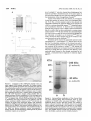
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Protein phosphorylation wikipedia , lookup
Cytokinesis wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Mechanosensitive channels wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
SNARE (protein) wikipedia , lookup
Magnesium transporter wikipedia , lookup
Protein moonlighting wikipedia , lookup
Signal transduction wikipedia , lookup
Cell membrane wikipedia , lookup
Endomembrane system wikipedia , lookup
Photoreceptor Membrane Proteins, Phototransduction, and Retinal Degenerative Diseases The Friedenwald Lecture Robert S. Molday R od and cone photoreceptor cells are specialized neurons of the vertebrate retina that function in the primary events of vision. Rod cells are highly sensitive to light and operate under dim lighting conditions. Cones are less sensitive and function in bright light and in color vision. Both photoreceptors are elongated cells consisting of several morphologically and functionally distinct regions (Fig. 1). The photoreceptor outer segment located adjacent to the retinal pigment epithelial (RPE) cell layer is a specialized compartment uniquely designed to carry out phototransduction, i.e., the capture of light and its conversion to an electrical signal. This membrane-rich organelle undergoes a continual renewal process in which newly synthesized membrane is added at the base of the outer segment, whereas packets of outer segment membrane are phagocytized at the distal end by the RPE cells. '"3 A thin, nonmotile cilium links the outer segment to the inner segment, a cellular compartment that contains the mitochondria, endoplasmic reticulum, golgi apparatus, and other subcellular organelles. Adjoining the inner segment is the cell body containing the nucleus. This region further extends into the synaptic region where the electrical signal generated in the photoreceptor cell is transmitted to other neurons of the retina. Photoreceptor outer segments have been extensively studied at a cellular level/'"8 In the rod cell, the outer segment appears as a cylindrical structure up to 60 /xm in length and 1.5 to 2 jam in diameter for mammals and 6 to 8 jam in diameter for amphibians. The cone outer segment is generally shorter and often is tapered or conical. The outer segment consists of hundreds of flattened disk membranes assembled in an ordered axial array (Fig. 1). In rod cells, the stack of closed disks is surrounded by a separate plasma membrane over the entire length of the outer segment except at its base. Here, newly formed disk membranes evaginate from the ciliary membrane to form a folded membrane system.8 Each rod outer segment (ROS) disk consists of two closely spaced lamellar membranes circumscribed by a hairpin From the Department of Biochemistry and Molecular Biology, University of British Columbia, Vancouver, BC, Canada. Supported by Grant EY02422 from the National Eye Institute, Bethesda, Maryland; The RP Research Foundation-Fighting Blindness, Canada; the Medical Research Council of Canada; and an Alcon Research Institute Award, Fort Worth, Texas. Reprint requests: Robert S. Molday, Department of Biochemistry and Molecular Biology, 2146 Health Sciences Mall, Vancouver, BC, Canada V6T 1Z3. E-mail: [email protected]. Investigative Ophthalmology & Visual Science, December 1998, Vol. 39, No. 13 Copyright © Association for Research in Vision and Ophthalmology loop called the rim region. The continuous disk membrane separates an intradiskal space or lumen from the cytoplasm. The perimeter of the rod disk is interrupted by one or more incisures that penetrate toward the center of the disk. Cone outer segments also consist of a stack of disk membranes, but unlike ROS disks, cone disks remain open and do not contain incisures.9'10 Fibrous elements extend from the rim regions of disks to adjacent disks and to the plasma membrane.""13 This cytoskeletal-like system serves to maintain the precise distances between adjacent disk and plasma membranes and stabilize the outer segment structure. ROS Isolation and Characterization Although ultrastructural features of ROS had been described in considerable detail by the early 1970s, relatively few studies dealt directly with the molecular composition of this lightsensitive organelle. This was due largely to the lack of a pure, well-characterized ROS preparation and to technical difficulties inherent in the molecular analysis of biological membranes, and in particular membrane proteins. A significant advancement came in 1974 when David Papermaster, working as a postdoctoral fellow in William Dreyer's laboratory at California Institute of Technology in Pasadena, developed a highly pure preparation of bovine ROS.14 This procedure, based in part on earlier protocols,1516 takes advantage of the fragile nature of the connecting cilium, the stability of intact ROS in sucrose solutions, and the low density of ROS relative to other subcellular organelles. Briefly, retina tissue immersed in sucrose solution is gently agitated to detach the ROS. Differential velocity centrifugation and sucrose density gradient sedimentation is then used to isolate ROS, largely free of contaminating subcellular organelles. Minor modifications to this procedure have been introduced over the years, but in general these preparations exhibit similar protein and lipid compositions. A typical profile of ROS proteins fractionated on a sodium dodecyl sulfate (SDS) polyacrylamide gel is shown in Figure 2 (lane a). Rhodopsin represents the major stained band, accounting for more than 70% of the total ROS protein. Numerous less abundant proteins of various molecular weights are also observed. Isolated ROS in sucrose solutions retain their soluble and weakly associated membrane proteins. Herman Kiihn, also a former postdoctoral fellow at Cal-Tech and later a research scientist at the Neurobiology Institute in Jiilich, Germany, developed several procedures to selectively extract soluble and 2493 2494 Molday JOVS, December 1998, Vol. 39, No. 13 Incisure Disks Rod outer segment plasma membrane Plasma Membrane- \ Rod outer segment Connecting Cilium Golgi complex Rod inner segment Endoplasmic reticulum > Cell body Synaptic terminal Synaptic vesicles 1. Left: schematic diagram of a rod photoreceptor cell. Right: diagram of a rod outer segment. Adapted with permission from Young RW. Visual cells. Set Am. 1970;223:81-91- FIGURE weakly associated membrane proteins for biochemical analysis. 1718 One such procedure involves hypotonic lysis of darkadapted ROS followed by centrifugation to separate the supernatant fraction from the membrane pellet. 1719 As shown in Figure 2 (lanes b, c), a distinct protein profile is observed for each fraction. The supernatant contains numerous proteins ranging in molecular mass from approximately 10 kDa to more than 100 kDa. Many of these proteins have been characterized by various laboratories and shown to function as key enzymes and regulatory proteins in phototransduction and metabolism (Table 1). The membrane fraction of ROS is dominated by rhodopsin. In addition, numerous less abundant membrane proteins are observed in the range of 30 kDa to 240 kDa (see Table 2). Separation of ROS Disk and Plasma Membranes My laboratory has focused primarily on the molecular analysis of integral membrane proteins of ROS and elucidation of their role in phototransduction, outer segment structure and metabolism, and retinal degenerative diseases. As a first step, it was important to analyze the protein composition of the ROS disk and plasma membrane. One view at the time held that the two membranes are identical in composition. This was supported by electron microscopic studies showing that the disk and plasma membranes arise from the same newly formed membranes at the base of the outer segment8 and immunocytochemical studies revealing the presence of high levels of rhodopsin in both membranes.20"22 Furthermore, several laboratories reported that both the disk and plasma membrane contain cGMP-gated channel activity and Na/Ca exchange activity.23"25 In contrast, biochemical labeling studies indicated that several proteins present in the ROS plasma membrane are absent in disks, suggesting that the two membranes may differ in protein composition.26'27 To resolve this issue, we developed a ricin-gold affinity density perturbation method that can effectively separate the plasma membrane, constituting ~5% of the ROS membrane, from the more abundant disk membranes.19'28 In this procedure, isolated ROS are first treated with neuraminidase to remove terminal sialic acid residues on surface sialoglycopro- The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 kDa 205 - 97 - 68 - 45 - - PDE(a,p) .^ Rhodopsin Kinase \ ^ Arrestin _ Rhodopsin I _- Transducin i a FIGURE 2. Sodium dodecyl sulfate (SDS) gel electrophoresis of bovine rod outer segment (ROS) proteins. Dark adapted, purified ROS (lane a) were lysed in hypotonic buffer, and the membrane fraction (lane b) was separated from the soluble fraction (lane c) by high-speed centrifugation. Fractions were run on a 9% SDS polyacrylamide gel and stained with Coomassie blue. Bands corresponding to rhodopsin, phosphodiesterase sub-units (PDEa,|3), rhodopsin kinase, arrestin, and transducin (a- and j3-subunits) are indicated.19 teins.27 Exposed galactose residues are then specifically labeled with the plant lectin ricin conjugated to gold particles. The small uniform gold particles (~10 nm diameter) are physically dense and therefore, increase the density of the plasma membrane to which they bind. They are also electron dense and serve as excellent visual markers for electron microscopy (EM). The gold-labeled ROS are lysed in hypotonic buffer to produce fragments consisting of unlabeled disks radiating from the cytoplasmic surface of the gold-labeled plasma membrane. Trypsin is then used to detach the disks from the plasma membrane, and sucrose gradient centrifugation is used to separate the unlabeled disks from the labeled plasma membrane. The fractionation procedure, as elegantly carried out by Laurie Molday, is documented in the electron micrographs in Figures 3A, 3B, 3C, and 3D. SDS gel electrophoresis and functional assays indicate that disk and plasma membranes differ in protein composition1928 (see Fig. 5A), Although both membranes contain rhodopsin as the major membrane protein, many proteins present in the plasma membrane are absent in disk membranes (Table 2). Furthermore, the plasma membrane exhibits cGMP-gated channel activity29 and Na/Ca-K exchange activity,30 whereas disk membranes do not. Differences in lipid composition between the disk and plasma membrane have also been reported.31-32 MONOCLONAL ANTIBODIES TO ROS MEMBRANE PROTEINS To begin to identify, characterize, and localize photoreceptor membrane proteins, highly sensitive and specific reagents are 2495 required. Monoclonal antibodies are particularly valuable since they typically bind to a well-defined site or epitope consisting of four to eight amino acids, can be produced in unlimited quantities, and have broad applications with highly sensitive immunochemical techniques. For example, monoclonal antibodies can be used to (1) detect and identify proteins by western blot analysis; (2) isolate proteins and peptides by immunoaffinity chromatography; (3) map the cellular and subcellular distribution of proteins by light and EM; (4) screen cDNA expression libraries for cloning and sequence analysis; (5) analyze membrane protein topology; (6) resolve proteinprotein interactions; (7) identify specific cells for developmental and transplantation studies; and (8) detect protein expression in gene therapy protocols. Monoclonal Antibodies to Rhodopsin Our studies on the generation and characterization of monoclonal antibodies began in the early 1980s. Using ROS as an immunogen, we were able to