of these compartments are much larger and denser than individual nucleoids of the superficial net shown in Fig. 2B. We believe that Fig. 5D of Montgomery and Pollak (1988) of a stout, twisted cord of nucleoplasm in the center of what the authors regard as a developing daughter cell may represent part of a knot of cords of chromatin within its compartment similar to what we have observed at the tips of B morphotypes. We can but speculate what may be the function of this organelle that is generally present at both poles of the B morphotype. Aware that daughter cells of some morphotypes of “Epulopiscium” arise in the tips of mother cells [see, e.g., Fig. 6 of Montgomery and Pollak (1988)], we propose that the terminal compartments with their prominent cords of chromatin may be primordia of daughter cells. Among the DAPI-stained B-morphotype cells scanned, many harbored long, slender daughters that invariably carried strongly fluorescing chromatin compartments at both ends, while no such compartments were found in the remaining cytoplasmic space of their mother organism even when, as in Fig. 1C, the mother cell’s own superficial nucleoids were still vividly fluorescing. That is what would be expected if, as suggested above, activated terminal compartments transform themselves into daugh- 230 Fig. 2A, B Morphotype B (HCl-Giemsa). Views of the same organism at two different levels of focus. A Median optical section. Note the straight cell wall of even density. The nucleoids appear as a line of pieces of chromatin just beneath the cell wall. The dark, lens-shaped body at the tip is of a kind regularly found at both poles of growing and of mature B morphotypes. Normally these compartments contain stout cords of chromatin (see Fig. 4, A–C). However, during the short interval between taking the photographs of A and B, the dark stain diffused away from the compartment and revealed in B – for reasons unknown – that the compartment was empty. B Overview of the superficial reticulate sheet of nucleoids. The shreds of membranous materials clinging to this cell are probably remains of the wall of its mother organism (bars 5 µm) ter cells. For direct evidence of this proposed course of events it would, however, be necessary to examine Btypes from much earlier phases of the growth cycle than those represented in the samples that have been available to us. The DAPI preparations have also revealed instances of cells that had, in addition to polar chromatin compartments, one or more brightly fluorescing clusters of chromatinic cords along their sides (Fig. 1, C and E). Such lateral clusters could perhaps account for certain B morphotypes mentioned by Clements et al. (1989) that harbor more than two daughter cells. A look at 3-year-old Feulgen preparations in London revealed there, too, three instances of solidly Feulgen-positive, lateral chromatin clusters among twenty B organisms. DAPI fluorescence also drew our attention to the fact that even fully grown type-B twin daughter cells often continue to cling together for some time within the faintly illuminated remains of the cell wall of their mother organism. Such remains are Fig. 3 Morphotype B (HCl-Giemsa). Superficial net of clusters of tightly packed nucleoids. The group indicated with an arrow is in best focus (bar 5 µm) seen clinging to much of the surface of the morphotype B of Fig. 2B. Nucleoids of morphotype A In A morphotypes, the nucleoids were evenly distributed over the entire subsurface plane of the cells (Fig. 5, A and B). The nucleoids were not arranged there in any particular order, but were a remarkably uniform distance from each other and quite possibly exist as distinct and separate entities; differences in the size of individual nucleoids may reflect their having been preserved at different stages of their replication cycle. Near the tip of A symbionts, nucleoids tend to be more closely packed together than over the rest of the body of these organisms, and some may extend from the plane of the main nuclear layer and enter deeper regions of the cytoplasm. The nucleoids were strongly Feulgen-positive, and most of them were slightly larger than nucleoids of cells from fast growing cultures of Bacillus mycoides and Bacillus megaterium [see Robinow and Kellenberger (1994)]. Chromatin of internally generated daughter cells The developing type-A daughter cells we have encountered were of two kinds. There were those whose chromatin presented as a tubular shell of seemingly solid chromatin perforated by large irregular holes (Fig. 5B). In the 231 Fig. 4A–C B morphotypes A, C HCl-Giemsa and B Feulgen. Examples of polar compartments containing stout cords of chromatin (bars 5 µm) other kind, which we regard as having been in a more advanced state of development, the chromatin was disposed in much the same way as it was in the surface layer of the mother cell except that the daughter’s chromatin tended to stain more deeply than that of its mother organism. The fluorescence of the chromatin of B morphotypes harboring juvenile cells seemed approximately equally bright in mother organisms and offspring (Fig. 1C). Light microscopy of the structure of the surface of B and A morphotypes Morphotypes B and A differ markedly in the nature of the boundary