Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
History of genetic engineering wikipedia , lookup
Quantitative trait locus wikipedia , lookup
Genomic library wikipedia , lookup
Pathogenomics wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Gene expression profiling wikipedia , lookup
Genome editing wikipedia , lookup
Microevolution wikipedia , lookup
Minimal genome wikipedia , lookup
Genomic imprinting wikipedia , lookup
Epigenetics of human development wikipedia , lookup
Public health genomics wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Gene expression programming wikipedia , lookup
Designer baby wikipedia , lookup
Genome (book) wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
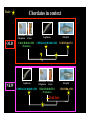
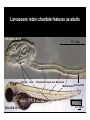
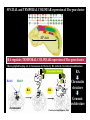
La rv ac ea ns a di As ci s ns hi p Am u ox y e pr k r ha m La S Ze br is af h f ol W m u H an Limbs Vertebrates Jaws Paired appendages Cephalochordates Urochordates Stem chordates Big brain Neural crest Placodes Dorsal nerve cord Notochord Gill slits Large-scale Gene Duplications “What characterizes the living world is both its diversity and its underlying unity” PARADOX: “Phenotypic diversity versus genetic unity” http://drujohnwigsinkiwi.files.wordpress.com/2007/01/fly2.jpg http://en.wikipedia.org/wiki/Image:Mus_Musculus-huismuis.jpg http://www.mun.ca/biology/desmid/brian/BIOL3530/DB_Ch15/figB4_A.jpg Evolutionary Genomics allows us to test that paradox • We have access to all genes present in the genome of an organism • We also can also detect those genes that are missing INVERSE PARADOX: “Genetic diversity versus phenotypic unity” Chordates in context Urochordates Olfactores Trait: Chordates in context Larvaceans Ascidians Oikopleura Ciona OLD UROCHORDATES (Tunicates) Amphioxus Zebrafish CEPHALOCHORDATES VERTEBRATES ? NEW Amphioxus CEPHALOCHORDATES Larvaceans Ascidians Oikopleura Ciona UROCHORDATES (Tunicates) Trait loss Zebrafish VERTEBRATES Larvaceans retain chordate features as adults Oikopleura larva 0.1 mm Gill slits Anus Dorsal hollow nerve cord Notochord Muscle blocksPost-anal Tail Zebrafish larva 0.1 mm absence of Midbrain in urochordates Zebrafish Oikopleura dioica Ciona intestinalis FOREBRAIN Oikopleura larva HINDBRAIN VERTEBRATES UROCHORDATES (Cañestro, Bassham and Postlethwait, 2005 Dev Biol) FOREBRAIN HINDBRAIN SPINAL CORD FOREBRAIN MIDBRAIN HINDBRAIN Zebrafish larva SPINAL CORD Midbrain, vertebrate innovation or urochordate simplification? Larvaceans Ascidians Oikopleura Ciona Amphioxus CEPHALOCHORDATES UROCHORDATES (Tunicates) Zebrafish VERTEBRATES Trait loss? ? Trait: presence of midbrain Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. (Cañestro and Postlethwait, 2007 Dev Biol) Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. Retinoic acid (RA) •RA is a derivative of vitamin A •RA basic role is to inhibit cell proliferation, and to activate mechanisms of cell differentiation and apoptosis. •In humans, as in other vertebrates, RA plays important physiological roles: Early embryogenesis (axial patterning), the formation of limbs, eyes Respiratory system, nervous system, reproductive system, hematopoietic system •Alterations in the RA metabolism or the pathways of RA action can lead to pathologic conditions such as cancer, dermatologic diseases, osteoporosis, rheumatoid arthritis, and premature births. •The broad range of RA functions has led to RA being targeted to develop applications, ranging from the use of RA anti-proliferative properties in therapeutic treatments against cancer, to the use of antiaging properties of RA in the industry of the skin care. •Therefore, the study of