generate a panel of antibodies to rhodopsin and other ROS membrane proteins.33 Our initial studies focused on the analysis of anti-rhodopsin antibodies.33"39 In collaboration with Paul Hargrave at the University of Florida and Bob Hodges at the University of Alberta, we were able to precisely map the epitopes for a number of anti-rhodopsin antibodies.37"39 Four antibodies have been widely used in studies carried out in my laboratory and in other research laboratories throughout the world. These include the rho4D2 and rho3A6 antibodies generated by David Hicks, a former postdoctoral fellow in my laboratory,21 and rholD4 and rho4B4 antibodies produced by Don MacKenzie, a former graduate student.35"37 The rho4D2 antibody, directed against a highly conserved epitope near the N terminus of rhodopsin, cross-reacts with rhodopsin from a broad range of vertebrate species.21'33'39 It has been widely used to study the cellular and subcellular distribution of rhodopsin in rod cells and as a rod cell-specific marker in retinal developmental and transplantation studi e s 2],33,<fo,<n ^ n e rnolrj>4 antibody reacts with the conserved eight amino acid C terminus of rhodopsin.57'38 This antibody has been routinely used to purify rhodopsin from ROS and heterologous cells for structure-function analysis.'""43 The rho4B4 antibody specifically binds to an internal loop connecting helices 4 and 5 and has been used to confirm the organization of rhodopsin in disk membranes.33'34'30" Finally, the rho3A6 antibody shows more limited cross-reactivity, binding to bovine and human rhodopsin, but not mouse rhodopsin.36'38 This property has been used effectively to monitor the expression and distribution of human rhodopsin in transgenic mice that serve as models for retinitis pigmentosa and other retinal Monoclonal Antibodies to Other ROS Proteins As part of our studies, we have generated a library of monoclonal antibodies against other ROS disk and plasma membrane proteins including peripherin/rrfs, rom-1, the cGMP-gated channel, guanylate cyclase, the Na/Ca-K exchanger, glyceraldehyde-3-phosphate dehydrogenase, ABCR/RIM protein, and others (see Table 2). Applications of some of these antibodies in the identification, characterization, and localization of specific photoreceptor proteins are described below. 2496 TABLE Molday IOVS, December 1998, Vol. 39, No. 13 1. Partial List of Soluble and Membrane-Associated ROS Proteins Protein Approximate Mr Phosphodiesterase Function Phototransduction a-subunit jS-subunit y-subunit 8-subunit Rhodopsin kinase Arrestin p44 arrestin variant Transducin a-subunit j3-subunit y-subunit RGS protein Phosducin G lyceraldeliyde-3-phosphate dehydrogenase Other glycolytic enzymes Creatine kinase Phosphatase 2A Guanylate kinase Nucleotide diphosphokinase Hexose monophosphate enzymes 88,000 84,000 11,000 17,000 68,000 48,000 44,000 f-GARP t-GARP Recoverin 65,000 32,000 26,000 Calmodulin 20,000 GCAP1 and GCAP2 24,000 Pyrophosphatase Actin 36,000 45,000 Tubulin Myosin Protein kinase C Others 205,000 85,000 39,000 37,000 8,000 57,000 33,000 38,000 43,000 38,000 54,000 Rhodopsin phosphorylation Rhodopsin inactivation Rhodopsin inactivation Phototransduction Activator of transducin GTPase activity Interaction with transducin subunits Glucose metabolism and energy production Glucose metabolism and energy production ATP regeneration Regulation Nucleotide metabolism Nucleotide metabolism Glucose metabolism and regeneration of NADPH Unknown Unknown Calcium-dependent modulator of rhodopsin kinase Calcium-dependent modulator of the cGMPgated channel Calcium-dependent modulator of guanylate cyclase Pyrophosphate hydrolysis Cytoskeletal protein localized at the base of the outer segment Component of microtubules Connecting cilium Protein regulation Mr, molecular weight; GARP, glutamic acid-rich protein. PHOTOTRANSDUCTION AND THE CYCLIC GMPGATED CHANNEL OF ROD PHOTORECEPTORS The cGMP-gated channel plays a central role in phototransduction by controlling the flow of Na+ and Ca2+ ions into the outer segment in response to light induced changes in intracellular cGMP concentrations (Fig. 4). 46 -' 18 Briefly, in darkadapted rod cells, Na+ and Ca2+ ions flow into the ROS through cGMP-gated channels maintained in their open state by a relatively high concentration of cGMP. K+ ions flow out of the inner segment through voltage-gated K+ channels, thereby completing a dark current loop. Na+ and K+ gradients are maintained by an active Na,K ATPase localized in the plasma membrane of the inner segment, and the Ca2+ concentration in the outer segment is kept at ~400 nM by the balanced efflux of Ca2+ through the Na/Ca-K exchanger in the ROS plasma membrane. Under these conditions, the rod cell is in its depolarized state (—40 mV), and there is a constant release of the glutamate transmitter from the synaptic region of the photoreceptor cell.49 Photoexcitation is initiated when a photon converts the ll-cis retinal chromophore of rhodopsin to its all-trans isomer. This reaction leads to the formation of Meta II rhodopsin or R* and activation of the visual cascade.30"53 R* catalyzes transducin activation via the exchange of GDP for GTP on its a-subunit (Ta). This in turn leads to the activation of phosphodiesterase (PDE) and the hydrolysis of cGMP to 5'-GMP. The decrease in intracellular cGMP causes the cGMP-gated channels to close and the rod cell to become hyperpolarized. Under this condition, glutamate release at the synaptic region of the rod cell is inhibited. The closure of cGMP-gated channels also causes Ca2+ levels in the outer segment to decrease since the Na/Ca-K exchanger continues to extrude Ca2+ from the outer segment. After photoexcitation, the photoreceptor cell returns to its dark state by the shutdown of the visual cascade system and resynthesis of cGMP.51'54 Rhodopsin is inactivated by ATPdependent phosphorylation at its C terminus and the subsequent binding of arrestin. Transducin and PDE are inactivated by the hydrolysis of GTP to GDP on Ta by its intrinsic GTPase activity. Guanylate cyclase, the enzyme responsible for the The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 TABLE 2497 2. Major ROS Plasma Membrane and Disk Membrane Proteins Protein Plasma membrane proteins Rhodopsin cGMP-channel a-subunit Approximate Quantity (%) Mr 36,000 (38,000) 60 63,000 (79,600) 7 Function Phototransduction Monoclonal Antibodies RholD4 Rho4D2 Phototransduction PMclDl PMc6E7 /3-subunit 240,000(155,000) 7 Ca 2+ homeostatis 50,000 4 Glucose transport 36,000 (38,000) 85 Phototransduction Guanylate cyclase (RetGCl) 112,000 1 Phototransduction Peripherin/rds 35,000 (39,000) 4 Outer segment structure Rom-1 37,000 (37,000) Na/Ca-K exchanger Glucose transporter 230,000 (130,000) Others Disk Membrane Proteins Rhodopsin ABCR/RIM Retinol dehydrogenaset Others 220,000 257,000 33,500 Outer segment structure 3 Transporter? Retinal reduction PMs5El 1 PMblC9 PMe2D9 PMelB3 Polyclonal antibody RholD4 Rho4D2 GC16G7 GC12H6 Per2B6 Per3B6 RomlD5 RomlC5 Rim3F4 mAbA.. ROS, rod outer segment; Mr, molecular weight estimated by sodium dodecyl sulfate gel electrophoresis. * Values in parentheses are determined from sequence. t Only the cone retinol clehydrogenase (retSDRl) has been cloned183; localization in ROS has not yet been determined. synthesis of cGMP from GTP, is activated by the decrease in intracellular Ca 2+ after photoexcitation, a process that is mediated by GCAP proteins.55"57 As the cGMP concentration increases, the cGMP-gated channels reopen, and the photoreceptor cell is returned to its depolarized state. A corresponding increase in intracellular Ca 2+ restores guanylate cyclase to its basal level of activity. Calcium feedback is also thought to facilitate photorecovery through the regulation of rhodopsin phosphorylation by recoverin58 and modulation of the channel sensitivity for cGMP by calmodulin.59 Since the initial patch clamp studies of Fesenko et al.,60 many laboratories have studied the physiological properties of the cGMP-gated channel of rod photoreceptors (for review, see Refs. 46 and 47). Generally, the rod channel is cooperatively activated by cGMP with a K l/2 of 10 to 50 /xM and a Hill coefficient of 1.7 to 35 and permeable to a wide range of monovalent and divalent cations including Na + , K+, Li+, Cs + , Rb"1", Ca 2+ , Mg 2+ , Mn 2+ , and Ba2+. The pharmacological agent, l-cis diltiazem is an effective inhibitor of channel activity and divalent cations, such as Ca 2+ and Mg2+ are known to significantly decrease the conductance of the channel. Molecular Characterization and Subcellular Distribution of the cGMP-gated Channel Molecular characterization of the rod cGMP-gated channel began in the late 1980s when Neil Cook and Benjamin Kaupp isolated a 63-kDa protein from bovine ROS that exhibited cGMP-dependent channel activity when reconstituted into lipid vesicles or planar bilayers.61'62 These studies, however, were not without considerable controversy since rhodopsin,63 a 39-kDa protein64 and a 250-kDa protein65 were also reported to be the rod channel. At the same time, Delyth Reid, a postdoctoral fellow in my laboratory, had generated a monoclonal antibody, designated PMclDl, which reacted with a 63-kDa protein in ROS plasma membranes. In collaboration with Benjamin Kaupp's group in Germany, we were able to show that this antibody recognizes the 63-kDa channel protein in both ROS and purified channel preparations.66 Furthermore, the PMclDl antibody immobilized on Sepharose quantitatively immunoprecipitated the 63-kDa protein and cGMP-dependent channel activity.66'67 These studies provided compelling evidence that the 63-kDa protein, and not the other candidate proteins, is a component of the rod cGMP-gated channel. Using isolated disk and plasma membrane preparations, we further showed by western blot analysis and activity measurements that the 63-kDa channel protein is present in the ROS plasma membrane, but absent in disks (Fig. 5A, 5B).66 Immunohistochemical techniques have confirmed that the cGMP-gated channel and the Na/Ca-K exchanger are targeted to the ROS plasma membrane and are not present in detectable quantities in disk membranes or in other retinal neurons (Fig. 5C) 3O(56-70 Molecular cloning and expression studies of Kaupp et al.7' provided the most direct evidence that the 63-kDa protein is a subunit of the rod channel and, in addition, yielded information about its primary structure. The cloned protein having a molecular mass of —79.6 kDa is considerably larger than the 63-kDa channel protein of ROS. This is attributed to the absence of the N-terminal 92 amino acids in die ROS channel, 2498 Molday IOVS, December 1998, Vol. 39, No. 13 V CV i DV FIGURE 3. Electron micrographs showing the separation of rod outer segment (ROS) disk and plasma membranes by the ricin gold affinity density perturbation method.2** (A) Isolated neuraminidase-treated ROS densely labeled with ricin gold particles (10 nm diameter). (B) Membrane fragments obtained after hypotonic lysis of ROS. Unlabeled disks are seen radiating from the cytoplasmic surface of the gold-labeled plasma membrane. (C) Disk membrane fraction obtained after sucrose density centrifugation. (D) Ricin gold-labeled plasma membrane fraction obtained after sucrose density centrifugation. presumably due to photoreceptor-specific posttranslational proteolysis.6* When expressed in Xenopus oocytes or HEK 293 cells, the protein assembles into a functional channel that is activated by cGMP.7172 These results led to the early view that the rod cGMP-gated channel is an oligomeric complex consisting of identical a-subunits. The first indication that the rod channel contains a second subunit was obtained from the cloning studies of King Wai Yau and coworkers.73 They isolated a cDNA encoding a 102-kDa polypeptide that is 30% identical in sequence to a-subunit and possesses similar structural features. Although this subunit, referred to as subunit 2, does not assemble into a functional channel when expressed by itself, coexpression with the rod a-subunit produces a functional channel that exhibits physio- logical properties characteristic of the ROS channel. These include rapid opening and closing orflickeringbehavior, inhibition by l-cis diltiazem and modulation by Ca-calmodulin.73'74 Questions regarding the true nature of the second subunit arose, however, since a 102-kDa polypeptide is not observed in isolated channel preparations from bovine or human ROS.6774 Instead, a larger polypeptide is observed that migrates on SDS polyacrylamide gels with an apparent molecular mass of 240 kDa (Fig. 6). To begin to characterize this channel-associated protein, we isolated a number of proteolytic peptides derived from the 240-kDa protein. The sequence of several peptides matched sequences present in subunit 2, suggesting this subunit is part of the 240-kDa protein.74 However, some peptide sequences were not found in subunit 2: but instead were The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 2499 Exchanger 4 RGS jr GDPT\ --*-?