between themselves and their environment. Optical sections such as that of Fig. 2A show that the type-B symbiont is surrounded by a proper wall of geometrical neatness and unvarying density. A narrow, translucent zone intervenes between the wall and the nucleoid layer. In contrast to this unexceptional pattern, we find the A morphotypes of Fig. 5 (A and B) surrounded by a thick “cortex”, to borrow a term adopted by Montgomery and Pollak (1988) for the membranous outer boundary of “E. fishelsoni” as a sign that they found it differed significantly from the cell walls of plants and bacteria. Note that in our usage “cortex” will refer to all of the region exterior to the layer of nucleoids. The type-A cortex of Fig. 5 (A and B) seems to be of the nature of a soft rind that lacks the sharply defined outer and inner contours of a proper wall, varies slightly in thickness along the length of the cell, and is not set off by a distinct gap from the nucleoid layer. The complexity of this cortex was revealed by electron microscopy (Figs. 6, 7). Fig. 5A, B Two A morphotypes (Feulgen). A Surface view of shallow layer of evenly spaced nucleoids. Note along both edges the rind-like, thick cortex. Out of focus in the interior is the dense chromatin of a developing daughter cell. B Optical section of another A morphotype. In the polar region some nucleoids have entered deeper layers of the cytoplasm. Over the rest of the cell the nucleoids are still seen as forming a shallow layer just beneath the thick, translucent cortex. In the interior is seen the fenestrated tube of dense chromatin of a developing daughter cell (bars 5 µm) 232 Fig. 6 Electron micrograph of a section of the cortex of another A morphotype. The cloud-like, faintly speckled shapes near the bottom are profiles of nucleoids (n). Above these is a labyrinth of interconnected spaces. Further up there is yet another labyrinth where the open spaces are lined with much denser membranes (d) than are those of the main labyrinth below. The outer border of the cortex appears to be formed by roughly circular profiles of irregularly stacked vesicles (bar 1.0 µm) Electron microscopy of the cortex of A morphotypes An overview of the periphery of a typical A morphotype is provided in Fig. 6. Up from the micrograph’s lower edge are seen profiles of nucleoids of low density and irregular contours similar to what has been found in E. coli preserved only with gluteraldehyde, where, according to Hobot et al. (1985), “… coarse aggregates of DNA were present within a seemingly empty dispersed nucleoid.” Continuing upwards, three levels of the outer cortex can be seen. Prominent is an extensive layer of labyrinthine design reminiscent of the maze of interconnected spaces found by Clements and Bullivant (1991) just inside a symbiont’s cell wall. Its upper reaches here are formed by a shallow layer of open spaces whose lining membranes are much denser than those of the main labyrinth below. Higher still, a region of stacked ovoid or spherical structures is observed, and beyond these a layer of tightly packed, ill-defined, perhaps fibrous materials are seen. Finally, exterior to this blurred horizon, a mat of bent and broken filaments are observed; these have also been encountered at the periphery of “Epulopiscium” morphotypes studied by Clements and Bullivant (1991) and, on the basis of their fine-structure, have been identified by them as bacterial flagella. Figure 7 shows a stretch of cortex similar to that of the previous figure but at higher magnification. What is seen in the higher levels of this micrograph and was found regularly in other sections of long stretches of uniformly structured cortex of A morphotypes, we regard as profiles of stacks of spherical or ovoid vesicles, each one enclosed within a spacious, thin, dense membranous capsule. Finely fibrous material appears to fill the space between neighboring capsules. The vesicles within, with diameters in the order of 0.11 µm, we regard as belonging to the “decorated” or “bristle-coated” kind familiar to cell biologists. The closely packed decorated vesicles resemble the “fluid segregating organelles” of Paramecium (McKanna 1976). These take the form of fascicles of narrow tubules, blind at one end, that are situated proximal to the canals 233 Fig. 7 High magnification of a short stretch of the outermost region of the cortex shows stacks of the characteristic profiles (thin arrows) of “decorated” or “bristle-coated” vesicles, each one surrounded by the profile of its thin but dense, spacious membranous capsule. Below, thick arrowheads point to vesicles preserved in the process of being encapsulated (bar 0.2 µm) conducting fluid to the pulsating vacuoles. The constituent tubules of the fascicles bear an array of minute pegs that endow cross-sections of them with a strong likeness to the bristle-coated vesicles of the outer cortex of the A morphotype. Discussion Mode of reproduction of the A morphotype The A morphotypes, like “E. fishelsoni” itself, reproduce by the internal generation of daughter organisms (Fishelson et al. 1985; Montgomery and Pollak 1988; Clements et al. 1989), but to date we have not found out where this process initiates. Most probably the development of daughter morphotypes