Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Magnesium transporter wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Paracrine signalling wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Western blot wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Point mutation wikipedia , lookup
Metalloprotein wikipedia , lookup
Signal transduction wikipedia , lookup
Genetic code wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Peptide synthesis wikipedia , lookup
Two-hybrid screening wikipedia , lookup
Ribosomally synthesized and post-translationally modified peptides wikipedia , lookup
Biosynthesis wikipedia , lookup
Anthrax toxin wikipedia , lookup
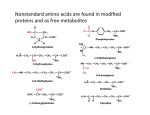
1 AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE Akul Y. Mehta INTRODUCTION: Ever since the discovery of alpha-fetoprotein (AFP) by Bergstrand and Czar in the year 1956, extensive research has been carried out on this molecular wonder of nature to elucidate its functions, its role and its widespread presence. This fetal component which was not commonly in adults was first found as a post albumin migrating protein in fetal serum. Its functions are varied from development of the fetus, where it acts as a major transport protein, to a serum marker in cancer, and its uses in the detection of congenital defects. The following discussion takes a closer look at the molecule with closer insight towards its structure, its role played in the human body, its biochemical functions, and the potential uses of the AFP molecule. PROTEINS In order to understand the structure and functions of AFP it is important to revise the basics of protein structure and function. Proteins are working molecules of a cell that carry out the ‘program’ of activities encoded onto them by genes. The ‘program’ which is nothing but the cell function, requires the coordinated effort of many different types of protein which work in synchronous with each other to provide the desired effect. [1] Classification of Proteins: Proteins are classified into several broad classes based on the functional role played by them within the body. • Structural proteins- which provide structural rigidity to the cell. • Transport proteins- those which control the flow of materials across the body and cellular membranes e.g. albumin and AFP. • Regulatory proteins- they act as sensors and switches to control protein activity and gene function. • Signaling proteins- including cell-surface receptors and other proteins that transmit external signals to the cell interior. • Motor proteins- they are those which cause motion. • Enzymes- specialized proteins which are capable of catalyzing an incredible range of intracellular and extracellular chemical reactions. Hierarchical Proteins: Structure of Although proteins are constructed by polymerization of only 20 different amino acids into linear chains, proteins carry out an incredible array of diverse tasks. Only when AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 2 a protein is in its correct three dimensional structure, or conformation is it able to function efficiently. A key concept in understanding how proteins work is thatfunction is derived from three dimensional structure, and the three dimensional structure is in turn specified by the amino acid sequence. The structure of proteins can be considered at four levels of organization starting with their monomeric building blocks, the amino acids as shown in Fig. 2 Fig.3 Peptide bonds (yellow) link the amide nitrogen atom (blue) of one amino acid(aa) with the carbonyl carbon atom (gray) of an adjacent one in the linear polymers known as peptides or polypeptides depending on their lengths. THE SECONDARY STRUCTURE: Fig. 2 The linear sequence of amino acids (primary structure) folds into helices or sheets (secondary structure) which pack into a globular or fibrous domain (tertiary structure). Some individual proteins selfassociate into complexes (quaternary structure) that can consist of tens to hundreds of subunits (supramolecular assemblies) They are the core elements of protein architecture. The various spatial arrangements resulting from the folding of localized parts of a polypeptide chain are referred to as the secondary structures. When stabilizing hydrogen bonds are formed between residues, parts of the backbone fold into one or more well defined periodic structures such as the alpha helix, the beta sheet also called the beta pleated sheet, and a set of turns (See Fig.4). THE PRIMARY STRUCTURE: It is the linear arrangement of the amino acid sequences present in the protein. It is nothing but the formation of a peptide bond combining the carboxylic acid moiety of one amino acid with an amino moiety of another amino acid. Thus one end of the protein has a free (unlinked) amino group (the Nterminus) and the other end has a free carboxyl group (the C-terminus). The sequence of a protein chain is conventionally written with its N-terminal amino acid on the left and its C-terminal amino acid on the right. A short chain of amino acids (20-50 amino acid residues) linked by peptide bonds and having a definite sequence is called a peptide, while longer chains are referred to as polypeptides or proteins (up to 4000 amino acid residues). Fig.3 shows the primary structure of a set of amino acids and the formation of the peptide bonds between them. