Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Deoxyribozyme wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Western blot wikipedia , lookup
Proteolysis wikipedia , lookup
Two-hybrid screening wikipedia , lookup
Genetic engineering wikipedia , lookup
Transcriptional regulation wikipedia , lookup
Cyanobacteria wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Signal transduction wikipedia , lookup
Biosynthesis wikipedia , lookup
Biochemistry wikipedia , lookup
Transformation (genetics) wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Microbial metabolism wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
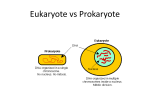
Microbiology Ch 3 18-37 Biology of Infectious Agents Prokaryotic and Eukaryotic Pathogens – bacteria are prokaryotes, whereas fungi, protozoa, and worms are eukaryotes -prokaryotes are haploid and do not carry out endocytosis and are incapable of ingesting liquid droplets -most eukaryotes are diploid with many chromosomes; Archaea are distinct from bacteria, but are also single celled and lacking nuclei and internal organelles -prokaryotes lack a nuclear membrane which allow transcription and translation to be coupled to rapidly synthesize new mRNA chains: which cannot be linked directly in eukaryotes Problems of Living Freely – the demands made on microbes are nutrition, occupancy (need to remain in a particular habitat), and resistance (to damaging agents) Feast or Famine – microbes frequently run out of nutrients; bacteria can rapidly metabolize nutrients when they arrive, but they have adapted to the lack of nutrients during periods of starvation and are poised for action until nutrients become available -two themes emerged in evolution: efficiency and adaptability Colonization and Occupancy – survival in intestinal tract depends on ability to remain in a given place and avoid being swept away by liquid currents -bacteria adhere to teeth by elaborating adhesive polysaccharides formed into a dental plaque (biofilm); in the intestine, microbes that adhere to wall differ from microbes living in the lumen because they face different nutritional problems Resistance to Damaging Agents – microbes encounter chemical/physical agents that threaten them, and they have evolved mechanisms to cope with those challenges -microbes have structural devices that protect against membrane-damaging chemicals, heat, and DNAdamaging radiation Small Size and Metabolic Efficiency – large numbers of bacteria can be packed in small volumes and is in the order of 1um in diameter -every human carries 10-100 trillion bacteria in the large intestine (more than eukaryotic cells) -being small allows microbes to have high metabolic rates because surface to volume ratio increases as size of cells decreases; rate of biochemical reactions limited by diffusion, and since bacteria are in intimate contact with external nutrients, they can achieve high metabolic rates -some bacteria double every 20 minutes -one measure of rapidity is that small metabolites (amino acids, sugars) constitute only 1% of bacteria’s dry weight Complex Envelopes and Appendages – bacteria are surrounded by layers and appendages that differ in composition from species to species -bacteria possess a cytoplasmic membrane, a cell wall, and some have outer membrane, flagella, pili, and a capsule -these structures account for 10-20% of dry weight of the cell -bacteria need these layers because they are constantly under stress -E. coli are exposed to bile salts that would degrade cell membrane if not protected -strongest antibody response to bacteria is on surface antigen Protection of Cytoplasmic Membrane – bacteria use 3 ways to protect cytoplasmic membrane from the environment (low osmotic pressure or presence of detergents) -these are gram-positive, gram-negative, and acid-fast bacteria -gram staining depends on ability of bacteria (gram-positives) to retain a complex of purple dye and iodine when challenged with alcohol wash -gram negatives do not retain the dye and can be counterstained after alcohol dye with a red dye, safranin -Procedure for Gram Staining – Fix specimen to slide add crystal violet (wash with H2O) add iodine fixative (wash with H2O) decolorize with lipophilic solvent (ethanol) counterstain with safrinin (pink stain) Gram-Positive Solution – protects cytoplasmic membrane with a thick cell wall composed of unique sugars and amino acids called murein, or peptidoglycan -Murein is critical component in maintaining shape and rigidity of both gram-positives and gramnegatives, but plays a LARGER role in gram-positives -Murein is composed of glycan chains that are cross-linked to one another by peptides, which wraps around the length and width of bacterium to form a sac -Shape of murein sac makes cell look different: rods (bacilli), spheres (cocci), or helices (spirilla) -Rigid murein corset allows bacteria to survive in media of lesser osmotic pressure than that of their cytoplasm -If you break apart murein with lysozyme, it will lyse the cell; lysozyme treated bacteria in isoosmotic medium do not lyse and become spherical spheroplasts -cell wall of gram-positive bacteria is composed of many layers of murein, which impede passage of hydrophobic compounds -sugars/charged amino acids make this structure hydrophilic, making cell resistant to the hydrophobic compounds of the intestine like bile salts -Gram-positive walls also contain teichoic acids, which are chains of tibitol/glycerol linked by phosphodiester bonds as well as lipoteichoic acids (signature molecule for gram-positive) -crystal violet can intercalate into cell wall and stain purple Gram-Negative Solution – these bacteria build a different protection mechanism than gram-positive bacteria; containing an outer membrane outside the murein cell wall, which is different from usual membranes and can resist damaging chemicals -the outer membrane is bilayered, with the outer leaflet containing lipopolysaccharide (LPS), which is not found anywhere else in nature. LPS has 3 components: 1. Lipid A – anchors LPS in outer leaflet, composed of disaccharides with fatty acids and phosphate groups attached 2. Core – consists of short sugars: ketodeoxyoctanoic acid and heptose 3. O antigen – long carb chain up to 40 sugars; hydrophilic chains cover bacterial surface and exclude hydrophobic compounds; mutants become sensitive to bile salts -exclusion of hydrophobic compounds in gram-negative bacteria relies on surrounding cells with hydrophilic polysaccharides -classic Gram-Negative Antigens: -K Antigen – Type of Capsule -H Antigen – Type of Flagella -O Antigen – long carb chain in LPS -outer membrane of gram-negatives also disallows hydrophilic substances due to lipid nature -gram-negative bacteria still need nutrients, and they obtain it through channels that permit passive diffusion of hydrophilic compounds (sugars, AA, ions) -channels consist of proteins with holes, called porins, which allow small hydrophilic molecules to pass through the porins -some hydrophilic molecules are too big to pass through porins, such as vitamin B12, sugars larger than trisaccharides, and Fe in the form of chelates; all of these cross by unique permeation mechanism that uses special proteins to translocate the compound -dual membrane creates a compartment called periplasmic space (periplasm) on outside of the inner membrane which contains the murein layer and a gel-like solution to facilitate nutrition -periplasm has degradative enzymes like phosphatases, nucleases, proteases, which break down large and impermeable molecules to digestible size -periplasm also contains binding proteins to help soak sugars and amino acids from medium -it also contains enzymes to inactivate antibiotics such as penicillins and cephalosporins, the βlactamases -periplasm rarely exists in gram-positive bacteria, but would be between the peptidoglycan and cytoplasmic membrane -bacteriophages sometimes use outer membrane as attachment sites, but at the same time, the outer membrane protects against many antibiotics -gram-negative bacteria are more resistant to antibiotics, especially penicillin -LPS of gram-negative membrane is reactive in the host -Lipid A component elicits fever and activates immunological and biochemical events that lead to mobilization of host defense mechanisms -in large quantities, LPS is called endotoxin, and can cause shock and death -the O-antigen portion is highly antigenic and come in many varieties Acid-Fast Solution – a few bacterial types, especially tubercle bacillus have developed a 3rd way to confront environmental challenges -cell walls of these bacteria contain large amounts of waxes, complex long-chain hydrocarbons with sugars and other modifying groups -these organisms are impervious to many harsh chemicals, including acids -dye introduced to these cells cannot be removed with HCl, unlike with other bacteria -the waxy coat is interlaced with murein, polysaccharides and lipids -coat allows the bacteria to avoid being killed by white blood cells -cost of protection is that these organisms grow very slowly (once every 24 hours) -mycobacteria are neither gram-positive or gram-negative, and instead have waxy coat; need to do an acid-fast stain to visualize them -add lipophilic dye to intercalate into cell coat and doesn’t allow for easy extraction of dye -TWO BACTERIA THAT DO NOT GRAM STAIN ARE Rickettsia and Chlamydia, they have to live INSIDE the cell, and are referred to as energy-parasites because they live off of energy in eukaryotic cell -spirochetes do NOT gram-stain; gram-negative like organism with a flagella TRAPPED in