starts at some point of the superficial nucleoid layer with its profusion of discrete nucleoplasms, but unambiguous evidence of such an event has not been encountered in our samples. Among A morphotypes are found many that bear a single, short daughter cell in the cytoplasm of the middle region of the mother organism. Thus, a count performed on two Feulgenstained slides of A morphotypes from the same sample yielded 27 that had single daughters varying from 28 to 40 µm in length and only 13 that bore two, long, overlapping daughter cells. As to the origins of these pairs, there would seem to be two possibilities. One scenario would be that of two short daughter cells of the same length, hence probably initiated at the same time (of which we have seen examples), lying some distance apart, both growing, eventually overlapping, and occupying most of the available space. Another conceivable origin of overlapping pairs would be their having arisen from a single short cell that had grown to great length and had, in the end, divided by binary fission before extrusion from the mother organism. To allow for the possibility of binary fission of a single internally generated daughter cell would not be unreasonable. It has, in fact, been observed to take place in the cells of long, segmented filamentous bacteria attached to the lining of the ileum of mice (Ferguson and Birch-Andersen 1979). Then again, two of Montgomery and Pollak’s (1988) striking electron micrographs show a single, snake-like cell emerging from a torn wall of the mother organism. If these scenes had been part of a process of reproduction, then binary fission would, presumably, in such instances have taken place soon after extrusion. If not, what is observed here are instances of mere rejuvenation and not propagation since the mother organism is destroyed during the process. Uncertain is also the life story, past and future, of the occasionally encountered instances of B morphotypes bearing a single daughter within a mother organism whose own nucleoid layer is still visible outside that daughter cell, as in Fig. 1D. Coated vesicles of “E. fishelsoni” The A morphotypes from surgeonfish around the Great Barrier Reef (Australia), are not the only ones that are equipped with coated or decorated vesicles. In electron micrographs accompanying their ground-breaking paper on “E. fishelsoni,” the giant enteric symbiont of surgeonfish of the Red Sea, Montgomery and Pollak (1988) describe “reticulate membranes and tubules at the cell periphery.” A close look at their work has convinced us that the structures referred there to are what used to be called “alveolate” vesicles or tubules. Some vesicles there have 234 a spiny circumference. Both aspects have been described by Leedale (1967) in his writing about alveolate vesicles near the pulsating vacuole of Euglena spirogyra. As he put it: “Alveolate vesicles are distinctive structures, their walls carrying a well defined alveolate patterning which in sectioned vesicles appears as hairs radiating from the surface”. We are aware that in current work in the field of molecular cell biology, alveolated/reticulated vesicles would be referred to as being “clathrin-coated,” but for the time being we feel it appropriate to refer to such structures in the manner of the authors whose work we are here discussing. Coated vesicles are known to perform a variety of functions not only in protists but also in the cells of certain tissues of plants and animals. Electron micrographs do not reveal what coated vesicles contribute to the maintenance of the giant bacteria. Tentatively we propose that their function is excretory, that in A morphotypes new vesicles are steadily produced at the base of the vesicle zone (Fig. 7) and are gradually pushed up to more peripheral levels and disintegrate there. The employment of the bristle-coated kind of vesicles for the removal of waste products may be a strategy that allows the A morphotype to overcome obstacles that Koch (1996) sees as preventing bacteria from attaining large size. The continuous, outward-moving supply of broken vesicles and fragments of their membranous capsule, all seemingly embedded in a fibrous matrix, may provide the cohesion required of a layer that has to function as the equivalent of a cell wall. Similar events seem to be happening in “E. fishelsoni.” Montgomery and Pollak (1988) mention that the numerous tubules of this symbiont “frequently touch the inner cortical surface” (i.e., the inner surface of the organism’s bounding membrane), and so they do. But close scrutiny of the relevant electron micrographs has provided clear evidence that at several points the reticulated/alveolated tubules actually fuse with and enter the substance of the cortex and (perhaps only temporarily) impart to stretches of it an alveolar pattern. Here, then, is another example of vesicle/tubule-coating materials contributing something to the building of a structure capable of performing the functions of a wall. The course of events proposed here as taking place near the surface of two kinds of enteric symbiont morphotypes may well seem improbable, but (in morphological terms) it resembles the dynamics of the steady loss of cell wall materials to the outside and their renewal