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 3 together. Since the stabilizing forces are weak the tertiary structure of a protein is not rigidly fixed, but it undergoes continual and minute fluctuation. This variation in structure has important consequences in the function and regulation of proteins. There are two important substructures within the tertiary structure which are of interest to study: Fig.4 Shows the Alpha Helix which is above with the red backbone, and the beta pleated sheet below with the blue backbone. THE TERTIARY STRUCTURE: The overall folding of the polypeptide chain yields its tertiary structure. The tertiary structure refers to the overall conformation of a polypeptide chain-that is the three dimensional arrangement of all its amino acid residues. The tertiary structure is primarily stabilized by hydrophobic interactions between the non-polar side chains, by hydrogen bonds between the polar side chains and by peptide bonds. These stabilizing forces hold elements of the secondary structure- alpha helices, beta strands, turns and random coils- compactly 1. Motifs: They are particular combinations of secondary structures. In some cases, motifs are signatures for a specific function. E.g. of motifs are - the helix-loop-helix which is a calcium binding motif found in more than hundred calcium binding proteins. - zinc-finger motif: it is an alpha helix and two beta strands held together by a zinc ion. This type of a motif is most commonly found in proteins that bind to DNA or RNA such as steroid hormone receptors. - coiled coil: two or more alpha-helices orient themselves around each other in a coiled coil manner. Fig. 5 gives a visual idea of how each of these motifs looks. Fig. 5 shows various kinds of motifs commonly found in proteins. (a) two helices connected with a helix-loop-helix motif commonly found in calcium-binding and DNA-binding regulatory proteins. (b) The zinc-finger motif which is present in many DNA-binding proteins that help regulate AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 4 transcription. (c) the parallel two stranded coiled-coil motif characterized by two alpha-helices wound one around the other and is stabilized by interactions between hydrophobic side chains. 2. Domains: A domain is a compactly folded region of a polypeptide. Often a domain is characterized by some interesting structural feature, such as, an unusual abundance of a particular amino acid (e.g. a proline rich domain), sequences common to many proteins (e.g. the Epidermal Growth Factor domain) or a particular secondarystructure domain (e.g. the zinc finger domain). Fig. 6 shows the quaternary structure of haemoglobin which is a tetramer of two alpha and two beta subunits. Domains are sometimes defined in functional terms on the basis of observations that an activity of a protein is localized to a particular region along the length of the protein (e.g. DNA binding domain). Experiment: Functional domains are often identified experimentally by whittling down a protein to its smallest active fragments with the aid of proteases, enzymes that cleave the polypeptide backbone- and then checking individual fragments for particular activity. Alternatively, the DNA, encoding a particular protein, can be subjected to mutagenesis so that segments of the protein’s backbone are removed or changed. The activity of the truncated or altered protein product synthesized from the mutated gene is then monitored and serves as a source of insight about which part of the protein is critical to its function. THE QUARTENARY STRUCTURE Multimeric proteins consist of two or more polypeptides or subunits. Quaternary structure of a protein describes the number (stoichiometry) and relative positions of the subunits in the multimeric proteins (e.g. Haemoglobin is a tetramer consisting of two alpha and two beta subunits as can be seen in Fig.6) [1] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 5 ALPHA FETOPROTEIN • History • Structure • Functions HISTORY Historically alpha fetoprotein’s presence was observed in both normal fetuses as well as in abnormal conditions. In 1956 Bergstrand and Czar found a fetal component not commonly found in adults which was first detected as a post albumin migrating protein in fetal serum using paper electrophoretic techniques. Subsequently Masopust and Kotal assigned to the unknown developmental protein of Bergstrand and Czar the name “fetoprotein”. Gitlin and coworkers devised the name “alpha- fetoprotein” (AFP) for the electrophoretic alpha-1 migrating human fetal protein. Then in 1963 Abelev and co-workers found a protein that migrated in the alpha-1 region of an electrophoretogram in hepatoma bearing mice. In 1965 Tatarinov described a similar protein in sera of humans bearing hepatomas. In the early 1970s, Brock and co-workers reported elevated AFP levels in human amniotic fluids and in maternal serum that correlated with the presence of neural tube defects in the fetus. In 1972 Gitlin and co-workers demonstrated that AFP was synthesized by fetal liver and yolk sac. A subsequent study indeed demonstrated that AFP was synthesized by a multitude of tissues, especially