••—.. Ca 2 *t Transducin y Light ^> N * Channel GD Rhodopsin^ Disk Membrane Plasma Membrane FIGURE 4. Diagram depicting the principal reactions in phototransduction. Light initiates the isomerization of 11-cis retinal to all-trans retinal resulting in the activated (Meta II) state of rhodopsin. Meta II rhodopsin catalyzes the exchange of GDP for GTP on transducin and the dissociation of the a-subunit (Tot) from the j37-subunits (T/3y). To: interacts with phosphodiesterase (PDE) to release the inhibitory constraint on the enzyme. Activated PDE catalyzes the hydrolysis of cGMP to 5'GMP. The decrease in intracellular cGMP concentration causes the channel to close and the rod cell to hyperpolarized. Intracellular Ca2+ levels decrease as the Na/Ca-K exchanger continues to extrude Ca2+ from the outer segment. Photorecovery is initiated by the shutoff of the visual cascade and the calcium mediated feedback mechanism. The visual cascade system is inactivated through (1) the phosphorylation of rhodopsin by rhodopsin kinase (RK) and the subsequent binding of arrestin; (2) hydrolysis of GTP to GDP on a-subunit of transducin, a reaction activated by RGS protein. PDE also returns to its inactive state as a result of this reaction; and (3) reassociation of the a-subunit of transducin with its jBy-subunits to form the inactivated transducin heterotrimer. Low intracellular Ca2+ concentrations lead to (1) the activation of guanylate cyclase, a process that is mediated by the calcium-binding protein GCAP; and (2) an increase in the sensitivity of the channel to cGMP as a result of the dissociation of calmodulin from the channel. As cGMP concentration increases, the channels reopen and the cell is returned to its depolarized state. The increase in Ca2+ also converts guanylate cyclase to its inactive or basal level of activity. The solid arrows show the photoexcitation process; dashed arrows show the photorecovery process. identical with sequences present in a previously cloned retinal glutamic acid-rich protein called GARP.75'76 The relationship between subunit 2 and GARP was resolved when, in collaboration with Benjamin Kaupp's laboratory, we were able to show that the 240-kDa polypeptide represents the full length j3-subunit of the cGMP-gated channel and contains both subunit 2 and GARP.75 Molecular Structure and Regulation of the cGMPGated Channel Topological models for the a- and /3-subunits of the rod channel as developed from sequence analysis and immunochemical labeling studies are shown in Figures 7A and 7B/ 5877 - 80 The core structural unit consists of six membrane-spanning segments (S1-S6) followed by a cGMP binding domain. A voltage sensor-like motif comprising the S4 segment and a pore region of approximately 20 to 30 amino acids located between the S5 and S6 transmembrane segments are also evident,81 A negatively charged glutamate residue in the pore region of the a-subunit has been shown to be responsible for external diva- lent cation blockage.82"3 Since the S4 segment, the pore region, and the folding pattern of the cGMP-gated channel subunits are characteristic features of voltage-gated cation channels, it has been suggested that cyclic nucleotide-gated channels and voltage-gated channels are members of a superfamily of cation channels that have evolved from a common primordial channel.81 The |3-subunit contains an unusual bipartite structure (Fig. yB-j 75,8'f T h e c-terminal region or 0' part of approximately 800 amino acids contains the core structural unit of the channel required for activity. The N terminus contains the GARP part, a region having a high content of glutamic acid and proline residues. Two shorter spliced variants of GARP, called f-GARP and t-GARP, are also present in ROS.84'85 The functions of the various GARP variants are currently under investigation. The channel is a heterotetrameric complex,86 most likely consisting of two a- and two |3-subunits (Fig. 7C). The pore regions of the individual subunits are oriented toward the central cavity of the channel where they serve as the ion selectivity filter and gate.87 Recent high-resolution x-ray analy- 2500 Molday IOVS, December 1998, Vol. 39, No. 13 Coomassie Blue PMC1D1 205976845 29- r a b 63 kDa c a b c B 0,02- cGMP Plasma Membrane AA 0.01- Disk Membrane 50 100 sis of a related K + selective channel from Streptomyces lividans has provided a detailed structure of the pore region of this channel and insight into how such pore regions function in the translocation of ions through these channels. 88 As part of our studies on the rod channel, we investigated factors that regulate the activity of the rod cGMP-gated channel. Yi-Te Hsu, a former graduate student, first showed that calmodulin modulates the apparent affinity of the channel for cGMP in a calcium dependent manner. 5 9 8 9 Using both native and reconstituted channels, he showed that Ca-calmodulin shifts the dose-response curve to higher cGMP concentrations (Fig. 8) : an effect that has been reproduced in various experimental systems. 74 ' 7590 He also showed that calmodulin-Sepharose can be used to purify the channel and that the binding site for calmodulin is localized on the £-subunit. 59 ' 88 More recently, this site has been mapped near the N terminus of the core structural unit of the j3-subunit.9192 Modulation of the channel by calmodulin operates under physiological Ca 2+ concentrations 59 ' 89 This has led us to suggest that Ca-calmodulin regulation of the channel may play a role in facilitating photorecovery after bleaching by increasing the sensitivity of the channel to cGMP. 5979 The finding that calmodulin modulates the rod channel has led to the important discovery by King Wai Yau's group that the olfactory cyclic nucleotide-gated channel is also strongly modulated by calmodulin, a mechanism that is important in olfactory adaptation. kDa 205- - 240 kDa (p-subunit) 97- 6845FIGURE 5- Localization of die cGMP-gated channel to the rod outer segment (ROS) plasma membrane. (A) Sodium dodecyl sulfate polyacrylamide gel stained with Coomassie blue (left) and the corresponding western blot analysis labeled with monoclonal antibody PMclDl against the 63-kDa osubunit of the channel.66 Lane a: rod outer segment (ROS) membranes; lane b: isolated disk membrane fraction; lane c: isolated plasma membrane fraction. (B) cGMP dependent channel activity. Purified disk and plasma membrane fractions solubilized in CHAPS detergent were reconstituted into Ca2+-containing lipid vesicles. The release of Ca 2+ was initiated with 150 fjM cGMP and detected by a change in absorbance (AA) using Arsenazo in as an indicator. Only the plasma membrane exhibits channel activity.66 (C) Electron micrograph of bovine ROS membranes labeled with antibodies against the cGMP-gated channel. ROS membrane fragments were labeled with the PMc6E7 anti-channel antibody followed by immunogold particles (10 nm diameter). The cytoplasmic surface of the inside out plasma membrane vesicles (arrowheads) is densely labeled. The disk membranes radiating from the plasma membrane vesicles are not labeled.68 63 kDa (ct-subunit) 29- FIGURE 6. Immunoaffinity purification of the channel from rod outer segment (ROS). ROS membranes (lane a) were solubilized in CHAPS buffer and passed through a PMc6E7Sepharose affinity column. After the column was washed to remove unbound proteins, the channel (lane b) was eluted with a peptide corresponding to the epitope for the PMc6E7 monoclonal antibody. The samples were run on a 9% sodium dodecyl sulfate polyacrylamide gel and stained with Coomassie blue. The purified channel contains two prominent polypeptides: the 63-kDa a-subunit and the 240-kDa 0-subunit. The Friedenwald Lecture 2501 IOVS, December 1998, Vol. 39, No. 13 Pore Region Pore Region S1 S2 S3 S4S5I S6 \ \ I / / | I RinHinn ^ite Binding Site 6 g o Garp Part CHO (V-Part f-Garp t-Garp Pore Region Pore Region COOH p-Part a - Subunit Garp Part - Subunit a 7. Schematic models of the rod cGMP-gated channel. (A) Linear representation (top) and topological model (bottom) of the rod a-subunit. The membrane spanning segments (S1-S6), the pore region containing a glutamate residue (0), the N-linked glycosylation site (CHO), and the cGMP binding site are shown. The N-terminal 92 amino acids (dashed line) is missing in the channel present in rod outer segment (ROS). (B) Linear representation (top) and topological model (bottom) of the rod j3-subunit. In addition to many structural features found in the a-subunit, the j3-subunit contains a calmodulin (CaM) binding site and an extended N-terminal region called the GARP part. Two additional spliced variants of GARP, called f-GARP and t-GARP, are also present in ROS.84 Unlike the a-subunit, the /3-subunit does not contain an N-ltnked glycosylation site or a glutamate residue in the pore region. (C) Model for the cGMP-gated channel complex. Two cc-subunits and two /3-subunits assemble into a tetrameric complex. The S6 and S5 segments are visualized to line the central cavity of the channel. The pore regions joining the S5 and S6 segments near the extracellular surface of the membrane extend toward the center of the cavity where they can function as a gate and ion selectivity filter. The glutamate residue (0) in the pore region of the a-subunit is responsible for external divalent cation blockage of the channel. Although the channel is shown with identical subunits across from each other, it is possible that the two identical subunits may be adjacent. FIGURE IOVS, December 1998, Vol. 39, No. 13 amounts of ATP are produced by anaerobic glycolysis to sustain cGMP levels in dark-adapted ROS.102 Glucose is also used by the hexose monophosphate pathway in ROS to regenerate NADPH for retinal reduction after the photobleaching of rhodopsin and glutathione reduction for the protection of outer segments from oxidative stress.102 A phosphocreatine shuttle system most likely serves as an additional source of energy. Biochemical pathways involved in the production of energy and reducing equivalents in ROS are depicted in Figure 10. The importance of anaerobic glycolysis in retinal function has been addressed in considerable detail in the physiological and biochemical studies of Barry Winkler and coworkers.104'105 RETINAL DEGENERATIVE DISEASES 60 cGMP Concentration FIGURE 8. Modulation of the rod cGMP-gated channel by Cacalmodulin. Rod outer segment (ROS) membrane vesicles loaded with Arsenazo III were placed in a calcium-containing buffer in the absence (•) or presence (•) of calmodulin, and calcium uptake was initiated by the addition of cGMP. The relative initial velocity (VJVm:ix) is plotted against cGMP concentration. The solid line is the sigmoidal binding curve calculated using a Michaelis constant (Km~) of 19 /xM and a Hill coefficient of 3-7 in the absence of calmodulin and a Km of 33 /xM and a Hill coefficient of 3-5 in the presence of calmodulin.59 In 1990, Ted Dryja and coworkers first reported that a P23H mutation in rhodopsin is responsible for a form of autosomal dominant retinitis pigmentosa (ADRP).106 Since this time, more than 70 different rhodopsin mutations have been linked to autosomal dominant and recessive forms of retinitis pigmentosa, and congenital stationary nightblindness (CSNB).107'109 GLYCOLYTIC ENZYMES AND GLUCOSE METABOLISM IN ROS AND CONE OUTER SEGMENTS Phototransduction and related outer segment processes require considerable amounts of energy and reducing equivalents in the form of ATP, GTP, and NADPH. Over the years, questions arose as to whether these cofactors are generated in the outer segments or produced in the inner segment and shuttled to the outer segment. In early studies, glycolytic and hexose monophosphate enzyme activities were detected in outer segment preparations,95"98 but the activities were low in comparison to other retinal cell layers. As a result, it has been unclear whether these enzyme activities come from endogenous outer segment enzymes or alternatively, from contaminants of the preparations. A report indicating that outer segments contain considerable amounts of creatine kinase also led to the possibility that energy in the form of ATP can be transferred from the inner segment to the outer segment by a phosphocreatine shuttle pathway.99 Our interest in glucose metabolism arose when Shu-Chan Hsu, a former graduate student, identified an abundant 38-kDa ROS plasma membrane-associated protein as glyceraldehyde3-phosphate dehydrogenase, a key enzyme in anaerobic glycolysis. 