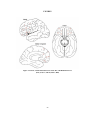
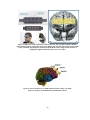
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Executive functions wikipedia , lookup
Embodied cognitive science wikipedia , lookup
History of neuroimaging wikipedia , lookup
Cognitive neuroscience wikipedia , lookup
Affective neuroscience wikipedia , lookup
Aging brain wikipedia , lookup
Neurolinguistics wikipedia , lookup
Neuroeconomics wikipedia , lookup
Neuroesthetics wikipedia , lookup
Lateralization of brain function wikipedia , lookup
Neurophilosophy wikipedia , lookup
Speech synthesis wikipedia , lookup
Speech perception wikipedia , lookup
Neurocomputational speech processing wikipedia , lookup
Embodied language processing wikipedia , lookup
University of Vermont ScholarWorks @ UVM Graduate College Dissertations and Theses Dissertations and Theses 2015 FNIRS Measures of Prefrontal Cortex Lateralization During Stuttered and FluencyEnhanced Speech in Adults Who Stutter Danra M. Kazenski University of Vermont, [email protected] Follow this and additional works at: http://scholarworks.uvm.edu/graddis Part of the Communication Commons, Communication Sciences and Disorders Commons, and the Psychology Commons Recommended Citation Kazenski, Danra M., "FNIRS Measures of Prefrontal Cortex Lateralization During Stuttered and Fluency-Enhanced Speech in Adults Who Stutter" (2015). Graduate College Dissertations and Theses. Paper 303. This Dissertation is brought to you for free and open access by the Dissertations and Theses at ScholarWorks @ UVM. It has been accepted for inclusion in Graduate College Dissertations and Theses by an authorized administrator of ScholarWorks @ UVM. For more information, please contact [email protected]. FNIRS MEASURES OF PREFRONTAL CORTEX LATERALIZATION DURING STUTTERED AND FLUENCY-ENHANCED SPEECH IN ADULTS WHO STUTTER A Dissertation Presented by Danra M Kazenski to The Faculty of the Graduate College of The University of Vermont In Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy Specializing in Psychology January, 2015 Defense Date: October 8, 2014 Dissertation Examination Committee: Barry Guitar, Ph.D., Advisor Michael Cannizzaro, Ph.D., Chairperson William Falls, Ph.D. Jom Hammack, Ph.D. Alice Schermerhorn, Ph.D. Cynthia J. Forehand, Ph.D., Dean of the Graduate College ABSTRACT The present study compared lateralization of cortical activation patterns in the prefrontal cortex (PFC) of adults who stutter (AWS) and typical speakers (TS) as measured with functional near infrared spectroscopy (fNIRS) in habitual and fluencyenhanced speaking conditions. Participants were AWS (n = 11) and gender- and age-matched TS (n = 11) who completed speaking tasks in three condition blocks: (1) habitual speech using no speaking strategy (2) prolonged speech after receiving short-term training in fluency-shaping strategy-use (3) syllable-timed speech after being trained to speak in rhythm with a metronome at 92 beats per minute. The three primary dependent variables were mean change in HbO (oxygenation) relative to resting baseline in the right and left PFC hemispheres and a Laterality Index (L-R)/(L+R) calculated from these values. Two primary hypotheses were tested: (1) AWS will present with greater right-hemisphere PFC oxygenation relative to TS in a habitual or everyday speaking task (2) AWS will present with reduced right-hemisphere PFC activation (leftward shift in laterality more similar to TS) during fluency-enhanced speech strategy tasks relative to a habitual speech task. Real-time stuttered speech measures using fNIRS indicated greater effortfulness of speech production in AWS when speaking fluently and disfluently as measured by greater bilateral change in PFC HbO relative to TS. AWS laterality did not differ from TS during everyday conversation and did not significantly change when using fluencyenhancing strategies, which was counter to the hypotheses. The TS group presented with significantly greater leftward PFC HbO in the metronome condition compared to AWS. Prolonged speech and metronome-timed speech seem to be associated with different activation patterns in the PFC for AWS and for TS. Results suggest an alternative explanation for compensatory activation in AWS during speech production, such that AWS present with greater overall activation in both PFC hemispheres relative to TS which results in greater right-sided laterality than TS. Future long-term studies on adults receiving prolonged speech treatment and examination of similar measures in young children who stutter may reveal more about the compensatory versus causal nature of stuttering. DEDICATION This dissertation is dedicated in loving memory to Mildred Kazenski and Drina Vurbic (SoS). ii ACKNOWLEDGEMENTS I am immensely grateful for the constant stream of support and encouragement from those who helped me navigate through the enlightening and often mystifying dissertation process. I am especially thankful for my Advisor, Dr. Barry Guitar for his warm and caring approach as a mentor and guide through the world of stuttering and beyond. Barry’s wife, Carroll, A.K.A. Vermont mom, was also an essential grounding force in my life after moving back from Montreal, and I am eternally appreciative of their generosity as part of my Burlington family. Surviving this project and the Ph.D. process could not have been accomplished without the support of Dr. Edward Conture as our external advisor and grant-writing guru. My committee Chair, Dr. Michael Cannizzaro shared critical expertise about the new fNIRS technology as we tackled this totally novel approach together. Dr. Annie Murray-Close provided invaluable feedback on earlier drafts of the manuscript, and the other committee members Dr. William Falls, Dr. Jom Hammack and Dr. Alice Schermerhorn brought key insights from the psychological perspective which provided needed balance to this project. Cheers to Alan Howard who patiently helped me plow through the multiple levels of statistical analyses and to Lucy Greenberg who was a stellar all around research assistant. Thanks to Claudia Abbiati who stepped in as a trusty sub and helped with all videos. Jackson Mathews graciously recorded his voice as the narrator for the protocol slides, and Alexis St. Croix and Emma Feldman helped transcribe the speech samples. Giant thanks to the whole Communication Sciences & Disorders and Psychology iii departments as I was the “guinea pig” for the new crossover Ph.D. which required many adjustments to be a successful venture. Thank you to all the members of the stuttering community who participated in this project and who have shared personal experiences so honestly and openly as part of the support groups over the years. Your perspectives have shaped all of my interactions with kiddos and adults who stutter, and I thank you for your candor and constant bravery to step outside your comfort zones! Hugs to my wonderful friends, Lauren & “A”, Nancy & Jeremy, Gwen, Team Mediocre (Rob & Kathy), Amy & Steve, Lizzy, Jess L., Sarita, Amanda H., Mia & Adam (and sweet Merin and Tessa), Wendy C., Lisa D., Jasmine, the Dartmouth volleyball crew, and all of the staff at Kahnawake and the Montreal Fluency Centres. Salute to my fellow Carnimores, especially Don Calamari and Secretary of Steak, for providing a jovial social “meating” outlet during intense points in the process. Of course, heaps of thanks to my parents, Lois and Bob, and my brothers and sister-in-law, Keith, Robb and Greta. We had quite a bit of struggle and loss in the past four years, and it was only manageable thanks to your strength and love. Finally, to my loving other half, Hauke, your patience and calmness throughout multiple data analyses panic attacks was impressive to say the least. I look forward to being on the “other side” of this process with you and Sophie, and thank you for being my infallible best friend and counterpart. iv TABLE OF CONTENTS Page DEDICATION ............................................................................................................................... ii ACKNOWLEDGEMENTS …………………………………………………………………….iii ABBREVIATIONS ...................................................................................................................... vii LIST OF TABLES ...................................................................................................................... viii LIST OF FIGURES ....................................................................................................................... x CHAPTER 1: INTRODUCTION ................................................................................................. 1 1.1. Literature Review ................................................................................................................. 3 1.1.1. Laterality of Activation During Speech Production..................................................... 3 1.1.2. Fluency-enhancing Speech Strategies ......................................................................... 9 1.1.2.1 Metronome-timed Speech ..................................................................................... 9 1.1.2.2. Prolonged Speech (PS) ....................................................................................... 13 1.1.3. Prefrontal Cortex ...................................................................................................... 17 1.1.4. PFC Activation During Effortful Strategy-Application ............................................ 20 1.1.5. Neuroimaging Measures of Activation Change During Speech Production ............ 24 1.1.6. Real-time Activation Measures During Spontaneous Speech with Functional Near Infrared Spectroscopy (fNIRS) ................................................................................. 25 1.2. Purpose ............................................................................................................................... 29 1.3. Hypotheses ......................................................................................................................... 30 CHAPTER 2: METHODS .......................................................................................................... 31 2.1. Inclusion and Exclusion Criteria ........................................................................................ 31 2.2. fNIRS Recording ................................................................................................................ 32 2.3. Procedures .......................................................................................................................... 32 2.3.1. Stimulus Delivery........................................................................................................ 34 2.3.2. Speaking Blocks .......................................................................................................... 34 2.3.2.1. Non-strategy Condition (Everyday Speech)........................................................ 36 2.3.2.2. Fluency-Enhancing Strategy Conditions (Prolonged and Metronome-timed Speech) ................................................................................................................ 36 2.3.3. Compensation and Follow-up Questionnaire .............................................................. 38 2.4. Questionnaires .................................................................................................................... 39 2.5. Cognitive Screener ............................................................................................................. 40 2.6. Withdrawal Procedures....................................................................................................... 41 CHAPTER 3: DATA ANALYSIS .............................................................................................. 42 3.1. Power Analysis ................................................................................................................... 42 3.2. fNIRS Data Processing ....................................................................................................... 42 3.3. Speech Sample Analyses .................................................................................................... 44 v 3.4. Hypotheses-Driven Analyses…………………………………………..…………………46 CHAPTER 4: RESULTS ............................................................................................................ 48 4.1. Descriptive Analyses .......................................................................................................... 48 4.1.1. Participants .................................................................................................................. 48 4.1.2. Stuttering Severity ....................................................................................................... 48 4.1.3. Stuttering Severity Measurement Reliability .............................................................. 50 4.1.4. Speech Rate ................................................................................................................. 51 4.1.5. Speech Effort Self-Ratings .......................................................................................... 51 4.1.6. Montreal Cognitive Assessment (MoCA; Nasreddine, 2010) ..................................... 52 4.1.7. State-Trait Anxiety Inventory (STAI) .......................................................................... 53 4.1.8. Overall Assessment of the Speaker’s Experience of Stuttering (OASES) ................... 53 4.1.9. Handedness ................................................................................................................. 54 4.1.10. Family History of Stuttering ..................................................................................... 54 4.1.11. Years of Treatment .................................................................................................... 55 4.1.12. Medications ............................................................................................................... 55 4.2. Hypotheses-Driven Analyses of Lateralization .................................................................. 56 4.2.1. Hypothesis 1: Between-group (AWS vs. TS) Differences in Laterality of Activation During Conversation in the Everyday (Non-strategy) Condition................................ 56 4.2.2. Hypothesis 2a: Within-group Shifts in Laterality from the Non-strategy-use (Everyday) Condition to the Strategy-use Conditions (Prolonged and Metronometimed Speech) .............................................................................................................. 57 4.2.3. Hypothesis 2b: Between-group (AWS vs. TS) Laterality Differences During Strategy-use in the Prolonged Speech and Metronome Conditions ............................ 59 4.3. Ancillary Analyses of AWS and TS (n = 3 each) Removed from Main Comparisons ...... 61 CHAPTER 5: DISCUSSION ...................................................................................................... 64 5.1. Main Finding #1: Non-significant Between-Group Differences in Laterality During Everyday Speech ................................................................................................................ 65 5.2. Main Finding #2: Within-group Differences in Laterality from Everyday to Prolonged Speech and Metronome Conditions .................................................................................... 68 5.3. Main Finding #3: Between-Group Differences in Laterality During Speech StrategyApplication (Prolonged and Metronome Speech) .............................................................. 71 5.4. General Discussion ............................................................................................................. 72 5.5. Caveats ............................................................................................................................... 77 CHAPTER 6: CONCLUSIONS ................................................................................................. 79 TABLES ........................................................................................................................................ 81 FIGURES ...................................................................................................................................... 95 REFERENCES........................................................................................................................... 111 vi ABBREVIATIONS The following abbreviations are referred to throughout this document: aPFC ACC AWS BA BG bpm CN CW CWS CWNS DAF DCS dlPFC dmPFC DTI EEG ERPs fMRI fNIRS HbO HHb IFC IFG LI Met MEG PET PFC PS SI SMA SPECT spm SPM SN STG THb TS vlPFC vmPFC VF VFT WM anterior prefrontal cortex anterior cingulate cortex adults who stutter Brodmann area basal ganglia beats per minute caudate nucleus continuous wave children who stutter children who do not stutter delayed auditory feedback diffuse correlation spectroscopy dorsolateral prefrontal cortex dorsomedial prefrontal cortex diffusion tensor imaging electroencephalography event-related potentials functional magnetic resonance imaging functional near infrared spectroscopy oxygenated hemoglobin deoxygenated hemoglobin inferior frontal cortex inferior frontal gyrus laterality index metronome-timed speech magnetoencephalography positron emission tomography prefrontal cortex prolonged speech syllable initiation supplementary motor area single positron emission computerized tomography syllables per minute Statistical Parametric Mapping substania nigra superior temporal gyrus total hemoglobin typical speakers (adults who do not stutter) ventrolateral prefrontal cortex ventromedial prefrontal cortex verbal fluency verbal fluency task working memory vii LIST OF TABLES Table Page Table 1: Neural regions associated with speech/language production in AWS and TS ………………………………………………………………………………………..81-82 Table 2: Brodmann Areas (BAs) and functional connectivity corresponding to the prefrontal cortex regions to be measured by fNIRS …………………………………….83 Table 3: Speaking task blocks counterbalanced across speech strategy conditions…......83 Table 4: Participant characteristics of Adults Who Stutter (AWS) and Typical Speakers (TS)………………………………………………………………………...84-85 Table 5: Between-group speech sample analyses (percent syllables stuttered; %SS) for AWS (n = 11) and TS (n =11) across conditions…………………………………….86 Table 6: Inter- and intra-rater reliability for stuttering severity by SSI-4 (Riley, 2009) categorical scale and numerical score…………………………………………………....86 Table 7: Between-group speech rate (syllables per minute) in pre-testing phone and Everyday condition tasks for AWS (n = 11) and TS (n = 11)…………………..……….87 Table 8: AWS (n = 11) stuttering severity self-ratings across daily-life speaking situations ……………………………………………………………………...................87 Table 9: Between-group self-reported speech effort after each condition block in AWS (n = 11) and TS (n = 11)………………………………………………………………....88 Table 10: Between-group (AWS vs. TS) pairwise comparison following linear mixed model of mean left and right PFC HbO relative to baseline, Laterality Index and mean difference HbO between AWS (n = 11) and TS (n = 11)………………………………..89 Table 11: Within-group pairwise comparison following linear mixed model of mean difference in PFC HbO activation relative to baseline in left (channels 1-8) and right (channels 9-16) hemispheres and Laterality Index for AWS (n = 11) and TS (n = 11) ……………………………………………………………………………………………90 Table 12: Laterality Indices for all subjects (AWS and TS) across three conditions (Everyday, Prolonged Speech, Metronome)……………………………………………..91 viii Table 13: Relative change in Laterality Indices (LIs) for all subjects (AWS and TS) from the Everyday condition to the Prolonged Speech and Metronome conditions…….92 Table 14: Ancillary analyses of Laterality Indices (LIs) for subjects removed from main analyses (AWS and TS; n = 3 each) across three conditions (Everyday, Prolonged Speech, Metronome)………………………………………………………….93 Table 15: Ancillary between-group (AWS vs. TS) pairwise comparison following linear mixed model of mean left and right PFC HbO relative to baseline, Laterality Index and mean difference HbO between AWS (n = 3) and TS (n = 3) removed from main analyses…………………………………………………………………………….93 Table 16: Ancillary within-group pairwise comparison following linear mixed model of mean difference in PFC HbO activation relative to baseline in left (channels 1-8) and right (channels 9-16) hemispheres and Laterality Index for AWS (n = 3) and TS (n = 3) removed from main analyses…………………………………………………….94 ix LIST OF FIGURES Figure Page Figure 1: Lateral, ventral and medial views of the PFC with Brodmann Areas (BAs) 9, 10, 11, and 46 (Fuster, 2002) ………………………………………………………....95 Figure 2: fNIR sensor with 4 light sources and 10 detectors (top left) and 16 optode (channel) measurement locations registered on sensor (left, middle) and on brain surface image (right). fNIR sensor positioned on participants’ head (left bottom). Brain surface image from University of Washington, Digital Anatomist Project (Ayaz et al., 2012)………………………………………………………………………..96 Figure 3: Source of detectors of fNIRS 100CW measure (Obrig et al. 2000; http://www.biopac.com/fNIR300A-Data-Stimulation-System) ………………………...96 Figure 4: Order of optode positions with channels 1-8 measuring left hemisphere HbO and channels 9-16 measuring right hemisphere HbO (Tak & Ye, 2011)………………..97 Figure 5: Approximate BIOPAC source detector localization coordinates of fNIRS optodes (León-Carrion, 2008)……………………………………………………………97 Figure 6: Between-group (AWS and TS; n = 11 each) mean left hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome)…………...98 Figure 7: Between-group (AWS and TS; n = 11 each) mean right hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome)…………...98 Figure 8: Between-group (AWS and TS; n = 11 each) Laterality Indices across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality …………...............99 Figure 9: Between-group (AWS and TS; n = 11 each) Laterality Indices across Conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality…………................99 Figure 10: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices in the Everyday condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality……………………………....100 x Figure 11: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices in the Prolonged Speech condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality; *TS #8 leftward LI dominance……………………………………………………………………….......101 Figure 12: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices in the Metronome condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality………………………………102 Figure 13: Mean HbO across all 16 voxels for AWS and TS (n = 11 each) in the Everyday conversation task (SE bars) ……………………………………...………….103 Figure 14: Mean HbO across all 16 voxels for AWS and TS (n = 11 each) in the Prolonged Speech conversation task (SE bars)………………………………………....104 Figure 15: Mean HbO across all 16 voxels for AWS and TS (n = 11 each) in the Metronome conversation task (SE bars)……………………………..