those of gastrointestinal origin. [2] Fig 7 showing a normal electrophoretogram of human serum and the alpha-1 region STRUCTURE OF AFP protein (DBP), AFP, and alpha-albumin (alpha-ALB). [2] Mammalian AFP is a single-chain glycoprotein. Its molecular masses ranging from 66 to 72kDa and a 3%-5% carbohydrate (glycan) content. AFP is a tumor associated fetal protein classified as a member of a three-domain albuminoid gene family that currently consists of four members- Albumin (ALB), vitamin-D binding When compared to albumin, AFP is quite similar in that it also binds to and transports a multitude of ligands, including bilirubin, fatty acids, retinoids, steroids, heavy metals, dyes, flavonoids, phytoestrogens, dioxins and various other organic drugs. However, unlike ALB, high concentrations of hydrophobic ligands (such as estrogens and AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 6 fatty acids) have been reported to induce an irreversible conformational change in the tertiary structure of AFP. [2] Structurally AFP is divided into 3-domains in a U-shaped configuration. The albuminoid gene family is structurally characterized by cysteine residues that are folded into layers that form loops dictated by disulphide bridging. A hinge region is present in the Domain 2 of AFP. The hinge concept developed from the observation that HAFP has two disulphide bridges fewer than human albumin, providing it with means of molecular flexibility. [4] There also exists a carbohydrate side chain attached to asparagine-232 in AFP. [5] Fig 8 Molecular configurations of human AFP (Left) and human albumin (Right) based on the predicted secondary structures. The amino acid residues participating in the formation of alpharespectively. helices, beta- sheets, beta-turns, and random coils are indicated by The loops formed by disulfide bonding are filled in black. Stars indicate extra turns introduced in human AFP at amino acid residues 195-198 and 504-507 where the probabilities of beta turn occurrence were higher. Black squares represent the carbohydrate residues attached to asparagine-232 in AFP. The first loop at the amino terminus in AFP is formed assuming cysteines 18 and 67participate in a disulfide linkage. [5] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 7 Fig 9 Primary and secondary structure amino acid sequence of HAFP. Note that the HAFP molecule is composed of three domains in a U-shaped configuration. HAFP belongs to the albuminoid gene family, which is structurally characterized by cysteine residues that are folded into layers that form loops dictated by disulfide bridging. The hinge concept developed from the observation that HAFP has two disulfide bridges fewer than human albumin, providing it with a means of molecular flexibility. [4] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 8 FUNCTIONS OF AFP FATTY ACID BINDING DURING DEVELOPMENT Presence of fatty acids on HAFP (human AFP) was first reported in 1978, enumerating six fatty acids namely- palmitic, stearic, oleic, linoleic, arachidonic (AA), and docosahexaenoic (DHA) acids. [2] AFP plays a role in the intracellular delivery of poly-unsaturated fatty acids (PUFA’s) into developing cells. Experiment- labeled AFP was found to enter the cells via coated pits and receptosomes and to move to tubular elements of the transreticular portion of the golgi apparatus. Fatty acids bound to AFP are transferred into cells within 5 minutes at 37°C, and, following fatty acid release, AFP can be recycled back across the cell surface. In short it followed receptor mediated endocytosis. Data revealed that the fatty acids bound to AFP were mainly incorporated into cell phospholipids and that 25%-40% of the incorporated AFP was secreted and released un-degraded after 60 minutes of incubation. The AFP first binds to an AFP cell surface receptor, and then the fatty acid is endocytosed and transferred within the cell by a specific fatty acid binding protein. During pregnancy and early infancy, the biological role of HAFP in binding and trafficking of PUFA’s is now well established. Human AFP is known to both regulate and facilitate the entry of fatty acids (especially arachidonic acid and docosahexaenoic acid) into cells undergoing differentiation. HAFP reversibly binds docosahexaenoic acid with high affinity and transports the fatty acid mainly during the fetal, perinatal and neonatal periods. HAFP itself undergoes transplacental passage to the maternal circulation and tissues. Fatty Acid Binding Sites: One major fatty acid binding site for long chain fatty acids has been documented to lie between residues 210 and 227 on HAFP on domain 2. Lysine residues, especially Lys-223, appears to be essential for the fatty acid binding at this site. This is because lysine contains a free amino group which can form ionic bonds with the fatty acid carboxylic acid functionality. The fatty acid binding site at domain 1 residing at residues 36-69 shows an amino acid homology to fatty acid synthetase. Upon comparison of the domain 1 and domain 2 fatty acid binding sequences a few points of interest emerge: First it can be seen that each of the 20 amino acid sequence stretches contain three or more lysine residues, which are essential for fatty acid binding. Second, one