the periplasmic space, and it spirals because flagella tries to beat -mycoplasma (different from mycobacteria) lacks a cell wall and incorporates sterols into membrane Murein and Antibiotics that Inhibit its Synthesis – drugs that block formation of murein lead to lysis and death of susceptible bacteria -the penicillins, cephalosporins, and carbapenems (collectively, β-lactams) act by inhibiting murein synthesis -murein is synthesized from nucleotide-bound building blocks: uridine diphosphate + either Nacetylglucosamine (GlcNAc) or N-acetylmuramic acid (has a peptide chain attached to it) -monomeric subunits made in cytoplasm and transferred from uridine disphosphate to a lipid carrier in the membrane; disaccharides are then linked to growing murein chain in periplasm -this step is inhibited by vancomycin -regeneration of lipid carrier is inhibited by bacitracin -final reaction in murein synthesis is transpeptidation, where long chains of disaccharides are crosslinked to make a 2D network, requiring formation of peptide bond between D-alanine on one chain and free N terminus of lysine or a diaminopimelic acid on the other -link is formed with the subterminal D-alanine, the terminal D-alanine being cleaved away in the process -reaction is exchange of one peptide bond (between two alanines) with another – a true transpeptidation -penicillins and cephalosporins inhibit this reaction -in presence of penicillin, transpeptidase becomes confused and makes a lethal penicilloylenzyme complex instead of an intermediate D-alanine-enzyme complex -antibiotics that inhibit murein synthesis kill bacteria by lysing them; in contrast to lysozyme, these antibiotics do not affect the murein itself, only its synthesis -without murein, cell continues to make cell contents to increase mass; the enlarged cytoplasm is not restrained by proper murein, and the cell contents extrude and the cell lyses -cells that are NOT growing are NOT lysed by penicillin because they are not increasing their mass -penicillin isn’t a perfect drug, there are always cells in a colony that do not grow (called persisters) and are not lysed with penicillin -for certain types of bacteria, penicillin is bacteriostatic and bactericidal; these bacteria are called tolerant, due to a deficiency in autolysin, a bacterial enzyme that cleaves murein -this enzyme breaks open bonds of murein at the septum to permit separation of daughter cells during division; normally autolysin is tightly controlled; treatment with penicillin may arouse it to more-unrestrained action -pneumococci are normally susceptible to lysis, but autolysin-defective mutants are found among penicillin-resistant mutants; strong detergents do NOT lyse these mutants -mycoplasmas are a group of bacteria without murein and are not rigid/have no shape and they are resistant to penicillin -Mycoplasma pneumonia contain sterols in their membranes Cytoplasmic Membrane – most critical role is uptake of substrates from the medium; bacteria take up mainly small-molecular-weight compounds and rarely macromolecules -compounds are hydrolyzed by enzymes in the periplasm or surrounding medium to peptides, oligosaccharides, nucleosides, and phosphate -cytoplasmic membrane contains permeases which facilitate entry of most metabolites; usually facilitates equilibration of a compound inside and outside the cell; uses energy Transport Across Cytoplasmic Membrane – 1. Facilitated Diffusion – glycerol can be carried across membrane down a concentration gradient, does not concentrate molecules inside the cell relative to outside environment; uptake is driven by intracellular use of the compound 2. Group Translocation – also known as phosphorylation-linked transport, this energy-dependent mechanism is used to transport certain sugars; substances are normally chemically altered in the process (sugar binds carrier in membrane, metabolized and internalized) 3. Active Transport – energy is used to drive the accumulation of substrate a. Lactose is concentrated unchanged in the cell, making transport of additional molecules unfavorable b. To drive the transport of molecules, cell uses energy from electrochemical gradient of protons, the proton motive force i. Gradient is generated by extrusion of protons from cell (resulting from oxidation of metabolic intermediates like NADH or ATP hydrolysis ii. Lactose is accumulated intracellularly by coupling the energetically unfavorable transport with the energetically favorable reentry of proteins (symport) -some proteins aid the process by modifying or concentrating substrates in the periplasmic space of gram-negatives; these binding proteins are specific for sugars, nucleotides, etc.. -periplasmic space also contains nucleotidases, nucleases, peptidases, proteases, and other hydrolytic enzymes -actual transport