……………........105 Figure 16: AWS participant #2 significant HbO activation in Everyday (left), Prolonged Speech (middle) and Metronome (right) conditions; imaged acquired using fnirSoft (Ayaz, 2010)…………………………………………………………………...106 Figure 17: TS participant #10 significant HbO activation in Everyday (left), Prolonged Speech (middle) and Metronome conditions; image acquired using fnirSoft (Ayaz, 2010)……………………………………………………………………………106 Figure 18: Mean HbO per voxel (16 voxels per condition) across Everyday (left), Prolonged Speech (middle), and Metronome (right) conversation blocks for AWS participant #7…………………………………………………………………………...107 Figure 19: Ancillary between-group (AWS and TS; n = 3 each) mean left hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome)…....108 Figure 20: Ancillary between-group (AWS and TS; n = 3 each) mean right hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome)…....108 Figure 21: Ancillary between-group (AWS and TS; n = 3 each) Laterality Indices (LIs) across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality…...109 Figure 22: Ancillary between-group (AWS and TS; n = 3 each) Laterality Indices (LIs) across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality…...109 xi Figure 23: Ancillary Individual AWS (left graphs; n = 3) and TS (right graphs; n = 3) Laterality Indices in Everyday (top), Prolonged Speech (middle) and Metronome (bottom) conditions of participants removed from main analyses; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality………….110 xii CHAPTER 1: INTRODUCTION Developmental stuttering is a neurologically-based motor speech disorder (e.g., Kleinow & Smith, 2000) which typically emerges between the ages of 2-4 years and persists into adulthood in approximately 1% of the general population (Bloodstein, 1995; Yairi & Ambrose, 1999). In general, speech motor control in adults who stutter (AWS) is more variable, less automatic and less efficient relative to typical speakers (TS)(Chang & Zhu, 2013; Ingham et al., 2012). While some neuroimaging studies have revealed atypical right-dominant lateralization of activation in AWS (e.g., Biermann-Ruben et al. 2005; Ingham et al., 2012), the neural basis for stuttering remains largely a mystery. The majority of evidence suggests that the right-dominant atypical lateralization for speech and language in AWS represents compensatory rather than causal activation for less efficient or automatic speech language processing in the left hemisphere (e.g., Beal et al., 2013; Chang & Zhu, 2013). However, there is ongoing discussion in the field regarding whether the observed atypical right lateralization in AWS is a cause or the symptom of years of stuttering, as competition with or compensation for less efficient left hemisphere motor speech regions. While practice of fluency-enhancing strategies in real-world stuttering treatment frequently takes place in these conversational speech contexts, many brain imaging treatment studies (e.g., fMRI, EEG) of AWS are limited in the ability to measure lateralization of activation during naturalistic speaking situations (i.e., spontaneous conversation). Therefore, real-time brain imaging in conversational contexts is needed to 1 reveal more about the underlying neural mechanism associated with change in stuttering behavior when strategies are used to speak fluently. Treatment studies have revealed important information about the plasticity of the lateralization of speech and language in AWS. For example, atypical right-lateralized activation has been shown to “normalize” or shift more bilaterally/leftward after training with fluency-enhancing strategies known to reliably reduce stuttering, such as metronome-timed or prolonged speech (e.g., Braun et al., 1997; Neumann et al., 2003). One possible mechanism for the reduction of stuttering with such speech strategy use is that the altered speech patterns restore activation to left-sided motor speech planning areas throughout the cortex by increasing on-line self-monitoring. The altered speech patterns may compensate for reduced speech automaticity and may thus optimize coordination of articulatory, phonatory and respiratory systems (De Nil & Kroll, 2001). The prefrontal cortex (PFC) is an important region of such expected neural change during strategy-application in conversation. As the locus of executive control and self-monitoring of speech, the PFC is activated during the generation of complex and novel spontaneous discourse (e.g., Blank et al., 2002; Coelho et al., 2012). Given the topdown control of the PFC with strong interconnections to motor speech planning, timing and execution areas, PFC activation would be expected to shift bilaterally/leftward following training of fluency-enhancing strategies in AWS (Neumann et al., 2003). Because of the PFC’s role in strategy-use during conversation, functional near infrared spectroscopy (fNIRS) technology is well-suited to quantify such change in real-time as AWS speak spontaneously in a relatively naturalistic clinical setting. 2 The present study will therefore measure functional PFC hemispheric activation and lateralization in AWS and matched controls using fNIRS optical imaging technology during spontaneous speech generation tasks to determine (1) to what degree PFC activation and lateralization is different during habitual everyday speech in TS and AWS and (2) if lateralization of PFC activation shifts and “normalizes” when AWS are trained to use fluency-enhancing strategies (e.g., speaking with a prolonged speech pattern or with a metronome). It is anticipated that the results of the present study will provide a clearer understanding of lateralization of PFC cortical activity related to both stuttered and fluency-enhanced speech and strategy application in real time. Specifically, comparing differences in PFC activation during stuttered versus fluency-enhanced speech will reveal more information about the underlying neurobiology of self-regulation of fluency in AWS and may reflect a possible mechanism for changes in stuttering behavior during treatment. 1.1. Literature Review 1.1.1. Laterality of Activation During Speech Production Neuroimaging evidence indicates that laterality of brain activation during speech production is different in adults who do and do not stutter. Typical speakers (TS) present with left-lateralized activation during speech production, and speech is seemingly automatic and effortless (Price, 2012; Ries et al., 2011). Some key brain areas activated during “normal” speech production in TS are (see Table 1): the left inferior frontal gyrus (IFG) including Broca’s area for motor speech planning and production, the auditory cortex, the prefrontal cortex (PFC), the bilateral superior temporal gyri (STG) including 3 Wernicke’s for speech/language comprehension, the premotor and primary motor cortices, and the supplementary motor area (SMA; e.g., Hickok & Poeppel, 2007), as well as the contralateral right cerebellum (e.g., Brown et al., 2005). The stronger left lateralization for language in TS appears to be primarily associated with left-sided processing of speech sounds in the auditory cortex (Kell et al., 2011) and with semantic decision-making, such as determining which of two presented words is a better match for another word (Binder et al., 2009; Hickok, 2009; Josse et al., 2008). Further, invasive measures such as Wada testing (i.e., the temporary blockade of lateralized hemispheric functions using barbituate agents) in TS has indicated that lateralization for speech/language in more posterior regions of the cortex is significantly correlated with activation in frontal lobes in the same hemisphere, with the strongest correlation in the left dorsolateral PFC (Lehericy et al., 2000). The evidence suggests that posterior language lateralization is reflected ipsilaterally in the prefrontal lobes (Deblaere et al., 2004), except for the cerebellum which is contralaterally activated. In contrast to TS, neuroimaging findings consistently demonstrate that adults who stutter (AWS) present with relatively greater right hemisphere activation in areas homologous to the speech-motor and auditory processing regions listed for TS (e.g., De Nil et al., 2000; Foundas et al., 2001; Fox et al., 2000; Ingham et al., 2000; Loucks et al., 2011). In addition to relatively right-sided overactivations, AWS present with left hemisphere deactivations in Broca’s and Wernicke’s areas during stuttered speech, which are key regions for auditory processing and motor speech execution (Wu et al., 1995). In other words, right-hemisphere motor areas in AWS have been shown to be atypically 4 activated in the right hemisphere and left-hemisphere speech planning and production areas are relatively underactivated compared to TS (see Table 1). However, less is known about how lateralization of neural activation shifts in real-time just before and during stuttering episodes. Braun et al. (1997) identified realtime atypical activation patterns in the left hemisphere in AWS in a positron emission tomography (PET) study. Results indicated that the right hemisphere was activated during stuttered speech while the left hemisphere was hyperactive during fluent speech. Another real-time direct measure of brain function to examine laterality in AWS is magnetoencephalography (MEG)(Biermann-Ruben et al., 2005; Kikuchi et al., 2011; Salmelin et al., 1998, 2000). A recent case study using MEG (Sowman et al., 2012) indicated a significant decrease in activation of the left IFG (BA 47) with an increase in activation in the same region of the right hemisphere immediately prior to blocked/stuttered speech in an AWS. Similar to the findings of Braun et al. (1997), there was a significant increase in the left IFG just prior to successful initiation of fluent speech and a compensatory pattern of shifted activity from left to right IFG just before stuttered speech. The results were also consistent with other imaging studies indicating relative deactivation of the left IFG in AWS (e.g., Fox et al., 1996), an area associated with recovery from stuttering (Kell et al., 2009). A meta-analysis of fMRI and PET studies (Brown et al., 2005) confirmed these robust right-lateralized differences during speech production in AWS. For example, the right frontal operculum (RFO), which is the homologue for Broca’s area and functions in language repair and self-monitoring, was shown to be relatively overactive in AWS 5 (Giraud et al., 2008) and was negatively correlated with stuttering (Preibisch et al., 2003a). The compensatory RFO activation was hypothesized to be associated with less efficient left-sided Broca’s area function in persistent stutterers. Support for this hypothesis has been demonstrated by reduced white matter integrity in the area surrounding the left Broca’s (e.g., Chang et al., 2008; Kell et al., 2009) and a less developed superior longitudinal fasciculus III in AWS, which transmits and receives sensory premotor planning information to and from Broca’s (Cykowski et al., 2010; Makris et al., 2005). Evidence therefore suggests that AWS have both structural and connectivity differences in key left-sided motor speech networks, which are associated with reduced efficiency when planning and producing rapid speech as well as de-automatization of speech movements (De Nil & Bosshardt, 2001). Generally, such rightward lateralization of speech/language in AWS is thought to be compensatory rather than causal (e.g., Brown et al., 2005; Chang et al., 2011; Lu et al., 2010ab; Preibisch et al., 2003ab; Sommer et al., 2002). Importantly, this atypical compensatory activation can shift with improved fluency in AWS, demonstrating some ability to “rescue” and “normalize” lateralization of function (Neumann et al., 2005 p. 31 & 34). Similar observations of compensatory activation, or shifting of functional lateralization, are well-established in lesion studies in people with aphasia (see review Anglade, Thiel, & Ansaldo, 2014). Such relative differences in lateralization of speech production and speech motor control circuitry in AWS have been identified after years of persistent stuttering. Whether or not these patterns are also present in young children who stutter (CWS) just after 6 stuttering onset is less understood. For example, a recent MEG study (Sowman et al., 2014) comparing 12 preschool-age CWS and 12 controls indicated left lateralization in both groups as measured during a picture-naming task. The authors concluded that lateralization in CWS was not different from controls. The lateralization found in AWS was suggested to be a result of “neuroplastic adaptation” (p. 1) over time with persistent stuttering. However, these results were measured during simple, single word productions, which resulted in consistently fluent speech. It is unclear how these results would differ if measured during longer duration spontaneous conversation including stuttered speech. In terms of neurodevelopmental differences, young CWS have been shown to present with less developed grey and white matter tracks interconnecting the basal ganglia-thalamocortical (BGTC) circuit and regions underlying speech motor control (Beal et al., 2013; Chang & Zhu, 2013). If the BGTC circuit is not functioning efficiently early in development, this can alter fine motor speech sequence timing accuracy in the interconnected SMA and the PFC. These areas regulate timing for motor movements and mediate cognitive control of motor behavior respectively during subsequent development (e.g., Brown et al., 2005; Ries et al., 2013). Thus, disruption of this circuit for fine motor control in young CWS has been proposed to set the stage for reduced efficiency and fluency of speech planning and execution in older AWS with persistent stuttering (Chang & Zhu, 2013; Pollok et al., 2005; Wiener, Turkeltaub & Coslett, 2010). Further evidence for the effect of early disruption of the BGTC circuitry has been shown in a case study of an AWS with persistent developmental stuttering whose stuttering disappeared after a bilateral thalamic infarction as an adult. Specifically, the 7 infarct severed the connection between the dorsomedial (DM) and centromedian (CM) nucleus of the thalamus, the lateral PFC and the SMA (Muroi et al., 1999). The subsequent disruption of the two sub-networks of the BGTC (SMA-CM and DM-PFC) eliminated the stuttering, suggesting that motor speech control function in these interconnected networks was associated with stuttering behavior. Group differences between AWS and TS in motor circuit efficiency extend to non-speech motor movements as well. For example, AWS have less left-lateralized activation during timed finger-movements relative to TS (Neef et al., 2011) and reduced finger tapping performance during concurrent verbal tasks (Greiner, Fitzgerald & Cooke, 1986). Webster (1998, 2004) suggested that this effect may be due to a “fragile” left SMA in AWS which is vulnerable to interference from other brain activity. Alm et al. (2013) further found a rightward shift of non-speech motor activation in AWS, which the authors suggested could be associated with “monohemispheric” (p. 1) motor impairment in the left hemisphere and consequently reflect instability in bilateral speech motor control (due to difficulties initiating complex motor sequences). Overall, reduced connectivity and efficiency in speech motor regions in AWS appears to be related to atypical neural development from a young age which can affect (1) timing of internally-initiated motor movements, (2) motor control during complex motor sequences and (3) inhibition of extraneous speech movements, all of which have been shown to be associated with stuttering (Chang & Zhu, 2013). Specific evidence suggests that stuttering is related to difficulty with successful initiation of motor programs in the extended BGTC circuit and relative overactivation in the speech motor 8 timing network of adults with persistent stuttering (Brown et al., 2005; Chang & Zhu, 2013; Max & Gracco, 2005). The evidence bolsters the argument that speech production in AWS seems to be more effortful, or less automatic, with less efficient motor coordination relative to TS with compensatory right-sided activation in response to an inefficient left hemisphere speech-motor circuit. Therefore, speech production and general motor control appear to be more effortful for AWS with relatively greater right hemispheric activation compared to TS. The collective results suggest that AWS present with reduced automaticity of motor movements and timing instability for both speech and non-speech tasks (Zelanic, Smith, Franz & Ho, 1997). This difference appears to be associated with atypically right-sided lateralization in AWS, perhaps as a compensation for the inefficient left-hemisphere structures. Such neural activation has plasticity though, as lateralization can be shifted with improved fluency, demonstrating some ability to approximate more “typical” activation patterns of TS (Fox et al., 1996). Activation shifts have also been demonstrated when AWS speak using known fluency-enhancing strategies, such as speaking in time with the support of external timing cues (e.g., a metronome), to compensate for less efficient initiation of speech motor movements (Packman, Code & Onslow, 2007) as will be discussed below. 1.1.2. Fluency-enhancing Speech Strategies 1.1.2.1 Metronome-timed Speech Motor control of speech, internal timing synchronization and other manual movements appear to be less efficient in AWS relative to TS. Given these difficulties in 9 internal timing, certain speaking conditions provide compensatory external timing cues or alter typical speech production patterns (i.e., reduce demands to produce rapid sequential timing of speech motor movements) and can reliably enhance fluency in AWS. These include: choral reading/speaking, presentation of masking noise while talking, singing, listening to delayed auditory feedback (DAF) of speech, and paying greater attention to speech production as when speaking with an accent/dialect (e.g., Bloodstein & Ratner, 2008; Bothe, 2004; Guitar, 2013; Kalinowski & Dayalu, 2002). Speaking in time with a metronome is also known to reliably and immediately induce fluency in AWS with little training (e.g., Packman, Onslow & Menzies, 2000 for a review). The metronome provides an external timing signal (e.g., Davidow, 2013), which acts as a “pacemaker” of sorts (Buchel & Sommer, 2004). The external timing cue of a metronome is hypothesized to compensate for impaired internal cueing for automatic, self-timed speech motor movements in the basal ganglia (BG) of AWS (Alm, 2004, 2005; Taniwaki et al., 2006; Wu et al., 2011). The BG includes the putamen, globus pallidus and caudate as well as the substantia nigra, and is highly interconnected with the PFC and motor circuitry, all of which are involved in speech timing, sequencing and sensorimotor integration (e.g., Nagy et al. 2006). Typically, movements are coordinated by the BG sending “go” signals to the SMA (Alm, 2005), and the BG has been shown to be less active during habitual speech in AWS (Brown et al., 2005). However, BG activation levels were shown to shift closer to those of controls in a metronome-timed speech study of AWS and TS (Toyomura et al., 2011). Wu et al. (2011) demonstrated that external timing cues provided by the 10 metronome supported fluent speech in AWS by bypassing the less efficient BG-SMA pathway and activating the cerebellar-premotor cortex system instead. Thus, external auditory pacing with a metronome may compensate for anomalous internal timing cues in the motor circuit of AWS described previously. Activation shifts away from the BG, thus facilitating integration of rhythm perception and production via the external signal (Alm, 2004; Haslinger et al., 2001; Smits-Bandstra & De Nil, 2007). The changes in the motor circuit of AWS as a result of metronome-timed speech are relevant for the present investigation of PFC laterality. Fluency will be elicited using this strategy, and laterality changes in the motor circuit will be reflected in the PFC, which is directly interconnected with the BG (Alexander et al., 1986). Specifically, the left dorsolateral (dlPFC; BAs 46/9) has projections connecting to the SMA via the BG, the thalamus, ACC and the cerebellum, all of which are involved in language processing and speech motor control (Miller & Cohen, 2001; Strick, Dum & Picard, 1995). Functionally, the BG regulates top-down projections from the PFC to control which motor signals gain access to the PFC (Stocco & Lebiere, 2012). The lateral PFC regulates motor inhibition by gating motor commands via the thalamus to the motor cortex. Together, the PFC resists goal-irrelevant movements and the BG is involved in taskswitching and flexible control of movement (Van Schouwenburg, Aarts & Cools, 2010), such as movement coordination during complex speech production. These motor circuit interconnections with the PFC have been demonstrated in AWS by “normalization” of BG activation during metronome-timed speech. Metronometimed speech acts by pacing (and consequently slowing) speech, enhancing fluency by 11 reducing language processing and motor demands in the fronto-temporal regions of the left hemisphere. This strategy also improves auditory perception and processing, as reflected by an increase in activation in the left auditory cortex in AWS during fluencyenhanced speech (Salmelin et al., 1998). Specifically, activation was shown to increase in the BG and bilateral/left-sided fronto-temporal speech regions, which are typically right hemisphere dominant in AWS, during metronome-timed speech (Toyomura et al., 2011). The effect was found even during spontaneously generated connected speech tasks (Braun et al., 1997; Stager, Jeffries & Braun, 2003). Therefore, metronome-timed speech alters neural activation in AWS relative to habitual stuttered speech, with more bilateral and leftward shifted activation in speech-language and motor regions. Several theories have been proposed to describe the mechanism underlying the fluency-enhancing effect of metronome-timed speech. One theory refers to the previous discussion that AWS have less efficient sensorimotor function than TS, and spontaneous language generation of novel and personal narratives stresses this underlying predisposition (Braun et al., 1997). Similarly, Stager et al. (2003) proposed that metronome-timed speech increases efficiency and integration of the auditory-motor system of AWS to self-monitor and to guide complex speech production. Another explanation for the effectiveness of metronome-timed speech is the syllable initiation (SI) theory, a later version of the Vmodel (Packman, Onslow, Menzies, 2000; Packman, Code & Onslow 2007). The SI theory builds from Webster’s (2004) model in which the SMA, which controls and initiates planning and production of spontaneous speech and connects 12 directly to the BG, is “vulnerable” in AWS. The PFC and pre-SMA also interconnect with the BG, and activation is greater in both regions during spontaneous speech generation tasks (Blank et al., 2002). Stuttering is then indirectly influenced by the increased motor demands of spontaneous speech due to high linguistic stress variability, which can affect speech motor control in the unstable SMA-BG circuit. Metronometimed speech acts to reduce this linguistic stress variability across syllables, and concurrently the amount of short phonated intervals. As a result, longer voiced sounds are produced more frequently and stuttering behavior is reduced (Davidow, 2013). Thus, SI theory suggests that metronome-timed speech facilitates speech coordination and timing while compensating for unstable sensorimotor integration in AWS (Packman, Code & Onslow, 2007). The reduced motor variability during metronome-timed speech allows for stabilization of the speech motor system during speech production. This is reflected by a reduction in self-reported speech effort required during metronome-timed relative to habitual (stuttered) speech in AWS (Ingham et al. 2009, 2012). Overall, results suggest that metronome-timed speech can alter BG, PFC and SMA activation in AWS and induce more bilateralized/leftward neural activation in motor speech regions to compensate for impaired internal rhythm generation and sensorimotor integration in the BG. 1.1.2.2. Prolonged Speech (PS) While metronome-timed speech reliably enhances fluency in AWS with minimal training and reported effort, treatment gains do not generalize well once the stimulus is removed, and speech quality is often rated as “unnatural” depending on the 13 speed of production (Ingham et al., 2012). Prolonged speech (PS) is another strategy that reliably enhances fluency in AWS and has been found to be one of the most effective treatments for AWS with long-term effects (e.g., Andrews et al., 1980; Herder et al., 2006). In general, PS therapy involves training AWS to learn and consistently use a manner of speaking that is incompatible with stuttering. The strategies include reducing speech rate, prolonging vowels, stretching out words, maintaining continuous vocalization, having soft contacts of articulators and easing into the first sound in words (Packman, Onslow & Menzies, 2000). Similar to metronome-timed speech, PS reduces variability of linguistic stress, and thus motor effort, while also slowing articulatory movements (Civier, Tasko & Guenther, 2010). A standardized PS therapy is the Camperdown program (O’Brian et al., 2003). Instead of training specific speech patterns such as modified rate and gentle onsets, the Camperdown program uses exemplar videos of clinicians or an adolescent speaking with PS (http://sydney.edu.au/health-sciences/asrc/health_professionals/asrc_download.shtml). The adolescent video model speaks at approximately 33 syllables per minute (spm), which is significantly slower than the normal adult speech rate from 180-220 spm (Ingham & Andrews, 1971). The AWS who is receiving treatment is instructed to imitate the model and practice with clinician support to start to utilize this speech pattern in the clinic. Eventually, practice is transferred to everyday speaking situations with progressively more natural-sounding speech. Typically, PS training requires long-term practice (weeks to months) to maintain fluency gains. The current protocol will involve short-term training (~6 minutes) of this speech pattern. 14 In terms of neurological changes observed following PS therapy, activation shifts are proposed to reflect differences in speech effort, automaticity and attention during speech production, as speech in AWS is generally more effortful and less automatic relative to TS (e.g., De Nil & Kroll, 2001). Specifically, lateralization has been shown to shift from more right-sided activation pre-treatment to bilateral/left-sided activation post-treatment (e.g., Neumann et al., 2005). Leftward increases in activation were also observed in “typically” activated temporal speech-related regions recruited by TS (IFG and STG) where anomalous white matter had been detected. Neumann et al. (2005) also found increased, and more distributed, neural activation bilaterally in the premotor and motor cortex one year after maintenance treatment (in the direction of TS). Some right-dominant lateralization reemerged two years post-treatment with an increase in stuttering behavior. The authors suggested that PS therapy increased self-monitoring of speech articulation while reducing demands on the AWS’s speech motor systems, allowing for “rescued” and “normalized” left-sided activation via compensatory neural reorganization (Neumann et al., 2005 p. 31 & 34). Neurological changes specific to BG activation have also been revealed pre- and post-treatment after three weeks of intensive PS treatment in AWS (Giraud et al., 2008). As was discussed in the metronome-timed speech section, the BG is interconnected with the PFC to facilitate self-generated movements and to inhibit extraneous movements. In the Giraud et al. (2008) study, pre-treatment stuttering severity was positively correlated with bilateral activation in the caudate of the BG and negatively correlated with the left substantia nigra, as these two areas operate antagonistically (Gerfen et al., 1990). 