or more lysines are located at the amino-terminal side of the sequence amino acid stretch. Third, the crucial amino acid for complexing to the carboxy group of the bound fatty acid has been identified as lysine position 13-14 amino acids from the amino terminal lysine. It must be noted that the third AFP fatty acid binding site which resides on domain 3, apparently overlaps or lies directly adjacent to the documented estrogen-binding site on RAFP (Rodent AFP), according to competitive binding reports. Binding, Spectral and immunological studies have demonstrated that conformational changes in the tertiary structures of rodent and human AFP’s can be induced by a high free fatty acid environment. The fatty acids induced a rapid and reversible conformational change in the AFP molecule. [2] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 9 Fig 10 shows the location of the fatty acid binding sites on the AFP molecule. Fig 11 the amino acid sequences of the Fatty Acid Binding Sites seen in detail. AFP IN DEVELOPING BRAIN Human fetal brain tissues obtained at autopsy and stained by immunohistochemical procedures revealed positive AFP staining in nerve cells of the cerebral wall, brain stem nuclei, and the epithelial layers of the choroids plexus. The presence of AFP within the cells of the choroids plexus suggested that the fetal protein was transudated from the blood to the cerebrospinal fluid via a cellular route across the choroids plexus epithelial layers. A proposal was thus made that the presence of AFP in the brain plays a role in neuronal differentiation and/or development. Large tissue areas and groups of cells in many regions of the developing brain, stained positively for AFP at various time intervals during development. Intracellular labeling localized AFP to the cytoplasm of the neuronal cells and extending into these cell’s axonic and dendritic extensions. AFP was found mainly in the cytoplasm of differentiating neurons at the axonal pole of the cell and in the dendritic processes of pyramidal cells. Studies in a 9 week old fetal baboon brain further demonstrated the presence of AFP in neural tube and neural crest derivatives and in the ventricles, which displayed an intracytoplasmic staining pattern. The AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 10 staining localization of the baboon AFP appeared to decline as myelination and glialcell development progressed. Uriel and coworkers proposed that the binding and transport of poly-unsaturated fatty acids by AFP could explain the presence of this fetal protein in the developing nervous system. As it is known that the myelin sheath of the neuronal cells are made of a lipid bilayer containing protein myelin, and this bilayer requires PUFAs for development. [2] AFP IN REPRODUCTIVE FLUIDS Alpha fetoprotein has long been detected in the biological fluids of the reproductive tract. Electrophoretic studies of oviductal fluids have demonstrated that the mammalian oviduct secretes a variety of proteins such as ALB, Beta-Globulins, and AFP. The production of proteins in the human oviductal fluid was found to be under hormonal control and the levels of such protein secretion were found to correlate with the estrogen peak of the menstrual cycle. The proteins derived from human oviductal fluid have two sources: firstly proteins originating from serum transudation, and secondly those which are synthesized and secreted by the uterine tubule mucosa. One such secreted protein designated as human oviductin-1 (HOV-1) displayed a molecular mass of 54kDa and an isoelectric pH of 4.5 and contained a carbohydrate moiety. Experiment: fresh donated human sperm were incubated with 1) a mixture of Human Oviductal Fluid (HOF) specific proteins 2) HOV-1 Then using indirect immunofluorescence, the investigators studied the ability of the HOF proteins to bind to the human sperm. While the mixture of human oviductal fluid proteins bound diffusely over the entire surface of the sperm, cell HOV-1 binding was restricted to the head region of the sperm. The investigators stated that the HOV-1 protein acted as an acrosome-stabilizing factor, serving to prevent premature acrosome activation. So why are we talking so much about HOV-1? In subsequent purifications of HOV-1 protein, determination of amino acid and carbohydrate composition of HOV-1 confirmed that HOV-1 was identical to HAFP. However, it’s molecular mass suggested that HOV-1 was a cut or truncated form of AFP. Thus HOV-1 was a globular, noncollagenous protein with carbohydrate attachment via an N-glycosidic linkage between N-acetyl-glucosamine and asparagines as in the case of HAFP. It was proposed that secreted AFP as a constituent protein of Human oviductal fluid may serve to mediate sperm survival and motility by decreasing the response of the acrosomal reaction so as to prolong sperm viability and function. [2] INTERACTION OF ESTROGEN DEVELOPMENT AFP WITH DURING In human AFP two estrogen binding regions are present. The first region lies between amino acids 423-444, which represents a major hydrophobic binding pocket on HAFP. The second region is from amino acids 445480. Both the regions display overlapping binding sites for fatty acids, diethyl stilbesterol, retinoids, warfarin, coumarin