10° She further showed by immunofluorescence, western blot analysis and enzyme activity measurements that other glycolytic enzymes and a GLUT-1 glucose transporter are also present in ROS and cone outer segments.101 Moreover, quantitative measurements indicated that isolated ROS preparations have the capacity to take up glucose and convert glucose to lactate by anaerobic glycolysis (Figs. 9A, 9B).101"103 Sufficient 0 50 100 150 Time (seconds) 200 B to O ac 400 CD 200 - 5 10 Time (min) FIGURE 9- 15 A Uptake of glucose into isolated bovine rod outer segment (ROS). The uptake of 45 /xM external 3-O-[l4C]methylglucose (3-O-MG), a nonmetabolizable analogue of glucose, was measured in the absence (•) and presence (O) of 0.05 mM cytochalasin B, an inhibitor of glucose transport.101 (B) Glycolytic flux in isolated bovine ROS. Time course of lactate production by isolated bovine ROS in the presence (•) and absence (V) of 5 mM glucose.102 The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 GP _ GSSG Glutathione redox cycle GSH OS PHOTOTRANSDUCTION IS FIGURE 10. Diagram showing the role of glycolysis, the hexose monophosphate pathway, and the phosphocreatine shuttle in ATP, GTP, and NADPH production. In photoreceptor outer segments (OS), glucose is transported across the plasma membrane by a GLUT-1-type glucose transporter (GT). Glucose is metabolized to lactate by anaerobic glycolysis resulting in the production of ATP and GTP for phototransdviction and other energy requiring processes. Glucose can also be used by the hexose monophosphate pathway (HMP) for the regeneration of NADPH. The latter is required for the reduction of all-trans retinal to all-trans retinol by retinol dehydrogenase (RDH) after the bleaching of rhodopsin and for glutathione reduction by glutathione reductase (GR) for protection of the outer segment against oxidative damage. Additional energy is obtained from the inner segment (IS). High-energy phosphocreatine (PCr) is transferred to the outer segment by the PCr and used to generate ATP. This reaction is catalyzed by a brain-type creatine kinase (Ckb).102 In addition, positional cloning and candidate gene approaches have led to the identification of large number of genes associated with retinitis pigmentosa and other retinal degenerative diseases (Table 3).108-109 Many genes code for photoreceptorspecific proteins that play key roles in phototransduction and outer segment morphogenesis. Our interest in this area has focused on the peripherin/rds-rom-1 complex and the ABCR/ RIM protein, two disk membrane proteins implicated in multiple retinal degenerative diseases. PERIPHERIN/RDS—ROM-1 COMPLEX Molecular Structure and Subcellular Distribution Peripherin/rds and rom-1 are homologous subunits of an oligomeric membrane protein found in disks. Peripherin/TYfe, 2503 formerly called peripherin, was first detected as a 33- to 35-kDa protein in bovine disk membranes with monoclonal antibodies110 and subsequently cloned from a retinal expression library by Greg Connell, a former graduate student in my laboratory. 1 " Several years later, Rod Mclnnes's laboratory in Toronto cloned rom-1, a protein that is 30% identical in sequence to peripherin/rds."2 Immunochemical labeling studies and sequence analysis has led to a topological model for these proteins (Fig. II). 1 1 1 "" 4 Characteristic structural features include four membrane- spanning segments (M1-M4), an extended carboxyl terminal domain exposed on the cytoplasmic side of disks and a large intradiskal loop (L3-L4) containing seven conserved cysteine residues. Peripherin/rds and rom-1 associate to form a tetrameric complex as determined by immunoprecipitation studies and hydrodynamic measurements."2"1'6 Since dimers of peripherin/rds and rom-1 are routinely observed by SDS gel electrophoresis under nonreducing conditions," 0 "" 3 it was initially thought that a disulfide-linked homodimer of peripherin/rcls interact noncovalently with a disulfide-linked homodimer of rom-1 to form a heterotetramer. " 4 " " 6 More recently, however, Chris Loewen, a graduate student in my laboratory, has shown that the peripherin/rd.s-rom-1 tetramer does not contain intermolecular disulfide bonds.117 Instead, intermolecular disulfide bonds, mediated by Cys 150 within the large intradiskal loop,118 link individvial tetramers into higher order oligomers, a mechanism that may underlie disk morphogenesis.117 The peripherin/rds-rom-1 complex has a unique subcellular distribution. Preembedding and postembedding immunogold labeling studies have localized peripherin/rds to the rim region of ROS and cone outer segment disks (Figs. 12A, 1 2 B ) iio,iu,u<f, 119 i mt i a i studies suggested that rom-1 is only COOH Cytoplasmic side Intradiscal side L3-4 FIGURE 11. Topological model for peripherin/rcfe and rom-1 based on sequence analysis and immunochemical studies. Both proteins are shown to contain four transmembrane segments (M1-M4), cytoplasmic N and C termini, and a large intradiskal loop (L3-4). Perpherin/rds contains an N-linked oligosaccharide chain (hexagons') that is not present in rom-1. Stretches of conserved amino acids and conserved cysteine residues are indicated.1' 1~113 2504 TABLE Molday IOVS, December 1998, Vol. 39, No. 13 3- Proteins Associated with Various Retinal Degenerative Diseases Protein Localization Disease Rhodopsin Phosphodiesterase O,/3) cGMP-gated channel (a) Guanylate cyclase (Ret GC1) GCAP1 Arresttn Rliodopsin kinase Transducin (a) Peripherin/r^/s Rod cells Rod cells Rod cells Cone and rod cells Cone and rod cells Rod cells Rod cells Rod cells Rod and cone cells ADRp'CKS.107 , m Rom-1 ABCR/RIM RPGR CRALBP CRX (transcriptional factor) RPE65 TIMP3 XLRS1 Myosin VIIA Bestrophin Rod and cone cells Rod cells Rod cells RPE and Miiller cells Rod and cone cells RPE cells Bruch's membrane Extracellular (photoreceptors) Photoreceptors/RPE RPE Digenic ADRP 137 ' 138 Stargardt's MD, 149 ' 152 ARRP,15'' CRD,' 55 and AMD?153 X-linked RP (RP3) 172 ' 182 ARRP15y LCA and CRD 1 5 7 1 5 8 LCA and ARRP160'161 Sorby's macular dystrophy 162 X-linked retinoschisis 170 Usher syndrome (USH1)171 Best's MD(VMDd2) 163164 ARRP 168169 and CSNB177 ARRP166 LCA167 and CRD1*0 Cone dystrophy 165 CSNB (Oguchi disease) 173 CSNB (Oguchi disease) 174 CSNB (Nougaret disease) 175 ADRP, 126 - 128 ' 130 ' 131 MD, 1 3 1 1 3 4 1 3 6 pattern dystrophy, 132133 and digenic ADRP 137 ' 138 ADR]*, autosomal dominant retinitis pigmentosa; ARRP, autosomal recessive retinitis pigmentosa; CSNB, congenital stationary nightblindness; X-linked RP, X-linked retinitis pigmentosa; MD, macular dystrophy; AMD, age-related macular dystrophy; CRD, cone-rod dystrophy; LCA, Leber's congenital amaurosis. present in ROS disks. 112 However, Orson Moritz, a former graduate student in my laboratory, has clearly shown that rom-1 is present along the rim region of cone and rod disk membranes as shown in Figure 12C. 113 B Role of Peripherin/rrfs in Outer Segment Morphogenesis and Retinal Degeneration in the RDS Mouse The importance of peripherin/r^ in outer segment morphogenesis originated from the cellular and molecular analysis of the retinal degeneration slow or rds mouse. Sanyal and coworkers 120 ' 121 showed that mice homozygous for the rds mutation fail to develop outer segments and the photoreceptor cells undergo slow degeneration such that few cells remain 1 year after birth. Heterozygous rds mice exhibit short, highly disorganized outer segments that often appear as whorls of membrane. 122 In the late 1980s, Grabriel Travis and colleagues 123 used subtractive hybridization techniques to identify a mutation in a photoreceptor cell-specific gene that is responsible for the rds phenotype. We subsequently showed that this gene codes for peripherin/refe.124 Finally, the research groups of Gabriel Travis and Dean Bok have reported that introduction of the normal peripherin/rds gene into a homozy- ROS COS CIS FIGURE 12. Localization of peripherin/rrfs and rom-1 to the rim region of rod outer segment (ROS) and cone outer segment (COS) disks. (A, B) Electron micrographs of an isolated bovine ROS disk and a group of disks labeled with Per 3B6 antiperiphcrin/rds monoclonal antibody and immunogold particles (10 nra diameter). The gold particles are localized to the rim region of the disks. (C) Electron micrograph of an ROS, a COS, and a cone inner segment (CIS) labeled for rom-1. Bovine retina was embedded in LR White resin. Sections were then labeled with the anti-rom-1 antibody (Rom L4) and immunogold particles. Gold particles are distributed along the peripheral region of both the ROS and COS and the incisures of ROS (arrow- head)^ The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 330. 2505 346 Cytoplasmic side OOH Trp25FS Intradiskal side Lysl 53Arg Lysl 53Del Pro2l6Leu Pro2l6Ser *Cys2l4Ser Ser212Gly Pro210Arg Pro210Ser Lysl93Del 13- Location of peripherin/rds mutations associated with retinal degenerative diseases. Unframed mutations have been linked to monogenic autosomal dominant retinitis pigmentosa (ADRP). Mutations framed in black have been implicated macular and pattern dystrophies. The mutation framed in gray is linked to digenic ADRP. Insert/RDS is the site of an insertion mutation in the rds mouse; FS indicates a frameshift resulting in a change in reading frame and premature stop. FIGURE gous rds mouse results in outer segment formation and suppression of photoreceptor degeneration.125 Taken together, these studies indicate that peripherin/rds is plays an essential role in ROS and cone outer segment morphogenesis, and the absence or reduced amounts of this membrane protein result in photoreceptor degeneration. Role of Peripherin/rds in ADRP and Macular Degeneration The finding that a mutation in the peripherin/rds gene causes photoreceptor degeneration in the rds mouse prompted many laboratories to determine whether mutations in the human gene are responsible for inherited retinal diseases. This has proved to be the case and to date more than 30 mutations in the peripherin/rds gene have been implicated in various retinal degenerative diseases (Fig. 13).126"130 Interestingly, whereas some peripherin/rds mutations cause ADRP, others result in macular degeneration and pattern dystrophies that go under such names as, butterfly-shaped pigment dystrophy, fundus flavimaculatus, cone-rod dystrophy and Bull's Eye maculopathy. l31~136 Similar efforts to link mutations in rom-1 with retinal degenerative diseases have been less successful. However, several mutations in the rom-1 gene have been linked to a novel form of digenic ADRP.137"138 To begin to understand the mechanism by which mutations in peripherin/rds cause specific disease phenotypes, Andy Goldberg, a former postdoctoral fellow in my laboratory, developed a heterologous cell expression system to study the molecular properties and subunit interactions of peripherin/rds and rom-1. 