15 However, these correlations were no longer significant with post-treatment fluency gains, suggesting that BG activation “normalized” in more severe AWS. Concurrent posttreatment increases in the left motor and auditory cortices and putamen also suggested improved connections between Broca’s speech motor planning area and the motor cortex (Giraud et al., 2008). The shift in activation was proposed to reflect lateralized deviant motor functions in support of Alm’s (2004) hypothesis that atypical BG function is associated with disturbed timing of speech production. Thus, AWS have been shown to present with atypically right-sided BG activation, and PS therapy can act directly and significantly on BG (and therefore PFC) function to correct this activation (Giraud et al., 2008). Similar post-treatment leftwardshifts of neural activation have also been shown in EEG studies (Boberg et al., 1983; Moore, 1984) and PET studies (De Nil et al., 2003; De Nil & Kroll, 2001) after stuttering-reduction therapies. The collective results suggest that practice of the PS strategy can lead to a bilateral/leftward shift in brain activation in homologous tissue which reflects compensation for (1) less dense white fibers in the speech-motor circuit (2) atypical BG and motor circuit function including the PFC and (3) relatively increased activations in the right-hemisphere during speech in AWS (Braun et al., 1997; De Nil, Kroll, Kapur & Houle, 2000). Because PS therapy emphasizes constant ongoing selfmonitoring of speech, requires focused attention, and slows articulatory movements, the post-therapy results seem to indicate that PS increases speech motor control and reduces motor effort in AWS (De Nil et al., 2003; Packman, Code & Onslow, 2007). 16 Prolonged speech (PS) and speaking with a metronome therefore induce fluency in AWS in overlapping ways. Both can induce bilateral/leftward shifts in neural activation in speech motor regions to “normalize” activation in the BG, SMA and PFC loop, by reducing demands on motor control (Packman et al., 1996; Toyomura et al., 2011). Metronome-timed speech does so by providing an external timing cue to compensate for and “normalize” impaired internal rhythm generation and sensorimotor integration in the BG of AWS (Neumann et al., 2005). Effort required to initiate syllables is also reduced during metronome-timed speech due to reduced variability in linguistic stress (Packman, Code & Onslow, 2007). Intensive PS practice seems to improve motor function by allowing the speaker time to self-monitor articulation, focus attention on fluent speech production and subsequently utilize more left-lateralized speech areas. These two fluency-enhancing speech strategies will be trained in the present study to compare PFC neural activation patterns during narrative discourse generation in AWS and TS when speaking normally (stuttering) versus when fluent. 1.1.3. Prefrontal Cortex The PFC is the target region of interest in the present study to examine lateralization during conversational speech tasks. In general, the PFC is known to be implicated in higher-order cognitive or “executive” functions, such as decision-making, inhibition, task-shifting, and attention regulation (Miller & Cohen, 2001; Ridderinkhof et al., 2004). As mentioned previously, the PFC is highly interconnected with speech motor regions known to be relatively overactivated (e.g., pre-SMA) or underactivated (e.g., BG) in AWS compared to TS during speech production (e.g., Miller & Cohen, 2001). If an 17 individual is relatively right-dominant for language as has been observed in AWS, the right PFC will also reflect this lateralization pattern (Deblaere et al., 2004). In contrast, spoken language is typically associated with activation in the left PFC in TS (e.g., Price, 2012). The PFC is therefore a key target region because laterality of activation of the PFC is associated with: (1) speech strategy-use generally and (2) laterality of speech and language production and motor control (Gabrieli, Poldrak & Desmond, 1998). Three specific PFC subregions will be targeted in the present study: the anterior (aPFC), dorsolateral (dlPFC), and ventromedial (vmPFC)(see Table 2 and Figure 1). The largest region of activation to be measured is the aPFC (BA 10). The aPFC is involved in multitasking, or tasks requiring coordination of two or more executive tasks (Gilbert et al., 2006; Ramnani & Passingham, 2001), execution of practiced tasks, generation and monitoring of internally produced and maintenance of attention (Ramnani & Owen, 2004; Burgess, Quayle & Frith, 2001). The BA 10 region is also activated during “default mode,” such as baseline “mind wandering” at rest (Ramnani & Owen, 2004) with rightlateralized resting state functional connectivity in the inferior frontal and middle frontal gyrus (e.g., Medvedev, 2014). The other two PFC networks (1) dlPFC and (2) vmPFC (see Table 2) dynamically adjust attention and cognitive resources as task demands change (Glascher et al., 2012). The first network, the dlPFC (BA 9/46), is involved in cognitive control such as conflict monitoring, attention switching, and response inhibition to maintain information in a highly active state in working memory (WM), especially in the presence of interference (Glascher et al., 2012; Kane & Engle, 2002; Wagner et al., 2001), and when learning 18 verbal strategies (Savage et al., 2001). The dlPFC connects to the sensorimotor cortices and implements cognitive control by regulating behaviorally-relevant attention, rulebased action selection information and executive functions when task demands are increased (Bunge et al., 2001; Goldman-Rakic, 1987). Injury to the left dlPFC impairs spontaneous discourse with reduced sentence and grammatical complexity and story coherence (Coelho et al., 2012). The final network, the vmPFC (BA 10/11), is involved in emotion-based decision-making (Bechara, Damasio & Damasio, 2000; Ridderinkhof et al., 2004) by regulating emotional responses and habits via connections to the amygdala, hypothalamus and nucleus accumbens (Ghashghaei & Barbas, 2002; Price, Carmichael & Drevets, 1996). Right vmPFC activation has been associated with increased threat vigilance (Rauch et al., 1997), negative affect (Davidson et al., 1990), and anticipation of positive and negative consequences when public speaking (Davidson et al., 2000). In contrast, the left vmPFC activation is associated with inhibition of the amygdala (Davidson, 1998) and increased positive affect (Davidson et al., 1990; Sutton et al., 1997). Therefore, the PFC is functionally diverse with widespread interconnectivity to posterior cognitive, motor, emotion and attention networks, which makes it an excellent target for the present study, which will study lateralization during strategy-use in complex spontaneous speaking tasks. The next question is how PFC activation would then change when strategies are applied to reduce stuttering and the effortfulness of speech in AWS. In order to answer this question, lateralization of PFC activation during 19 verbal strategy-use more generally needs to be understood. A summary of PFC function during strategy-application will be presented prior to reviewing what is known about PFC activation during complex discourse production in TS relative to AWS. 1.1.4. PFC Activation During Effortful Strategy-Application General strategy learning and application initially requires effort and executive function, with recruitment of the PFC for top-down control (Elfgren & Risberg, 1998). For verbal strategy-use, executive and semantic function are primarily measured using verbal fluency tasks (VFTs)(e.g., Henry & Crawford, 2004). Phonemic VFTs include asking a participant to generate multiple single words that start with the same letter (e.g., “f”) in a set time period. Semantic VFTs involve generation of as many words as possible in a category, such as animals. Both require greater language processing demands than for typical speech in terms of vocabulary knowledge, access and selection, executive function and working memory (Blomgren et al., 2003). They also require vigilance to follow and maintain rules as well as to self-monitor performance and productivity (Elfgren & Risberg, 1998). AWS differ from TS in VFT performance and neural activation during these tasks. Specifically, VFTs are typically associated with greater left-lateralized PFC activation as demonstrated in focal lesion studies in TS (e.g., Robinson et al., 2012). Further, the left IFG and dlPFC (BAs 9/46) have been shown to be more active during phonemic VFTs in TS (Frith et al., 1991; Herholz et al., 1996; Raichle et al., 1994; Robinson et al., 2012; Warburton et al., 1996; Wood et al., 2001). In contrast, a study of phonemic VF skills in school-age children and adolescents who stutter indicated 20 significantly lower VFT performance in these groups compared to controls, suggesting specific executive function impairment including the frontal lobes (Bahrami, Nejati & Pooretemad, 2014). AWS have also been shown to have impaired performance in tasks which require focused and shifted attention, which are skills needed to efficiently and successfully complete VFTs (Heitmann, Asbjornsen & Helland, 2004). For example, Blomgren et al. (2003) demonstrated greater right-hemisphere activation in right prefrontal regions in AWS relative to TS during a covert verb and noun generation task. The collective results provide evidence that AWS may present with lower performance and greater right hemispheric frontal activation on cognitively demanding tasks relative to controls. Additionally, there are differences in activation depending on the effortfulness of tasks. For TS, activation levels increase in the right dlPFC when word retrieval becomes more difficult (Howard-Jones et al., 2005; Sachs et al., 2011). Relatively greater bilateral activation has also been reported in the dlPFC (BAs 9/46) and middle frontal gyrus (BAs 10/11) during cognitively demanding executive function tasks, such as the Stroop task (León-Carrion et al., 2008) and the non-speech Wisconsin Card Sort Task (WCST; e.g., Buchsbaum et al., 2005; Monchi et al., 2001). Thus, there is an even wider bilateral recruitment of neural regions as task difficulty and effort increases when engaged in cognitively challenging tasks for TS, which require effort for individuals to apply strategies, maintain attention and monitor performance to generate correct responses (Barch et al., 1997; Cadoret, Pike & Petrides, 2001). 21 Discourse, which is complex language production in real-world contexts, similarly engages prefrontal and speech-motor planning networks. Simply processing or comprehending organized narratives activates a widely distributed bilateral network including the vmPFC (BA 8, 9, 10), thalamus, putamen, caudate, cerebellum and amygdala (Xu et al., 2005). More complex discourse tasks, such as spontaneous production of narratives or monologues, involve higher-level processes of attention, thought selection, semantic knowledge, working memory, grammar formulation, phonetic planning and speech production which engage prefrontal regions (e.g., Bajaj, 2007). For example Coelho et al. (2012) reported that TS with left dlPFC (BA 9) damage had specific difficulties with narrative coherence when asked to retell a story. Neuroimaging studies have also shown that activation during discourse in TS is dynamic and can shift rapidly or differentially reorganize depending on cognitive task demand and context, such as when single words or complex narratives are presented (e.g., Stephan et al., 2003). For example, Blank et al. (2002) found that the medial and lateral PFC were activated more during propositional speech, or novel formulation of a message using semantic and episodic memory (e.g., answering the question “tell me about your last holiday”), compared to automatic, less effortful non-propositional speech (e.g., counting or recitation). Therefore, the bilateral dlPFC and ventromedial PFC are regions of interest which are expected to be activated during more complex discourse processing and production in TS. Lateralization of PFC activation during complex speech generation is expected to be different in AWS relative to TS. For example, Watson et al. (1994) found that some 22 AWS are more likely to present with atypical right-sided activation in inferior frontal regions as a result of less efficient cognitive, motor and linguistic processing during complex discourse tasks. Further, Braun et al. (1997) demonstrated relatively greater bilateral prefrontal cortex (PFC) activation in the dlPFC (BAs 8, 9), vmPFC (BA 11) and medial anterior PFC (aPFC, BA 10) in AWS during habitual (“everyday” stuttered) speech compared to a simple oro-motor movement task. The results are consistent with motor skill learning generally, such that there are increases in PFC activation during initial performance of novel motor tasks (Jenkins et al., 1994). With practice, there are subsequent decreases in the left dlPFC (BA 46), primary motor cortex, cerebellum and BG (Floyer-Lea & Matthews, 2005). The attenuated PFC activation has been demonstrated in AWS, with less activity in the left dlPFC, striatum, and bilateral vmPFC cortices (associated with error detection and speech motor activity regulation) during metronome-timed speech relative to habitual speech (Braun et al., 1997). The reduced activation suggested that the metronome strategy decreased the effort required for speech production in AWS, with a decline in the need for PFC control motor performance (Braun et al., 1997). Overall, evidence suggests that AWS present with more bilateral/rightward PFC activation during speech production relative to TS. Cognitive effort is also associated with differences in PFC activation for both AWS and TS. Training both groups to use speech strategies (prolonged and metronome-timed speech) to enhance fluency during discourse production is expected to elicit different activation relative to habitual (“everyday”) speech depending on the effortfulness of the strategy-application. Strategy- 23 application is expected to be associated with greater bilateral recruitment of prefrontal regions in TS and reduced bilateral recruitment in AWS with overall PFC activation levels shifting leftward as effortfulness or cognitive difficulty increases. Finally, total activation is expected to be positively correlated with self-reported speech effort for both AWS and TS. The manner in which these proposed neural activation patterns will be measured will be discussed in the following section. 1.1.5. Neuroimaging Measures of Activation Change During Speech Production While change in brain activity of AWS during speech production has been measured using a variety of neuroimaging modalities including fMRI (e.g., Chang et al., 2009), PET (e.g., Braun et al., 1997), EEG (e.g., Özge et al., 2004), SPECT (e.g., Pool et al., 1991), MEG (e.g., Kikuchi et al., 2011, Sowman, 2014), and DTI (e.g., Chang et al., 2011), these different imaging methods are limited in the ability to make real-time measures of naturalistic connected speech (Irani et al., 2007). For example, fMRI is the current ‘gold standard’ in imaging research, but this method is restrictive in that it is loud, expensive and susceptible to movement artifacts, thus limiting measurement to covert speech production, or reading/saying short utterances. Therefore, there are only a few studies in the stuttering literature that report real-time activation changes during connected stuttered speech in more naturalistic speaking contexts. For instance, MEG and PET have been used to make real-time measures, but PET requires use of a radioactive tracer and both methods are expensive and restrictive with bulky imaging equipment (e.g., Braun et al., 1997; Ingham et al., 2000; Sowman et al., 2012, 2014; Stager et al., 2003). 24 Thus, there is a need for more empirical research utilizing neuroimaging methods that allow for real-time measures of activation during connected speech, which can also be safely used to image children, using much less expensive, portable and less restrictive imaging technologies. Optical imaging technologies may provide a possible solution to this need, as an innovative, safe way to measure such real-time hemodynamic changes in blood flow using non-invasive near-infrared light (650-900 nm). Two of these optical technologies, functional near infrared spectroscopy (fNIRS) and diffuse correlation spectroscopy (DCS), have been used to measure relative change in neural oxygenated hemoglobin (HbO), deoxygenated hemoglobin (HHb) and total hemoglobin (THb) concentration in typical and clinical populations. 1.1.6. Real-time Activation Measures During Spontaneous Speech with Functional Near Infrared Spectroscopy (fNIRS) A fNIRS system is a portable optical neuroimaging research tool capable of monitoring the brain's hemodynamics, when performing cognitive tasks in naturalistic contexts. Both DCS and fNIRS have been validated for use in human research (see Yu, 2012 and Quaresima, Bisconti & Ferrari, 2011 for reviews), and cross-validation studies comparing fNIRS with other imaging modalities have been completed with good concordance of fMRI signals (e.g., Alderliesten et al., 2013; Gervain et al., 2011; Kennan et al., 2002; Lindauer et al., 2010). In terms of clinical applicability, fNIRS (LED units especially) can be used in any setting such as in a clinic, at the bedside, or in an office/laboratory. Further, fNIRS has been successfully used to safely study language and cognition in over a hundred empirical studies in children and adults, in persons with and 25 without neurological disorders (e.g., ADHD, schizophrenia, dyslexia, depression, Alzheimer’s, anxiety, traumatic brain injury, autism) in a variety of naturalistic settings (see reviews Arenth, Ricker & Schultheis, 2007; Ehlis et al., 2013; Homae, 2014; Irani et al., 2007; Kovelman et al., 2009; Quaresima et al., 2012; Shalinsky et al., 2009). Dieler, Tupak and Fallgatter (2012) and Rossi et al. (2012) provide extensive reviews of fNIRS studies specifically involving lateralized speech and language-related tasks. Thus, fNIRS technology has been successfully utilized to measure lateralized neural change during spontaneous speech and language tasks in naturalistic speaking contexts as well as inter- and intra-hemispheric connectivity during rest (e.g., Medvedev, 2014). Because the present study aims to measure PFC activation change during highly fluent and stuttered speech during spontaneous speech generation in AWS, optical imaging using fNIRS technology is the ideal choice to make such real-time measures. Functionally, the fNIRS technology employs flexible optic fibers that carry wavelengths of infrared light (via lasers or light-emitting diodes, LEDs) to (source) and from (detector) tissue in an optical frequency window that easily passes through a few centimeters of most tissue (see Figures 2 & 3). This allows continuous monitoring of oxygenated hemoglobin (HbO) changes by detecting back-scattered light intensity. The attenuated changes of light intensity across time are measured by fNIRS and interpreted as relative changes in HbO and HHb (deoxygenated hemoglobin), calculated using a modified Beer-Lambert law (Cope & Delpy, 1988; Hoshi, 2005), thereby providing information about neural activity and total blood flow based on diffuse light scatter. Generally, higher relative oxygenation in cerebral blood flow is indicative of greater 26 cognitive effort. The current study will use a LED-based fNIRS unit to measure attenuation or change in concentration of HbO relative to baseline in the PFC during speech-generation tasks (see Dieler et al., 2012 and Scholkmann et al., 2014 for reviews). Specifically, the anterior, medial, superior, ventral and dorsolateral PFC (BAs 9, 10, 11 and 46) will be measured in AWS and TS (see Table 2 and Figures 1 & 2). To the author’s knowledge, only one fNIRS study (Sato et al., 2011) and one pilot DCS study (Tellis, Mesquita & Yodh, 2011) have been conducted in AWS. Both studies found changes in relative blood flow that were consistent with other fMRI and brain imaging studies in AWS indicating increased activation of motor areas and reduced activation in auditory processing areas relative to typical speakers. Tellis et al. (2011) used DCS to measure brain blood flow differences in Broca’s area (BA 44), the left PFC and the right PFC region (as a control) during speaking tasks (e.g., counting, singing, reading aloud and choral reading, monologue) in one AWS and a matched control. Results indicated statistically significant changes in cerebral blood flow in the left PFC during monologue in the control participant while the AWS did not show this change. The AWS also presented with atypical right-hemisphere laterality in the PFC during the monologue task. The findings were consistent with other studies indicating greater motor region activation with only marginal changes in speech/language and auditory processing regions (Blomgren et al., 2003; Brown et al., 2005). Thus, real-time blood flow measures are consistent with other imaging studies (fMRI) in AWS and can be utilized to measure lateralization during various speaking tasks. The single known fNIRS study of people who stutter (Sato et al., 2011), 27 measured functional lateralization of auditory speech processing in preschool-age (n = 6), school-age (n = 7) and adults (n = 10) who stutter and matched controls using a laser fNIRS localized bilaterally over temporal areas. All three stuttering groups in demonstrated an atypical lack of the left hemispheric advantage observed in matched controls during auditory processing of phonemic (different final vowels) relative to prosodic (different pitch contours) stimuli. Further, severe stuttering was positively correlated with atypical rightward lateralization for the phonemic contrast conditions (Sato et al., 2011). This study demonstrated that (1) fNIRS is a practical means to gather lateralization data in people who stutter across development, (2) right-sided laterality in auditory processing of speech is present in preschool-age CWS, and (3) laterality in AWS is possibly related to the onset of stuttering rather than a consequence of years of stuttering. The results of Sato et al. (2011) are similar to those found by Weber-Fox, Wray and Arnold (2013), which showed atypical lateralization in preschool-age CWS during language processing as measured by event-related potentials (ERPs). Thus, fNIRS studies have revealed right-lateralized activation results which are consistent with other imaging technologies of AWS. Certain participant-specific factors need to be considered when examining speech/language generation using fNIRS. For example, age has been found to be a factor in fNIRS studies, with more bilateral or right-sided activation during narrative comprehension in elderly subjects, which is typically left-lateralized in younger subjects (Scherer et al., 2007). Further general decline in activation in elderly subjects relative to 28 younger participants has been shown with reduced hemispheric asymmetry during VFTs (Herrmann et al., 2006), and a significant decline in dlPFC activation during working memory tasks (Kwee & Nakada, 2003). Additionally, grey and white matter volumes decrease in the PFC with age, which can affect overall cognitive performance (Raz, 2000). Holtzer et al. (2011) also found that young adults presented with greater increase in brain activation in response to increased cognitive task demands relative to older individuals. Thus, age-related decline and shift in activation must be accounted for when examining language performance in fNIRS studies. Further, gender differences have also been demonstrated with fNIRS, such that women generally present with more bilateral representation than men (e.g., Okada, Takahashi & Tokumitsu, 1996) and greater overall oxygenation changes have been found in men during VFTs tasks (Kameyama et al., 2004). Handedness has also been shown to significantly correlate with laterality of fNIRS response (e.g., Watanabe et al., 1998). For these reasons, only right-handed adults matched by gender and age (between ages 18-55) will be enrolled in the present study. 1.2. Purpose Based on the above discussion, there is persuasive evidence to warrant making real-time measures of PFC activation changes in AWS during spontaneous discourse production compared to speaking while using fluency-inducing strategies. The primary aim of the present study is to use fNIRS to measure functional PFC hemispheric lateralization in AWS and TS during speech generation tasks to determine if lateralization is shifted by “turning on” trained fluency-enhancing strategies (e.g., speaking with a 29 prolonged speech pattern and with a metronome). Thus, the current study will compare between- and within-group PFC neural activation during stuttered (internally-timed) and near-fluent speech in an externally-timed metronome condition and a highly selfmonitored prolonged speech condition. 