and other drugs (found by competitive binding studies). Site 1 estrogen binding site on the HAFP is a high affinity binding site located at residues 424-439. The 5 amino acids crucial to binding on RAFP were found to be glycine-425, methionine-427, isoleucine-430, alanine-432, and threonine-433. These amino acids match precisely on the MAFP residues glycine-428, methionine-430, isoleucine-433, alanine-434, and threonine435. In humans three out of the five of these amino acids show substitutions namely glycine to arginine, isoleucine to threonine, and serine to alanine. Amino acid sequences of human and rodent AFPs were compared to an estrogen binding region on the human estrogen receptor (HER: residues 419-431). It was seen that 4 out of 5 of these crucial amino acids for binding estrogen on rodent AFP matched and aligned to those on HER. The remaining hydrophobic amino acids in human and rodent AFP are also present in HER, namely AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 11 the leucines and valine. In human AFP multiple (five) alanines have replaced many of the crucial amino acids present in both RAFP and HER. However a single alanine retained in all species is probably significant for binding. Thus, this 15-amino acid region on the various AFPs seems to represent the high-affinity binding site for rodent and to a lesser extent human AFP. The Site 2 estrogen binding site on the AFP molecule is a low affinity binding site located from amino acids 454-468 on the HAFP. The site is highly hydrophobic (leucines, isoleucines, alanines). Various AFP molecules display a common cysteine, unlike HER, but otherwise they show similar arginine and histidine positioning. In both the AFPs and HER, the composition and placement (position) of leucines, isoleucines, arginines, and histidine as the dominant amino acid are shared. The HER segment is devoid of glycines and cysteine in its secondary binding site, in contrast to human and rodent AFPs. Cysteine does not seem to be required for estrogen binding at the secondary site. Thus AFP site 1 is primarily committed to estrogen binding and ligation. And it is proposed that site 2 is thought to serve as a docking site for proteins of heat-shock proteins (HSPs) family, such as HSP-70 and HSP-90 as in HER. Such docking sites for HSP’s are also known to be involved in protein folding/unfolding activities. Misfolded proteins are believed to be the cause of various diseases such as Alzheimer’s, and neural tube defects such as Down’s syndrome. [2] Fig. 12 Shows the Estrogen binding sites present on the AFP molecule AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 12 Fig. 13 Shows amino acid sequences of the estrogen binding sites present on HER(human estrogen receptor); RAFP (rat alpha-fetoprotein); MAFP (mouse alpha-fetoprotein); the numbers indicate amino acid sequence numbers of the protein. [2] AFP AND INSULIN: In the late 1990’s studies of AFP-derived peptide fragments from the amino-terminal side of AFP Domain 1 were found to have glucose/insulin related activities. A synthetic peptide, duplicating amino acids 13-19 (LDSYQC) of HAFP Domain 1, was reported to influence glucose uptake by human red blood cells is a clinical laboratory. The LDSYQC peptide was found to stimulate the entry of glucose into red blood cells from insulin-dependant diabetic children invitro. It is of interest that an amino acid sequence on the insulin alpha chain (residues 17-21) has been found to be structurally homologous to the AFP-derived LDSYQC sequence. [2] Fig. 14 shows the insulin like segment present in Domain-1 of the AFP molecule. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 13 AFP AND THYROID HORMONES In developing and neoplastic cell cultures AFP and thyroid hormones display an inverse relationship in that cells exposed to increasing concentrations of triiodothyronine (T3) show decreased secretion of measurable AFP. In vitro studies have demonstrated that thyroxine (T4) treated newborn rodents displayed depressed serum AFP levels that were attributed to blockage of hepatic AFP synthesis. In human newborn studies, a reciprocal relationship between serum AFP levels and congenital hypothyroidism also exists. Thus AFP and thyroid hormone levels have consistently displayed an inverse relationship in both invitro and in vivo model systems. [2] AFP AND CELL SEQUENCES ADHESION Intercellular and cell-extracellular matrix (ECM) interactions are of great significance in many biological processes of growth, apoptosis, differentiation and cell migration, as well as cancer cell invasion and dissemination or metastasis. These functions are mediated by many cell adhesion molecules and cell surface receptors. Several families of adhesion molecules have been identified and their synthesis and expression on the cell membrane studied in relation to the invasive and metastatic phenotype. Results of studies on tumor metastasis have demonstrated that cell adhesion plays an important role in various steps of the metastatic cascade and that dysregulation of adhesion mechanisms contributes to the formation of metastasis. On the other hand, certain adhesive interactions may diminish the metastatic process. For example, adhesion molecules that promote homotypic cell