Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Neural coding wikipedia , lookup
Neural oscillation wikipedia , lookup
Development of the nervous system wikipedia , lookup
Neural modeling fields wikipedia , lookup
Optogenetics wikipedia , lookup
Holonomic brain theory wikipedia , lookup
Artificial neural network wikipedia , lookup
Biological neuron model wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Central pattern generator wikipedia , lookup
Metastability in the brain wikipedia , lookup
Nervous system network models wikipedia , lookup
Node of Ranvier wikipedia , lookup
Convolutional neural network wikipedia , lookup
Catastrophic interference wikipedia , lookup
Hierarchical temporal memory wikipedia , lookup
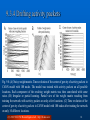
9. Continuous attractor and competitive networks Fundamentals of Computational Neuroscience, T. P. Trappenberg, 2002. Lecture Notes on Brain and Computation Byoung-Tak Zhang Biointelligence Laboratory School of Computer Science and Engineering Graduate Programs in Cognitive Science, Brain Science and Bioinformatics Brain-Mind-Behavior Concentration Program Seoul National University E-mail: [email protected] This material is available online at http://bi.snu.ac.kr/ 1 Outline 9.1 9.2 9.3 9.4 Spatial representations and the sense of direction Learning with continuous pattern representations Asymptotic states and the dynamics of neural fields ‘Path’ integration, Hebbian trace rule, and sequence learning 9.5 Competitive networks and self-organizing maps (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 2 9.1 Spatial representations and the sense of direction Auto-associative attractor models General memory states in mind The shape of objects, their smell, texture, or color Point attractor neural networks (PANNs) Memory represented by independent vectors Continuous attractor neural networks (CANNs) The training patterns represent continuous Spatial location of an object Topographic maps (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 3 9.1.1 Head direction The sense of direction Representation of body or head direction A mechanism to update this information without visual cues Fig. 9.1 (A) Experimental response of a neuron in the subiculum of a rodent when the rodent is heading in different directions in a familiar maze. The dashed line represents the new head properties of the same neuron when the rodent is placed in the new unfamiliar maze. The new response properties will normally be similar to the previous one, that is, head direction cells try to maintain approximately their response properties to specific head directions. However, the results shown were produced in experiments with a rodent that had cortical lesions that weakened the ability to maintain the response properties after the rodent was transferred into a new environment. (B) Neuronal response from many hippocampal neurons in a rodent that responded to the subject’s location (places) in a maze. The figure shows the firing rates of the neurons in response to a particular place. whereby the neurons were placed in the figure so that neurons with similar response properties were placed adjacent to each other. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 4 9.1.2 Place fields Head direction representations The spatial representation of a one-dimensional feature space in the brain Apply equally to higher-dimensional representations Neurons in the hippocampus of rats Fire in relation to specific locations within a maze A specific topography of neurons within the hippocampal tissue with respect to their maximal response to a particular place has not been found The rearrangement (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 5 9.1.3 Spatial representations in network models A possible solution to representing head directions Fig. 9.2 A proposal as to how the activity of nodes, for clarity arranged into a circle, can represent head directions. With the 20 nodes of this model we can represent head directions with a resolution of 18 degree when using a single binary node as a representation of a head direction. The single active node in the figure, represented as a solid circle, indicates a head direction of 72 degrees in this example. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 6 9.1.4 Graded winner-take-all models Winner-take-all Only one node or one activity packet of nodes The dynamic equation for the networks 1 dhi (t ) 1 hi (t ) dt N ext i (t ) j (9.1) h(x, t ) hi (x, t ) w(x, y )r (y, t )dy1 yd I ext (x, t ) y1 yd (9.2) t 1-dimension ij j Neural field equation w r (t ) I h( x, t ) hi ( x, t ) w( x, y )r ( y, t )dy I ext ( x, t ) y t (9.3) The discretization rules x i x (9.4) dx x h(ix, t ) hi (t ) (9.5) x 1 / N (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 7 9.2 Learning with continuous pattern representations Recurrent neural networks Represent a continuous set of patterns Hebbian learning Hebbian rule for the excitatory weights In the neural field representation w E ( x, y ) r ( x, t ) r ( y , t ) (9.6) The firing rate r μ is the firing rate of the neural field while dominated by the training example of a pattern μ presented to the network The inhibition from inhibitory interneurons w wE c (9.7) (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 8 9.2.1 Learning Gaussian head direction patterns ri e i 2 / 2 2 (9.8) The external input to a node i, Gaussian profile around a preferred direction The displacement between The head direction αHD provided by the external input The optimal firing direction of the cell αi i min(| i HD |,360 | i HD |) (9.9) (9.10) A contribution to each weight component w ri rj e E ij ( i2 2j ) / 2 2 E HD i ) e (9.11) wij ( ( i j ) 2 / 2 2 E HD i n ) e (9.12) wij ( (9.13) w Ne E ij (9.14) [( i j ) 2 ( i j )( n ) 2 ] / 2 2 for a node with a preferred direction different from the direction of the training