115 He showed that coexpression of peripherin/rds and rom-1 results in a heterotetrameric complex having properties similar to those of the complex derived from ROS. However, many mutations in the large intradiskal segment, including the C214S peripherin/rds mutant-linked ADRP,130 result in abnormal protein folding and the inability of peripherin/rds to associate with rom-1." 8 From these studies, we have concluded that segments of the large intradiskal loop are important in subunit-subunit interactions, a property important in normal outer segment formation. We have also examined the R172W peripherin/rds mutant linked to a mild form of macular degeneration.131136 Interestingly, this mutant folds normally and assembles with rom-1 into a nativelike tetrameric complex.139 Apparently, subtle differ- 2506 Molday ences in the structure and/or stability of this complex result in preferential cone degeneration.136 Recently, a novel digenic form of ADRP linked to a L185P mutation in peripherin/nfc and a null mutation in rom-1 has been described in several families. 137138 Individuals who coinherit both mutations (double heterozygotes) exhibit an ADRP disease phenotype, whereas family members who inherit only one of these mutations (single heterozygotes) are essentially normal. We postulated that this disease pattern might arise from defective peripherin/rdsrom-1 subunit interactions. To investigate the role of subunit assembly in this form of digenic RP, Andy Goldberg analyzed the hydrodynamic properties of the complexes formed when either wild-type peripherin/rds or the L185P mutant is expressed in COS-1 cells in the presence and absence of rom-1. l39 The results of this study are diagramed in Figure 14. Coexpression of the L185P peripherin/rds mutant with rom-1 (simulating an individual who inherits only the peripherin/n/s mutation) results in a heterotetrameric complex similar to the wild-type complex. Wild-type peripherin/ rds in the absence of rom-1 (simulating an individual who inherits a rom-1 null mutation) self-assembles into a homotetrameric complex. In contrast, L185P peripherm/rds in the absence of rom-1 (simulating individuals who inherit both mutations) fails to self-assemble into a tetramer, but instead remains in a dissociated state. From these studies we have concluded that peripherin/rds-containing tetramers are crucial in outer segment formation and stability. Reduced levels of this complex can lead to abnormal and unstable outer segments and progressive photoreceptor degeneration.139 Implicit in this model is the hypothesis that peripherin/rc/s homotetramers can effectively substitute for peripherin/rdsrom-1 heterotetramers and promote outer segment disk morphogenesis. Recently, we have had the opportunity to test this prediction in a Rom-1 knockout mouse produced by Geoff Clark and Rod Mclnnes.l4° Biochemical and ultrastructural analyses of these mice, indeed, indicate that peripherin/rds self-assembles into a homotetramer in the absence of rom-1, and this complex supports ROS and cone outer segment formation (Goldberg A, Molday L, Molday R, Clark G, Mclnnes R, unpublished results, 1997). Comparative analysis of the rom-1 knockout mouse and the rds mouse leads to the conclusion that peripherin//Y/s is the dominant subunit required for disk morphogenesis. Rom-1 is relegated to a more minor role, perhaps enhancing the stability of the outer segment and/or fine tuning the structure of disks. THE ABCR/RIM PROTEIN AND STARGARDT'S MACULAR DEGENERATION More recent studies in my laboratory have focused on the identification and characterization of the abundant high-molecular-weight rim protein first identified by David Papermaster and colleagues in frog photoreceptors"""' 2 and subsequently detected in mammalian ROS.1''314'1 Michelle Illing, as part of her graduate research in my laboratory, purified and cloned the bovine 220-kDa rim protein and showed that this protein is a member of the superfamily of ABC (ATP binding cassette) transporters.145 This extensive family of proteins include the cysticfibrosistransmembrane regulator (CFTR) linked to cystic IOVS, December 1998, Vol. 39, No. 13 Molecular Complex B D Phenotype WT Per/rds WT Rom-1 Normal Ll 85P Per/rds WT Rom-1 "Normal" Wt Per/rds Null Rom-1 "Normal" L185PPer Null Rom-1 Digenic ADRP 14. Subunit assembly model for digenic autosomal dominant retinitis pigmentosa (ADRP) linked to a L185P mutation in peripherin/rds and a null allele in Rom-1. Left: molecular complexes of peripherin/rds (dark) and rom-1 (light) determined by velocity sedimentation measurements.l39 Right: corresponding phenotypes for individuals who inherit one or both mutations. (A) Wild-type (WT) peripherinAv5fo-rom-l complex characteristic of normal individuals. (B) L185P peripherin/r^5-rom-l complex predicted to exist in individuals who inherit only the L185P mutation. This tetrameric complexes is suggested to support outer segment formation resulting in a borderline "Normal" phenotype. (C) Peripherin/rrfs homotetrameric complex predicted to exist in individuals who inherit a null mutation in rom-1. This complex can take the place of the peripherin/rc/s-rom-1 heterotetramer and support outer segment morphogenesis and structure. A borderline "Normal" phenotype can result from a small net reduction in the amount of peripherin/rcfc containing tetramers. (D) Dissociated L185P peripherin/r<3?s mutant in the absence of rom-1. This species is not expected to support outer segment morphogenesis or structure. A significant reduction in the level of peripherin/rc/.s containing tetramer will result in disorganized, unstable outer segments, a condition which underlies photoreceptor degeneration and a ADRP phenotype. FIGURE fibrosis, P-glycoprotein involved in multidrug resistance in cancer, TAP1 and TAP2 proteins that serve as peptide transporters in lymphocytes, prokaryotic permeases, and others.146 Like CFTR and P-glycoprotein, the rod ABC protein consists of two structurally related halves, each of which contains a multiple membrane-spanning domain followed by a cytoplasmic ATP binding cassette. On the basis of sequence analysis and biochemical studies, we have developed a working model for the topological organization of this glycoprotein in disk membranes as shown in Figure 15. The Friedenwald Lecture 10VS, December 1998, Vol. 39, No. 13 2507 N1614FS G818E Del WAIC INTRADISKAL SPACE R2106C CYTOPLASMIC SPACE L2027F I R2038W \R2077W V2050L E1087K L962FS V1072A N965S A1028V S1071FS E1036K FIGURE 15- Topological model for ABCR/RIM based on sequence analysis, location of N-linked oligosaccharides, and analogy with other ABC transporters. The model shows 12 transmembrane segments, 2 ATP binding cassettes (ABC) on the cytoplasmic side of the membrane, and 2 N-linked oligosaccharide chains (hexagons') on the intradiskal side. 145 Several mutations linked to Stargardt's macular dystrophy are indicated. 149152 The cellular and subcellular distribution of this ABC protein has been investigated using both immunochemical and molecular biology techniques. The protein is abundantly expressed in the outer segments of rod photoreceptors, but is absent in cone cells. 1 4 5 1 4 7 ' 1 '' 8 This is in agreement with in situ hybridization and northern blot analysis, indicating that mRNA expression is restricted to rod cells. 149 At a higher resolution, postembedding, immunogold labeling studies carried out by Laurie Molday have shown that the bovine rod ABC transporter, like its frog counterpart, is localized to the rim region of rod disk membranes (Fig. l 6 ) . 1 4 1 1 4 5 During the course of our studies, Allkimets and coworkers elegantly showed that mutations in the ABCR gene coding for a retinal rod-specific ABC protein are responsible for Stargardt's disease, an autosomal recessive macular dystrophy with a juvenile onset. 149 This disease, characterized by progressive loss in central vision, bilateral atrophy of the macular region of the retina and RPE layer, and the appearance of orange-yellow flecks, accounts for up to 7% of the human retinal degenerative diseases. 150151 Analysis of the amino acid sequences of human, bovine, and mouse proteins indicate that the ABCR protein and the rim protein are one in the same. 1 4 5 ' 1 4 7 ' 1 4 8 ' 1 5 2 The location of several reported Stargardt's mutations l 4 y 1 5 2 in context to our working model of rod ABC protein, now referred to as the ABCR/RIM protein, is shown in Figure 15. More recent studies suggest that mutations in the ABCR gene can also cause other disease phenotypes. Allkimets et al. 153 have reported that some mutations in the ABCR gene are associated age-related macular degeneration, a result that has generated considerable controversy. Homozygous and compound heterozygous mutations in the ABCR gene resulting in null alleles have also been implicated in autosomal recessive RP and cone-rod dystrophy. 154 ' 155 The function of the ABCR/RIM protein is not currently known. Other members of this superfamily are known to mediate the active transport of a wide range of compounds including drugs, metabolites, peptides, and lipids. l46 A related protein, ABC1, has also been implicated in the engulfment of cell corpses after apoptosis. l5<5 It is possible that the ABCR/RIM ROS ROS RIS 8 FIGURE 16. (A, B) Localization of ABCR/RIM protein to the rim and incisures of rod outer segment (ROS) disks. Electron microgniphs of bovine rod photoreceptor cells embedded in LR White resin, sectioned, and labeled with an anti-ABCR/RIM antibody and immunogold particles. Labeling is restricted to the peripheral region and incisures (arrowhead) of the ROS, a pattern that is characteristic of disk rim labeling. The rod inner segment (RIS) is not labeled. 