the synthesis of RA and the mechanisms of RA action is important not only to better understand the physiological roles of the RA and develop new applications, but also to prevent these applications from becoming toxic by altering the physiological levels of RA and posing a risk for the human health. Action of Retinoic Acid Genetic Machinery Vitamin A (Retinol) (Raldh) RARE “Collinearity” of Hox-cluster gene expression: •spatial •temporal http://www.mun.ca/biology/desmid/brian/BIOL3530/DB_Ch15/figB4_A.jpg SPATIAL and TEMPORAL COLINEAR expression of Hox gene cluster AP axis RA regulates TEMPORAL COLINEAR expression of Hox gene cluster Choreographed looping out of chromosome territories by RA-induced chromatin decondensation. Transcription factors Hoxb1 Hoxb9 RA chromosome RA (Chambeyron and Bickmore, 2004) SPATIAL and TEMPORAL COLINEAR expression of Hox gene cluster AP axis RA regulates TEMPORAL COLINEAR expression of Hox gene cluster Choreographed looping out of chromosome territories by RA-induced chromatin decondensation. 1 Hoxb1 Transcription factors Hoxb9 RA chromosome RA (Chambeyron (Chambeyron and and Bickmore, Bickmore, 2004) 2004) SPATIAL and TEMPORAL COLINEAR expression of Hox gene cluster AP axis RA regulates TEMPORAL COLINEAR expression of Hox gene cluster Choreographed looping out of chromosome territories by RA-induced chromatin decondensation. 1 Hoxb1 1 Hoxb9 RA RA Transcription factors 2 RA Chromatin structure Genomic architecture chromosome (Chambeyron and Bickmore, 2004) RETINOIC RETINOIC ACID ACID crucial for AnteroPosterior CNS patterning in Chordates RA-induced Homeotic transformation (POSTERIORIZATION) Midbrain Hindbrain Forebrain Hindbrain Fore/Midbrain RA RA RA (modified from Schilling and Knight, 2001) CEPHALOCHORDATES UROCHORDATES (Ascidians) VERTEBRATES “Innovation” of RA-genetic machinery was a milestone for the origin of chordate bodyreviewed plan.in Fujiwara (2006) Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. RA-genetic machinery ALDH1A CYP26 RAR ? ? ? ? ? ? AMBULACRARIA ? ? ? ? ? CHORDATES Echinoderms Hemichordates Cephalochordates Urochordates AscidiansLarvaceans Vertebrates 1. RA-genetic machinery is NOT a chordate innovation ALDH1A CYP26 RAR AMBULACRARIA CHORDATES Echinoderms Hemichordates Cephalochordates Urochordates Vertebrates AscidiansLarvaceans + (Cañestro, Postlethwait, Gonzalez-Duarte, Albalat 2006 Evo Dev) 2. RA genetic machinery have been lost in Oikopleura ALDH1A CYP26 RAR AMBULACRARIA CHORDATES Echinoderms Hemichordates Cephalochordates Urochordates Vertebrates AscidiansLarvaceans ALDH2 prediction: ALDH1A1/2/3 AP axial patterning in Oikopleura is independent of RA “ALDH2” (Cañestro, Postlethwait, Gonzalez-Duarte, Albalat 2006 Evo Dev) Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. Endogenous β-galactosidase activity Amphioxus (Cañestro et al., 2001) Oikopleura DMSO * Figure 3 DMSO * Pharynx Posterior gut 10-6 M RA Pharynx Homeotic posteriorization 10-6 M RA RA does NOT cause homeotic posteriorization in Oikopleura Hox1 Pax2/5/8a RA DMSO Otx Figure 4 Figure 5 Figure 6 AP axial patterning in Oikopleura is independent of RA Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. • “genetic diversity”: In contrast to Oikopleura, Ascidians have a full RA-genetic machinery •“phenotypic unity”: Oikopleura and ascidians share a basic AP axial patterning Hox1 CNS Ciona Hox1 Epidermis (Nagatomo & Fujiwara, 2003) CNS Oikopleura Epidermis (Cañestro et al., 2005) Evo-Devo “inverse paradox”: “similar morphology” despite “different genetic machinery” (Cañestro, Yokoi and Postlethwait, 2007 Nature Reviews Genetics) Ascidians palp cells HrETR-1 DMSO DMSO RA HcPax2/5/8 RA DMSO RA HrHox1 RA posteriorization RA DMSO (Katsuyama et al., 1995) (Katsuyama et al., 1995, 1998) “Shortened head” or “Moonfaced” (Yagui and Makabe, 2002) RA affects palp morphogenesis (Hinman and Degnan, 2000) NO posteriorization In agreement with our results in Oikopleura, RA does NOT seem to AP axial patterning became independent of RA in stem urochordate induce homeotic posteriorization in ascidians. Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. Abstract Developmental signaling by retinoic acid (RA) is thought to be an innovation essential for the origin of the chordate body plan. The larvacean urochordate Oikopleura dioica maintains a chordate body plan throughout life, and yet its genome appears to lack genes for RA synthesis, degradation, and reception. This suggests the hypothesis that the RA-machinery was lost during larvacean evolution, and predicts that Oikopleura development has become independent of RA-signaling. This prediction raises the problem that the anterior–posterior organization of a chordate body plan can be developed without the classical morphogenetic role of RA. To address this problem, we performed pharmacological treatments and analyses of developmental molecular markers to investigate whether RA acts in anterior–posterior axial patterning in Oikopleura embryos. Results revealed that RA does not cause homeotic posteriorization in Oikopleura as it does in vertebrates and cephalochordates, and showed that a chordate can develop the phylotypic body plan in the absence of the classical morphogenetic role of RA. A comparison of Oikopleura and ascidian evidence suggests that the lack of RA-induced homeotic posteriorization is a shared derived feature of urochordates. We discuss possible relationships of altered roles of RA in urochordate development to genomic events, such as rupture of the Hox-cluster, in the context of a new understanding of chordate phylogeny. Conclusions Trait: AP axial patterning is dependent of RA “Genetic variability” vs “Phenotypic unity” Cephalochordates Urochordates Ascidians Vertebrates Larvaceans Loss of RA-machinery Trait loss: AP axial patterning independent of RA Conclusions (Ikuta et al. 2004) 1 2 3 4 5 6 7 8 9 1011121314 Cephalochordates 1 2-4 10 6-5 1213 (Seo et al. 2004) 1 2 4 9 9’10111213 Urochordates Ascidians 1 2 3 4 5 6 7 8 9 10111213 A B C D Hox gene organization Vertebrates Larvaceans Loss of RA-machinery 1.AP axial patterning independent of RA 2.Hox-cluster rupture & Loss of temporal collinearity Conclusions (Ikuta et al. 2004) 1 2 3 4 5 6 7 8 9 1011121314 Cephalochordates 1 2-4 10 6-5 1 2 4 9 9’10111213 Urochordates Ascidians Oikopleura no temporal colinearity 1213 (Seo et al. 2004) 1 2 3 4 5 6 7 8 9 10111213 A B C D Hox gene organization Vertebrates Larvaceans Loss of RA-machinery 1.AP axial patterning independent of RA 2.Hox-cluster rupture & Loss of temporal collinearity Conclusions 1 2 3 4 5 6 7 8 9 1011121314 Cephalochordates 1 2-4 10 6-5 1213 1 2 4 9 9’10111213 Urochordates Ascidians 1 2 3 4 5 6 7 8 9 10111213 A B C D Vertebrates Larvaceans Loss of RA-machinery 1.AP axial patterning independent of RA 2.Hox-cluster rupture & Loss of temporal collinearity 3.Genome diminution (e.g. Gbx loss) Urochordate Determinative development/ rapid embryogenesis Conclusions 1 2 3 4 5 6 7 8 9 1011121314 Cephalochordates 1 2-4 10 6-5 1213 1 2 4 9 9’10111213 Urochordates Ascidians 1 2 3 4 5 6 7 8 9 10111213 A B C D Vertebrates Larvaceans Loss of RA-machinery 1.AP axial patterning independent of RA 2.Hox-cluster rupture & Loss of temporal collinearity 3.Genome diminution (e.g. Gbx loss) So ... now, what do you think? Is the midbrain a vertebrate innovation? or maybe a urochordate simplification?