1.3. Hypotheses Hypothesis #1 was that AWS would present with greater right-hemisphere PFC oxygenation relative to typical speakers in everyday speech tasks in which no speech strategies were applied. Hypothesis #2 was that AWS would present with reduced righthemisphere PFC activation (leftward shift in Laterality Index) during fluency-enhanced speech strategy tasks (prolonged and metronome-timed speech) relative to everyday (stuttered) speech tasks. 30 CHAPTER 2: METHODS The current study utilized a prospective design to examine lateralization of PFC activation during stuttered and fluency-enhanced speech in adults who stutter (AWS) and typical speakers (TS) during discourse tasks (e.g., spontaneous conversation) as measured in real time using fNIRS technology. 2.1. Inclusion and Exclusion Criteria Inclusion criteria to be enrolled in the study: males and females between 18-55 years old, English as the first language, and right-handedness (determined by self-report and the Edinburgh Handedness Inventory--Short Form, modified from Oldfield, 1971 by Veale, 2013). Exclusion criteria: reported traumatic brain injury (including concussion) with loss of consciousness for more than 5 minutes, reported history of substance abuse, attention deficit-hyperactivity disorder, obsessive compulsive disorder, psychiatric illness, language or learning disability (other than stuttering), cerebral vascular accident or neurological disease. Group classification of AWS or TS was based on self-report of persistent developmental stuttering (childhood onset fluency disorder), diagnosis by a certified speech-language pathologist (the author), and >2% syllables stuttered based on a pretesting phone conversation. TS presented with less than 1% syllables stuttered on the same screening conversation and self-identified as fluent speakers without any personal history of stuttering. 31 2.2. fNIRS Recording The primary measurement involved a continuous wave (CW) functional near infrared optical brain imaging unit fNIR100-ACK-W (BIOPAC Systems, Inc.; www.biopac.com), a stand-alone functional brain imaging system that includes a control unit and sensor for CW-fNIRS and Cognitive Optical Brain Imaging (COBI) Studio software (Ayaz & Onaral, 2005). The fNIRS sensor detects the oxygen levels in the PFC and provides values for oxygenated (HbO) and deoxygenated hemoglobin (HHb) in realtime. The sensor is mounted in a flexible band worn on the forehead that includes four IR light sources (730 and 850 nm wavelengths) and ten detectors with a fixed sourcedetector separation of 2.5 cm (channels 1-8 = left hemisphere; 9-16 = right hemisphere; see Figures 2 & 3 for CW fNIRS equipment and sensor locations). The COBI studio software allowed for viewing of the PFC hemodynamic response in real time and saved data for post-acquisition analysis. FnirSoft software (Version 3.1; Ayaz, 2010) was used for signal processing and to prepare data for statistical analysis using IBM SPSS (Version 22, IBM SPSS, Inc.). 2.3. Procedures This study’s protocol was approved by the University of Vermont (UVM) Committee on Human Research in the Behavioral Sciences. Individuals who expressed interest in the study by contacting the principal investigator (PI) by phone or email were screened during a telephone conversation. During this phone screening, the participant was given a brief summary of the study, similar to the description in the purpose section of the consent form. They were informed of the kinds of tasks they would be asked to 32 complete and the time commitments of the study, as well as general information about the fNIRS technology. Participants were asked to provide verbal responses to elicit some basic background information and to determine eligibility. Participants gave verbal consent for the examiner to audio record the conversation (at least 300 syllables) for later analysis of percent syllables stuttered. If the participant verbally consented and was eligible, s/he was sent (via email) a Case History Form and the study consent forms. Only the AWS group was sent a standardized questionnaire, the Overall Assessment of the Speaker's Experience of Stuttering (OASES)(Yaruss & Quesal, 2008; ~15 minutes to complete; see 2.4.). To maintain confidentiality, participants were coded using letters and numbers depending on group/enrollment order (e.g., FS1= fNIRS participant who stutters #1; FC4= fNIRS control participant who does not stutter #4). Participants then scheduled a one hour appointment which took place in Pomeroy Hall on UVM campus. Each participant’s completed an OASES questionnaire and Case History were reviewed by the examiner prior to the fNIRS testing to verify eligibility. Each subject received an oral and a written explanation of the purposes and procedures, and written informed consent was obtained by the PI. Participants then completed the Montreal Cognitive Assessment (MoCA; Nasreddine, 2010) a one-page 30-point cognitive screen administered by the author (see 2.5.). If he/she passed this screening (scored 26 or above), participants then completed the State-Trait Anxiety Inventory (STAI; Spielberger et al., 1983; see 2.4.). 33 Participants were then trained by the examiner for some of the cognitive paradigms that would be presented during the fNIRS protocol (e.g., a “+” on the screen = a 15 second silent fixation period before and after each individual speaking task). The participant then cleaned his/her forehead with an alcohol wipe and was fitted with the brain activation-monitoring headband on the forehead, which was secured with a soft stretchy cloth (to block out light and keep the band comfortably on his/her forehead). A 10-second baseline condition was then recorded as the participant passively watched an aquarium video on the computer (which was repeated prior to each of the three testing blocks). The participant then completed the three counterbalanced blocks of three to five speaking tasks each (45 seconds - 1 minute each), as summarized in Table 3 and discussed in detail in section 2.3.2. Two verbal fluency tasks (VFTs) were performed only after the first Everyday speech block, but these results were not included in the analysis of this study and are not discussed further in this manuscript. 2.3.1. Stimulus Delivery Visual stimuli were delivered via one of 12 different (counterbalanced) narrated Powerpoint presentations with standardized durations for each slide. The entire protocol was video and audio recorded for later transcription and analysis. 2.3.2. Speaking Blocks The 10-second silent resting baseline (watching a video of an aquarium with fish swimming slowly) was collected prior to each of the three conditions: (1) Everyday speech, (2) Prolonged Speech (PS), and (3) Metronome-timed speech (Met). The first condition completed was always the non-strategy Everyday condition for all participants. 34 The order of the second and third strategy conditions, Met and PS, were counterbalanced. Additionally, the three speaking Task Blocks summarized as tasks 1-3 in Table 3 were counterbalanced so that each Task Block was equally distributed across strategy/nonstrategy conditions. Each Task Block (Table 3) included three speaking tasks in this order: conversation, describing a complex procedure, and memorizing and telling a joke. Each condition had the three speaking tasks delivered in the same order but with differences in the specific question asked. For example, telling a memorized joke was always the last speaking task in each Task Block, but the participant was asked to memorize and retell a different joke for each block. Participants passively watched a cross (+) on the computer screen for 15 seconds prior to and following each speaking task within each block as a period of rest. Participants were assured that there were no right or wrong ways to perform the tasks. Following each condition, participants were prompted to stop using the speech strategy learned in the previous condition and were asked to have a brief conversation using their everyday speech prior to starting the next baseline to ensure any trained strategies were discontinued. After each speaking block (Everyday, PS, Met), participants briefly rated the effortfulness of speech production using a 9 point Speech Effort Self-Rating Scale (1 = very effortless, 9 = very effortful) from Ingham et al. (2006). The total time wearing the headband and completing the speaking tasks below was approximately 30 minutes depending on the initial adjustments required to ensure the headband sensors made secure contact with the participant’s forehead. 35 Two spontaneous discourse generation tasks (conversation and procedural description) were included because Tellis et al. (2011) found significant differences in blood flow as measured with DCS only when the speech task was of sufficient length (one minute or longer). The description task (e.g., the steps to plan a surprise party) was also selected to assess changes in PFC activation per Xu et al. (2005) and Cannizzaro et al. (2012). One speech stress task (telling a memorized joke) was included in all blocks because AWS who attend the Stuttering Support Group at UVM have subjectively reported difficulty with timing and delivery of jokes and there is a question on the OASES (Yaruss & Quesal, 2008) regarding “How difficult is it for you to communicate in the following social situations?” specific to joke-telling. The participants were provided one minute to memorize the joke and had 45 seconds to tell the joke to the examiner. The joke-telling task was expected to elicit the most stuttering, and the participants were cutoff after 45 seconds to increase the possibility that the participants did not finish the joke prior to the standardized recording time. 2.3.2.1. Non-strategy Condition (Everyday Speech) The non-strategy, or Everyday speech, condition was always administered first. The condition included the three speaking tasks from either Task Block 1, 2, or 3 (summarized in Table 3). 2.3.2.2. Fluency-Enhancing Strategy Conditions (Prolonged and Metronome-timed Speech) Two counterbalanced fluency-enhancing strategy conditions (PS and Met), were presented following the non-strategy condition. The examiner started a new experiment 36 for each strategy-use condition and collected another 10-second resting baseline. Participants were trained to correctly utilize each strategy immediately prior to each condition, and the examiner did not start the fNIRS recording until after the participant had acquired the trained speech strategy. The strategy-use conditions included the same three counterbalanced categories of speaking tasks (45 seconds - 1 minute each) from either Task Block 1, 2, or 3 as described in 2.3.2. (see Table 3), except the participants were prompted to speak with the trained fluency-enhancing strategy throughout the entire condition. For the Prolonged Speech (PS) condition training, participants viewed an exemplar video of an adolescent modeling prolonged speech from the Camperdown Program, a standardized PS program with evidence for efficacy in randomized control trials (e.g., O’Brian, Onslow, Cream & Packman, 2003). The exemplar video is freely available at: http://sydney.edu.au/health-sciences/asrc/videos/Adolescent_exemplar.mp4. The examiner trained the participant by having him/her watch, imitate and spontaneously produce the speech sample using a provided transcript at a rate of ~33 syllables per minute (spm; 75 syllables in 2 minutes 15 seconds; ~ 6 minutes of PS training). Participants were considered to have met the criteria to begin the PS strategy block once they imitate the model at a rate of no more than 37.5 spm (completing the target paragraph in ≥ 2 minutes). The examiner prompted the participants to try to maintain the PS strategy throughout the speaking tasks, and each narrated directions slide on the powerpoint provided a brief model of the correct PS speech pattern to cue continued use of the strategy. 37 For metronome-timed speech training, a model of how to speak in time with the 92 beats per minute auditory stimulus was presented via a narrated powerpoint. Participants then practiced speaking with the metronome while saying the days of the week and the months of the year. The trained examiner followed-up to ensure the strategy was acquired (one syllable per beat) prior to initiating the tasks. The metronome beat was played at all times during task completion to provide a consistent external timing cue. 2.3.3. Compensation and Follow-up Questionnaire Following the protocol, the examiner removed the headband from the participants. All participants then filled out a Compensation Form with his/her address to receive his/her $25 compensation. Only the AWS participants then answered the following additional questions about their stuttering in writing: List members of the immediate or extended family with a history of stuttering (N/A if not applicable; note if maternal or paternal and if the stuttering is persistent or recovered) Detail previous treatment history for stuttering (number of years of stuttering treatment) Where were these services provided? Which fluency techniques have you been taught and are you currently using? Have you participated in the Stuttering Support Group at UVM? (Describe) The AWS also rated their typical stuttering in specific daily life speaking situations using the following 1-10 scale: 1 = no stuttering and 10 = the most stuttering imaginable for anyone. Situations included: speaking to strangers, telling jokes, making 38 phone calls, conversation with strangers, conversation with a familiar person (family/friends), typical speech severity across the entire day, and most and least severe stuttering. The family history of stuttering for the TS group was acquired separately via post-testing communication. 2.4. Questionnaires The two standardized questionnaires used in this study are well-established and are given to every client who receives fluency treatment or attends a diagnostic at the Eleanor M. Luse Center clinic. The first was the Overall Assessment of the Speaker’s Experience of Stuttering (OASES)(Yaruss & Quesal, 2008), which takes approximately 15-20 minutes to complete. The OASES examines aspects and situations of stuttering and the emotions associated with those moments. It is divided into four sections including: general information, reactions to stuttering, communication in daily situations and quality of life. Each section contains 20-25 questions totaling 100 items overall. A rating scale is used from 1-5, which varies in each section corresponding to the area being measured and the question being asked. The total score for each of the four sections are added together to reveal an overall score indicating the impact of the speakers stuttering. The OASES has “strong” reliability and validity with Pearson product-moment correlations of test-retest impact scores ranging from 0.90 to 0.97 and 100% consistency in overall impact rating categories from test to retest (Yaruss & Quesal, 2006). The second questionnaire, the State-Trait Anxiety Inventory (STAI; Spielberger et al., 1983) took approximately 5 minutes to complete and was filled out by all participants in the research room just before the fNIRS headband was attached. The STAI measures 39 anxiety, feelings of apprehension, tension, nervousness, and worry in adults. The STAI specifically differentiates between the temporary condition of "state anxiety" and the more general and long-standing quality of "trait anxiety". Participants self-rated based on the following choices: 1 = “not at all”; 2 = “somewhat”; 3 = “moderately so”; 4 = “very much so.” Sample item: “I feel calm.” The state anxiety measure was collected to account for possible associations between anxiety, PFC activation and stuttering severity during the speaking tasks (e.g., Ezrati-Vinacour & Levin, 2004). STAI internal consistency alpha coefficients range from 0.86 to 0.95 and test-retest coefficients range from 0.65 to 0.75 over a two month testing interval (Spielberger et al., 1983). 2.5. Cognitive Screener The Montreal Cognitive Assessment (MoCA)(Nasreddine, 2010; Nasreddine et al., 2005) is a rapid 10-minute screening instrument for mild cognitive dysfunction. The cognitive domains assessed include: attention and concentration, executive function, memory, language, visuoconstructional skills, conceptual thinking, calculations and orientation. The total possible score is 30 points with 26 or above being considered within the normal range (average score for normal controls = 27.4). The MoCA has been validated for 55-85 year olds with strong sensitivity to distinguish normal controls from those with mild cognitive impairment. All participants were administered this screening as part of eligibility for the study. The test, reliability/validity and administration instructions are freely accessible for clinicians at www.mocatest.org. 40 2.6. Withdrawal Procedures Participants were informed that they may withdraw from the study at any time they wished without penalty or choose not to answer some questions on the questionnaires. All enrolled participants chose to continue with the protocol as described above. 41 CHAPTER 3: DATA ANALYSIS 3.1. Power Analysis A power analysis was conducted using G*Power 3.1.6 (Faul et al., 2009) to determine the number of participants needed in this study. The primary model was examined using an F test repeated measures design with within- and between-group interactions (2 groups and 3 measures). To achieve a power of 0.80 and a medium effect size at α = 0.05, a sample size of 14 was required per group. In previous work by Cannizzaro, Dumas, Prelock & Newhouse (2012), clear and robust differences were found in the neural activation related to conditions of narrative discourse processing with 12 subjects. A second G*Power analysis was conducted for independent t-tests for between-group comparisons. To achieve a power of 0.80 and a medium effect size at α = 0.05, a sample size of 10 was required per group. 3.2. fNIRS Data Processing Voxel locations were spatially registered using standardized locations acquired from Ayaz, et al. (2006; see Figures 4 & 5 for optode locations). Using modified BeerLambert Law (Cope & Delpy, 1988), changes in concentrations of oxygenated hemoglobin (HbO) relative to the initial 10-second resting baseline (which sets the activation level to zero) prior to each condition were extracted from the raw fNIR data (16 optodes x 2 wavelengths). The unit of measurement of concentration HbO change relative to baseline (zero) was micro molar units, or uM. Raw HbO data collected using COBI software (Ayaz & Onaral, 2005) were preprocessed and analyzed using fnirSoft software (Ayaz, 2010). HbO values at each voxel were manually evaluated to exclude 42 individual optode channels (if any) due to motion artifact, saturation or noise. Based on these criteria, channel #8 for AWS was removed for participant #2 in the everyday condition and channel #8 was removed for TS participant #1 in the prolonged speech condition (Table 4). For all data, a linear phase, low-pass FIR (finite impulse response) filter was applied with an order of 20 to attenuate the high frequency components of the signal as well as respiration and cardiac cycle effects (Ayaz, 2010). Mean changes in baseline-corrected concentration of HbO were calculated from the attenuation of the CW LEDs hemodynamic response across the three speaking conditions. Because the hemodynamic response evolves over 10-12 seconds after task onset (Buckner, 1998) and increases roughly linearly over time (Boynton, 1991), the first 10 seconds were removed from each 1-minute speaking task for all participants. All directions and + rest fixation periods were also removed from analysis to isolate the ~50 seconds of spontaneous conversation in each condition block. Mean voxel-wise HbO were calculated at each of the 16 channels for each participant in each condition block. Interpolated-bordered voxel-wise statistical HbO activation maps and real-time videos of the frontal view (at p < 0.05) were generated to visualize significant activation on a brain surface image (Ayaz et al., 2006). For each participant, the calculated mean HbO values for the conversation speaking task were grouped and averaged corresponding to left (channels 1-8) and right (channels 9-16) hemisphere measurements in the PFC (mean left and right activation change respectively). These mean HbO values per hemisphere of the averaged baselinecorrected HbO responses were used to calculate a Laterality Index, LI = (L-R)/(L+R), for 43 each participant and each condition (Everyday, PS and Met). The LI ranged from -1 to 1 with indexes of +0.15 to 1 arbitrarily defined as indicating left hemisphere dominance, -0.15 to +0.15 indicating bilaterality and -0.15 to -1 indicating right hemisphere dominance (Baicu, 2005; Bisconti, 2012). 3.3. Speech Sample Analyses There were three speaking tasks (conversation, description and joke) in each condition block (Everyday, PS and Met). Each speech sample was analyzed for percent syllables stuttered (%SS) and speech rate (syllables per minute; spm) and was later reliability-checked (see 4.1.3 and Table 6). A stuttering severity score was calculated using the Stuttering Severity Instrument- 4th Edition (SSI-4; Riley, 2009) based on the initial phone conversation screening and the three speech tasks from the Everyday speech portion of the research protocol. The SSI-4 measures frequency, duration and physical concomitants associated with stuttering for a total overall score. Descriptive statistics (%SS and spm) for each speaking condition are presented in Tables 5 & 7. Because the focus of the present study was to compare laterality differences during highly stuttered versus highly fluent speech, stuttering frequency (%SS) in each individual Everyday speaking task for the AWS group was calculated. The comparison determined whether all three speaking tasks should be collapsed or whether one task should be isolated for main analyses. The conversation, description and joke task %SS were all significantly positively correlated (see Table 5; Bivariate Pearson) at p < 0.01. AWS stuttered most on the joke (Mean = 10.46% SS; SD = 7.14), then the conversation (Mean = 9.11% SS; SD = 8.69) followed by the description task (Mean = 7.69% SS, SD 44 = 8.10). Paired samples t-tests indicated that Everyday conversation and description mean %SS were not statistically significantly different (p = 0.28), and conversation and joke were not significantly different (p = 0.33). However, Everyday description and joke %SS were significantly different at p = 0.02. Therefore, the description task was eliminated from further analyses because it elicited the least mean stuttering and was significantly different from the joke task. Even though the conversation %SS was not as high as the joke, it was not statistically significantly different. It was then determined that the ~1 minute conversation rather than the joke task in each condition would be the primary focus of data analysis (Everyday, PS, Met) because: (1) between-group mean speaking time during the joke telling task was significantly different (p = 0.001) for AWS (Mean = 40.86 secs., SD = 4.85) and TS (Mean = 31.71 secs., SD = 7.55) in the Everyday condition, such that the TS group told the joke much faster than the AWS group with subsequently less speaking/activation time for fNIRS analysis, (2) the between-group comparison of speaking time in the Everyday conversation task was not significantly different (AWS mean = 59.7 secs., SD = 1.07; TS mean = 59.9 secs., SD = 0.27; p = 0.39) and (3) the Everyday joke task was significantly (p < 0.001) shorter and more variable in time than the conversation task across all participants (conversation length Mean = 59.82 secs., SD = 0.77; joke length Mean = 36.29 secs., SD = 7.77). The conversation task also allowed for analysis of a longer, spontaneous, openended speech sample of more consistent duration across participants. In contrast, the joke task required memorizing a relatively short joke and repeating it, which was not 45 spontaneously generated speech. Subsequent fNIRS data analysis therefore focused on the conversational speaking task in each condition block (Everyday, PS, Met), and the description and joke tasks were eliminated from subsequent analyses. 3.4. Hypotheses-Driven Analyses Three 2 (group) x 3 (condition) Repeated Measures (RM) analyses using a linear mixed-effects model design, a conservative analysis method for detecting between conditions differences with small sample sizes were conducted. The term of interest was the group x condition interaction, and all analyses were completed with IBM SPSS Statistics software (IBM Corp., 2013). The three target dependent variables (DVs) were: (1) mean Left HbO in the left PFC channels during conversation (2) mean Right HbO in the right PFC channels during conversation and (3) a Laterality Index calculated from these mean Left and Right hemispheric HbO values (LI= (L-R)/(L+R); Sato et al., 2011). The significance criterion was Bonferroni corrected to p < 0.01 for all analyses. Following the linear mixed models design, five hypotheses-specific pairwise comparisons were conducted to determine differences in the three target DVs both within and between groups. For Hypothesis #1, pairwise comparison #1 analyzed between-group differences in the non-strategy conversational speaking task (Everyday AWS vs. Everyday TS). For Hypothesis #2, pairwise comparisons #2 & #3 analyzed within-group (AWS) differences in the Everyday speech condition relative to the fluency-enhanced speech conditions (2 = Everyday versus PS and 3 = Everyday versus Met). Pairwise comparisons 46 #4 & #5 analyzed between-group differences in the strategy conditions (4= PS AWS versus PS TS and 5 = Met AWS versus Met TS). Additional t-tests were performed to compare between-group (AWS versus TS) differences in speech rate, mean %SS per speaking task in each condition, overall SSI-4 score, self-reported speech effort, handedness, MOCA scores, age, STAI-State, STAI-Trait (see Table 4 for group and individual mean results). Post hoc ancillary analyses were performed on three participants per group (AWS and TS) who were removed from the primary analysis because they did not meet the stuttering severity criteria. See section 4.1 for a summary of this analysis. 