is carried out by membrane-bound carriers called permeases, which are involved in all three types of transport -permeases have high affinity for substrate on the outside and low affinity on the inside Uptake of Iron – free iron is scarce in the blood and many tissues because it is bound to transferrin or ceruloplasmin; iron is essential for the growth of bacteria, and they have adapted to obtain iron -they excrete chelating compounds called siderophores that bind Fe with great avidity; some can directly extract iron from transferrin Other Functions of Bacterial Membrane – cytoplasmic membrane is where cytochromes are located and oxidative metabolism is carried out, performing the role of mitochondria in eukaryotic cells -membrane is also there for excretion of nascent proteins or membrane-incorporation proteins -some bacteria secrete 10% of all the proteins they make, including toxins and virulence factors -nascent polypeptides that have hydrophobic signal sequences are translocated from ribosomes across cytoplasmic membrane by an energy-requiring mechanism -bacteria can take up long DNA molecules DNA and Chromosome Mechanics – gemone is a single, circular chromosome of double stranded DNA; some species have a single chromosome; E. coli chromosome has 5 million base pairs and 2500 genes -DNA is coiled into a central, irregular structure called a nucleoid, where it is twisted into supercoils needed for organization, transcription, and replication -supercoiling depends on two topoisomerases; DNA gyrase (introduces supercoils into circular DNA) and Topoisomerase I (relaxes the supercoils by making single strand nicks) -DNA replication has 3 stages: initiation, elongation, and termination -DNA replication starts at a precise place called the replicative origin and proceeds in both directions -the two polymerases meet halfway around the chromosome -to replicate, E. coli must unwind and rotate at 6000rpm -at any given temp, rate of DNA polymerase movement is independent of growth rate of cells -in E. coli, replication takes 40 minutes whether cell is dividing slowly or quickly -in fast growing bacteria, frequency of initiation is increased, so there can be multiple origins that begin synthesizing DNA at the same time -most drugs targeting DNA replication are too toxic for clinical use however metronidazole is a drug normally inert but can become active through modification by some bacteria (molecule contains a nitro group which must be partially reduced -partially reduced metronidazole is incorporated into DNA, making it unstable (lethal synthesis) -metronidazole is a drug that is effective against anaerobic bacteria and amoebas -other DNA inhibitors are nalidixic acid (inhibitor of DNA gyrase) and fluoroquinolones (interfere with DNA gyrase or topoisomerase and cause double stranded DNA breaks) Gene Expression: Uniqueness of Prokaryotic RNA Polymerase and Ribosomes – bacterial cytoplasm is composed largely of proteins (40% dry weight) and RNA (35% dry weight) -bacterial ribosomes have smaller subunits and RNA molecules (70S 50S and 30S) than eukaryotic (80S), and there are 2 subunits of the ribosomes that have 21 and 35 different proteins -protein synthesis is the chief biosynthetic activity of bacteria -RNA is made at a rate proportional to the number of RNA polymerase molecules engaged in transcription, and protein synthesis rate is proportional to the cellular concentration of ribosomes -synthesis of principal macromolecules of bacteria is regulated by the frequency with which each chain is initiated and not by altering rate of manufacture of each molecule (speed of chain elongation) -antibiotics can act selectively at initiation or elongation of macromolecular synthesis -rifampin – an inhibitor of transcription, acts at the initiation step by binding to RNA polymerase freely floating in cytoplasm but much less strongly to RNA polymerase bound to DNA that has initiated transcription -mammalian RNA polymerase does not bind rifampin, so the drug is non-toxic -largest class of clinically useful antibiotics consists of those that inhibit protein synthesis -some work by inhibiting ribosomes (by binding to either large or small subunits); because prokaryotic ribosomes are different from eukaryotic ribosomes -ribosomally active antibiotics are chloramphenicol and macrolides (erythromycin), which block formation of peptide bonds by binding near aminoacyl tRNA binding site on large ribosomal subunit -then antibiotic is removed, free ribosomes can start translation again; these drugs are more bacteriostatic than bactericidal -a group of protein synthesis inhibitors, the aminoglycosides, is bactericidal -aminoglycosides include streptomycin, gentamicin, and neomycin which all bind the 30S subunit of bacteria; it enhances binding of 30S and 50S and