example For infinite resolution of the model, i.e. Δ α → 0: for a node with a preferred direction equal to that of training example ( i j )2 / 2 2 The weight matrix has a Gaussian shape and the same width as the receptive fields of the nodes The continuous notations wE ( x, y) wE (| x, y |) : weight matrix depends only on the distance between nodes (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 9 9.2.2 Gaussian interaction profiles in the brain An effective interaction structure Short-distance excitation and long-distance inhibition Columnar organizations in the cortex The superior colliculus from cell recordings in monkeys Fig. 9.3 Data from cell recordings in the superior colliculus in a monkey that indicate the interaction strength ρw between cells in this midbrain structure. The solid line displays the corresponding measurement from simulations of a CANN model of this brain structure. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 10 9.2.3 Self-organized interaction structures in CANNs Fig. 9.4 A recurrent associative attractor network model, similar to the model shown in Fig. 9.2, where the nodes have been arbitrarily placed in the physical space on a circle. The relative connection strength between the nodes is indicated by the thickness of the lies between the nodes. Each node responds during learning with a Gaussian firing profile around the stimulus that excites the node maximally. Each node is assigned a center of the receptive field randomly from a pool of centers covering the periodic training domain. (A) Before training all nodes have the same relative weights between them. (B) After training the relative weight structure has changed with a few strong connections and some weaker connections. (C) The regularities of the interactions can be revealed when reordering the nodes so that nodes with the strongest connections are adjacent to each other. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 11 9.3 Asymptotic states and the dynamics of neural fields The asymptotic states (attractors) The weight matrix is shift invariant after training the network on continuous Gaussian patterns Local cooperation and global competition Activity packet A collection of nodes to be active Shift invariant The activity packet can be stabilized at any location in the network depending on an initial external stimulus Dynamic competition (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 12 9.3.1 Attractor regimes The different regimes in the CANN model depend on the level of inhibition c 1. Growing activity The inhibition is weak compared to the excitation 2. Decaying activity The inhibition is strong compared to the excitation 3. Stable activity packet In an intermediate range of the strength of inhibitions relative to that of the excitations Fig. 9.5 (A) Time evolution of the firing rates in a CANN model with 100 nodes. Equal external inputs to nodes 30-70 were applied at t = 0. This external input was removed at t = 10τ. The inhibition was set to three times the average firing rate of a node when driven by a Gaussian external input like that used for training the network. (B) The solid line represents the firing rate profile of the simulation shown in (A) at t = 20τ. The dashed line corresponds to the firing rate profile in a similar simulation with reduced (by a factor of three) inhibition. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 13 9.3.2 Formal analysis of attractor states A threshold activation function g(x) = 1/exp(0.007x) x h ( x ) The stationary state of the dynamics eqn 9.3 x w( x, y)dy (9.15) x h(x1)=h(x2)=0, x w( x1 , y)dy 0 (9.16) x2 x1 E N erf ( ) c( x2 x1 ) For the weighting function w = w - c, 2 2 (9.17) 2 1 2 1 Fig. 9.6 (A) Plot of the functions (9.17) and two linear functions with slope c = 1 and c = 0.4. The intersection of the functions (other than at x2 – x1 = 0) gives the solutions we are seeking of eqn 9.17. (B) The solution of eqn 9.17 as a function of the inhibition constant. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 14 9.3.3 Stability of the activity packet The stability of the activity packet with respect to its movement Calculate the velocity of the boundaries h dx The velocity dt x t (9.18) The centre x (t ) 12 ( x (t ) x (t )) (9.19) The velocity of the centre of the activity packet dxc 1 dt 2x1 c x2 1 2 w( x1 , y )dy 1 2x2 x2 Fig. 9.7 (A) Two Gaussian bell curves x1 x1 (9.20) centered around two different values x1 and x2. The striped and dotted areas are the same due to the symmetry of the bell curve. The integrals from x1 to x2 over the two different curves are therefore the same. This is not true if the two curves are not symmetric and have different shapes. (B) The dashed line outlines the shape of an activity packet from a simulation. The symmetry of this activity packet makes the gradients of boundaries equal except for a sign. 15 (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr w( x2 , y )dy 9.3.4 Drifting activity packets Fig. 9.8 (A) Noisy weight matrix. Time evolution of the center of gravity of activity packets in CANN model with 100 nodes. The model was trained with activity packets on all possible locations. Each component of the resulting weight matrix was then convoluted with some noise. (B) Irregular or partial learning. Partial view of the weight matrix resulting from training the network with activity packets on only a few locations. (C) Time evolution of the center of gravity of activity packets in CANN model with 100 nodes after training the network on only 10 different locations. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 16 9.3.5 Stabilization of the activity packet The drift in the activity packet can be stabilized by a small increase in the excitability of neurons once they have been recently activated NMDA receptor Voltage-dependent nonlinearity An increases of the voltage-dependent nonlinearity would make more states stable Fig. 9.8 (D) ‘NMDA’-stabilization. The network trained o the 10 locations was augmented a stabilization mechanism that reduces the firing threshold of active neurons. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 17 9.4 ‘Path’ integration, Hebbian trace rule, and sequence learning The possibility of ‘updating’ the state A subject might not have an absolute value available Rotate a subject with closed eyes Path integration Calculate the new position from the old position 9.4.1 Path integration with asymmetric weighting functions The path integration problem involves using such asymmetries in a systematic way The strength of the asymmetry to the velocity of the movement Idiothetic cues (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 18 9.4.2 Idiothetic update of head direction representations Fig. 9.9 Model for path integration in CANNs. The central nodes are part of the network with collateral connections as used to represent head directions (Fig. 9.2). The rotation nodes represent collections of neurons that signal rotation velocities proportional to their activity. The afferents of these rotation cells can modulate the collateral connections within the head direction network. We symbolized this with synapses close to the synapses of the collateral connections. Each rotation cell can synapse on to each synapse in the head direction network. The separation of the connections, as indicated by the solid and dashed lines in the figure, is self-organized during learning (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 19 9.4.3 Self-organization of a rotation network Biologically realistic model Self-organize the network Clockwise rotation Clockwise synapse Short-term memory Trace term ri (t 1) (1 )ri (t ) ri (t ) (9.21) The weight between rotation nodes with Hebbian rule rot wijk kri rj rkrot (9.22) The rule strengthens the weights between the rotation node and the appropriate synapses in the recurrent network (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 20 9.4.4 Updating the network after learning The dynamics of the model h( x, t ) h( x, t ) weff ( x, y, r rot )r ( y, t )ri rot (t )dy (9.23) y t rot rot (9.24) wijeff ( wij c)(1 wijk rk ) Fig. 9. 10 (A) Simulation of a CANN model with idiothetic updating mechanisms. The acitivity packet can be moved with idiothetic inputs in either clockwise or anti-clockwise directions, depending on the firing rates of the corresponding rotation cells. (B) The different weighting functions from node 50 to the other nodes in the network after learning. w, solid line; wrot, dashed line, weff, dotted line. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 21 9.4.5 Sequence learning Sequence learning Apply the generic mechanisms of asymmetric weighting functions Including a trace term (in pattern space) in the canonical learning rule wij 1 N ( i j i j ) 1 (9.25) For a sufficient strength of the asymmetric component Using the strength parameter The network is able to jump between the patterns in sequences (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 22 9.5 Competitive networks and self-organizing maps 9.5.1 Two-dimensional SOM Two-dimensional feature vectors r1in r in r2 (9.26) in Fig. 9.11 Architecture of a twodimensional self-organizing map. Each of the two input values rin1 and rin2 , each representing one of two feature components, is mapped on the map network with individual weight values win. The nodes in the map network are arranged in a two-dimensional sheet with collateral connections (not shown) corresponding to the distances between nodes in this two-dimensional sheet. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 23 9.5.2 Simplifying winner-take-all description The response of the network A Gaussian firing rate around the node that receives the strongest input Wining node and label it with ‘*’ (( i i ) ( j j ) ) / 2 (9.27) The firing rate of other nodes, rij e Hebbian learning rule, wxini krx (riin wxini ) (9.28) * 2 * 2 2 w inx* The weight vector of the wining node is closest to the corresponding input vector, w inx w inx for all x (9.29) * Fig. 9.12 Experiment with two-dimensional self-organizing feature maps. (A) Initial map with random weight values. (B) and (C) Two examples of the resulting feature map after 1000 random training examples with different random initial conditions. These simulations are discussed further in Chapter 12. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 24 9.5.3 Other competitive networks (1) Fig. 9.13 Another example of a twodimensional self-organizing feature map. In this example we trained the network on 1000 random training examples from the lower left quadrants. The new training examples were then chosen randomly from the lower-left and upper-right quadrant. The parameter t specifies how many training examples have been presented to the network. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 25 9.5.3 Other competitive networks (2) Fig. 9. 14 Example of categorization (vector quantization) of two-dimensional input data (two-dimensional training vectors). The training data are represented as dots, and the input vector that would best evoke a response of one of the three output nodes is represented by a cross. (A) Before training there is no correspondence between the group of input data and the output node representing category. (B) After training we have a ‘preferred vector’ for each node that corresponds to each of the clusters in the training data set. (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 26 Conclusion The continuous attractor model Spatial representation Winner-take-all models Hebbian learning with continuous pattern Gaussian interaction Self-organization models Attractor regimes Path integration and Sequence learning Competitive networks (C) 2012 SNU CSE Biointelligence Lab, http://bi.snu.ac.kr 27