47 CHAPTER 4: RESULTS 4.1. Descriptive Analyses 4.1.1. Participants Twenty-nine total participants (AWS n = 15, 2 female; TS n = 14, 2 female) completed the full protocol. As will be described in sections 4.1.2 and 4.1.6, one AWS participant did not pass the cognitive screener (not included in Table 4) and three other AWS did not meet the inclusion criteria for percent syllables stuttered (%SS) based on post-testing analyses. These participants and the matched controls were excluded from the main analyses for a total of 11 AWS (1 female; Mean age 33.2 years, SD = 10.8) and 11 TS (1 female; Mean age 32.8 years, SD = 11.2). See Table 4 for other participant demographics such as handedness, results of the MoCA screening and the OASES and STAI questionnaires, stuttering severity, treatment and family history, and medications. 4.1.2. Stuttering Severity To calculate stuttering severity scores with the SSI-4 (Riley, 2009), the phone conversation and three in-clinic speaking samples in the Everyday condition were transcribed for each participant. All TS met the inclusion criteria of ≤1% syllables stuttered (SS). Three of the AWS met the criteria to be entered into the study based on individual samples of > 2% SS, but their overall mean %SS was < 2% and their SSI-4 scores fell in the ≤ very mild severity category based on post-testing analyses. Because this study’s primary hypotheses revolve around change in lateralization patterns while stuttering and while highly fluent, these three participants and the matched TS were excluded from the main analyses. With these subjects removed (n = 11; 1 female), the 48 mean SSI-4 score for AWS was 26.1 (SD = 6.46), which corresponds to “moderate” severity. The TS group (n =11) SSI-4 severity score was 1.27 (SD = 1.0), which fell below the lowest severity category possible on the SSI-4 (< very mild). As another metric of severity, percent syllables stuttered (%SS) was also calculated for each individual speaking task (Table 5). Overall group mean %SS, based on the phone and three Everyday speech samples for each participant, was significantly different between the AWS (Mean = 12.1% SS, SD = 6.89) and TS groups (Mean = 0.13% SS, SD = 0.19; p = 0.000). The groups were also significantly different (Bonferroni corrected p < 0.01) in the Everyday conditions not including the phone call (AWS Mean = 11.26% SS, SD = 7.08; TS = 0.11% SS, SD = 0.17). Group %SS did not significantly differ (Bonferroni corrected p > 0.01) in the Met condition (AWS Mean = 0.32% SS, SD = 0.68; TS Mean = 0.00% SS, SD = 0.00) or the PS condition (AWS Mean = 0.32% SS, SD = 0.44; TS Mean = 0.00% SS, SD = 0.00) with all means falling below 1% SS. The significantly reduced stuttering <1% SS in the AWS group during both the PS and Met conditions indicated successful application of these strategies. The total %SS and SSI-4 scores were significantly positively correlated (rho = .95, p < 0.01), such that increases in stuttering severity measured by %SS were associated with a similar increase in SSI-4 scores. To further gauge stuttering severity, AWS participants self-rated their stuttering severity across different speaking situations in their daily lives (e.g., conversation with strangers, telling a joke, least and most stuttering) using a 1-10 scale in which 1= no stuttering and 10 = the most severe stuttering imaginable. See Table 8 for a summary of 49 these self-reported stuttering severity means, SDs and ranges for the AWS group. The mean typical SR for AWS (n = 11) was 4.18 (SD = 1.33) with a range from 3-7, which corresponds to mild-moderate stuttering severity. 4.1.3. Stuttering Severity Measurement Reliability Three of the 11 AWS participants were randomly selected to assess both intraand inter-rater measurement reliability. Intra-rater reliability of SSI-4 stuttering severity scores was carried out by the first author who reanalyzed the phone conversation and three conversation samples audio and video recorded in the clinic (mean absolute difference from original SSI-4 score/mean original SSI-4 score). Inter-rater reliability was completed by an experienced certified speech-language pathologist with a specialty in stuttering using the same samples. Reliability was conducted for (1) SSI-4 categorical severity ratings (e.g., very mild, mild, moderate, severe, very severe) and (2) SSI-4 numerical score (frequency or percent syllables of stuttering from two in-clinic samples, average duration of the three longest stuttering moments plus the physical concomitants subscale scores). Cohen’s Weighted Kappa was 0.80 for intra-rater reliability and 0.625 for interrater reliability (both of which are considered to be “good” strengths of agreement) for SSI-4 categorical severity ratings. Overall SSI-4 numerical score intra-rater agreement (mean absolute difference from original SSI-4 score/mean original SSI-4 score) was 94.8% (range 88.89 – 100%) and inter-rater agreement was 89.6% (range 82.6 – 97.3; see Table 6). Therefore, measurement reliability was good for categorization of participant stuttering severity. 50 4.1.4. Speech Rate Each speaking sample across all conditions and participants was analyzed for speech rate in syllables per minute (spm) to make group comparisons and to ensure the target speech rates were met in the strategy conditions (Table 7). The AWS group (n = 11; Mean = 170.8 spm, SD = 48.3) spoke significantly slower than the TS group (n = 11; Mean = 265.7 spm, SD = 26.9) in the Everyday speech condition (Bonferroni corrected p < 0.01). This was in part due to the AWS having stuttered speech during these samples which reduced the number of syllables spoken in the allotted time. The target speech rate for the Prolonged Speech condition was 37.5 spm or less (based on the video exemplar), and the metronome was set to 92 beats per minute. The groups did not significantly differ (p > 0.01) in mean speech rate in the PS condition (AWS Mean = 48.2 spm, SD = 8.5; TS Mean = 41.7 spm, SD = 9.7) or the Met condition (AWS Mean = 89.4 spm, SD = 10.4; TS Mean = 93.8 spm, SD = 2.7). The AWS group spoke significantly faster (p < 0.01) than the target rate in the PS condition. However, both the PS and Met strategies were effective in minimizing stuttering to near zero levels in both groups (see 4.1.2), indicating successful application of the strategies regardless of the target rate. 4.1.5. Speech Effort Self-Ratings The participants rated the effortfulness of speech production following each condition (Everyday, PS and Met) using the Speech Effort Self-Rating Scale in which 1 = very effortless and 9 = very effortful (see Table 9; Ingham et al., 2006). AWS reported significantly more speech effort in the Everyday condition (Mean = 3.27, SD = 1.85, p < 51 0.01) than the TS group (Mean = 1.45, SD = 0.69), suggesting that the AWS group’s habitual speech production was more effortful than TS. AWS and TS were not significantly (p > 0.01) different in self-reported speech effort for the PS (AWS Mean = 4.55, SD = 2.95; TS Mean = 5.09, SD = 2.12) or Met conditions (AWS mean = 3.91, SD = 2.21; TS Mean = 3.18, SD = 1.33). Paired sample t-tests indicated that the AWS group’s ratings of effortfulness did not significantly differ across speaking conditions (Bonferroni corrected p > 0.16), meaning that strategy-application did not significantly change perceived effortfulness of speech production. Everyday speech was the least effortful for AWS and TS, followed by the Met condition with the PS condition being the most effortful. While none of the AWS group’s effortfulness ratings were statistically significantly different between conditions, each of the TS within-group speech effort ratings was significantly different from the other (p < 0.016) across the three conditions. Speech effort in the PS and Met conditions was significantly correlated (rho = 0.54, p < 0.01) suggesting that the two strategy-application conditions were associated with similar amounts of self-rated effort for both groups. Further, speech effort in the Everyday condition was significantly correlated with both %SS (rho = .48, p < 0.05) and SSI-4 scores (p < 0.01), such that effortfulness increased with increasing stuttering severity in that condition. 4.1.6. Montreal Cognitive Assessment (MoCA; Nasreddine, 2010) As mentioned previously, one AWS participant scored 21 out of 30 on the Montreal Cognitive Assessment (MoCA), which was below the normal limits for adults 52 set at a score of 26 or above. His data were excluded from all analyses (not reported in Table 4 or elsewhere in this document). One other AWS (participant #7 on Table 4), a college student majoring in engineering, scored a 24 on the MoCA. This score falls in the upper range of what is considered “mild cognitive impairment”. However, research for this severity rating has not yet been established (www.mocatest.org/FAQ.asp), and the lower end of the normative range is a score of 25.2. This participant met all other inclusion criteria and was included in the experiment. The group means on the MoCA (n = 11 per group) did not significantly differ at p > 0.05 (AWS Mean = 27.82, SD = 1.72; TS Mean = 28.73, SD = 1.01). Scores on the MoCA were significantly positively correlated with age (rho = 0.49, p < 0.05) such that cognitive scores increased as participant age increased. 4.1.7. State-Trait Anxiety Inventory (STAI) The two groups (n = 11 each) did not significantly differ (p > 0.05) on the STAIState (AWS Mean = 43.4, SD = 4.5; TS Mean = 45.3, SD = 6.1) or the STAI-Trait (AWS Mean = 49.5, SD = 4.9; TS Mean = 50.8, SD = 8.9) standard scores. Both mean scores fell within the normal range for adults (see Table 4). The results suggest that the groups’ self-reported levels of anxiety just prior to being fitted with the fNIRS headband were comparable. 4.1.8. Overall Assessment of the Speaker’s Experience of Stuttering (OASES) Only the AWS group (n = 11) completed the OASES (Yaruss & Quesal, 2008), a questionnaire about the degree to which stuttering affects daily life. The group mean of 2.35 (SD = 0.44; see Table 4) corresponded to an overall impact rating of “moderate” as 53 the average of four sections (General Information, Your Reactions to Stuttering, Communication in Daily Situations, and Quality of Life). Impact scores ranged from a low of 1.72 to a maximum of 3.11. The STAI-State scores were significantly positively correlated with the OASES (rho = 0.71, p < 0.05), such that higher self-reported state anxiety prior to testing was associated with higher self-reported impact of stuttering on the AWS’s daily life. 4.1.9. Handedness All participants self-reported as right-handed and completed the Edinburgh Handedness Inventory--Short Form (modified from Oldfield, 1971 by Veale, 2013). A resulting Laterality Quotient from +60 to +100 indicated right-handedness, -60 to +60 indicated mixed handedness, and -60 to -100 indicated left-handedness. Mean group handedness (n = 11) did not significantly differ (AWS Mean = 95.45, SD = 8.43; TS Mean = 93.27, SD = 15.13; p > 0.05). One TS participant (#8 in Table 4) scored a +50 on the Edinburgh Handedness Inventory--Short Form (e.g., hand preference during writing, throwing, tooth brushing and using a spoon), suggesting mixed/right handedness. However, this participant verbally confirmed right-handed dominance overall in his daily life. He met all other inclusion criteria and was included in all analyses. 4.1.10. Family History of Stuttering Two of the AWS (n = 11) reported having an immediate family history of stuttering (e.g., sibling, parent), three AWS reported an extended family history of stuttering (e.g., uncle, grandparent), and six AWS reported no family history (see Table 4). One included TS (n = 11) reported an extended family history of stuttering and the 54 other 10 TS reported no family history of stuttering. The one TS who reported an immediate family history of stuttering was one of the three participants who were removed from the main data analysis (due to the matched AWS not achieving the %SS criteria). Family history of stuttering was positively correlated with %SS (rho = 0.49, p < 0.05) and SSI-4 scores (rho = 0.95, p < 0.01) such that stuttering severity was higher for those who reported having family members (immediate or extended) who stutter or stuttered. 4.1.11. Years of Treatment The AWS group (n = 11) presented with a mean of 8.2 years of previous treatment (SD = 8.50, Range 2-30; see Table 4). Participants specified a number of treatment approaches previously learned including: prolonged speech, fluency-shaping (easy onsets, light contacts, flexible rate, pauses), stuttering modification (pull-outs, voluntary stuttering), proprioception and relaxation and breathing strategies. 4.1.12. Medications Two of the included AWS participants (n =11) reported regular medication use: participant #5 Wellbutrin and #10 insulin for Type I Diabetes. Two of the included TS participants (n = 11) reported current medication use: #4 Citalopram (20 mg/day) for mild seasonal affective disorder and #10 Zoloft. 55 4.2. Hypotheses-Driven Analyses of Lateralization 4.2.1. Hypothesis 1: Between-group (AWS vs. TS) Differences in Laterality of Activation During Conversation in the Everyday (Non-strategy) Condition Results of pairwise comparison #1 following the RM linear mixed model analysis indicated no significant difference (p > 0.01) between AWS and TS on the measured variable of hemispheric laterality index (LI which varied continuously from -1 or pure right hemisphere dominance to +1 or pure left hemisphere dominance) in the Everyday conversation condition. Both groups presented with bilateral HbO in the Everyday condition with AWS being slightly rightward (Mean = -0.009) and TS being more leftward (Mean= 0.02, SE = 0.02; see Table 10 and Figures 8-10). Six of the 11 AWS (55%) and five of the 11 TS (45%) presented with rightward/bilateral LI (below zero; see Table 12 and Figure 10). For mean activation (HbO) levels in the left and right hemispheres in the Everyday condition, AWS presented with positive activation relative to baseline in both hemispheres (Left Mean = 1.97; Right Mean = 2.13, SE = 0.68). Results indicate bilateral PFC activation which was non-significantly higher in the right hemisphere than the left for AWS during stuttered conversational speech. Both right and left hemisphere mean activation was significantly different from baseline (zero) for the AWS in the Everyday condition (see Table 10 and Figures 6-7). The TS group presented with less activation (non-significant at p > 0.01) in both hemispheres relative to the AWS group in Everyday conversation. TS’s left hemisphere HbO (Mean = 1.57, SE = 0.68) was significantly higher than baseline (zero), while right 56 hemisphere activation (Mean = 1.25, SE = 0.68) was not significantly different from baseline in Everyday conversation. The results indicate greater leftward activation relative to rightward activation in the control group when speaking normally. Neither of the between-group differences in hemispheric activation in the Everyday conversation task reached significance (p > 0.01; see Table 10 and Figures 6-10). 4.2.2. Hypothesis 2a: Within-group Shifts in Laterality from the Non-strategy-use (Everyday) Condition to the Strategy-use Conditions (Prolonged and Metronome-timed Speech) Results of pairwise comparisons #2 & 3 following the RM linear mixed model analysis indicated no statistically significant (p > 0.01) within-group differences in laterality of the AWS group measured by LI and left/right mean HbO from the (a) Everyday to PS condition and the (b) Everyday to Met condition (see Table 11 and Figures 6-12). AWS presented with a statistically non-significant leftward shift in LI in the Met (Mean = 0.002, SE = 0.01) and PS (Mean = 0.004; SE = 0.02) speech strategy conditions relative to Everyday speech (Mean = -0.009; SE = 0.02). All values indicated bilateral activation across all conditions (strategy-use and non-strategy-use) for AWS. While six of the 11 (55%) AWS presented with LIs below zero (indicating rightward/bilateral activation) in the Everyday condition, four were below zero in the PS condition (36%) and five were below zero in the Met condition (45%; see Table 12). Eight of the 11 AWS (73%) presented with a non-significant leftward shift in LI in the PS condition relative to the Everyday condition, while six AWS (55%) had a leftward LI shift in the Met condition relative to Everyday LI (Table 13). 57 In terms of shift in hemispheric laterality of PFC HbO, AWS presented with a statistically non-significant decrease in mean left HbO while speaking with trained strategies (Left Mean PS = 1.42, SE = 0.54; Left Mean Met = 1.58, SE = 0.72) relative to Everyday speech (Mean left = 1.97, SE = 0.68; Table 10). AWS presented with a decrease in mean right HbO in the PS (Mean = 1.32, SE = 0.51) and Met (Mean = 1.49, SE = 0.68) conditions relative to the Everyday condition (Mean right = 2.13, SE = 0.68; Table 10). The results indicate non-significantly lower overall PFC activation in AWS during strategy-use in both hemispheres (even with self-reported increase in speech effort) and non-significantly more leftward/bilateral activation shift while speaking using these fluency-enhancing strategies. None of the AWS within-group differences in hemispheric activation reached significance (p > 0.01) or were significantly different from baseline (Table 11). Within the TS group, there was a statistically significant (p < 0.01) greater mean left HbO level in the Everyday condition (Mean = 1.57) relative to the PS condition HbO which was negative relative to baseline (Mean = -0.27; SE = 0.46; Mean diff. = -1.84, SE = 0.46; see Tables 10 & 11 and Figures 6-12 & 14). The TS group also presented with a statistically significant decrease in right HbO from the Everyday relative to the PS (Mean diff. = -1.85, SE = 0.48) and Met (Mean diff. = -1.72, SE = 0.53; Table 11) conditions. The results suggest decreased bilateral PFC activation for TS when applying newly learned speech strategies relative to speaking normally. All TS group LI values indicated bilateral activation across all conditions (Everyday, PS, Met), which was non-significantly in the leftward in direction (Everyday 58 Mean = 0.02; PS Mean = 0.016; Met Mean = 0.05, SE = 0.01; see Tables 10 & 12 and Figures 10-11). Individual participant TS #8 presented with a LI of 0.22 in the PS condition indicating leftward dominance (> 0.15 cutoff; Figure 11). Six of the 11 TS (55%) presented with leftward/bilateral LIs (above zero) in the Everyday condition, four had LIs above zero in the PS condition (36%) and ten presented with a leftward LI in the Met condition (91%; see Figures 10-12 and Table 12). Four of the 11 TS (36%) presented with a non-significant leftward shift in LI in the PS condition relative to the Everyday condition LI, while eight TS (73%) had a leftward LI shift in the Met condition relative to Everyday LI (see Table 13). Thus, while none of the within-group measures of AWS’s left and right HbO activation across tasks was statistically significantly different across condition blocks (strategy-use vs. non-strategy-use), the TS group presented with significantly greater left HbO in the Everyday condition relative to the PS condition and significantly greater right HbO in the Everyday condition relative to both strategy-use conditions (PS and Met; see Table 11). 4.2.3. Hypothesis 2b: Between-group (AWS vs. TS) Laterality Differences During Strategy-use in the Prolonged Speech and Metronome Conditions Results of pairwise comparisons #4 & 5 following the RM linear mixed model analysis indicated a significant (p < 0.01) between-group difference in LI in the Met condition, such that TS presented with significantly greater leftward activation (Mean = 0.047) relative to AWS (Mean = 0.002; SE = 0.01; see Table 10). The only LI score that significantly differed from zero (no laterality) was the TS group’s mean LI in the Met 59 condition, which was in a leftward direction but did not surpass the +0.15 threshold for leftward dominance. LI was not statistically different (p > 0.01) between-groups in the PS condition (AWS Mean = 0.004; TS Mean = 0.016, SE = 0.02) with the TS group being non-significantly slightly more leftward in laterality (Table 10). In terms of left and right hemisphere HbO, AWS presented with statistically significantly (p < 0.01) greater right HbO activation (Mean = 1.32, SE = 0.51) relative to TS (Mean = -0.60) in the PS condition (see Table 10 and Figures 7 & 11). Betweengroup differences in right HbO when talking with the metronome were non-significant, with greater right activation in AWS (Mean = 1.49) relative to TS (Mean = -0.48, SE = 0.68; p = 0.06; see Table 10 and Figures 7 & 12). There were no significant betweengroup differences (p < 0.01) in mean left HbO across speaking conditions. However, there was a trend towards less left HbO of TS in the PS condition (Mean = -0.27, SE = 0.54) relative to AWS (Mean = 1.42, p = 0.04; see Table 10 and Figure 6). Both groups presented with bilateral LIs across all conditions, being significantly more leftward when talking with the metronome in the TS group relative to AWS (Table 10 and Figures 8 & 9). However, only the TS group presented with significant withingroup differences in hemispheric activation when speaking with trained strategies relative to talking normally (Table 11). The Met condition brought about the most leftward LIs for the TS group (with only one subject being below zero; see Figure 12). AWS activation levels were not significantly different when they were stuttering compared to when speaking fluently (Table 11). 60 Overall, AWS presented with relatively higher mean bilateral activation than TS in all conditions. Between-group differences in activation were significant in the PS condition (AWS greater in the right hemisphere) and for LI in the Met condition (TS more leftward). Left and right hemisphere HbO was statistically different from zero (baseline) in all three speaking conditions for AWS, while controls had activation that was significantly different from baseline only in the left hemisphere during Everyday conversation (Table 10). The relatively higher mean left and right PFC activation in adults with persistent stuttering compared to controls across all conditions suggests more cognitive effort when speaking normally as well as when using fluency-enhancing speech strategies. 4.3. Ancillary Analyses of AWS and TS (n = 3 each) Removed from Main Comparisons Three of the original 14 participants per group were removed from the analyses due to the AWS participants not meeting the stuttering severity criteria and scoring ≤ “very mild” on the SSI-4. Ancillary linear mixed model analyses were performed to compare the three removed AWS (1 female) and three matched TS (1 female) to determine if left/right PFC HbO differed between these groups. Results indicated that these “very mild” AWS presented with laterality which was not significantly different from their matched controls (n = 3) across all comparisons, measures and conditions (See Tables 14-16 and Figures 19-23). However, the ancillary comparison (n = 3 versus n = 11 of the main analyses) was underpowered and may not accurately reflect group differences. 61 Specifically, HbO activation levels in the left and right hemispheres and Laterality Indices (LIs) in the three speaking conditions did not differ in the within- and betweengroup comparisons across the three conditions: Everyday, PS and Met. The three AWS presented with non-significantly higher mean HbO overall in both left (Mean Everyday = 3.03, SE = 1.87; Mean PS = 1.17, SE = 1.35; Mean Met = 1.55, SE = 1.49) and right hemispheres (Mean Everyday 1.90, SE = 1.35; Mean PS = 1.05, SE = 1.08; Mean Met = 0.93, SE = 1.47) relative to TS (Table 15), similar to the full sample comparisons (n = 11). However, the AWS presented with a non-significantly more leftward LI (Mean = 0.048, SE = 0.029) relative to TS (Mean = 0.038, SE = 0.029) in the Everyday condition, while the full (n =11) sample Everyday LI was more rightward (Mean = - 0.009). The results suggest that these “very mild” AWS have PFC lateralization patterns which are more similar to TS (leftward) than more severe AWS when speaking normally and when using speech strategies. Further ancillary analyses were conducted by combining the three removed participants with the other 11 per group for a linear mixed model comparison of all subjects (n = 14). In the combined samples, LI in the Met condition was significantly more leftward for TS (Mean = 0.045; p < 0.01) relative to AWS (Mean = 0.007; SE = 0.008), which was a similar result to the main (n = 11) analysis described previously. Also consistent with the main results, mean left HbO was significantly greater in the Everyday condition relative to the PS condition in the combined TS group (Mean Difference = 1.86; SE = 0.52; p < 0.01). 