from below that has been demonstrated in grampositive, rod-shaped bacteria (Koch and Doyle 1985; Beveridge and Kadurugamuwa 1996). We have not obtained unambiguous electron micrographs of B morphotypes, but have looked at enough of them to allow us to say that the outer region of the cortex there is not composed of coated vesicles but of more sophisticated durable structures and that B morphotypes have a distinct, twoply, coherent wall. The samples of intestinal symbionts we examined were from those segments of their life cycle through which they pass during the hours of daylight. Thus, the few observations that we have been able to make leave much of the life cycle of these giant symbionts unaccounted for. We hope that others will collect over a stretch of 24 h a set of anatomical/cytological observations on “E. fishelsoni” and some of its morphotypes that would complement the circadian set of numerical data on growth and reproduction collected by Montgomery and Pollak (1988). Meanwhile, it is obvious that the nucleoids of the symbiont morphotypes we studied resemble those of common bacteria, those of type A more obviously so than the strangely small ones of the type-B symbionts. The validity of our suggestion that the polar compartments containing stout cords of chromatin may be primordia of daughter cells depends on the proof or disproof that can only be provided by future life cycle studies. Until this information is available, we find support in the fact that, despite their large size, these polar compartments resemble the passing stage in the development of spores in Bacillus cells in which distinct and variously coiled cords of chromatin are found within a small compartment at the tip of rod-shaped cells (Robinow 1960). Lastly, while the task of alveolate vesicles and tubules of “E. fishelsoni” and bristle-coated vesicles of the A-type cortex can at present only be guessed at, it is of interest to have found that the employment of such organelles is not limited to eukaryotes. Acknowledgements The authors are much indebted to K. D. Clements, now at the University of Auckland (New Zealand), for catching surgeonfish and sending us samples for genetic analysis and microscopy. For electron micrographs, the authors thank D. Moyles, now at the University of Guelph (Ontario, Canada). C. Robinow owes thanks to R. G. E. Murray for putting him on the tracks of E. Angert’s work with the giant bacteria. References Angert ER, Clements KD, Pace NR (1993) The largest bacterium. Nature 362:239–241 Angert ER, Brooks AE, Pace NR (1996) Phylogenetic analysis of Metabacterium polyspora: clues to the evolutionary origin of Epulopiscium spp. the largest bacteria. J Bacteriol 178 : 1451– 1456 Beveridge TJ, Kadurugamuwa JL (1996) Periplasm, periplasmic spaces and their relationship to bacterial wall structure: novel secretion of selected periplasmic proteins from Pseudomonas aeruginosa. Microb Drug Resistance 2:1–8 Chatton É, Pérard C (1913) Schizophytes du caecum du cobaye. 2. Metabacterium polyspora n.g., n.s. C R Hebd Soc Biol (Paris) 74:1232–1234 Clements KD, Bullivant S (1991) An unusual symbiont from the gut of surgeonfishes may be the largest known prokaryote. J Bacteriol 173:5359–5362 Clements KD, Sutton DC, Choat JH (1989) Occurrence and characteristics of unusual protistan symbionts from surgeonfishes (Acanthuridae) of the Great Barrier Reef, Australia. Marine Biol 102:403–412 Ferguson DJP, Birch-Andersen A (1979) Electron microscopy of a filamentous, segmented bacterium attached to the small intestine of mice from a laboratory animal colony in Denmark. Acta Pathol Microbiol Scand B 87:247–252 Fishelson L, Montgomery WL, Myrberg AA (1985) A unique symbiosis in the gut of tropical herbivorous surgeonfish (Acanthuridae: Teleostei) from the Red Sea. Science 229:49–51 Harry EJ, Pogliano K, Losick R (1995) Use of immunofluorescence to visualize cell-specific gene expression during sporulation in Bacillus subtilis. J Bacteriol 177:3386–3393 235 Hobot JA, Villiger W, Escaig J, Maeder M, Ryter A, Kellenberger E (1985) Shape and fine structure of nucleoids observed on sections of ultrarapidly frozen cryosubstituted bacteria. J Bacteriol 162:960–971 Koch AL (1996) What size should a bacterium be? A question of scale. Annu Rev Microbiol 50:317–348 Koch AL, Doyle RJ (1985) Inside-to-out growth and the turnover of the gram-positive rod. J Theor Biol 117:137–157 Leedale GF (1967) Euglenoid flagellates. Prentice Hall, Englewood Cliffs, NJ McKanna JA (1976) Fine structure of the fluid segregation organelles of Paramecium contractile vacuoles. J Ultrastruct Res 54:1–10 Montgomery WL, Pollak PE (1988) Epulopiscium fishelsoni n.g., n.s., a protist of uncertain taxonomic affinities from the gut of an herbivorous reef fish. J Protozool 35:565–569 Piekarski G (1937) Zytologische Untersuchungen an Paratyphusund Coli Bakterien. Arch Mikrobiol 8:428–439 Robinow CR (1960) Morphology of bacterial spores, their development and germination. In: Gunsalus IC, Stanier RY (eds) A treatise on structure and function. Academic Press, New York London, pp 207–248 Robinow CR, Kellenberger E (1994) The bacterial nucleoid revisited. Microbiol Rev 58:211–232 Smith GM (1956) Cryptogamic botony, vol 1, 2nd edn. McGraw Hill, New York