adhesion among homotypic tumor cells in a primary tumor site will likely diminish the metastatic potential. In that context, down regulation of these adhesion molecules has been shown to correlate with a higher propensity of tumor cells to detach from the primary site and to spread. On the other hand up regulation of other adhesion molecules correlates with a higher potential of tumor cells to metastasize. Preferential adhesion of metastatic tumor cells to vascular endothelial cells of certain organs has been demonstrated in experimental tumors and it may be explained in part by the organ-specific expression pattern of some adhesion molecules. The second domain of AFP contains short peptide sequences common to extra cellular matrix(ECM) proteins (like laminins, fibronectin, collagen etc) bearing cellular adhesion motifs (CAMs). These short sequences are not found commonly in Albumin, Alpha-Albumin and DBP. Adhesive macromolecules (such as laminins, fibronectin etc) have a potential utility in developmental state and disease state involving growth, developmental state and disease state involving growth, differentiation, cell-migration and tumor metastasis. Using synthetic peptides derived from ECM’s, the functional significance of such short signal peptide sequences has been identified from one or more domains of these macromolecules. On inspection of the matched peptide sequences, it is observed that frequent amino acid matches of AFP with CAM-like sequences occur in the second domain of AFP (amino acids 190394). The physiological functions of adhesion sequences of CAMs and the cell types involved are listed in the table below. Some of these activities include cell adhesion, migration, differentiation, growth, neurite outgrowth, tumor spread, enzyme activity, angiogenesis, and heparin, fibrin, collagen interactions. Some adhesion proteins are known to contain two signals of opposing activities e.g. YIGSR and SIKVAV of laminin. This demonstrates that additional fine-tuning controls are in place on the same molecule to even further regulate adhesion activities. Two such sites may not be simultaneously available on a protein due to ligand and or conformational masking, and could progressively be unveiled during a cascade such as clotting anticlotting network. Such a diverse array of peptide recognition signals on AFP suggests that AFP might share functional properties with adhesion molecules. It is proposed that AFP may play AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 14 a role in fine-tuning the architectural interstitial growth patterns in developing organisms. However all adhesion sites may not be concurrently accessible since AFP is thought to express different functions at various developmental stages. Some sites may be unmasked or made conformationally available during progressive stages of embryonic and fetal development depending upon ligand binding and stress condition of the protein. [3] Fig. 15 shows the locations of some cell-adhesion motif sequences present in the AFP molecule. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 15 Table 1 Cell Adhesion Sequences in Extracellular Matrix Proteins Compared With Sequences on AFP, Albumin, Vitamin-D Binding Protein, and Alpha Albumin. [3] THE GROWTH INHIBITORY PEPTIDE (as a temporary halt may be required until fetal homeostasis is achieved in the stress/shock condition). Alpha fetoprotein known largely as a growth promoting agent, also possesses a growth inhibitory motif recently identified as an occult segment of the molecule. The 34 amino-acid stretch termed the growth inhibitory peptide (GIP) has been chemically synthesized, purified and characterized. The GIP segment lies buried in a molecular crevice and can be exposed following a conformational change in HAFP. The hidden segment is an amino acid stretch on AFP that can potentially be exposed by extreme ligand concentrations and possibly by stress or shock conditions during fetal development HISTORY OF THE GIP In the early 1970’s and 1980’s extensive studies were carried out to determine the growth regulatory properties of AFP. Studies involving anti-AFP antibodies were carried out. When mouse AFP was used as a control for the antibody experiments, it was found that the fetal protein, incubated with estradiol (E2) in 130-fold molar excess was capable of growth regulation in the prepuberal mouse uterus. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 16 In 1983 the first report of an AFP-E2 complex-induced inhibition of rat mammary tumors appeared. During 1983-1985 reports detailed and documented use of immature mouse uterine bioassay to measure and quantify the inhibition of growth in normal tissues. At the same time, E2 addition/depletion experiments showed that minute amounts of AFP complexed with E2 were capable of blocking E2-supported growth of MTW9A breast tumors in rats. It was further proposed that AFP might be present in adult cancer tissues as a means to regulate growth. In 1990 it was found that both mouse AFP and HAFP activated by complexing with E2 were capable of growth inhibition of MCF-7 human breast cancer xenografts. During this period an AFP “cassette” concept was proposed in which isolated stretches of amino acids on the AFP molecule were predicted