62 There were no significant between-group differences in mean left HbO or withingroup differences for AWS (n = 14) alone. For mean right HbO, TS presented with significantly higher right activation in everyday speech relative to PS (Mean Difference = 1.93; SE = 0.47; p < 0.01) and the Met condition (Mean Difference = 1.75, SE = 0.51; p < 0.01). A significant between-group difference was observed for the PS condition, with AWS presenting with greater mean right HbO than TS (Mean Difference = 1.86, SE = 0.63; p < 0.01). None of the other between- or within-group comparisons were significant. All of the combined n = 14 results that reached significance (p < 0.01) were consistent with the n = 11 main analyses across groups and conditions. 63 CHAPTER 5: DISCUSSION The purpose of this study was to investigate possible between-group (AWS versus TS) and within-group differences in PFC oxygenation in everyday stuttered conversation compared to fluency-enhanced speech using prolonged and metronome-timed speech strategies to achieve significantly less stuttering. In general, these comparisons are relevant to a comprehensive understanding how hemispheric laterality is associated with persistent developmental stuttering. Examining laterality shifts when applying wellknown speech strategies, especially prolonged speech which is used in real-world fluency therapy, is relevant to an understanding more about the underlying neural changes which occur in association with strategy-application in treatment. Specifically, the results add to the growing literature attempting to parse out whether AWS present with atypical right-sided lateralization relative to TS and whether such lateralization is causal or compensatory (e.g., Chang et al., 2011; Lu et al., 2010ab). Sowman et al. (2014) provided robust evidence which suggested the lateralization in AWS is compensatory because atypical laterality was not observed in the inferior frontal, temporal and parietal regions of interest in young preschool-aged CWS relative to matched controls. However, the authors added the caveat that many of the preschool-age participants would recover from their stuttering and may not have the atypical laterality observed in AWS whose stuttering persists long-term. Chang et al. (2008) provided additional evidence against the causal link, reporting no differences in laterality between 9-12 year old boys who stutter and their matched controls. The authors further demonstrated that laterality of recovered stutterers was closer to persistent stutterers than 64 controls, suggesting that the Sowman (2014) data represented the “true” laterality of the CWS. In contrast, Sato et al. (2011) provided evidence for right-lateralized auditory processing in preschool-age CWS which suggests that atypical neural activation may be present at early stages of stuttering. Tellis et al. (2011) further found atypical righthemisphere laterality in the PFC during a monologue task in an AWS as measured by DCS. The present study aimed to provide more information regarding laterality differences in AWS after years of persistent stuttering relative to controls and to identify how neural activation changes, if at all, when fluency is enhanced using speech strategies known to reduce stuttering to near-zero levels. The advantages of this protocol over most previous neuroimaging studies with AWS are that the subjects spoke freely for a sustained amount of time (~1 minute per task) with naturally stuttered and fluencyenhanced speech in a clinical context similar to that of therapy. The current study resulted in three main hypotheses-driven findings to be discussed below, followed by a brief general discussion, caveats and conclusions. 5.1. Main Finding #1: Non-significant Between-Group Differences in Laterality During Everyday Speech The first between-group main finding indicated that the AWS and TS groups (n = 11 each) did not significantly differ in laterality of activation in the PFC in a 1-minute everyday conversation, rejecting Hypothesis #1. Both groups presented with bilateral PFC activation while speaking normally, including naturally-occurring stutters for the AWS group. The AWS presented with non-significantly higher activation relative to 65 typical speakers in both PFC hemispheres, more so in the right hemisphere, while speaking normally. However, both the right and left HbO in AWS was significantly different from baseline (zero), while only the left hemisphere was significantly different for typical speakers. This discrepancy in overall activation may be due to the fact that the AWS group reported everyday speech to be significantly more effortful than typical speakers using the Speech Effort Self-Rating Scale (Ingham et al., 2006). The results support previous evidence that speech production is more effortful in adults with persistent stuttering (e.g., Chang & Zhu, 2013). The greater subsequent overall bilateral activation in the PFC, an area highly interconnected with the speech motor control circuit (BG and SMA), during stuttered speech may reflect relatively inefficient motor control in adults with persistent stuttering. The results may be related to AWS’s greater difficulties with successful initiation of motor programs in the extended BGTC circuit and relative overactivation in the speech motor timing network of AWS (e.g., Chang & Zhu, 2013; Max & Gracco, 2005) as previously discussed. The overall activation results also suggest a possible new interpretation to the compensatory hypothesis, in which AWS have been shown to recruit more of the right hemisphere to compensate for a less efficient speech production circuit in the left hemisphere. Instead, the results suggest that AWS may not have a right hemispherespecific switch during stuttered speech, at least in the PFC, but rather an overall increase bilaterally in activation across a wider network of prefrontal regions. In other words, the adults who stutter are recruiting more of their PFC in order to produce speech (whether 66 fluent or disfluent) than typical speakers, which is then measured in brain imaging studies as relatively greater right hemisphere activation. These group differences in and prefrontal activation seem to reflect group differences in the degree of effortfulness and cognitive resources required for speech production. The speech of AWS is more effortful than for TS, both when speaking normally and when applying speech strategies to be highly fluent. As an interesting ancillary analysis, the three AWS who were removed from group comparisons because of their “very mild” stuttering severity, had comparable activation patterns and were even slightly (not significantly) more leftward in laterality than the matched controls. This subgroup’s activation patterns suggest a possible association between their high levels of spontaneous fluency and leftward laterality in conversation relative to the AWS with more severe stuttering. Even though this AWS subgroup presented with persistent stuttering, these AWS had PFC activation which was more similar to typical speakers, which may be associated with their naturally higher levels of fluency. It is also possible that the PFC activation changes found in this study are not fully reflecting lateralization differences in key areas found to be different in young children who stutter (CWS). For example, the auditory cortex was shown to be more rightward lateralized during phonemic processing in preschool-age CWS by Sato et al. (2011), which was posited to reflect compensatory activation for a less efficient connections between left hemisphere language and auditory areas. Unlike the Sato et al. (2011) results, the present study (n = 11) did not find a significant correlation between stuttering 67 severity and atypical lateralization of PFC activation (except for the n = 3 ancillary analyses of < very mild AWS which was underpowered). However, the Sato et al. (2011) findings reflect differences in speech processing while the present results reflect lateralization during speech production. Given the preliminary nature of the present results using a relatively new technology, clearly more research is needed to clarify the findings. 5.2. Main Finding #2: Within-group Differences in Laterality from Everyday to Prolonged Speech and Metronome Conditions The second main finding indicated that PFC laterality did not significantly shift when AWS were trained in fluency-enhancing PS and metronome-timed speech, rejecting Hypothesis #2a. All AWS achieved near-fluent speech when using the trained strategies, and there was a slight leftward shift in laterality index in the predicted direction which was not significant. Overall activation in the left and right PFC of AWS was relatively reduced when speaking fluently compared to stuttered everyday speech. This effect was observed even though reported speech effort was greater when applying the strategies relative to typical conversation. The attenuated PFC activation is consistent with previous findings comparing metronome-timed speech relative to habitual speech (Braun et al., 1997) with a reduction in the need for PFC control of motor performance and enhanced sensory (auditory) processing. This may be due to improved speech/language monitoring in AWS as a result of the slowed output of production during metronome-timed and prolonged speech which enhanced auditory feedback and increased processing and motor planning time. The lack of significant change in 68 activation when speaking fluently versus stuttering may suggest trait differences in PFC activation in AWS during speech production regardless of fluency levels (Braun et al., 1997). Main finding #2 suggests that the strategies that elicited fluent speech were not associated with leftward laterality changes in the PFC for AWS as predicted, but they may have been associated with reduced effortfulness and increased automaticity of speech production as reflected by a bilateral drop in PFC activation. The majority of the AWS (9 out of the 11) had previous extended experience with the PS or other fluencyshaping strategies in treatment, which are relatively simple to acquire techniques conceptually. These factors may have allowed them to acquire and successfully use that strategy in the testing situation with relative ease. Further, the metronome’s external timing cue is reported to support fluent speech production by compensating for a less efficient internal timing cue in AWS, feasibly reducing effortfulness by reducing demands on motor control and changing linguistic stress patterns, even though the resulting speech is relatively unnatural. Both strategies slow speech down significantly and allow for greater preparation time for speech production, leading to improved sensorimotor integration which may have accounted for the significant drop in overall activation in AWS, especially in the PS condition. The metronome and PS strategies resulted in different activation patterns for the typical speakers (TS) even though both were newly-learned speech patterns. For both strategies, TS presented with relatively less activation in both hemispheres compared to speaking normally (see Figures 6-7). When TS spoke normally, they had a 69 bilateral/leftward laterality index which shifted (non-significantly) slightly rightward in the PS condition and significantly more leftward when talking with the metronome (Figures 8 & 9). Mean left and right PFC activation were actually negative relative to baseline during the PS task (Figures 6-7 & 14), suggesting reduced oxygenation overall in the PFC when using that strategy. Talking in time with the metronome was associated with negative right hemisphere activation and positive left hemisphere activation for TS (Figures 6-7 & 15), leading to a laterality index which was significantly leftward, more so than any other condition (Figure 8). All but one TS participant had leftward laterality in the metronome condition whereas the majority of TS (6 of the 11) had rightward laterality in the PS condition (Table 12). It is unclear what may be associated with the differential effects of the two trained speech strategies on PFC activation in TS. The PS strategy was reported to be the most effortful for the TS group, and the PFC is typically recruited when learning new motor patterns or when engaged in cognitively challenging tasks (e.g., Pike & Petrides, 2001), so it is counterintuitive that oxygenation was negative relative to resting baseline in both hemispheres during this task. It is possible that activation shifted to other posterior sensorimotor and auditory processing regions of the brain outside the PFC not specifically measured in this study (e.g., anterior frontal operculum, premotor cortex, SMA, Broca’s, Wernicke’s, posterior insula, ACC, primary auditory cortex, and/or cerebellar lobes), while the TS group exerted effort to learn and apply this new and unnatural speaking pattern (e.g., Braun et al., 1997). 70 Further, the metronome-timed speech rate was faster than in the PS condition by approximately 55 syllables per minute for the typical speakers. Perhaps the metronome speech approximated normal speech more than the significantly stretched out prolonged speech, and this factored in to the more leftward activation in the metronome condition, which was similar to everyday speech in TS. The external pacing cue of metronometimed speech may have altered the need for self-monitoring of speech similarly for TS and AWS, which may explain the relatively higher bilateral activation in the PS condition compared to speaking in time with the metronome. 5.3. Main Finding #3: Between-Group Differences in Laterality During Speech Strategy-Application (Prolonged and Metronome Speech) Hypothesis #2b predicted that the fluency strategies would “normalize” PFC activation and the between-group (AWS versus TS) lateralization patterns would be similar. The third main finding partially supported this prediction such that group Laterality Indices (LI) were not significantly different in the PS condition. However, between-group LI was significantly different in the metronome condition, with the TS group being more leftward than the AWS. Further, Hypothesis #2b was not confirmed in that the results did not represent a shift in the AWS group towards the TS group, since the LIs also did not significantly differ in the everyday condition. Additionally, AWS presented with significantly greater right HbO in the PS strategy condition relative to TS. Between-group left HbO was not significantly different in either strategy condition, and right HbO did not differ between groups in the metronome condition. Group laterality differences were even more apparent in the 71 metronome condition, with AWS maintaining more bilateral activation while the typical speakers shifted leftward. Overall activation was consistently greater in AWS relative to TS across all conditions. The results suggest that AWS have relatively greater bilateral PFC activation in general, especially the right side, compared to TS following short-term training to use speech strategies. The results provide supporting evidence that suggest AWS and TS have fundamental, trait differences in laterality of PFC activation during speech production. 5.4. General Discussion One finding of general relevance to the purpose of the present study is that adults with developmental stuttering have elevated, but not significantly different PFC activation than typical speakers when speaking normally. While we expected group laterality to differ only in everyday conversation (i.e., AWS being rightward and TS leftward) and be more alike when speaking fluently (i.e., both shift more bilateral/leftward), unexpected results were found. Specifically, the AWS group did not have a noticeable leftward change in PFC activation when using strategies compared to when speaking normally. Instead, they maintained a stable bilateral HbO activation level which was consistently higher than TS in all conditions. In contrast, TS presented with consistently leftward/bilateral activation across conditions, with the metronome condition being the most leftward. Below, eight possible accounts for the results are discussed. Individual differences. One possible interpretation of present findings is that there were appreciable individual differences in the degree of laterality as a result of learning the speech strategies. In other words, differences between LI may be associated with high 72 within- and between-group variability such that one participant may have had high relative activation in both hemispheres which was “washed out” when converted into the LI (see Tables 10 & 12 and Figures 10-12 and 16-17). For this reason, the mean left and right HbO activation were also analyzed to provide more information about the overall activation patterns measured. Each speech strategy affected individual participants differently as well. For example, AWS participant #7’s LI became less rightward during prolonged speech, but shifted to be the most leftward AWS during the metronome condition (see Figures 10-12, 18), suggesting that the two strategies could have differential effects depending on the participant. Inter-individual differences. It is possible that those AWS who had previously learned and utilized the PS strategy in previous treatment found it easier to acquire. Therefore, it may have required less mental effort overall for these AWS relative to the TS group and the other AWS who were never exposed to the PS strategy. Many of the participants verbally expressed the difficulty and effortfulness speaking with the two trained speech strategies, as was reflected by the higher speech effort ratings for both groups in the PS and metronome conditions. Nine of the 11 AWS participants had previously been trained to use fluency-shaping, or PS, strategies whereas none of the TS had this previous exposure. It is therefore possible that the laterality was not comparable associated across conditions for all participants based on previous experience using the PS strategy. Intra-individual Differences. Variable shifts in activation levels were observed for individuals across conditions. For example, participant #2 in the AWS group (see Figures 73 10-12 & 16) made a noticeable leftward shift in laterality when speaking with the PS and metronome strategies relative to his rightward activation in everyday conversation. This participant reported using fluency-shaping similar to PS in three years of treatment, which may have facilitated acquiring the PS pattern and therefore affected brain activation patterns more so than someone who was learning that strategy for the first time. That being said, AWS participant #5 (who reported 9 years of therapy including PS) had rightward/bilateral activation across all three speaking conditions, being even more rightward during PS (Figures 10-12). Similar variability was observed in the control group, such that TS participants 6 and 7 (see Figures 10-12 & 18) shifted from leftward in the Everyday condition to rightward in the PS condition and back leftward for the Metronome condition (Table 13). TS participant #9 had a comparable LI with the most rightward AWS in the Everyday condition, which shifted leftward in the PS and metronome conditions. Thus, inter- and intra-individual variability makes generalization of the collapsed group results difficult. Between-group differences in prior exposure to testing setting. We posit that the both groups responded similarly to the novel testing conditions. Although some of the AWS participants had been to the clinic before to attend Stuttering Support Group, the testing took place in an unfamiliar lab in an unfamiliar part of the building separate from the treatment rooms. Further, the STAI-State and Trait scores were not significantly different between groups, so pre-testing anxiety based on this self-report measure did not seem to indicate differences in participants’ reaction to the unfamiliar testing environment. While nine of the AWS reported previous exposure to the PS strategy or 74 fluency-shaping techniques of some kind, none of the participants reported previous experience speaking in time with the metronome. Within-group differences in stuttering severity. It is also possible that discrepancies in findings are associated with within-group differences in stuttering severity. Specifically, more severe AWS may present with stronger rightward laterality, while milder AWS present with laterality which is more similar to TS as was found in the ancillary analyses of the present study. However, the two “very severe” AWS (#4 and 9) were different in that #4 was relatively rightward in everyday and Met conditions, while shifting leftward in the PS condition (more bilateral activation; Figures 10-12). In contrast, #9 started with leftward laterality in everyday conversation and shifted to a slightly rightward bilateral LI when speaking fluently in the PS and Met conditions. Given the high variability in LI scores across conditions, it seems important for future studies using this technology to make subject-by-subject as well as group comparisons to account for the within-group variability across severity levels. Training duration. In other brain imaging studies measuring change in activation patterns following PS therapy (e.g., Giraud et al., 2008; Neumann et al., 2005), the amount of strategy practice was considerably more than in the present experiment. The participants received about six minutes of practice with the PS strategy in the current protocol, whereas the other studies made pre- and post-treatment measures after months or even a year of training and practice in multiple settings. Therefore, the lack of a significant shift in this experiment may be a result of limited strategy practice in a relatively short timeframe. 75 Strategy Mechanism Differences. While both metronome-timed and prolonged speech bring about fluency, they appear to do so in different ways. This is evidenced by the high variability in individual participants’ activation responses, with shifting both leftward and rightward relative to everyday speech. The only consistency was observed in the TS group for the metronome condition which was consistently leftward. The PS condition resulted in negative bilateral activation for the TS group, which may have been due to reallocation of activation outside the PFC as mentioned previously and is not possible to confirm. While both strategies significantly slowed speech (target of 33 spm for PS and 92 spm for the metronome condition), the differences in the metronome for TS might be due to the rhythmic production and its closer approximation to normal speech than the extremely stretched out prolonged speech. Speech-Related Anxiety. While the pre-testing state anxiety scores from the STAI did not differ between AWS and TS, it is feasible that there were individual and group differences in speech-related anxiety specific to the conversation tasks (e.g., Davidson et al., 2000). To assess these possible differences, concurrent measures of emotional reactivity or physiological responding such as galvanic skin response could have been collected throughout the protocol. Each participant could also self-rate his/her speechrelated anxiety after each condition block using a scale similar to the Speech Effort scale (1 = zero speech-related anxiety and 9 = the worst imaginable speech-related anxiety). The combined physiological and self-report information would have allowed for more detailed comparisons to account for the possibility that anxiety levels specific to speaking could vary widely among participants and subsequently affect PFC activation levels. 