145 2508 Molday IOVS, December 1998, Vol. 39, No. 13 Retina) Derivative Peptide Antioxidant Phospholipid Other A DP + Pi ABCR/RIM Peripherin/rds-Rom-1 Complex FIGURE 17. Diagram showing possible transport functions of the ABCR/RIM protein. Retinal derivatives, peptides, antioxidants, phospholipids, or other agents may be actively transported into or out of the disk membrane or translocated from one side of the disk lipid bilayer to the other using ATP hydrolysis as a source of energy. Pi, inorganic phosphate. Glutathtone Cycle Hexose Moitophosphate Pathway Retinal Reduclioi Glucose Transporter Glucose Rhodopsm Kmase CSMB Arnwtin CSNB cGMP-gated Channel Na/Ca-K Exchanger 4Na dopsin Guanylate Cyclase LCAI • " Disk Lamellar ADRP MD ABCR/RIM XLRS1 Slgdl MD O ' Disk Rim Plasma Membrane FIGURE 18. Diagram showing the distribution of various proteins in the rod outer segment (ROS). Retinal diseases linked to mutations in these proteins are also indicated. ADRP, autosomal dominant retinitis pigmentosa; ARRP, autosomal recessive retinitis pigmentosa; CSNB, congenital stationary night blindness; CD, cone dystrophy; CRD, cone-rod dystrophy; MD, macular dystrophy; Stgdt MD, Stargardt's macular dystrophy; XLRS1, X-linked retinoschists 1; XLRP3, X-linked retinitis pigmentosa 3. The Friedenwald Lecture JOVS, December 1998, Vol. 39, No. 13 is involved in the transport of retinal derivatives, phospholipids, peptides or other endogenous substrates across the disk membrane CFig- 17)- Molecular and biochemical approaches are now being used to define in more detail the structure and function of this unique disk protein and to understand how specific mutations cause various disease phenotypes. SUMMARY AND FUTURE DIRECTIONS Over the past two decades, remarkable progress has been made on the cellular and molecular characterization of ROS and cone outer segments. Many membrane and soluble proteins have been identified and localized within the outer segment, and their structural and functional properties have been studied in considerable detail. This has led to a comprehensive understanding of phototransduction and supporting metabolic pathways and a glimpse into mechanisms underlying outer segment structure, morphogenesis, and renewal. Recent studies have also indicated that mutations in many photoreceptor outer segment-specific proteins cause a multitude of inherited retinal degenerative diseases (Fig. 18). Molecular studies are now beginning to provide insight into how specific mutations in these proteins lead to various retinal diseases. Although we have learned a great deal about the molecular composition and function of ROS and cone outer segments, much remains to be learned. The staicture and function of many proteins associated with retinal degenerative diseases, such as the ABCR/RJM associated with Stargardt's disease, XLRS1 implicated in X-linked juvenile retinoschisis, and RPGR associated with X-linked RP3 remain to be determined. Regulatory mechanisms underlying phototransduction need to be understood in a more detailed, quantitative manner. Molecular mechanisms and protein-protein interactions that form and stabilize the unique ROS and cone outer segment structure and target proteins to the outer segment and more specifically to the outer segment disk or plasma membrane have to be understood at a molecular level. Although the future challenges are numerous, one only can be optimistic that the rapid advances in basic and clinical research will lead to a complete understanding of photoreceptor structure and function and detailed insight into the molecular basis for retinal diseases. From this knowledge, it should be possible to develop novel and effective treatments for most, if not all, the diseases that lead to the loss in vision. Acknowledgments I want to convey my sincere gratitude to the numerous past and present research colleagues who have contributed greatly to the research accomplishments embodied in this award and extend special thanks to Laurie Molday who carried out many of the electron microscopic and biochemical studies described in this article. Other postdoctoral fellows, graduate students, and research assistants who have made significant contributions include Jinhi Ahn, Steve Clark, Carole Colville, Greg Connell, Andrea Dose, Andrew Goldberg, David Hicks, Theresa Hii, Shu-Chan Hsu, Yi-Te Hsu, Michelle Tiling, Tom Kim, Dale Laird, Chris Loewen, Don MacKenzie, Orson Moritz, Igor Nassonkin, Delyth Reid, and Simon Wong. I also thank my friends and collaborators, King Wai Yau, Benjamin Kaupp, Barry Winkler and Bernhard Weber, and other prominent vision investigators with whom I have had an opportunity to collaborate and discuss research of mutual interest. These include Gobind Khorana, David Papermaster, Dan Oprian, Paul Hargrave, Ted Dryja, David Williams, Rod Mclnnes, Neal 2509 Cook, Heinz Wassle, Kris Palczewski, Karl Koch, Paul Bauer, Steve Fisher, and Jeremy Nathans. Finally, I gratefully acknowledge the longstanding grant support from the National Eye Institute, IIP Foundation of Canada, and the Medical Research Council. References 1. Young RW. The renewal of photoreceptor cell outer segments. J Cell Biol. 1967;33:6l-72. 2. Young RW, Bok D. Participation of the retinal pigment epithelium in the rod outer segment renewal process. / Cell Biol. 1969:42:392-403. 3. Bok D. Retinal photoreceptor-pigment epithelium interactions. Friedenwald lecture. Invest Ophthalmol Vis Sci. 1985;26:l659. 4. Dowling JE. The organization of vertebrate visual receptors. In: Allen JM, ed. Molecular Organization and Biological Function. New York: Harper & Row; 1967:186-210. 5. Cohen AI. Rods and cones. In: Fuortes MGF, ed. Handbook of Sensory Physiology. New York: Springer-Verlag; 1972;VII/2:63110. 6. Corless JM, Fetter RD, Zampighi OB, Costello MH, Wall-Buford DL. Structural features of the terminal loop region of frog retinal rod outer segment disk membranes, II: organization of the terminal loop complex. / Comp Neurol. 1987;257:9-23. 7. Eckmiller MS. Cone outer segment morphogenesis. Taper change and distal invaginations. / Cell Biol. 1987;105:2267-2277. 8. Steinberg RH, Fisher SK, Anderson DH. Disc morphogenesis in vertebrate photoreceptors. / Comp Neurol. 1980;190:501-518. 9. Cohen AI. Further studies on the question of the patency of saccules in outer segments of vertebrate photoreceptors. Vision Res. 197O;lO:445-453. 10. Laties AM, Liebman PA. Cones of living amphibian eye: selective staining. Science. 1970;l68:l475-l477. a 11. Roof DJ, Heuser JE. Surface of rod photoreceptor disk membrane components. / Cell Biol. 1982;95:487-500. 12. Usukura J, Yamada E. Molecular organization of the rod outer segment. A deep etching study with rapid freezing using unfixed frog retina. Biomed Res. 1981;2:77-193. 13. Fetter RD, Corless JD. Morphological components associated with frog cone outer segment disk margins. Invest Ophthalmol Vis Sci. 1987;28:646-657. 14. Papermaster DS, Dreyer WJ. Rhodopsin content in the outer segment membranes of bovine and frog retinal rods. Biochemistry. 1974;13:2438-2444. 15. McConnell DG. The isolation of retinal outer segment fragments. JCellBiol. 1965;27:459-473. 16. DeGrip WJ, Daemen FJ, Bonting SJ. Biochemical aspects of the visual process, XVIII: enrichment of rhodopsin in rod outer segment membrane preparations. Vision Res. 1972; 12:1697-1707. 17. Kiihn H. Light and GTP-regulated interaction of GTPase and other proteins with bovine photoreceptor membranes. Nature. 1980; 283:587-589. 18. Kiihn H. Interactions of rod cell proteins with the disk membrane: influence of light, ionic strength, and nucleotides. Curr Top Membr Transp. 1981 ;15:171-201. 19- Molday RS, Molday LL. Isolation and characterization of rod outer segment disk and plasma membranes. Methods Neurosci. 1993; 15:131-150. 20. Nir I, Papermaster DS. Differential distribution of opsin in the plasma membrane of frog photoreceptors: an immunocytochemical study. Invest Ophthalmol Vis Sci. 1983;24:868-878. 21. Hicks D, Molday RS. Differential immunogold-dextran labeling of bovine and frog rod and cone cells using monoclonal antibodies against bovine rhodopsin. Exp Eye Res. 1986;42:55-71. 22. Polaws AS, Altman, LG, Papermaster DS. Immunocytochemical binding of anti-opsin N-terminal specific antibodies to the extracellular surface of rod outer segment plasma membranes. / Histochem Cytochem. 1986;34:659-664. 23. Koch KW, Kaupp UB. Cyclic GMP directly regulates a cation conductance in membranes of bovine rods by a cooperative mechanism. J Biol Chem. 1985;260:6788-6800. 2510 Molday 24. Puckett KL, Goldin SM. Guanosine 3',5'-cyclic monophosphate stimulates release of actively accumulated calcium in purified disks from rod outer segments of bovine retina. Biochemistry. 1986;25:1739-1746. 25. Schnetkamp PPM. Sodium-calcium exchange in the outer segments of bovine rod photoreceptors./ Physiol. 1986;373:25-45. 26. Clark VM, Hall MO. Labeling of bovine rod outer segment surface proteins with I25I. Exp Eye Res. 1982;34:847-859. 27. Molday LL, Molday RS. Glycoproteins specific for the retinal rod outer segment plasma membrane. Biochim Biophys Acta. 1987; 897:335-340. 28. Molday RS, Molday LL. Differences in the protein composition of bovine retinal rod outer segment disk and plasma membranes isolated by a ricin-gold-dextran density perturbation method. J Cell Biol. 1987;105:2589-2601. 29. Cook NJ, Molday LL, Reid D, Kaupp UB, Molday RS. The cGMPgated channel of bovine rod photoreceptors is localized exclusively in the plasma membrane. / Biol Chem. 1989;264:69966999. 30. Reid DM, Friedel U, Molday RS, Cook NJ. Identification of the sodium-calcium exchanger as the major ricin-binding glycoprotein of bovine rod outer segments and its localization to the plasma membrane. Biochemistry. 1990;29:l601-l607. 31. Kamps KM, De Grip WJ, Daemen FJ. Use of a density modification technique for isolation of the plasma membrane of rod outer segments. Biochim Biophys Acta. 1982;687:296-302. 32. Boesze-Battaglia K, Albert AD. Phospholipid distribution among bovine rod outer segment plasma membrane and disk membranes. Exp Eye Res. 1992;54:821-82333. Molday RS. Monoclonal antibodies to rhodopsin and other proteins of rod outer segments. Prog Retinal Res. 1988:173-209. 34. MacKenzie D, Molday RS. Organization of rhodopsin and a high molecular weight glycoprotein in rod photoreceptor disc membranes using monoclonal antibodies. / Biol Chem. 1982;257: 7100-7105. 35. Molday RS, MacKenzie D. Monoclonal antibodies to rhodopsin: characterization, crossreactivity and application as structural probes. Biochemistry. 1983;22:653-660. 36. Molday RS, MacKenzie D. Inhibition of monoclonal antibody binding and proteolysis by light-induced phosphorylation of rhodopsin. Biochemistry. 1985;24:776-781. 37. MacKenzie D, Arendt A, Hargrave P, McDowell JH, Molday RS. Localization of binding sites for carboxyl terminal specific antirhodopsin monoclonal antibodies using synthetic peptides. Biochemistry. 1984;23:6544-6549. 38. Hodges RS, Heaton RJ, Parker JMR, Molday L, Molday RS. Antigenantibody interaction. Synthetic peptides define linear antigenic determinants recognized by monoclonal antibodies directed to the cytoplasmic carboxyl terminus of rhodopsin. / Biol Chem. 1988;263:11768-11775. 39. Laird DW, Wong SC, Molday RS. Antigenic determinants on rhodopsin as defined by monoclonal antibodies. In: Goheen SC, ed. Membrane Proteins. Richmond: BioRad Lab; 1987:45-70. 40. Hicks D, Barnstable CJ. Different rhodopsin monoclonal antibodies reveal different binding patterns on developing and adult rat retina. / Histochem Cytochem. 1987;35:1317-1328. 41. Doi T, Molday RS, Khorana HG. The role of the intradiscal domain in rhodopsin assembly and function. Proc Natl Acad Sci USA. 1990;87:4991-4995. 42. Oprian DD, Molday RS, Kaufman R, Khorana HG. Expression of a synthetic bovine rhodopsin gene in monkey kidney cells. Proc Natl Acad Sci USA. 1987;84:8874-8878. 43. Hsu Y-T, Connell GJ, Wong S, Molday RS. Structural and functional properties of rhodopsin from rod outer segment disk and plasma membrane. Biochim Biophys Acta-Biomembranes. 1993; 1145:85-92. 44. Olsson JE, Gordon JW, Pawlyk BS, et al. Retinal degeneration in transgenic mice with a mutant or wild-type copy of the human rhodopsin gene. Neuron. 1992;9:815-830. 45. Li T, Snyder WK, Olsson JE, Dryja TP. Transgenic mice carrying the dominant rhodopsin mutation P347S: evidence for defective vectorial transport of rhodopsin to the outer segments. Proc Natl Acad Sci USA. 1996;93:l4l76-l4l81. IOVS, December 1998, Vol. 39, No. 13 46. Yau K-W, Baylor DA. Cyclic nucleotide-activated conductance of retinal photoreceptor cells. Annu Rev Neurosci. 1989; 12:289327. 47. Yau K-W. Phototransduction mechanism in retinal rods and cones. The Friedenwald Lecture. Invest Ophthahnol Vis Sci. 1994;35:9-32. 48. Lagnado L, Baylor D. Signal flow in visual transduction. Neuron. 1992;8:995-1002. 49. Copenhagen DR, Jahr CE. Release of endogenous excitatory amino acids from turtle photoreceptors. Nature. 1989;34l:536539. 50. Stryer L. Cyclic GMP cascade of vision. Annu Rev Neurosci. 1986;9:87-H951. Stryer L. Visual excitation and recovery. / Biol Chem. 1991;266: 10711-10714. 52. Pugh EN, Lamb TD. Amplification and kinetics of the activation steps in phototransduction. Biochim Biophys Acta. 1993;ll4l: 111-149. 