to possess biological activities of their own. Thus scientists isolated many peptides from the AFP molecule and tested them for biological activities. These studies led to the identification, chemical synthesis of a 34-amino acid segment from the third domain of HAFP which was found to be a disrupter of endocrine associated growth in the rodent bioassay. Subsequent findings served to confirm that this amino acid stretch was an active site of E2 regulatory growth on the native AFP molecule. This 34 amino acid sequence was then designated as the Growth Inhibitory Peptide (GIP). [4] STRUCTURE OF THE GIP The 34-amino acid GIP was also termed the P149. Synthetic fragments of which have been designated as P149a (the amino terminal 12 amino acids), P149b (the hydrophobic midpiece consisting of 14 amino acids) and P149c (an eight amino acid peptide located near the carboxyl terminus). The molecular mass of the peptide determined by mass spectrophotometry was found to be 3573Da. The 34 amino acid sequence was found to exhibit complex aggregation behavior. Upon solubilization, the peptide formed trimers; however, at high concentrations (8mg/ml) the trimers clustered into large aggregates. At 0.2mg/ml the trimers slowly dissociated to form dimers (by intrapeptide disulphide bonds). It was found that the trimeric form was active and showed growth inhibitory activity while the dimeric form was lacking growth inhibitory activity. [4] Fig. 16 the three domains of human alpha-fetoprotein are shown in a bar configuration showing the 590 amino acid full-length protein. The telescoping segment displayed, from amino acids #445–477, is depicted in its single letter code amino acid sequence together with the three fragment constituents of P149 comprising P149a, P149b, and P149c. [6] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 17 Fig. 17 Shows the GIP segment highlighted on the AFP molecule. Fig. 18 shows the cyclized form of the GIP segment. EFFECT OF GIP ON MOUSE ASCITES MAMMARY TUMOR The growth suppression of the mammary ascites tumor by P149 was determined in non-estrogen supplemented assay which measured tumor cell growth and ascites accumulation of 6WI-1 mammary tumors transplanted in mice (isografts). Tumor cells inoculated were 0.3x106 cells. Mice inoculated with 6WI-1 mammary cells were injected with a previously determined optimal dose of 1.0 µg peptide per day or saline for 11 days. On day 12 following inoculation, the total accumulated ascites fluid volume and tumor cells in the peritoneal cavity of each animal were harvested and the total ascites cell count was determined. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 18 It was observed that the dose of 1µg/day of P149 for 11 days suppressed the tumor associated body weight gain and nor was there any increase in the ascites fluid volume. These mice further went on to live an additional 30 days before finally they were sacrificed. The tests were then also carried out on various peptide fragments of the P149 peptide. It was found that the P149 or its fragment P149c, but not P149a or P149b significantly suppressed accumulation of both cells and ascites. [6] Fig. 19 The serial transplantation in NYALAR mice of isograft 6WI-1 murine mammary tumor ascites cells is depicted at 3 different dose inocula. A1 is non-tumor treated animal. B1 is the ascites laden breast tumor inoculated mouse which has 0.3×106 cells inoculated. C1 is the group of mice which has the ascites breast cancer cells inoculated but is treated with 1.0 µg of peptide per day. [6] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 19 EFFECT OF GIP ON CELL LINES An experiment was carried out in which the GIP was challenged against 60 different cell culture lines representing a variety of human cancers. In a 6 day proliferation assay, the P149 peptide was seen to suppress growth in 38 out of 60 cancer cell lines, representing nine different cancer cell types including prostate, breast, and ovarian cancers. It was seen that the GIP peptide was seen to be active against both E2dependant and E2-independent growth. The table below shows the list of human tumor cell cultures and the growth suppressive screening results of HAFPderived peptide (P149) [4] Table 2 the growth-suppressive (cytostatic) screening results of HAFP-derived peptide (P149) for multiple types of human tumor cell cultures [4] PROPOSED MECHANISM OF ACTION Although the exact mechanism of action has not as yet been elucidated, a proposal has been made as to how GIP acts as a growth inhibitor. Following administration, E2 serves as a growth-promoting agent in immature tissues such as the uterus, or in transformational E2-dependant breast cancer cells. Likewise, administration of the GIP results in homing of this peptide to tissue areas of up-regulated growth, as seen in estrogen-targeted cells 24 hours later. By means of receptor-mediated endocytosis via integral cell membrane proteins, the GIP might gain access through the plasma membrane while encountering the membrane-associated caveolae. Since the estrogen receptor (ER) has been reported to localize to the inner membrane portion of the AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 20 caveolae, it is conceivable that the GIP binds to the membrane-tethered