76 5.5. Caveats Sample size was a limiting factor in this study after the three AWS participants with very mild stuttering (< 2%SS) were removed from the sample for a total of 11 participants in each group. Representation of females who stutter was also a factor as there was only one female per group included in the final analyses which makes generalization of results to both genders difficult. Testing fatigue and differences in discomfort wearing the headband were also factors. We minimized the number of tasks to limit the time wearing the headband to approximately 30 minutes, but some participants reported feeling discomfort by the end of testing if it took longer to ensure proper skin contact of the headband during the initial set-up. The extended wear time may have interfered with some participants’ cognitive performance and attention to the tasks later in the session. However, the conversation task used for the present analysis was the first task completed in each condition block, making it is unlikely that fatigue interfered with measured laterality. Further, the conditions were counterbalanced to account for possible testing fatigue effects when learning and applying the fluency strategies. It is possible that the short-term training time to learn the strategies (~6 minutes for prolonged speech and ~3 minutes for metronome-timed speech) was not enough to elicit significantly different neural activation patterns. Future studies could examine PFC activation change pre- and post-treatment following a longer course of prolonged speech training and practice in multiple real-world contexts. This follow-up could reflect the laterality differences found in fMRI studies after extended use of fluency-enhancing 77 speech strategies (e.g., Neumann et al., 2003). Finally, rather than averaging all 8 channels on each side, voxel-wise comparisons in target regions using tasks known to elicit particular PFC regions, such as the dlPFC during verbal fluency tasks, would provide interesting information about differences in specific lateralization patterns during executive tasks in AWS and TS. 78 CHAPTER 6: CONCLUSIONS The present study is the first to use fNIRS technology to measure real-time changes in activation in the PFC while adults who do and do not stutter speak normally and with fluency-enhancing strategies. The results are unique in that they revealed between- and within-group differences in activation when speaking normally in conversation, while most other brain imaging studies have been limited to one word responses, which are typically not stuttered. Present results suggest an alternate interpretation to the compensatory laterality hypothesis, such that there may be compensatory relative overactivation of the PFC during speech production in AWS, observed in both the left and right hemispheres. While an interesting finding, the answer to the questions about whether there are lateralization differences in adults with persistent developmental stuttering and if this reflects causal or compensatory activation established early in development remains unclear. The laterality measures collected in the current study focused on the PFC, which was considered a reflection of hemispheric activation during speech production in adjacent and interconnected temporal, parietal and cerebellar regions involved in speech motor control. The relatively greater overall bilateral activation in AWS compared to typical speakers, and activation levels consistently significantly above zero, suggests greater effortfulness and reduced automaticity of speech production when stuttering and, unexpectedly, when speaking fluently. AWS seem to need to recruit more PFC neural activation in order to produce speech during any form of speech production. The resulting bilateral activation patterns are reflected by relatively greater right activation than typical 79 speakers, which is consistent with fMRI studies examining between-group differences in laterality. The present study also demonstrated the practical usefulness of fNIRS technology to measure laterality differences in real-time in a naturalistic speaking environment. Future studies utilizing this technology may track change before, during and after achieving significantly greater fluency levels across many speaking contexts, to shed more light on the ongoing compensation versus causal laterality hypotheses. It will be necessary to examine laterality differences in younger children closer to the onset of stuttering (preschool-age), similar to Sowman et al. (2014) and Sato et al. (2011) but in connected speech as was done in the present study. Whole-head fNIRS neuroimaging would be ideal to measure activation levels in the PFC as well as in the temporal lobes where key regions are activated during speech and language tasks. All analyses should compare individual and group results, as there is a high degree of variability among all participants. These combined real-time assessments of lateralization may reveal more about the underlying neural changes accompanying fluency achieved after long-term practice of prolonged speech, as was measured after short-term training in the present study. 80 TABLES Table 1: Neural regions associated with speech/language production in AWS and TS BAs Neural Region Inferior Frontal Cortex/Gyrus Abbrev. Function IFC/IFG Speech planning and executive control of speech Error monitoring, suppress unintended motor activity Motor speech production, action monitoring L FO linked with selfmonitoring, language repair Prosody tasks; change vocal pitch; motor prep., breath control during speech AWS relative to TS Less grey matter L Neg. correlate of stutt. severity (Preibisch, 2003a) 32/24 Anterior cingulate cortex ACC 44, 45 Broca’s (pars operculus, pars triangularis) PO 47/45 Frontal operculum FO 45/13 Insula -- 6/4/43 Rolandic operculum RO Articulation and phonation Underactive L Overactive R Superior Temporal Cortex/Gyrus STC/ STG Timing self-timed motor sequences Underactivity bilat. -- Speech processing; comprehension 22 Wernicke’s 41, 42 Primary auditory cortex Overactive R Underactivity L *Overactive R Not seen in TS (Preibisch et al., 2003a) *Overactive R Underactivity bilat. Neg correlation with stutt. severity (Neumann et al., 2003) 6 PAC Cerebellum Vermis of Lobule III, IV -- Motor cortex -- Supplementary Motor Area SMA Auditory processing Self-monitoring, timing motor execution, silent articulatory rehearsal (sup.) Integrate articulatory planning and sensory feedback Internal generation of motor activity, repetitive movements (speech and fingers) 81 *Underactivity bilat. *Overactive bilat. Underactive L Overactive R Overactivation L 6 4 2/3 PreSupplementary Motor Area Primary motor cortex Sensorimotor cortex Subcortical PreSMA Response selection, sequence motor plan Overactivation bilat. -- Motor execution Overactive R -- Motor execution Underactive L BG Internal “pacemaker”; timing for motor movement, innervate vocal tract muscles, sequencing Caudate Nucleus CN Motor timing, sequencing, execution Putamen Put Basal Ganglia Underactivation (Brown et al., 2005) Overactive bilat.; positive correlation stutt. severity Underactive L No activation Timing of motor output Rate-dependent motor Globus pallidus GPall (Brown et al., 2005) processing Motor planning; Increase in R postSubstantia nigra SN reward-seeking treatment Set shifting; motor Thalamus Thal execution; behavioral Overactive bilat. flexibility *Key areas specific to AWS during speech production identified by Brown et al. (2005) BAs= Brodmann Areas; AWS= adults who stutter; TS = typical speakers; bilat. = bilaterally 82 Table 2: Brodmann Areas (BAs) and functional connectivity corresponding to the prefrontal cortex regions to be measured by fNIRS BAs Prefrontal Cortex Region Connectivity Function Multitasking Episodic memory Theory of mind Anterior PFC Dorsal ACC, insula, Default mode: activated at rest and 10 (aPFC) lateral parietal lobe during “mind wandering” Application of two unrelated rules simultaneously Practiced tasks BG (CN, SN, Flexible strategy-use GlobPall, putamen), Higher order cognitive processing inferior parietal Executive function Dorsolateral lobe, Episodic memory PFC 9/46 cerebellum, Top-down guidance of attention/thought (dlPFC) premotor cortex, Narrative coherence pre-SMA, ACC, Self-regulation amygdala, STG Novel instruction processing Planning Projections to vlPFC Affect-guided decision-making Ventromedial Amygdala, Processing speed 10/11 PFC hypothalamus, Performance monitoring in emotionally (vmPFC) nucleus accumbens, arousing contexts ventral striatum Social cognitive processes Gilbert et al., 2010; Luu et al., 2003; Petrides, 2005; Ramnani & Owen, 2004; Roy, Shohamy & Wager, 2012; Tanji & Hoshi, 2008; Wood & Grafman, 2003 Table 3: Speaking task blocks counterbalanced across speech strategy conditions Task Category Task # Task Block 1 Task Block 2 Task Block 3 Conversation 1 what did last weekend (1 min.) Description 2 Plan a surprise party (1 min.) Joke 3 Tell memorized joke #1 (~ 45 secs) what will do this weekend (1 min.) Organize a move to a new home (1 min.) Tell memorized joke #2 (~ 45 secs) describe perfect weekend (1 min.) Trip to New York city plans (1 min.) Tell memorized joke #3 (~ 45 secs) 83 84 Age 23 49 22 48 29 31 20 32 47 38 26 33.2 Hand. MoCA STAI-S STAI-T +100 29 43 46 +100 30 42 49 +88 27 37 41 +100 28 53 53 +100 29 43 47 +88 27 42 49 +100 24 37 44 +75 28 44 58 +100 29 45 51 +100 29 43 51 +100 26 48 55 +96 28 43.4 49.5 # Sex Age 1 M 22 2 M 48 3 M 21 4 M 51 5 M 28 6 M 30 7 M 20 8 M 31 9 M 46 10 F 38 11 M 26 Mean: 32.8 Hand. MoCA STAI-S STAI-T +100 28 52 47 +100 30 38 47 +100 29 39 57 +100 29 42 52 +100 28 48 44 +50 30 36 43 +100 28 48 59 +88 30 48 50 +100 28 51 42 +100 29 54 72 +88 27 42 46 +94 29 45.3 50.8 %SS 0.15 0.00 0.10 0.18 0.00 0.62 0.00 0.00 0.03 0.05 0.26 0.13 SSI-4 2 (<very mild) 0 (<very mild) 2 (<very mild) 2 (<very mild) 0 (<very mild) 2 (<very mild) 0 (<very mild) 0 (<very mild) 2 (<very mild) 2 (<very mild) 2 (<very mild) 1.3 (<very mild) Family Hx Other (Medications, Treatment) None None Extended None Meds: Citalopram (20 mg/day) None None None None None None Meds: Zoloft (75mg/day) None %SS SSI-4 OASES Yrs. Tx Family Hx Other (Medications, Treatment) 17.9 25 (moderate) 2.14 15 Extended Tx: FS, stutt. mod., prop., SSG 4.5 18 (mild) 1.72 3 None Tx: FS, SSG 10.2 23 (mild) 1.99 2 Immediate No Tx since age 10 27.8 39 (very severe) 3.11 30 Extended Tx: PS, stutt. mod., SSG 6.6 22 (mild) 2.60 9 None Tx: PS, FS, SSG; Meds: Wellbutrin; 7.4 21 (mild) 2.26 8 None Tx: FS, relaxation/breathing 10.9 27 (moderate) 2.23 3 None Tx: PS, stutt. mod., SSG 8.8 23 (mild) 2.85 3 None Tx: PS, FS 16.4 37 (very severe) 2.68 12 None SSG 6.2 27 (moderate) 1.78 2 Immediate Tx: PS, FS, stutt. mod.; Meds: insulin (Type I Diabetes) 16.0 25 (moderate) 2.50 3 Extended Tx: PS, FS, stutt. mod, prop., SSG 12.1 26 (moderate) 2.35 8.2 b. TS (n =11; 1 Female) Included in Main Analyses # Sex 1 M 2 M 3 M 4 M 5 M 6 M 7 M 8 M 9 M 10 F 11 M Mean: a. AWS (n = 11; 1 Female) Included in Main Analyses Table 4: Participant Characteristics of Adults Who Stutter (AWS) and Typical Speakers (TS) 85 SSI-4 OASES Yrs. Tx 9 (<very mild) 1.86 2 10 (very mild) 2.08 7 15 (very mild) 2.45 2 %SS 0.00 0.00 0.00 SSI-4 0 (<very mild) 0 (<very mild) 0 (<very mild) Family Hx None None Immediate Family Hx Immediate Extended None Other (Medications, Treatment) Meds: Celexa Other (Medications, Treatment) Tx: Stutt. mod., SSG; Meds: Advair (100/50), Zyrtec Tx: FS, stutt. mod., prop. Tx: PS, relaxation/breathing Note: * indicates participants who were removed from subsequent fNIRS analysis because AWS’s overall stuttering %SS fell below 2%SS; Mean = values based on n=11(1 female) in each group after 3 marked * participants were removed; Age (Years); Hand (Handedness) based on Laterality Quotient +60 to +100 indicates right handedness, -60 to +60 indicates mixed handedness, -60 to -100 indicates left handedness (Veale, 2013); MoCA (Montreal Cognitive Assessment) screening score; ≥26 = within normal range; STAI-S and STAI-T = State/Trait Anxiety Inventory respectively (Spielberger, 1983); %SS (mean percent syllables stuttered on phone screening and three everyday speaking tasks during testing); SSI-4 (Stuttering Severity Instrument-4th Edition; Riley, 2009); OASES = Overall Assessment of Speaker’s Experience of Stuttering (Yaruss & Quesal, 2008); Family Hx (Immediate) an individual in the participant’s immediate family (e.g., parent, sibling) stutters or stuttered based on self-report; (Extended) an individual in the participant’s extended family (e.g., aunt, grandparent) stutters or stuttered based on self-report; Other = medications, reported history of techniques learned in stuttering treatment (Abbreviations: Meds = medications; Tx = treatment; PS = Prolonged speech, FS = Fluency shaping, stutt. mod. = stuttering modification, prop. = proprioception, SSG = attends or attended Stuttering Support Group at UVM) # Sex Age Hand. MoCA STAI-S STAI-T 1* F 49 +100 26 50 50 2* M 21 +100 28 48 48 3* M 34 +88 29 48 47 d. TS (n =3; 1 Female) Removed from Main Analyses # Sex Age Hand. MoCA STAI-S STAI-T %SS 1* F 49 +75 28 49 59 1.2 2* M 21 +100 28 46 42 1.6 3* M 36 +88 30 43 50 1.5 c. AWS (n =3; 1 Female) Removed from Main Analyses 86 Intra-Rater Reliability Category Very Mild Mild Very severe Initial SSI-4 Score 18 23 37 Initial Category Rating Mild Mild Very severe Participant 2 3 9 Group AWS IntraRater SSI-4 Score 16 22 37 Inter-Rater Reliability Category Very Mild Moderate Very Severe % diff SSI-4 Score 11.11% 4.35% 0% InterRater SSI-4 Score 16 27 38 11.11% 17.4% 2.7% % diff SSI-4 Score Table 6: Inter- and intra-rater reliability for stuttering severity by SSI-4 (Riley, 2009) categorical scale and numerical score 87 88 89 90 91 92 Subject 1 2 3 4 5 6 7 8 9 10 11 Metronome Prolonged Speech TS AWS TS AWS L +0.08 L +0.01 L +0.04 L +0.03 L +0.05 L +0.09 -0.03 R L +0.11 L +0.06 0.00 0.00 L +0.04 -0.01 R -0.03 R -0.01 R L +0.03 L +0.02 L +0.03 -0.04 R -0.04 R L +0.01 L +0.03 -0.11 R L +0.01 L +0.01 L +0.07 -0.02 R L +0.01 -0.07 R -0.06 R L +0.10 -0.06 R L +0.08 -0.02 R L +0.07 -0.02 R L +0.07 L +0.03 L +0.06 L + 0.03 -0.01 R -0.01 R -0.10 R L + 0.02 3R 8L 4R 6L 6R 4L 3R 8L *Possible LIs range from -1 (total right dominance) to +1 (total left dominance); 0 = no laterality AWS = Adults Who Stutter; TS = Typical Speakers L = relative leftward Laterality Index (LI) change in HbO activation (from Everyday condition LI to PS and Met) R = relative rightward Laterality Index (LI) change in HbO activation (from Everyday condition LI to PS and Met) Table 13: Relative change in Laterality Indices* (LIs) for all subjects (AWS and TS) from the Everyday condition to the Prolonged Speech and Metronome conditions 93 AWS TS AWS TS AWS TS EV 3.03 2.36 1.17 0.42 1.55 0.67 Mean 1.91 2.10 0.75 0.88 0.71 0.70 1.35 1.49 2.65 SE Left HbO Activation Mean p SE Diff. 0.67 0.81 1.87 EV= Everyday condition PS = Prolonged Speech condition MET = Metronome condition MET PS Group Cond. Right HbO Activation Mean p SE Mean Diff. 0.26 0.90 1.35 1.90 1.64 1.65 0.34 1.08 1.05 -0.60 1.13 0.62 1.47 0.93 -0.20 0.048 0.038 0.008 0.045 0.027 0.039 1.91 2.08 1.52 Mean SE 0.03 0.01 -.04 -0.01 0.33 0.33 0.024 0.008 0.04 SE Laterality Index Mean p SE Diff. 0.01 0.82 0.029 Table 15: Ancillary between-group (AWS vs. TS) pairwise comparison following linear mixed model of mean left and right hemisphere PFC HbO relative to baseline, Laterality Index and mean difference HbO between AWS (n = 3) and TS (n = 3) removed from main analyses 94 FIGURES Figure 1: Lateral, ventral and medial views of the PFC with Brodmann Areas (BAs) 9, 10, 11, and 46 (Fuster, 2002) 95 Figure 2: fNIR sensor with 4 light sources and 10 detectors (left, top) and 16 optode (channel) measurement locations registered on sensor (left, middle) and on brain surface image (right). fNIR sensor positioned on participants' head (left, bottom). Brain surface image from University of Washington, Digital Anatomist Project (Ayaz et al., 2012) Figure 3: Source and detectors of fNIRS 100CW measure (Obrig et al., 2000; http://www.biopac.com/fNIR300A-Data-Stimulation-System) 96 Figure 4: Order of optode positions with channels 1-8 measuring left hemisphere HbO and channels 9-16 measuring right hemisphere HbO (Tak & Ye, 2011) Figure 5: Approximate BIOPAC source detector localization coordinates of fNIRS optodes (León-Carrion, 2008) 97 2.8 2.4 2 Left HbO 1.6 1.2 A… A… 0.8 0.4 0 -0.4 -0.8 -1.2 Everyday PS MET Figure 6: Between-group (AWS and TS; n = 11 each) mean left hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome) 2.8 2.4 Right HbO 2 1.6 1.2 AWS 0.8 AWNS 0.4 0 -0.4 -0.8 -1.2 Everyday PS MET Figure 7: Between-group (AWS and TS; n = 11 each) right hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome) 98 0.06 Laterality Index (LI) 0.05 0.04 0.03 0.02 AWS 0.01 AWNS 0 -0.01 -0.02 -0.03 Everyday PS MET PS MET Figure 8: Between-group (AWS and TS; n = 11 each) Laterality Indices across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality AWNS Everyday AWS -0.04 -0.02 0 0.02 0.04 0.06 Laterality Index (LI) Figure 9: Between-group (AWS and TS; n = 11 each) Laterality Indices across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality 99 2 5 6 Right Laterality 7 AWS Participants Everyday 10 4 11 1 Left Laterality 3 9 8 -0.08 -0.04 0 0.04 Laterality Index 0.08 0.12 9 Right Right Laterality Laterality 1 3 TS Participants Everyday 2 4 7 5 Left Left Laterality Laterality 11 10 6 8 -0.08 -0.04 0 0.04 Laterality Index 0.08 0.12 Figure 10: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices Everyday condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality 100 5 Right Laterality 6 7 9 AWS Participants PS 10 8 4 Left Laterality 11 1 3 2 -0.08 -0.04 0 0.04 Laterality Index 0.08 0.12 2 Right Laterality 11 6 3 TS Participants PS 4 7 5 Left Laterality 1 9 10 8 -0.08 -0.04 *.22 0 0.04 0.08 0.12 Laterality Index Figure 11: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices Prolonged Speech condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality; *TS #8 = leftward LI dominance 101 4 Right Laterality 5 11 AWS Participants MET 9 6 10 3 8 Left Laterality 1 2 7 -0.08 -0.04 0 0.04 Laterality Index 0.08 0.12 4 Right Laterality 9 2 TS Participants MET 7 3 1 Left Laterality 8 11 5 6 10 -0.08 -0.04 .14 0 0.04 Laterality Index 0.08 0.12 Figure 12: Individual AWS (top graph; n = 11) and TS (bottom graph; n = 11) Laterality Indices Metronome condition; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality 102 103 104 105 106 107 5 4 Left HbO 3 2 AWS 1 AWNS 0 -1 -2 Everyday PS MET Figure 19: Ancillary between-group (AWS and TS; n = 3 each) mean left hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome) 5 4 Right HbO 3 2 AWS 1 AWNS 0 -1 -2 Everyday PS MET Figure 20: Ancillary between-group (AWS and TS; n = 3 each) mean right hemisphere HbO activation across conditions (Everyday, Prolonged Speech and Metronome) 108 0.08 0.07 Laterality Index (LI) 0.06 0.05 0.04 0.03 AWS 0.02 AWNS 0.01 0 -0.01 -0.02 Everyday PS MET PS MET Figure 21: Ancillary between-group (AWS and TS; n = 3 each) Laterality Indices (LIs) across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality AWNS Everyday AWS -0.04 -0.02 0 Right 0.02 LI 0.04 0.06 Left 0.08 Figure 22: Ancillary between-group (AWS and TS; n = 3 each) Laterality Indices (LIs) across conditions (Everyday, Prolonged Speech and Metronome); possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality 109 Figure 23: Ancillary Individual AWS (left graphs; n = 3) and TS (right graphs; n = 3) Laterality Indices (LIs) in Everyday (top), Prolonged Speech (middle) and Metronome (bottom) conditions of participants removed from main analyses; possible LIs range from -1 (total right dominance) to 1 (total left dominance); 0 = no laterality 110 REFERENCES Alderliesten, T., De Vis, J.B., Lemmers, P.M.A., van Bel, F., Benders, M.J.N.L., Hendrikse, J., & Peterson, E.T. (2013). Simultaneous quantitative assessment of cerebral physiology using respiratory-calibrated MRI and near-infrared spectroscopy in healthy adults. NeuroImage. http://dx.doi.org/10.1016/j.neuroimage.2013.07.015. Alexander, G., DeLong, M., & Stuck, P. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annual Review of Neuroscience, 9, 357-381. Alm, P.A. (2004). Stuttering and the basal ganglia circuits: A critical review of possible relations. Journal of Communication Disorders, 37(4), 325-369. Alm, P.A. (2005). On the causal mechanisms of stuttering. Doctoral thesis. Department of Clinical Neuroscience, Lund University, Sweden. Alm, P.A., Karlsson, R., Sundberg, M., & Axelson, H.W. (2013). Hemispheric lateralization of motor thresholds in relation to stuttering. Public Library of Sciences One, 8(10), e76824. Andrews, G., Guitar, B., & Howie, P. (1980). Meta-analysis of the effects of stuttering treatment. Journal of Speech and Hearing Disorders, 45, 287-307. Anglade, C., Thiel, A., & Ansaldo, A.I. (2014). The complementary role of the cerebral hemispheres in recovery from aphasia after stroke: A critical review of literature. Brain Injury, 28(2), 138-145. Arenth, P.M., Ricker, J.H., & Schultheis, M.T. (2007). Applications of functional nearinfrared spectroscopy (fNIRS) to neurorehabilitation of cognitive disabilities. The Clinical Neuropsychologist, 21(1), 38-57. Ayaz, H. (2010). “Functional Near Infrared Spectroscopy based Brain Computer Interface”. PhD Thesis, Drexel University, Philadelphia, PA. Ayaz, H., & Onaral, B. (2005). Analytical software and stimulus-presentation platform to utilize, visualize and analyze near-infrared spectroscopy measures. Master’s Degree thesis, Drexel University. Ayaz, H., Izzetoglu, M., Platek, S. M., Bunce, S., Izzetoglu, K., Pourrezaei, K., & Onaral, B. (2006). Registering fNIR Data to Brain Surface Image using MRItemplates, 28th IEEE EMBS Annual International Conference, New York, p.2671-2674. doi:10.1109/IEMBS.2006.260835. 111 Ayaz, H., Shewokis, P.A., Bunce, S., Izzetoglu, K., Willems, B., & Onaral, B. (2012). Optical brain monitoring for operator training and mental workload assessment. NeuroImage, 59(1), 36-47. Bahrami, H., Nejati, V., & Pouretemd, H. (2014). A comparative study of phonemic and semantic in children and adolescents with developmental stuttering and normal peers. Zahedan Journal of Research in Medical Sciences, 16(5), 41-44. Baicu, M.V., Watson, J.M., Maccotta, L., McDermott, K.B., Buckner, R.L., Gilliam, F.G., & Ojemann, J.G. (2005). Evaluating functional MRI procedures for assessing hemispheric language dominance in neurosurgical patients. Neuroradiology, 47, 835-844. Bajaj, A. (2007). Analysis of oral narratives of children who stutter and their fluent peers: Kindergarten through second grade. Clinical Linguistics & Phonetics, 21, 227245. Barch, D.M., Braver, T.S., Nystrom, L.E., Forman, S.D., Noll, D.C., & Cohen, J.D. (1997). Dissociating working memory from task difficulty in human prefrontal cortex. Neuropsychologia, 35, 373–1380. Beal, D.S., Gracco, V.L., Brettschneider, J., Kroll, R.M., & De Nil, L.F. (2013). A voxel based morphometry (VBM) analysis of regional grey and white matter volume abnormalities within the speech production network of children who stutter. Cortex, 4, 2151-2161. Bechara, A., Damasio, H., & Damasio, A.R. (2000). Emotion, decision making and the orbitofrontal cortex. Cerebral Cortex, 10, 295-307. Biermann-Ruben, K., Salmelin, R., & Schnitzler, A. (2005). Right rolandic activation during speech perception in stutterers: a MEG study. NeuroImage, 25, 793–801. Binder, J.R., Desai, R.H., Graves, W.W., & Conant, L.L. (2009). Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cerebral Cortex, 19, 2767-2796. Bisconti, S., DiSante, G., Ferrari, M., & Quaresima, V. (2012). Functional near-infrared spectroscopy reveals heterogeneous patterns of language lateralization over frontoplar cortex. Neuroscience Research, 73, 328-332. Blank, S.C., Scott, S.K., Murphy, K., Warburton, E., & Wise, R.J.S. (2002). Speech production: Wernicke, Broca and beyond. Brain, 125, 1829-1838. 112 Blomgren, M., Nagarajan, S., Lee, J., Li, T., & Alvord, L. (2003). Preliminary results of a functional MRI study of brain activation patterns in stuttering and nonstuttering speakers during a lexical access task. Journal of Fluency Disorders, 28, 337-356. Bloodstein, O. (1995). A handbook on stuttering, 5th edition, San Diego: Singular Press. Bloodstein, O., & Ratner, N.B. (2008). A handbook on stuttering. 6th edition, Clifton Park, NY: Delmar Cengage Learning. Boberg, E., Yeudall, L. T., Schopflocher, D., & Bo-Lassen, P. (1983). The effect of an intensive behavioral program on the distribution of EEG alpha power in stutterers during the processing of verbal and visuospatial information. Journalof Fluency Disorders, 8, 245–263. Bothe, A.K. (2004). Evidence-based treatment of stuttering: Empirical bases and clinical applications. Lawrence Erlbaum Associates Inc. Boynton, G. M., S. A. Engel, G. H. Glover, & D. J. Heeger. (1991). Linear systems analysis of functional magnetic resonance imaging in humans V1. Journal of Neuroscience, 16(13):4207– 4221. Braun, A.R., Varga, M., Stager, S., Schulz, G., Selbie, S., Maisog, J.M., Carson, R.E., & Ludlow, C.L. (1997). Altered patterns of cerebral activity during speech and language production in developmental stuttering. An h2(15)o positron emission tomography study. Brain, 120(5), 761-784. Brown, S., Ingham, R.J., Ingham, J.C., Laird, A.R., & Fox, P.T. (2005). Stuttered and fluent speech production: An ale meta-analysis of functional neuroimaging studies. Human Brain Mapping, 25(1), 105-117. Buchel, C., & Sommer, M. (2004). What causes stuttering? PLoS Biology, 2(2), e46. Buckner, R. L. (1998). Event-related fMRI and the hemodynamic response. Human Brain Mapping, 6, 373–377. Bunge, S.A., Ochsner, K.N., Desmond, J.E., Glover, G.H., & Gabrieli, J.D.E. (2001). Prefrontal regions involved in keeping information in and out of mind. Brain, 124, 2074-2086. Burgess, P.W., Quayle, A., & Frith, C.D. (2001). Brain regions involved in prospective memory as determined by positron emission tomography. Neuropsychologia, 39, 545-555. 113 Buchsbaum, B.R., Greer, S., Chang, W.-L., & Berman, K.F. (2005). Meta-analysis of neuroimaging studies of the Wisconsin Card-Sorting Task and component processes. Human Brain Mapping, 25, 35-45. Cadoret, G., Pike, G.B., & Petrides, M. (2001). Selective activation of the ventrolateral prefrontal cortex in the human brain during active retrieval processing. European Journal of Neuroscience, 14, 1164-1170. Cannizzaro, M., Dumas, J., Prelock, P.A., & Newhouse, P. (2012). Organizational structure reduces processing load in the prefrontal cortex during discourse processing of written language: Implications for high-level reading issues after TBI. Perspectives on Neurophysiology and Neurogenic Speech and Language Disorders, 22(2), 67-78. Chang, S.