53. Liebman PA, Parker KR, Dratz EA. The molecular mechanism of visual excitation and its relation to the structure and composition of the rod outer segment. Annu Rev Physiol. 1987;49:765-791. 54. Kaupp UB, Koch K-W. Role of cGMP and Ca2+ in vertebrate photoreceptor excitation and adaptation. Annu Rev Physiol. 1992;54:153-175. 55. Koch, K-W, Stryer L. Highly co-operative feedback control of retinal rod guanylate cyclase by calcium ions. Nature. 1988;334: 64-66. 56. Palczewski K, Subbaraya I, Gorczyca WA, et al. Molecular cloning and characterization of retinal photoreceptor guanylyl cyclaseactivating protein. Neuron. 1994; 13:395-404. 57. Dizhoor AM, Olshevskaya EV, Henzel WJ, et al. Cloning, sequencing, and expression of a 24-kDa Ca(2+)-binding protein activating photoreceptor guanylyl cyclase. / Biol Chem. 1995;270:2520025206. 58. Kawamura S. Rhodopsin phosphorylation as a mechanism of cyclic GMP phosphodiesterase regulation by S-modulin. Nature. 1993;362:855-857. 59. Hsu Y-T, Molday RS. Modulation of the cGMP-gated channel of rod photoreceptor cells by calmodulin. Nature. 1993361:76-7960. Fesenko EE, Kolesnikov SS, Lyubarsky AL. Induction by cyclic GMP of cationic conductance in plasma membrane of retinal rod outer segment. Nature. 1985;313:31O-313. 61. Cook NJ, Kaupp UB. The cGMP-dependent cation channel from vertebrate photoreceptors: purification and functional reconstitution. Photobiochem Photobiophys. 1986;13:331-343. 62. Cook NJ, Hanke W, Kaupp UB. Identification, purification and functional reconstitution of the cyclic GMP-dependent channel from rod photoreceptors. Proc Natl Acad Sci USA. 1987;84:585589. 63. Clack JW, Stein PJ. Opsin exhibits cGMP-activated single-channel activity. Proc Natl Acad Sci USA. 1988;85:9806-9810. 64. Matesic D, Liebman PA. cGMP-dependenr cation channel of retinal rod outer segments. Nature. 1987;326:600-603. 65. Shinozawa T, Sokabe M. Terada S, Matsuska H, Yoshizawa T. Detection of cyclic GMP binding protein and ion channel activity in frog rod outer segments. J Biochem. 1987;102:281-290. 66. Cook NJ, Molday LL, Reid D, Kaupp UB, Molday RS. The cGMPgated channel of bovine rod photoreceptors is localized exclusively in the plasma membrane. / Biol Chem. 1989;264:69966999. 67. Molday RS, Cook NJ, Kaupp UB, Molday RS. The cGMP-gated cation channel of bovine rod photoreceptor cells is associated with a 240 kDa protein exhibiting immunochemical crossreactivity with spectrin./Biol Chem. 1990;265:18690-18695. 68. Molday RS, Molday LL, Dose A, et al. The cGMP-gated channel of the rod photoreceptor cell. Characterization and orientation of the amino terminus. J Biol Chem. 1991;266:21917-21922. 69. Kim T, Reid DM, Molday RS. Structure-function relationships and localization of the Na/Ca-K exchanger in rod photoreceptors. J Biol Chem. 1998;273:l656l-l6567. The Friedenwald Lecture IOVS, December 1998, Vol. 39, No. 13 70. Wiissle H, Griinert U, Cook NJ, Molday RS. The cGMP-gated channel of rod outer segments is not localized in bipolar cells of the mammalian retina. Neurosci Lett. 1992;134:199-202. 71. Kaupp UB, Niidome T, Tanabe T, et al. Primary structure and functional expression from complementary DNA of the rod photoreceptor cyclic GMP-gated channel. Nature. 1989;343:762766. 72. Dhallan RS, Macke JP, Eddy RL, et al. Human rod photoreceptor cGMP-gated channel: amino acid sequence, gene structure, and functional expression./. Neuroscience. 1992;12:3248-3256. 73. Chen T-Y, Peng Y-W, Dhallan RS, Ahamed B, Reed RR, Yau K-W. A new subunit of the cyclic nucleotide-gated cation channel in retinal rods. Nature. 1993;362:764-767. 74. Chen T-Y, Ming M, Molday LL, Hsu Y-T, Yau K-W, Molday RS. Subunit 2 (or j3) of retinal rod cGMP-gated cation channel is a component of the 240 kDa channel-associated protein and mediates Ca2+-calmodulin modulation. Proc Natl Acad Sci USA. 1994; 91:11757-11761. 75. Korschen HG, Ming M, Seifert R, et al. A 240 kDa protein represents the complete subunit of the cyclic nucleotide-gated channel from rod photoreceptor. Neuron. 1995; 15:627-636. 76. Sugimoto Y, Yatsunami K, Tsujimoto M, Khorana HG, Ichikawa A. The amino acid sequence of a glutamic acid-rich protein from bovine retina as deduced form the cDNA sequence. Proc Natl AcadSci USA. 1991;88:31l6-3119. 77. Wohlfart P, Haase W, Molday RS, Cook NJ. Antibodies against synthetic peptides used to determine the topology and site of glycosylation of the cGMP-gated channel from bovine rod photoreceptors. / Biol Cbem 1992;267:644-648. 78. Bonigk W, Altenhofen W, Muller F, et al. Rod and cone photoreceptor cells express distinct genes for cGMP-gated channels. Neuron. 1993;10:865-877. 79. Molday RS, Hsu Y-T. The cGMP-gated channel of photoreceptor cells: its structural properties and role in phototransduction. Behav Brain Set. 1995;18:44l-451. 80. Molday RS. Calmodulin regulation of cyclic-nucleotide-gated channels. Curr Opin Neurobiol. 1996;6:445-452. 81. Heginbotham L, Abramson T, MacKinnon R. A functional connection between the pores of distantly-related ion channels as revealed by mutant K+ channels. Science. 1992;258:1152-1155. 82. Root MJ, MacKinnon R. Identification of an external divalent cation-binding site in the pore of a cGMP-activated channel. Neuron. 1993; 11:459-466. 83- Eismann E, Muller F, Heinemann SH, Kaupp UB. A single negative charge within the pore region of a cGMP-gated channel controls rectification, Ca2+ blockage and ionic selectivity. Proc Natl Acad Sci USA. 1994;91:11O9-1H3. 84. Colville C, Molday RS. Primary structure and expression of the human /3-subunit and related proteins of the rod photoreceptor cGMP-gated channel./Biol Cbem. 1996;271:32968-32974. 85. Ardell MD, Makhija AK, Oliveira L, Miniou P, Viegas-Pequignot E, Pittler SJ. CDNA, gene structure and chromosomal localization of human GAR1 (CNCG3L), a homolog of the third subunit of bovine photoreceptor cGMP-gated channel. Genomics. 1995;28: 32-38. 86. Liu DT, Tibbs GR, Siegelbaum SA. Subunit stoichiometry of cyclic nucleotide-gated channels and effects of subunit order on channel function. Neuron. 1996;l6:983-990. 87. Sun Z-P, Akabas MH, Goulding EH, Karlin A, Siegelbaum SA. Exposure of residues in the cyclic nucleotide-gated channel pore: P-region structure and function in gating. Neuron. 1996;l6:l4l149. 88. Doyle DA, Cabral JM, Pfuetzner RA, et al. The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science. 1998;280:69-77. 89- Hsu Y-T, Molday RS. Interaction of calmodulin with the cyclic GMP-gated channel of rod photoreceptor cells. / Biol Cbem. 1994;269:29765-29770. 90. Gordon SE, Downing-Park J, Zimmerman AE. Modulation of the cGMP-gated ion channel in frog rods by calmodulin and an endogenous inhibitory factor. / Physiol. 1995;486:533-546. 91. Grunwald ME, Yu W-P, Yu H-H, Yau K-W. Identification of a domain on the /3-subunit of the rod cGMP-gated cation channel 92. 93. 94. 95. 96. 97. 98. 99. 100. 101. 102. 103. 104. 105. 106. 107. 108. 109. 110. 111. 112. 113114. 2511 that mediates inhibition by calcium-calmodulin. / Biol Cbem. 1998;273:9148-9157. Dietmar W, Zoche M, Muller F, et al. Calmodulin controls the rod photoreceptor CNG channel through an unconventional binding site in the N-terminus of the /3-subunit. EMBOJ. 1998;17:22732284. Chen T-Y, Yau K-W. Direct modulation by Ca2+-calmodulin of cyclic nucleotide-activated channel of rat olfactory receptor neurons. Nature. 1994;368:545-548. Liu M, Chen T-Y, Ahamed B, Li J, Yau K-W. Calcium-calmodulin modulation of the olfactory cyclic nucleotide-gated cation channel. Science. 1994;266:1348-1354. Lowry OH, Roberts NR, Schulz DW, Clow JE, Clark JR. Quantitative histochemistry of the retina, II: enzymes of glucose metabolism. J Biol Cbem. 196l;236:2813-2820. Futterman S. Metabolism of the retina, III: the role of reduced triphosphopyridine nucleotide in the visual cycle. / Biol Cbem. 1963;238:ll45-1150. McConnell DG, Ozga GW, Solze DA. Evidence for glycolysis in bovine retinal microsomes and photoreceptor outer segments. Biochim Biophys Acta. 1969; 184:11 -28. Futterman S, Hendrickson A, Bishop PE, Rollins MH, Vacano E. Metabolism of glucose and reduction of retinaldehyde in retinal photoreceptors. / Neurocbem. 1970; 17:149 -156. Wallimann T, Wegmann G, Moser H, Huber R, Eppenberger HM. High content of creatine kinase in chicken retina: compartmentalized localization of creatine isoenzymes in photoreceptor cells. Proc Natl Acad Sci USA. 1986;83:3816 - 3819. Hsu S-C, Molday RS. Glyceraldehyde-3-phosphate dehydrogenase is a major protein associated with the plasma membrane of retinal photoreceptor outer segments./ Biol Cbem. 1990;265:1330813313. Hsu S-C, Molday RS. Glycolytic enzymes and a GLUT-1 glucose transporter in the outer segments of rod and cone photoreceptor cells. / Biol Cbem. 1991;266:21745-21752. Hsu S-C, Molday RS. Glucose metabolism in photoreceptor outer segments. / Biol Cbem. 1994;269:17954-17959. Lopez-Escalera R, Li X-B, Szerencsei RT, Schnetkamp PPM. Glycolysis and glucose uptake in intact outer segments isolated from bovine retinal rods. Biochemistry. 1991 ;30:8970-8976. Winkler BS. Glycolytic and oxidative metabolism in relation to retinal function. / Gen Pbysiol. 1981;77:667-69l. Winkler BS, Dang L, Malinoski C, Easter SS. An assessment of rat photoreceptor sensitivity to mitochondrial blockade. Invest Opbtbalmol Vis Sci. 1997;38:1569 -1577. Dryja TP, McGee TL, Reichel E, et al. A point mutation of the rhodopsin gene in one form of retinitis pigmentosa. Nature. 1990;343:364-366. Berson EL. Retinitis pigmentosa. The Friedenwald Lecture. Invest Ophthalmol Vis Sci. 1993;34:l659-l676. Daiger SP, Sullivan LS, Rossiter BJF. RetNet Retinal Information Network Web site. www.sph.uth.tmc.edu/Retnet/home.htm. Dryja TP, Li T. Molecular genetics of retinitis pigmentosa. Hum Mol Genet. 1995;4:1739-1743. Molday RS, Hicks D, Molday LL. Peripherin: a rim-specific membrane protein of rod outer segment disks. Invest Ophthalmol Vis Sci. 1987;28:50-6l. Connell G, Molday RS. Molecular cloning, primary structure, and orientation of the vertebrate photoreceptor cell protein peripherin in the rod outer segment disk membrane. Biochemistry. 1990;29:4691-4698. Bascom RA, Manara S, Collins L, Molday RS, Kalnins VI., Mclnnes RR. Molecular cloning of the cDNA for a novel photoreceptorspecific membrane protein (rom-1) identifies a disk rim protein family implicated in human degenerative retinopathies. Neuron. 1992;8:1171-1184. Moritz OL, Molday RS. Molecular cloning, membrane topology and localization of bovine rom-1 in rod and cone photoreceptor cells. Invest Ophthalmol Vis Sci. 1996;37:352-362. Molday RS. Peripherin/rds and rom-1: molecular properties and role in photoreceptor cell degeneration. Prog Retinal Eye Res. 1994;13:271-299. 2512 Molday 115. Goldberg AFX, Moritz OL, Molday RS. Heterologous expression of photoreceptor peripherin/rds and rom-1 in COS-1 cells: Assembly, interactions and localization of multisubunit complexes. Biochemistry. 1995:34:14213-14219. 116. Goldberg AFX, Molday RS. Subunit composition of the peripherin/rds-rom-1 disk rim complex from rod photoreceptors: hydrodynamic evidence for a tetrameric quaternary structure. Biochemistry. 1996;35:6l44-6l49. 117. Loewen CR, Molday RS. Oligomerization of the peripherin/w/srom-1 complex: possible role in disk structure and retinitis pigmentosa. Invest Ophthalmol Vis Set. 1998;39(4):S963- Abstract nr 4446. 118. Goldberg AFX, Loewen CJ, Molday RS. Cysteine residues of photoreceptor peripherin/rds: role in subunit assembly and autosomal dominant retinitis pigmentosa. Biochemistry. 1998;37:680685. 119. Arikawa K, Molday LL, Molday RS, Williams DS. Localization of peripherin/rds in the disk membranes of cone and rod photoreceptors: relationship to disk membrane morphogenesis and retinal degeneration./ Cell Biol. 1992;116:659-667. 120. Sanyal S, Jansen HG. Absence of receptor outer segments in the retina of rds mutant mice. Neurosci Lett. 1981;21:23-26. 