inhibits elongation of peptides -action of this group of antibiotics accumulates free ribosomes as 70S subunits and not as 30S and 50S subunits; accumulation of 70S results in abortive attempts to initiate protein synthesis, but the atypical ribosomes block normal ribosomes from translating -effect is irreversible, and causes death of bacteria -point mutation in 30S causes resistance to these antibiotics Capsules, Flagella, and Pili – these structures are dispensable; important for survival under certain circumstances but not in others -the capsule is a slimy outer coating made by bacteria; it is not needed for survival; consist of high molecular weight polysaccharides that make bacteria slippery and ready to avoid white blood cells -flagella are long, helical structures that endow bacteria with motility to aid in their spread -can have one or many flagella -pili are also called fimbriae are involved in attachment of bacteria to cells and other surfaces Bacterial Chemotaxis – movement caused by flagella is used for chemotaxis toward substances that attract and away from substances that attract -flagella spin around from their point of attachment at cell surface -each flagellum has a counterclockwise helical pitch -when flagella rotate clockwise, bacteria get in the way of each other and cannot form bundles; thus, bacteria tumble in a random fashion -both swimming and tumbling account for bacterial chemotaxis -in the absence of attractants or repellents, bacteria alternate indifferently between swimming and tumbling; when attractant is sensed, swimming lasts longer than tumbling; swimming stops quickly when repellent is present Bacterial Adhesion and Pili – microbes are attracted to specific tissues, called tissue tropism, which involves attachment of surface components of organisms to specific receptors on certain cells -bacterial structures most often involved are pili, which are shorter than flagella and are distributed in large numbers over the surface -bacteria that can conjugate have specialized sex pili; longer structures linking donor (male) and recipient (female) cells during DNA transfer -attachment to eukaryotic cells involves attachment of the tip of the pili followed by their retraction into the cell (resembling grappling hooks); drugs are used to target pili -Some organisms can change AA composition of pili and put on disguises that enable them to outflank the immune system, called antigenic variation -Salmonella undergoes rapid changes between expression and nonexpression of genes that code for the protein of flagella, called phase variation Nutrition and Energy Metabolism – nutrition for bacteria must be provided by medium or synthesized by the organisms themselves; based on energy requirements, bacteria are divided into photosynthetic or chemosynthetic bacteria that subsist on CO2 and minerals, using light for energy. -the other type of bacteria include all the organisms that need preformed organic compounds; all pathogenic microbes fall within this group. E. coli only need glucose -other pathogenic bacteria are unable to make one or more essential metabolites- vitamins, AA, purines, pyrimidines -strict aerobes require oxygen to grow, such as tubercle bacillus which thrives in the lung -strict or obligate anaerobes cannot grow in the presence of oxygen, such as those that cause botulism -most important to medicine are those that can grow whether in O2 or not, the facultative anaerobes and include E. coli and other intestinal bacteria -strict aerobes perform respiration only, where O2 is the final electron acceptor in a set of oxidationreduction reactions -strict anaerobes carry out fermentation where the final electron acceptor is an organic molecule, such as pyruvate, which is reduced to lactate in the lactic acid fermentation, and acetyl CoA ethanol -facultative anaerobes are capable of either form of metabolism -respiration yields more energy than fermentation -E. coli can use glucose, lactose, fructose or AA for nutrition -Pseudomonas can grow on hundreds of organic compounds -it is possible to culture bacteria in minimal media containing water solutions of glucose, ammonia, phosphate, sulfate, and other minerals -bacteria can also be grown on nutrient broth containing meat extract and other complex proteins; adding agar to these creates solid media -some bacteria can only grow on complex media and have complex nutritional requirements, called nutritionally fastidious, such as highly parasitic species such as staphylococci or streptococci -some bacteria ONLY replicate INSIDE host cells and have never been cultured, such as chlamydia, they are known as obligate intracellular parasites Growing and Resting States – the time it takes for bacteria to divide is called generation time or doubling time; E. coli takes 20 minutes