Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
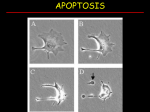
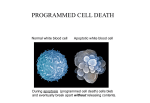
Chem*3560 Lecture 13: Caspases and Programmed Cell Death In a multicellular organism, there are a number of circumstances in which individual cells are sacrificed for the benefit of the organism as a whole. A process exists for the orderly dissolution of the cellular components so that they can be absorbed and used by neighbours. This is the process of programmed cell death or apoptosis (pronounced apo’ptosis, not apop’tosis). The Greek word means “falling off”, as of autumn leaves. Catastrophic cell death or Necrosis Programmed cell death or Apoptosis Pathological response to injury Normal physiological response to internal or external signals Cytoplasm shrinks without membrane rupture Cell membranes pinch off into smaller units Cell contents are packaged to be absorbed by neighbours; no inflammation Organelles swell and rupture Cell membrane lyses Cell contents spill out Inflammation When does apoptosis happen? In embryonic and fetal development: sculpting of embryonic form organization of the nervous system. In the adult: on stimulation by the immune system in response to DNA damage or abnormality, due to chemical or radiation damage, or viral infection on withdrawal of supporting hormones in tumours. How does apoptosis happen? Apoptosis occurs when proteolytic enzymes in the cell called Caspases become activated. Caspase stands for Cys-catalysed Aspartate targeting protease, i.e. Cysteine is the catalytic nucleophile, and attacks sequences at aspartate. Preferred target sequences follow the polar/non-polar pattern -Asp-Glu-Val-Asp-, with the cut coming after the second Asp. The sequence -Asp-Glu-Ala-Asp-, which in single letter codes reads -DEAD- is also a target. Mammalian cells contain 14 currently known caspases, but many of these are involved in inflammatory responses rather than apoptosis. Two subgroups of caspases are involved in apoptosis: Caspases 3 and 7 are designated as effector or executioner caspases because they carry out the cleavages that shut down the cell’s operations. Caspases 8 and 9 are designated as initiator caspases, because they activate caspases 3 and 7. Caspase activation Caspases are synthesized as inactive procaspases, from which the N-terminal region is cut. A second cut is critical for activation and divides the caspase into 20 kDa and 10 kDa segments which remain attached through H-bonds. The activating cut severs a loop (L2 in the figure), and reorients the catalytic Cys from an inaccessible position to one that can interact with substrate. The sequence at the activating cut site is a caspase target sequence, so caspases can activate procaspases. Caspase substrates The executioner caspases 3 and 7 are highly selective in their targets, but are geared to shut down the cell’s activities down irrevocably: Structural nuclear proteins DNA repair and replication proteins Cytoskeleton proteins actin, spectrin, keratin Many protein kinases, particularly those controlling cytoskeleton and cell proliferation One particular target is the Inhibitor of Caspase Activated DNase, ICAD. When the inhibitor is destroyed, its companion DNase sets about cutting DNA in the linker regions between nucleosomes, approximately every 200 base-pairs. The cut DNA appears as a “ladder” pattern of DNA fragments in gel electrophoresis, which is one of the key experimental indicators that apoptosis is occurring. The substrates of initiator caspases 8 and 9 are procaspases 3 and 7, as well as their own procaspases. This means that once the process starts, a positive feedback loop leads to escalating caspase activation. How is apoptosis induced? Apoptosis may be initiated internally, or signalled from outside the cell, frequently induced by the action of immune system cells such as T-lymphocytes. The internal signal activates procaspase 9, whereas the external signal activates procaspase 8. Whichever initiator caspase is first activated, the end result will be the activation of all the apoptosis-related caspases Internal pathway for inducing apoptosis Internal control over apoptosis is maintained by a family of proteins called Bcl-2 proteins which are closely associated with mitochondria. Bcl-2 itself is anti-apoptotic, but a related protein Bax is pro-apoptotic. So long as the balance is in favour of Bcl-2, the cell functions normally. If the balance is shifted in favour of Bax, apoptosis may be induced. When DNA damage is detected during replication, the damage response activates DNA repair processes, as well as the cell proliferation suppressor called p53. p53 is a transcription factor which controls expression of a number of proteins that halt cell cycle progression. These include the cyclin kinase inhibitor p21, which inhibits Cyclin A-Cdk2. This causes the cell cycle to stall while repairs are undertaken. Meanwhile Bax is being expressed. If DNA repairs take too long, Bax levels build up to the critical point of excess over Bcl-2, and this results in release of cytochrome c from mitochondria. When cytochrome c enters the cytoplasm, it causes aggregation of a protein called Apaf-1 (apoptosis promoting and activating factor). The N-terminus of Apaf-1 has a segment called the Caspase recruitment domain or CARD, and the CARD domain becomes exposed when Apaf-1 binds cytochrome C. CARD domains tend to stick to other CARD domains, and Procaspase 9 also has a CARD domain at its N-terminus (making it a CARD-carrying protein). The result is a localized build up of Procaspase 9 molecules that pair up around Apaf-1 molecules. The initiator procaspase has weak activity that is ineffective if the procaspase molecules are dispersed, but when held close together in pairs, eventually one will activate the other and set the whole apoptosis process off. Externally signalled apoptosis Cell surface membranes carry a variety of proteins that act as receptors for extracellular signals. A set of receptors called Tumor Necrosis Factor receptors (TNF-R), and a related receptor called FAS are found on many cell surfaces. The signals they respond to take the form of protein ligands, secreted by or carried on the membranes of cells of the immune system. In the absence of ligand, the receptors are randomly dispersed on the surface of the cell, and have no effect on the cell’s fate. When the ligand binds, this induces the receptors to cluster together. The portion of the receptor inside the membrane carries a protein binding domain called the death domain, which binds other proteins carrying death domains. Binding is too weak to act at single receptors, but becomes effective when receptors group in a cluster. Death domains are found on small adapter proteins that carry the death domain at one end and a death effector domain (DED) at the other end. Death effector domains are also protain binding domains that associate with like domains. Procaspase 8 carries a death effector domain at its N-terminus, so procaspase 8 molecules become paired up when they bind to the receptor-adapter complex. Procaspase 8 has weak activity that is ineffective if the procaspase molecules are dispersed, but when paired up, eventually one will activate the other and set the whole apoptosis process off.. IAPs (inhibitors of apoptosis) protect against accidents IAPs are proteins that bind to the catalytic site of caspases, and trick the caspase by inserting a peptide chain in the reverse direction relative to a normal caspase substrate. The amount of IAP is small, so it can cope with occasional accidental activation of a caspase, but if a real signal exists, too much caspase is produced for IAPs to deal with. The IAP-Caspase complex becomes a target for ubiquitin ligase, and the ubiquitin tag is added to the unwanted caspase, marking it for destruction.