121. Jansen HG, Sanyal S. Development and degeneration of retina in rds mutant mice. Electron microscopy. / Comp Neurol. 1984; 224:71-84. 122. Hawkins RK, Jansen HG, Sanyal S. Development and degeneration of retina in rds mutant mice: photoreceptor abnormalities in the heterozygotes. Exp Eye Res. 1985;4l:701-720. 123. Travis GH, Brennan MB, Danielson PE, Kozak CA, Sutcliffe JG. Identification of a photoreceptor specific mRNA incoded by the gene responsible for retinal degeneration slow (rds). Nature. 1989;338:70-73. 124. Connell G, Bascom R, Molday LL, Reid D, Mclnnes RR, Molday RS. Photoreceptor peripherin is the normal product of the gene responsible for retinal degeneration in the rds mouse. Proc Natl AcadSci USA. 1991;88:723-726. 125. Travis GL, Groshan KR, Lloyd M, Bok D. Complete rescue of photoreceptor dysplasia and degeneration in transgenic retinal degneration slow (rds) mice. Neuron. 1992;9:113-119. 126. Farrar GJ, Kenna P, Jardan SA, et al. A three-base pair deletion in the peripherin-rds gene in one form of retinitis pigmentosa. Nature. 1991 ;354:478- 480. 127. Kaijawra K, Hahn LB, Mukai S, Travis GH, Berson EL, Dryja TP. Mutations in the human retinal degeneration slow gene in autosomal dominant retinitis pigmentosa. Nature. 1991;354:480483. 128. Griming G, Milan, JM, Meins M, et al. Mutations in the human peripherin/RDS gene associated with autosomal dominant retinitis pigmentosa. Hum Mutat. 1994:321-323. 129. Keen TJ, Inglehearn CF, Kim R, Bird AC, Battacharya SS. Retinal pattern dystrophy associated with a 4 bp insertion at codon 140 in the RDS-peripherin gene. Hum Mot Genet. 1994;3:367-368. 130. Saga M, Mashima Y, Akeo K, Oguchi Y, Kudoh J, Shimizu N. A novel Cys-2l4-Ser mutation in the peripherin/RDS gene in a Japanese family with autosomal dominant retinitis pigmentosa. Hum Genet. 1993:92:519-521. 131. Wells J, Wroblewski J, Keen J, et al. Mutations in the human retinal degeneration slow (RDS) gene can cause either retinitis pigmentosa or macular dystrophy. Nat Genet. 1993:3:213-218. 132. Nicols BE, Sheffeld VC, Vandenburgh K, Drack AV, Kimura AE, Stone EM. Butterfly-shaped pigment dystrophy of the fovea caused by a point mutation in codon 167 of the RDS gene. Nat Genet. 1993:3:202-207. 133. Weleber RG, Carr RE, Murphey WH, Sheffield VC, Stone EM. Phenotypic variation including retinitis pigmentosa, pattern dystrophy and fundus flavimaculatus in a single family with a deletion of codon 153 or 154 of the peripherin/rds gene. Arch Ophthalmol. 1993;111:1531-1542. 134. Gorin MB, Jackson KE, Ferrell RE, et al. A peripherin/retinal degeneration slow mutation (Pro210Arg) associated with macular and peripheral retinal degeneration. Ophthalmology. 1995; 102:246-255. IOVS, December 1998, Vol. 39, No. 13 135. Fishman GA, Stone EM, Alexander KR, Gilbert LD, Derlacki DJ, Butler NS. Serine-27-Phenylalanine mutation within the peripherin/RDS gene in a family with cone dystrophy. Ophthalmology. 1997;104:299-306. 136. Jacobson SG, Cideciyan AV, Maguire AM, Bennett J, Sheffield VC, Stone EM. Preferential rod and cone photoreceptor abnormalities in heterozygotes with point mutations in the RDS gene. Exp Eye Res. 1996;63:603-608. 137. Kajiwara K, Berson EL, Dryja TP. Digenic retinitis pigmentosa due to mutations at the unlinked peripherin/RDS and ROM1 loci. Science. 1994;264:1604 -1608. 138. Dryja TP, Hahn LB, Kajiwara K, Berson EL. Dominant and digenic mutations in the peripherin/RDS and ROM1 genes in retinitis pigmentosa. Invest Ophthalmol Vis Sci. 1997:38:1972-1982. 139- Goldberg AFX, Molday RS. Defective subunit assembly underlies a digenic form of retinitis pigmentosa linked to mutations in peripherin/rds and rom-1. Proc Natl Acacl Sci USA. 1996;93: 13726-13730. 140. Clarke GA, Rossant J, Mclnnes RR. Rom-1 is required for outer segment morphogenesis and photoreceptor viability [ARVO Abstract]. Invest Ophthalmol Vis Sci. 1998;39(4):S962. Abstract nr 4442. 141. Papermaster DS, Schneider BG, Zorn MA, Kraehenbuhl JP. Immunocytochemical localization of a large intrinsic membrane protein to the incisure and margins of frog rod outer segment disks. J Cell Biol. 1978;78:4l5-425. 142. Papermaster DS, Reilly P, Schneider BG. Cone lamellae and red and green rod outer segment disks contain a large intrinsic membrane protein on their margins: an ultrastructural immunocytochemical study of frog retinas. Vision Res. 1982;22:l4l71428. 143. Molday RS, Molday LL. Identification and characterization of multiple forms of rhodopsin and minor proteins in frog and bovine rod outer segment disc membranes. / Biol Chem. 1979;254: 4653-4660. 144. Converse CA. The large intrinsic membrane protein in rod outer segments: in vitro synthesis in cattle, and comparison in humans and rabbits. Exp Eye Res. 1979;29:409-l6. 145. llling M, Molday LL, Molday RS. The 220 kDa rim protein of retinal rod outer segments is a member of the ABC transporter superfamily./Biol Chem. 1997;272:10303-10310. 146. Higgins CF. ABC transporters: from microorganisms to man. Annu Rev Cell Biol. 1992;8:67-113. 147. Sun H, Nathans J. Stargardt's ABCR is localized to the disc membrane of retinal rod outer segments. Nat Genet. 1997; 17:15-6. 148. Azarian SM. Travis GH. The photoreceptor rim protein is an ABC transporter encoded by the gene for recessive Stargardt's disease (ABCR). FEES Lett. 1997;409:247-252. 149- Allikmets R, Singh N, Sun H, et al. A photoreceptor cell-specific ATP-binding transporter (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet. 1997; 15:236-246. 150. Kaplan J, Gerber S, Larget-Piet, D, et al. A gene for Stargardt's disease (fundus flavimaculatus) maps to the short arm of chromosome I. Nat Genet. 1993;5:308-3U. 151. Blacharski PA. Fundus flavimaculatus. In: Newsome DA, ed. Retinal Dystrophies and Degeneration. New York: Raven Press; 1988:135-159. 152. Nassonkin 1, llling M, Koehler MR, Schmid M, Molday RS, Weber BHF. Mapping of the rod photoreceptor ABC transporter (ABCR) to Ip21-p22.1 and identification of novel mutations in Stargardt's disease. Hum Genet. 1998; 102:21-26. 153. Allikmets R, Shroyer NF, Singh N, et al. Mutation of the Stargardt disease gene (ABCR) in age-related macular degeneration. Science. 1997;277:1805-1807. 154. Martinez-Mir A, Paloma E, Allikmets R, et al. Retinitis pigmentosa caused by a homozygous mutation in the Stargardt disease gene ABCR. Nat Genet. 1998; 18:11 -12. 155. Cremers FP, van de Pol DJ, van Driel M, et al. Autosomal recessive retinitis pigmentosa and cone-rod dystrophy caused by splice site mutations in the Stargardt's disease gene ABCR. Hum Mol Genet. 1998;7:355-362. IOVS, December 1998, Vol. 39, No. 13 156. Luciani M-F, Chimini G. The ATP binding cassette transporter ABC1 is required for the engulfment of corpses generated by apoptotic cell death. EMBOJ. 1996;15:226-235. 157. Swain PK, Chen S, Wang QL, et al. Mutations in the cone-rod homeobox gene are associated with the cone-rod dystrophy photoreceptor degeneration. Neuron. 1998;19:1329-1336. 158. Freund CL, Gregory-Evans CY, Furukawa T, et al. Cone-rod dystrophy due to mutations in a novel photoreceptor-specific homeobox gene (CRX) essential for maintenance of the photoreceptor. Cell. 1997;91:543-553. 159- Maw MA, Kennedy B, Knight A, et al. Mutation of the gene encoding cellular retinaldehyde-binding protein in autosomal recessive retinitis pigmentosa. Nat Genet. 1997;17:198-200. 160. Marlhens F, Bareil C, Griffoin JM, et al. Mutations in RPE65 cause Leber's congenital amaurosis. Nature Genetics. 1997;17:139-l4l. 161. Morimura H, Fishman GA, Grover SA, Fulton AB, Berson EL, Dryja TP. Mutations in the RPE65 gene in patients with autosomal recessive retinitis pigmentosa or Leber congenital amaurosis. Proc Natl Acacl Sci USA. 1998;95:3088-3093162. Weber BHF, Vogt G, Pruett RC, Stohr H, Felbor U. Mutations in the tissue inhibitor of metalloproteinase-3 (TIMP3) in patients with Sorby's fundus dystrophy. Nat Genet. 1994;8:352-356. 163. Petrukhin K, Koisti MJ, Bakall B, et al. Identification of the gene responsible for Best macular dystrophy. Nat Genet. 1998;19:24l-247. 164. Marquardt A, Stohr H, Passmore LA, Kramer F, Rivera A, Weber BHF. Mutations in a novel gene, VMD2, encoding a protein of unknown properties cause juvenile-onset vitelliform macular dystrophy (Best's disease). Hum Mol Genet. 1998;7:1517-1525. 165. Payne AM, Downes SM, Bessant DA, et al. A mutation in guanylate cyclase activator 1A (GUCA1A) in an autosomal dominant cone dystrophy pedigree mapping to a new locus on chromosome 6p21.1. Hum Mol Genet. 1998;7:273-277. 166. Dryja TP, Finn JT, Peng Y-W, McGee TL, Berson EL, Yau K-W. Mutations in the gene encoding the a subunit of the rod cGMPgated channel in autosomal recessive retinitis pigmentosa. Proc Natl Acad Sci USA. 1995:92:10177-10181. 167. Perrault I, Rozet JM, Calvas P, et al. Retinal-specific guanylate cyclase gene mutations in Leber's congenital amaurosis. Nat Genet. 1996;l4:46l-464. 168. McLaughlin ME, Sandberg MA, Berson EL, Dryja TP. Recessive mutations in the gene encoding the /3 subunit of rod phosphodiesterase in patients with retinitis pigmentosa. Nat Genet. 1993; 4:130-134. 169. Huang SH, Pittler SJ, Huang X, Oliveira L, Berson EL, Dryja TP. Autosomal recessive retinitis pigmentosa caused by mutations in the a-subunit of rod cGMP phosphodiesterase. Nat Genet. 1995; 11:468-471. 170. Sauer CG, Gehrig A, Warneke-Wittstock R, et al. Positional cloning of the gene associated with X-linked juvenile retinoschisis. Nat Genet. 1997;17:l64-170. The Friedenwald Lecture 2513 171. Weil D, Blanchard S, Kaplan J, et al. Defective myosin VILA gene responsible for Usher syndrome type IB. Nature. 1995;374: 60-1. 172. Meindl A, Dry K, Herrmann K, et al. A gene (RPGR) with homology to the RCC1 guanine nucleotide exchange factor is mutated in X-linked retinitis pigmentosa (RP3). Nat Genet. 1996; 13:3542. 173. Fuchs S, Nakazawa M, Maw M, Tamai M, Oguchi Y, Gal A. A homozygous 1-base pair deletion in the arrestin gene is a frequent cause of Oguchi disease in Japanese. Nat Genet. 1995;10:360362. 174. Yamamoto S, Yamamoto S, Sippel KC, Berson EL, Dryja TP. Defects in the rhodopsin kinase gene in the Oguchi form of stationary night blindness. Nat Genet. 1997; 15:175178. 175. Dryja TP, Hahn LB, Reboul T, Arnaud B. Missense mutation in the gene encoding the alpha subunit of rod transducin in the Nougaret form of congenital stationary' night blindness. Nat Genet. 1996;13:358-360. 176. Rosenfeld PJ, Cowley GS, McGee TL, Sandberg MA, Berson EL, Dryja TP. A null mutation in the rhodopsin gene causes rod photoreceptor dysfunction and autosomal recessive retinitis pigmentosa. Nat Genet. 1992;l:209-213. 177. Gal A, Orth U, Baehr W, Schwinger E, Rosenberg T. Heterozygous missense mutation in the rod cGMP phosphodiesterase /3-subunit gene in autosomal dominant stationary night blindness. Nat Genet. 1994;7:64-68. 178. Dryja TP, Berson EL, Rao VR, Oprian DD. Heterozygous missense mutation in the rhodopsin gene as a cause of congenital stationary night blindness. Nat Genet. 1993;4:280-283. 179. Sieving PA, Richards JE, Naarendorp E, Bingham EL, Scott K, Alpern M. Dark-light: Model for nightblindness from the human rhodopsin Gly90Asp mutation. Proc Natl Acad Sci USA. 1995;92: 880-884. 180. Perrault I, Rozet JM, Gerber S, et al. A retGC-1 mutation in autosomal dominant cone-rod dystrophy. Am J Hum Genet. 1998;63:651-654. 181. Kelsell RE, Gregory-Evans K, Payne AM, et al. Mutations in the retinal guanylate cyclase (RETGC-1) gene in dominant cone-rod dystrophy. Hum Mol Genet. 1998;7:1179-H84. 182. Yan D, Swain PK, Breuer D, et al. Biochemical characterization and subcellular localization of the mouse retinitis pigmentosa GTPase regulator (mRPgr). / Biol Chem. 1998;273:1965619663. 183. Haeseleer F, Huang J, Lebioda L, Saari JC, Palczewski K. Molecular characterization of a novel short-chain dehydrogenase/reductase that reduces M-trans retinal. / Biol Chem. 1998;273:2179021799.