to double in nutrient broth and 1 hour in glucose-minimal media -growth will stop once population reaches maximal cell density or nutrients become exhausted Bacteria Growth Measurement – many ways have been devised to measure growth of bacteria; analyzers that measure numbers include hemocytometer and electronic particle analyzers, which measure the total count, and cannot discriminate between living and dead bacteria -to determine the number of living or viable bacteria, a colony count is used, which is carried out by placing an appropriate dilution on solid growth medium and waiting for colonies to form -colonies multiplied by the dilution factor is number of colony-forming units (CFU) -if bacteria grow in clumps, number of CFUs is an underestimate of total number of bacteria Law of Growth – Nt = N0ekt where Nt is number of bacteria at time t, and N0 is the initial number of bacteria at t=0, k is the growth rate constant -law of growth describes a geometric progression; death can be described by the same equation but with a negative constant Growth in the Real World – single bacterium diiding twice an hour would produce mass as large as earth in 2 days; when bacteria grow to a certain density, they exhaust required nutrients or run out of a carbon source, required inorganic compound or essential amino acids or vitamins -for aerobic bacteria, crowding leads to exhaustion of oxygen, which is poorly soluble in water -stage of culture where growth stops is called the stationary phase -explosiveness of exponential growth means that a small number of bacteria can initiate infection fast -even when E. coli exhaust source of glucose, they continue to carry out low level of protein synthesis sufficient to adapt to the use of other nutrients such as other sugars -bacteria are exposed to countless injuries and have developed special adaptive mechanisms to cope -damage to E. coli DNA activates genes that code for proteins to repair the damage, known as the SOS response -non-growing bacteria are still immunogenic and can elicit immune responses with both beneficial and detrimental results -production of toxins in non-growing bacteria often starts or accelerates when bacteria enter stationary phase, because sometimes the toxins provide bacteria with nutrients -cessation of growth of some bacterial species initiates sporulation resulting in production of metabolically inert spores resistant to chemical and physical insults where mother cell lyses -cytoplasmic contents then released contain large amounts of toxins, such as in tetanus, gas gangrene, and other diseases Mechanisms of Adaptation – inefficient bacterial strains are rapidly lost in competition with others that use their resources more effectively -metabolic efficiency is characterized by metabolic parsimony: bacteria tend not to make constituents they cannot use at the time Regulation of Enzyme Activity – stopping the flow of substrates, like in E. coli synthesis of leucine when grown on leucine medium, at very beginning of the pathway eliminates wastes of unusable metabolites; -first enzyme is affected due to feedback or end-product inhibition Regulation of Enzyme Synthesis – feedback inhibition stops synthesis of leucine in the leucine fed culture, but enzymes for synthesis of leucine are still synthesized -genes involved in metabolic pathway are often strung together in a multigenic segment of DNA called an operon, and transcription of all genes of an operon into mRNA can be turned on or off together by a single regulatory switch -one sequence is the promoter site where RNA polymerase binds to initiate synthesis of mRNA -operons involved in biosynthesis of amino acids, such as leucine are sometimes regulated by a mechanism called attenuation -small stretch of mRNA is synthesized from beginning of coding sequence of leucine operon, regardless of presence or absence of leucine. RNA polymerase encounters a region called the attenuator, where in the presence of leucine, transcription is terminated -this achieves the effect of not synthesizing enzymes of pathway if not needed -another mechanism for turning on or off the synthesis of enzymes is found in the case of enzymes involved in utilization of sugars -if lactose is the sole carbon source, bacteria must make the enzyme B-galactosidase necessary to convert lactose into glucose and galactose -at the beginning of the operon, past the promoter, there is a sequence called the operator where a protein called a repressor binds, inhibiting initiation of transcription -in the presence of lactose, repressor undergoes conformational change and stops binding to the operator and transcription can begin again -B-galactosidase is an inducible enzyme, made on demand; in contrast to a constitutive enzyme -for B-galactosidase, lactose is known as the inducer