ER, a peptide-to-ER binding interaction that has been demonstrated in vitro. It has been shown that ER alpha, soon after translation on the ribosome, translocates to the plasma membrane caveolae and resides there, attached as a complex with the SHc adaptor/scaffold protein. The caveolae or lipid rafts have been shown to be essential for insulin-like growth factor-1 receptor (IFG1R) signaling during the induction of cell activation, proliferation, and differentiation. The IGF-1 receptors were reportedly located in caveolar plasma membranes and are intimately involved in signal transduction cascades. Binding of the GIP to the ER could involve the steroid receptor segment region that recognizes the co-activator NRbox. It is conceivable that signal transduction, especially via MAP kinases, could also be blunted at this point. Alternatively, once the GIP has complexed with the ER, a cytoskeletal shape change could occur due to the juxtaposition of the ER with tubulin and actin. The ER-GIP complex might then be transported through the cell via the cytoskeletal network to the perinuclear endoplasmic reticulum (EPR) and the entire complex could position itself near the entrance of the nuclear pore. The EPR is known to be contiguous with the perinuclear region of the cytoplasm. Immunofluorescent micrographs presently demonstrate such a perinuclear location for the GIP in the uterine tissue sections. Since the GIP has sequence identity to both nuclear Lamin B and an inner-membrane protein of the nuclear pore, it is possible that the ER/GIP complex can be translocated and positioned at or near the nuclear pore channel to impede (gate) further entry of complex into the nucleoplasm. In effect, the ER would be denied contact with the transcriptional machinery of the nucleus, thus inhibiting immediate-early gene activation. [7] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 21 Fig. 20An artist’s conception of a possible cell penetration route of growth inhibitory peptide after approaching the plasma membrane at an angle. During the cell membrane insertion process, peptides are capable of interacting with integral membrane receptors/proteins to affect signal transduction. In this proposed model, P149 (shown along the extended arrow) might encounter and bind the estrogen receptor-alpha (ER-alpha) tethered to the insulin growth factor-1 receptor (IGF-1R)-SHc adaptor/scaffold complex. When incoming E2 binds normally to this tripartite complex via ER-alpha, the ER-alpha is released, after which it attaches to the cytoskeleton (grey background threads) for transcytosis to the perinuclear region of the endoplasmic reticulum and enters the nuclear pore region (see text). The ER-alpha-GIP bound complexes might blunt this response, either at the cell membrane or by plugging the nuclear pore. Heterochromatin = nuclear material consisting of DNA, histones, and associated proteins. Circled P = phosphorylation sites; MAPK= mitosis activated protein kinases; p110 and p85= phosphoinositol-3 kinase proteins; Lamins A, B, C = nucleoskeletal proteins near the nucleopores; HSP- 90, 70 = heat shock proteins that interact with the cytoskeleton. [7] AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE 22 CONCLUSION Alpha-fetoprotein is truly an amazing protein which is found in our bodies. The myriad of functions it performs in our bodies have still only been discovered in part. Many more theories have been released on it’s uses in the human body, but who knows what more functions this molecule performs. Further research behind this molecule is thus mandatory in order to understand the complete working of the molecule inside the body. The discovery of the GIP from the AFP has lead to the opening of the use of AFP and it’s peptides in development of cures for cancer. Furthermore the structure of the GIP itself may provide vital information for development of new drugs for a cure for cancer. I would like to conclude by saying that this truly is a superman molecule which finds use in almost every area of molecular biology. REFERENCES 1. Harvey Lodish [et al.]: Protein Structure and Function. In: Molecular Cell Biology (5th ed.). New York, W. H. Freeman and Company, 2003, p.59. 2. Mizejewski GJ. Biological Roles of AlphaFetoprotein During Pregnancy and Perinatal Development. Exp Biol Med 229:439-463, 2004. 3. Mizejewski GJ. Alpha-fetoprotein structure and function: relevance to isoforms, epitopes, and conformational variants. Exp Biol Med 226:377-408, 2001. 4. Mizejewski GJ, MacColl R: Alphafetoprotein growth inhibitory peptides: Potential leads for cancer therapeutics. Mol Cancer Ther 2: 1243–1255, 2003. 5. Morinaga T, Sakai M, Wegmann TG, Tomoaki T. Primary structures of human AFP and its mRNA. Proc Natl Acad Sci USA 80:4604– 4608, 1983. 6. Meuhlemann M., Miller KD, Dauphinee M, Mizejewski GJ. Review of Growth Inhibitory Peptide as a Biotherapeutic agent for tumor growth, adhesion, and metastasis. Cancer and Metastasis Reviews 24:441-467,2005. 7. Mizejewski G, Smith G, Butterstein G: Review and Proposed Action of alphafetoprotein growth inhibiting peptides as estrogen and cytoskeletal-associated factors. Intl Journal Cell Biology 28: 913–933, 2004. AN INTRODUCTION TO ALPHA-FETOPROTEIN AND THE GROWTH INHIBITORY PEPTIDE