-E., Erickson, K.I., Ambrose, N.G., Hasegawa-Johnson, M.A., & Ludlow, C.L. (2008). Brain anatomy differences in childhood stuttering. NeuroImage, 39(3), 1333-1344. Chang, S.-E., Horwitz, B., Ostuni, J., Reynolds, R., & Ludlow, C.L. (2011). Evidence of left inferior frontal-premotor structural and functional connectivity deficits in adults who stutter. Cerebral Cortex, 11, 2507-2518. Chang, S.-E., Kenney, M.K., Loucks, T.M., & Ludlow, C.L. (2009). Brain activation abnormalities during speech and non-speech in stuttering speakers. NeuroImage, 46, 201–212. Chang, S.-E., & Zhu, D.C. (2013). Neural network connectivity difference in children who stutter. Brain. doi: 10.1093/brain/awt275. Civier, O., Tasko, S.M., & Guenther, F.H. (2010). Overreliance on auditory feedback may lead to sound/syllable repetitions: Simulations of stuttering and fluencyinducing conditions with a neural model of speech production. Journal of Fluency Disorders, 35, 246-279. Coelho, C., Le, K., Mozeiko, J., Krueger, F., & Grafman, J. (2012). Discourse production following injury to the dorsolateral prefrontal cortex. Neuropsychologia, 50(14), 3564-3572. Cognitive Optical Brain Imaging (COBI) Studio. fNIR Devices, LLC. fNIR Imager & COBI Studio Manual. Drexel University Cope, M., & Delpy, D. (1988). System for long-term measurement of cerebral blood and tissue oxygenation on newborn infants by near infra-red transillumination. Medical & Biological Engineering & Computing, 26(3), 289-294. 114 Cykowski, M.D., Kochunov, P.V., Ingham, R.J., Ingham, J.C., Mangin, J.F., Riviere, D., Lancaster, J.L., & Fox, P.T. (2008). Perisylvian sulcal morphology and cerebral asymmetry patterns in adults who stutter. Cerebral Cortex, 18, 571–583. Cykowski, M. D., Fox, P.T., Ingham, R.J., Ingham, J.C., & Robin, D. A. (2010). A study of the reproducibility and etiology of diffusion anisotropy differences in developmental stuttering: a potential role for impaired myelination. Neuroimage, 52(4), 1495-1504. Davidow, J.H. (2013). Systematic studies of modified vocalization: the effect of speech rate on speech production measures during metronome-paced speech in persons who stutter. International Journal of Language & Communication Disorders. doi: 10.1111/1460-6984.12050 Davidson, R.J. (1998). Affective style and affective disorders: perspectives from affective neuroscience. Cognition & Emotion, 12, 307-330. Davidson, R.J., Ekman, P., Saron, C., Senulis, J., & Friesen, W.V. (1990). Approach/withdrawal and cerebral asymmetry: Emotional expression and brain physiology, I. Journal of Personality and Social Psychology, 58, 330-341. Davidson, R.J., Marshall, J.R., Tomarken, A.J., Henriques, J.B. (2000). While a phobic waits: Regional brain electrical and autonomic activity in social phobics during anticipation of public speaking. Biological Psychiatry, 47, 85-95. Deblaere, K., Boon, P.A., Vandemaele, P., Tieleman, A., Vonck, K., Vingerhoets, G., Backes, W., Defreyne, L., & Achten, E. (2004). MRI language dominance assessment in epilepsy patients at 1.0 T: region of interest analysis andcomparison with intracarotid amytal testing. Neuroradiology, 46, 413-420. Delis, D.C., Kaplan, E., & Kramer, J.H. (2001). Delis-Kaplan Executive Function System (D-KEFS). San Antonio, TX: The Psychological Corporation. De Nil, L. F., & Brossardt, H.G. (2001). Studying stuttering from a neurological and cognitive information processing perspective. In H.G. Brosshardt, J.S. Yaruss, & H.F.M. Peters (Eds.), Fluency disorders: Theory, research, treatment, and selfhelp. Proceedings of the third world conference of fluency disorders (pp. 53-58). Nijmegen, The Netherlands: Nijmegen University Press. De Nil, L.F., & Kroll, R.M. (2001). Searching for the neural basis of stuttering treatment outcome: Recent neuroimaging studies. Clinical Linguistics & Phonetics, 15,163– 168. 115 De Nil, L.F., Kroll, R.M., Kapur, S., & Houle, S. (2000). A positron emissiontomography study of silent and oral single word reading in stuttering andnonstuttering adults. Journal of Speech Language and Hearing Research, 43(4),1038-1053. De Nil, L.F., Kroll, R.M., Lafaille, S.J., & Houle, S. (2003). A positron emission tomography study of short- and long-term treatment effects on functional brain activation in adults who stutter. Journal of Fluency Disorders, 28(4), 357-380. Dieler, A.C., Tupak, S.V., & Fallgatter, A.J. (2012). Functional near-infrared spectroscopy for the assessment of speech related tasks. Brain & Language, 121, 90-109. Ehlis, A.-C., Schneider, S., Dresler, T., & Fallgatter, A.J. (2013). Application of functional near-infrared spectroscopy in psychiatry. Neuroimage. http://dx.doi.org/10.1016/j.neuroimage.2013.03.067. Elfgren, C.I., & Risberg, J. (1998). Lateralized frontal blood flow increases during fluency tasks: influence of cognitive strategy. Neuropsychologia, 36(6), 505-512. Ezrati-Vinacour, R., & Levin, I. (2004). The relationship between anxiety and stuttering: a multidimensional approach. Journal of Fluency Disorders, 29, 135-148. Faul, F., Erdfelder, E., Buchner, A., & Lang, A.-G. (2009). Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41, 1149-1160. Floyer-Lea, A., & Matthews, P.M. (2005). Distinguishable brain activation networks for short- and long-term motor skill learning. Journal of Neurophysiology, 94, 512518. Foundas, A.L., Bollich, A.M., Corey, D.M., Hurley, M., & Heilman, K.M. (2001). Anomalous anatomy of speech-language areas in adults with persistent developmental stuttering. Neurology, 57, 207–215. Fox, P.T., Ingham, R.J., Ingham, J.C., Hirsch, T.B., Downs, J.H., Martin, C., Jerabek, P., Glass, T., & Lancaster, J.L. (1996). A PET study of the neural systems of stuttering. Nature, 382, 158–161. Fox, P.T., Ingham, R.J., Ingham, J.C., Zamarripa, F., Xiong, J.-H., & Lancaster, J.L. (2000). Brain correlates of stuttering and syllable production: A PET performance-correlation analysis. Brain, 123, 1985-2004. Frith, C., Friston, K., Liddle, P., Frackowiak, R.S.J. (1991). A PET study of word finding. Neuropsychologia, 29, 1137–1148. 116 Fuster, J.M. (2002). Frontal lobe and cognitive development. Journal of Neurocytology, 31(3-5), 373-85. Gabrieli, J.D.E., Poldrack, R.A., & Desmond, J.E. (1998). The role of the left prefrontal cortex in language and memory. Proceedings of the National Academy of Science USA, 95, 906-913. Gerfen, C.R., Engber, T.M., Mahan, L.C., Susel, Z., Chase, T.N., Monsma, F.J. Jr., & Sibley, D.R. (1990). D1 and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science, 250, 1429-1432. Gervain, J., Mehler, J., Werker, J.F., Nelson, C.A., Csibra, G., Lloyd-Fox, S., Shukla, M., & Aslin, R.N. (2011). Near-infrared spectroscopy: A report from the McDonnell infant methodology consortium. Developmental Cognitive Neuroscience, 1, 2246. Ghashghaei, H. T., & Barbas, H. (2002). Pathways for emotion: interactions of prefrontal and anterior temporal pathways in the amygdala of the rhesus monkey. Neuroscience, 115, 1261–1279. Gilbert, S.J., Gonen-Yaacovi, G., Benoit, R.G., Volle, E., & Burgess, P.W. (2010). Distinct functional connectivity associated with lateral versus medial rostral prefrontal cortex: A meta-analysis. NeuroImage, 53, 1359-1367. Gilbert, S.J., Spengler, S., Simons, J.S., Steele, J.D., Lawrie, S.M., Frith, C.D., & Burgess, P.W. (2006). Functional specialization within rostral prefrontal cortex (area 10): A meta-analysis. Journal of Cognitive Neuroscience, 18(6), 932-948. Giraud, A.-L., Neumann, K., Bachoud-Levi, A.-C., von Gudenberg, A.W., Euler, H.A., Lanfermann, H., & Preibisch, C. (2008). Severity of dysfluency correlates with basal ganglia activity in persistent developmental stuttering. Brain andLanguage, 104(2), 190-199. Glascher, J., Adolphs, R., Damasio, H., Bechara, A., Rudrauf, D., Calamia, M., Paul, L.K., & Tranel, D. (2012). Lesion mapping of cognitive control and value-based decision making in the prefrontal cortex. Proceedings of the National Academy of Sciences, 109(36), 14681-14686. Goldman-Rakic, P.S. (1987). Circuitry of primate prefrontal cortex and regulation of behavior by representational memory. In F. Plum & U. Mouncastle (Eds.), Handbook of physiology, Vol. 5. (pp. 373–417). Washington, DC: The American Physiological Society. 117 Greiner, J., Fitzgerald, H., & Cooke, P. (1986). Speech fluency and hand performance on a sequential tapping task in left- and right-handed stutterers and nonstutterers. Journal of Fluency Disorders, 11, 55-69. Guitar, B. (2013). Stuttering: An Integrated Approach to Its Nature and Treatment 4th edition. Lippincott, Williams & Wilkins, Baltimore, MD. Haslinger, B., Erhard, P., Kampfe, N., Boecker, H., Rummeny, E., Schwaiger, M., Conrad, B., & Ceballos-Baumann, A.O. (2001). Event-related functional magnetic resonance imaging in Parkinson’s disease before and after levodopa. Brain, 124(3), 558-570. Heitmann, R.R., Asbjornsen, A., & Helland, T. (2004). Attentional focus in speech fluency disorders. Logopedics Phoniatrics Vocology, 29, 119-127. Henry, J.D., & Crawford, J.R. (2004). Meta-analytic review of verbal fluency performance following focal cortical lesions. Neuropsychology, 18(2), 284-295. Herder, C., Howard, C., Nye, C., & Vanryckeghem, M. (2006). Effectiveness of Behavioral Stuttering Treatment: A Systematic Review and Meta-Analysis. Contemporary Issues in Communication Science and Disorders, 33, 61-73. Herholz, K., Thiel, A., Wienhard, K. Pietrzyk, U., von Stockhausen, H.M., Karbe, H., Kessler, J., Bruckbauer, T., Halber, M., & Heiss, W.-D. (1996). Individual functional anatomy of verb generation. NeuroImage, 3, 185–194. Herrmann, M. J., Walter, A., Ehlis, A. C., & Fallgatter, A. J. (2006). Cerebral oxygenation changes in the prefrontal cortex: Effects of age and gender. Neurobiology of Aging, 27(6), 888–894. Hickok, G. (2009). The functional neuroanatomy of language. Physics of Life Reviews, 6(3), 121-143. Hickok, G., & Poeppel, D. (2007). The cortical organization of speech processing. Nature Reviews Neuroscience, 8, 393-402. Holtzer, R., Mahoney, J.R., Izzetoglu, M., Izzetoglu, K., Onaral, B., & Verghese, J. (2011). fNIRS study of walking and walking while talking in young and old individuals. Journal of Gerontology Academy of Biological Sciences and Medical Sciences, 66(8), 879-87. Homae, F. (2014). A brain of two halves: Insights into interhemispheric organization provided by near-infrared spectroscopy. NeuroImage, 85, 354-362. 118 Hoshi, Y. (2005). Functional near-infrared spectroscopy: Potential and limitations in neuroimaging studies. International Review of Neurobiology, 66, 237–266. Howard-Jones, P.A., Blakemore, S.J., Samuel, E.A., Summers, I.R., & Claxton, G. (2005).Semantic divergence and creative story generation: an fMRI investigation. Cognitive Brain Research, 25(1), 240-250. IBM Corp. Released 2013. IBM SPSS Statistics for Windows, Version 22.0. Armonk, NY: IBM Corp. Ingham, R.J. & Andrews, G. (1971). Stuttering: The quality of fluency after treatment. Journal of Communication Disorders, 4, 279-288. Ingham, R.J., Bothe, A.K., Jang, E., Yates, L., Cotton, J., & Seybold, I. (2009). Measurement of speech effort during fluency-inducing conditions in adults who do and do not stutter. Journal of Speech, Language, and Hearing Research, 52, 1286-1301. Ingham, R.J., Bothe, A.K., Wang, Y., Purkhiser, K., & New, A. (2012). Phonation interval modification and speech performance quality during fluency-inducing conditions by adults who stutter. Journal of Communication Disorders, 45(3), 198-211. Ingham, R.J., Fox, P.T., Ingham, J.C., & Zamarripa, F. (2000). Is overt stuttered speech a prerequisite for the neural activations associated with chronic developmental stuttering? Brain and Language, 75, 163–194. Ingham, R.J., Warner, A., Byrd, A., & Cotton, J. (2006). Speech effort measurement and stuttering: Investigating the chorus reading effect. Journal of Speech, Language, and Hearing Research, 49, 660-670. Irani, F., Platek, S.M., Bunce, S., Ruocco, A.C., & Chute, D. (2007). Functional near infrared spectroscopy (fNIRS): an emerging neuroimaging technology with important applications for the study of brain disorders. The Clinical Neuropsychologist, 21(1), 9-37. Jenkins, I.H., Brooks, D.J., Nixon, P.D., Frackowiak, R.S., & Passingham, R.E. (1994). Motor sequence learning: a study with positron emission tomography. Journal of Neuroscience, 14, 3775-3790. Josse, G., Seghier, M.L., Kherif, F., & Price, C.J. (2008). Explaining function with anatomy: language lateralization and corpus callosum size. Journal of Neuroscience, 28, 14132-14139. 119 Kalinowski, J. & Dayalu, V. (2002). A Common Element in the Immediate Inducement of Effortless, Natural-Sounding, Fluent Speech in Stutterers: "The SecondSpeech Signal". Medical Hypotheses, 58(1), 61-66. Kameyama, M., Fukuda, M., Uehara, T., & Mikuni, M. (2004). Sex and age dependencies of cerebral blood volume changes during cognitive activation: A multichannel near-infrared spectroscopy study. Neuroimage, 22(4), 1715–1721. Kane, M.J., & Engle, R.W. (2002). The role of the prefrontal cortex and working memory capacity, executive attention, and general fluid intelligence: An individualdifference perspective. Psychonomic Bulletin & Review, 9(4), 637-671. Kell, C.A., Morillon, B., Kouneiher, F., & Giraud, A.-L. (2011). Lateralization of speech production starts in sensory cortices- A possible sensory origin of cerebral left dominance for speech. Cerebral Cortex, 21, 932-937. Kell, C.A., Neumann, K., Von Kriegstein, K., Posenenske, C., Von Gudenberg, A.W., Euler, H., & Giraud, A. (2009). How the brain repairs stuttering. Brain, 132(10), 2747–2760. Kennan, R.P., Kim, D., Maki, A., Koizumi, H., & Constable, R.T. (2002). Non-invasive assessment of language lateralization by transcranial near infrared optical topography and functional MRI. Human Brain Mapping, 16(3), 183-189. Kikuchi, Y., Ogata, K., Umesaki, T., Yoshiura, T., Kenjo, M., Hirano, Y., Okamoto, T., Komune, S., & Tobimatsu, S. (2011). Spatiotemporal signatures of an abnormal auditory system in stuttering. NeuroImage, 55, 891–899. doi: 10.1016/j.neuroimage.2010.12.083. Kleinow, J., & Smith, A. (2000). Influences of length and syntactic complexity on the speech motor stability of fluency speech of adults who stutter. Journal of Speech, Language and Hearing Research, 43, 548-559. Kovelman, I., Shalinsky, M.H., White K.S., Schmitt, S.N., Berens, M.S., Paymer, N., & Petitto, L.-A. (2009). Dual language use in sign-speech bimodal bilinguals: fNIRS brain-imaging evidence. Brain and Language, 109(2-3), 112-123. Kwee, I.L. & Nakada, T. (2003). Dorsolateral prefrontal lob activation declines significantly with age. Journal of Neurology, 250(5), 525-529. Lehericy, S., Cohen, L., Bazin, B., Samson, S., Giacomini, E., Rougetet, R., Hertz Pannier, L., Le Bihan, D., Marsault, C., & Baulac, M. (2000). Functional MR evaluation of temporal and frontal language dominance compared with the Wada test. Neurology, 54, 1625-1633. 120 León-Carrion J., Damas-López, J., Martín-Rodríguez, J.F., Domínguez-Roldán, J.M., Murillo-Cabezas, F., Barroso Y. Martin, J.M. & Domínguez-Morales, M.R. (2008). The hemodynamics of cognitive control: the level of concentration of oxygenated hemoglobin in the superior prefrontal cortex varies as a function of performance in a modified Stroop task. Behavioural brain research, 193(2), 248256. Lindauer, U., Dirnagl, U., Fuchtemeier, M., Bottiger, C., Offenhauser, N., Leithner, C., & Royl, G. (2010). Pathophysiological interference with neurovascular coupling when imaging based on hemoglobin might go blind. Frontiers in Neuroenergetics, 2(25), 1-9. doi: 10.3389/fnene.2010.00025 Loucks, T., Kraft, S.J., Choo, A.L., & Sharma, H. (2011). Functional brain activation differences in stuttering identified with a rapid fMRI sequence. Journal of Fluency Disorders, 36, 302-307. Lu, C., Chen, C., Ning, N., Ding, G., Guo, T., Peng, D., Yang, Y., Li, K., & Lin, C. (2010a). The neural substrates for atypical planning and execution of word production in stuttering. Experimental Neurology, 221, 146–156. Lu, C., Peng, D., Chen, C., Ning, N., Ding, G., Li, K., Yang, Y., & Lin, C. (2010b). Altered effective connectivity and anomalous anatomy in the basal ganglia thalamocortical circuit of stuttering speakers. Cortex, 46, 49–67. Luu, P., Tucker, D.M., Derryberry, D., Reed, M., & Poulsen, C. (2003). Electrophysiological responses to errors and feedback in the process of action regulation. Psychological Science, 14, 47–53. Makris, N., Kennedy, D.N., McInerney, S., Sorensen, A.G., Wang, R., Caviness Jr., V.S., & Pandya, D.N. (2005). Segmentation of subcomponents within the superior longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study. Cerebral Cortex, 15, 854–869. Max, L. & Gracco, V.L. (2005). Coordination of oral and laryngeal movements in the perceptually fluent speech of adults who stutter. Journal of Speech, Language, and Hearing Research, 48(3), 524-542. Medvedev, A.V. (2014). Does the resting state connectivity have hemispheric asymmetry? A near-infrared spectroscopy study. NeuroImage, 85, 400-407. Miller, E.K., & Cohen, J.D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167-202. 121 Monchi, O., Petrides, M., Petre, V., Worsley, K., & Dagher, A. (2001). Wisconsin card sorting revisited: distinct neural circuits participating in different stages of the task identified by event-related functional magnetic resonance imaging. The Journal of Neuroscience, 21(19), 7733-7741. Moore, W.H. (1984). Hemispheric alpha asymmetries during an electromyographic biofeedback procedure for stuttering. Journal of Fluency Disorders, 17, 143-162. Muroi, A., Hirayama, K., Tanno, Y., Shimizu, S., Watanabe, T., & Yamamoto, T. (1999). Cessation of stuttering after bilateral thalamic infarction. Neurology, 53, 890-891. Nagy, A., Eordegh, G., Paroczy, Z., Markus, Z., & Benedek, G. (2006). Multisensory integration in the basal ganglia. European Journal of Neuroscience, 24, 917-924. Nasreddine, Z. (2010). The Montreal Cognitive Assessment (MoCA). Version 7.1. www.mocatest.org. Nasreddine, Z., Phillips, N.A., Bédirian, V., Charbonneau, S., Whitehead, V., Collin, I., Cummings, J.L., & Chertkow, H. (2005). The Montreal Cognitive Assessment (MoCA): A Brief Screening Tool For Mild Cognitive Impairment. Journal of the American Geriatrics Society, 53, 695-699. Neef, N.E., Jung, K., Rothkegel, H., Pollok, B., von Gudenberg, A.W., Paulus, W., & Sommer, M. (2011). Right-shift for non-speech motor processing in adults who stutter. Cortex, 47, 945-954. Neumann, K., Euler, H.A., Gudenberg, A.W.v., Giraud, A.-L., Lanfermann, H., Gall, V., & Preibisch, C. (2003). The nature and treatment of stuttering as revealed by fMRI: A within- and between-group comparison. Journal of Fluency Disorders, 28(4), 381-410. Neumann, K., Preibisch, C., Euler, H.A., von Gudenberg, A.W., Lanfermann, H., Gall, V., & Giraud, A.-L. (2005). Cortical plasticity associated with stuttering therapy. Journal of Fluency Disorders, 30(1), 23-39. O’Brian, S., Onslow, M., Cream, A., & Packman, A. (2003). The Camperdown Program: Outcomes of a new prolonged-speech treatment model. Journal of Speech Language and Hearing Research, 46, 933-946. Obrig, H., Wenzel, R., Kohl, M., Horst, S., Wobst, P., Steinbrink, J., Thomas, F., & Villringer, A. (2000). Near-infrared spectroscopy: does it function in functional activation studies of the adult brain? International Journal of Psychophysiology 35(2-3), 125-142. 122 Okada, F., Takahashi, M., & Tokumitsu, Y. (1996). Dominance of the ‘nondominant’ hemisphere in depression. Journal of Affective Disorders, 37(1), 13-21. Oldfield, R.C. (1971). The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia, 9, 97-113. Özge, A., Toros, F., & Çömelekoğlu, U. (2004). The Role of Hemispheral Asymmetry and Regional Activity of Quantitative EEG in Children with Stuttering. Child Psychiatry and Human Development, 34(4), 269-280 Packman, A., Code, C., & Onslow, M. (2007). On the cause of stuttering: integrating theory with brain and behavioral research. Journal of Neurolinguistics, 20, 353362. Packman, A., Onslow, M., Menzies, R. (2000). Novel speech patterns and the treatment of stuttering. Disability and Rehabilitation, 22(1-2), 65-79. Packman, A., Onslow, M., Richard, F., & van Doorn, J. (1996). Syllabic stress and variability: A model of stuttering. Clinical Linguistics & Phonetics, 10, 235-263. Petrides, M. (2005). Lateral prefrontal cortex: architectonic and functional organization. Philosophical Transactions of the Royal Society Biological Sciences, 360, 781795. Pollok, B., Gross, J., Muller, K., Aschersleben, G., & Schnitzler, A. (2005). The cerebral oscillatory network associated with auditorily paced finger movements. NeuroImage, 24(3), 646-655. Pool, K.D., Devous, M.D., Freeman, F.J., Watson, B.C., & Finitzo, T. (1991). Regional cerebral blood flow in developmental stutterers. Archives in Neurology, 48, 509512. Preibisch, C., Neumann, K., Raab, P., Euler, H.A., von Gudenberg, A.W., Lanfermann, H., & Giraud, A.L. (2003a). Evidence for compensation for stuttering by the right frontal operculum. NeuroImage, 20(2), 1356-1364. Preibisch, C., Raab, P., Neumann, K., Euler, H.A., von Gudenberg, A.W., Gall, V., Lanfermann, H., & Zanella, F. (2003b). Event-related fMRI for the suppression of speech-associated artifacts in stuttering. NeuroImage, 19(3), 1076-1084. Price, C.J. (2012). A review and synthesis of the first 20 years of fMRI studies of heard speech, spoken language and reading. NeuroImage, 62, 816-847. 123 Price, J.L., Carmichael, S.T. & Drevets, W.C. (1996). Networks related to the orbital and medial prefrontal cortex; a substrate for emotional behavior? Progress in Brain Research, 107, 523–536. Quaresima, V., Bisconti, S., & Ferrari, M. (2012). A brief review on the use of functional near-infrared spectroscopy (fNIRS) for language imaging studies in human newborns and adults. Brain and Language, 121(2), 79-89. Raichle, M.E., Fiez, J.A., Videen, T.O., Macleod, A.M.K., Pardo, J.V., Fox, P.T. & Petersen, S.E. (1994). Practice-related changes in human brain functional anatomy during nonmotor learning. Cerebral Cortex, 4(1), 8-26. Ramnani, N., & Owen, A. (2004). Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nature Reviews Neuroscience, 5(3), 184-194. Ramnani, N., & Passingham, R.-E. (2001). Changes in the human brain during rhythm learning. Journal Cognitive Neuroscience, 13, 1–15. Rauch, S.L., Savage, C.R., Alpert, N.M., Fischman, A.J., & Jenike, M.A. (1997). The functional neuroanatomy of anxiety: A study of three disorders using positron emission tomography and symptom provocation. Biological Psychiatry, 42, 446452. Raz, N. (2000). Aging and the brain and its impact on cognitive performance: integration of structural and functional findings. In F.I.M. Craik, & T.A. Salthouse (Eds.), The handbook of aging and cognition (pp. 1-90). Mahwah, NJ: Lawrence Erlbaum Associates. Ridderinkhof, K.R., Ullsperger, M., Crone, E.A., & Nieuwenhuis, S. (2004). The role of the medial prefrontal cortex in cognitive control. Science, 306, 443-447. Ries, S., Janssen, N., Dufau, S., Alario, F.-X., & Burle, B. (2011). General-purpose monitoring during speech production. Journal of Cognitive Neuroscience, 23(6), 1419-1436. Ries, S.K., Xie, K., Haaland, K.Y., Dronkers, N.F., & Knight, R.T. (2013). Role of the lateral prefrontal cortex in speech monitoring. Frontiers in Human Neuroscience, 7, 1-16. Riley, G.D. (2009). Stuttering Severity Instrument for children and adults—fourth edition (SSI-4). Austin, TX: Pro-Ed. Robinson, G., Shallice, T., Bozzali, M., & Cipolotti, L. (2012). The differing roles of the frontal cortex in fluency tests. Brain, 135, 2202-2214. 124 Rossi, S., Telkemeyer, S., Wartenburger, I., & Obrig, H. (2012). Shedding light on words and sentences: near-infrared spectroscopy in language research. Brain & Language, 121, 152-163. Roy, M., Shohamy, D., & Wager, T.D. (2012). Ventromedial prefrontal-subcortical systems and the generation of affective meaning. Trends in Cognitive Sciences, 16(3), 147-156. Sachs, O., Weis, S., Zellagui, N., Sass, K., Huber, W., Zvyagintsev, M., Mathiak, K., & Kircher, T. (2011). How different types of conceptual relations modulate brain activation during semantic priming. Journal of Cognitive Neuroscience, 23, 12631273. Salmelin, R., Schnitzler, A., Schmitz, F., & Freund, H. (2000). Single word reading in developmental stutterers and fluent speakers. Brain, 123(Part 6), 1184-1202. Salmelin, R., Schnitzler, A., Schmitz, F., Jancke, L., Witte, O. W., & Freund, H. J.(1998). Functional organization of the auditory cortex is different in stutterers and fluent speakers. Neuroreport, 9(10), 2225-2229. Sato, Y., Mori, K., Koizumi, T., Minagawa-Kawai, Y., Tanaka, A., Ozawa, E., Wakaba, Y., & Mazuka, R. (2011). Functional lateralization of speech processing in adults and children who stutter. Frontiers in Psychology, 2(70), 1-10. Savage, C.R., Deckersbach, T., Heckers, S., Wagner, A.D., Schachter, D.L., Alpert, N.M, Fischman, A.J., & Rauch, S.L. (2001). Prefrontal regions supporting spontaneous and directed application of verbal learning strategies: evidence from PET. Brain, 124, 219-231. Scherer, L.C., Ska, B., Giroux, F., Lesage, F., Senhadji, N., Marcotte, K., Tomitch, L.M.B, Benali, H., & Joanette, Y. (2007). Discourse comprehension in successful aging: A NIRS study. Brain and Language, 103(1-2), 228-229. Scholkmann, F., Kleiser, S., Metz, A.J., Zimmerman, R., Pavia, J.M., Wolf, U., & Wolf, M. (2014). A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology. NeuroImage, 85, 6-27. Shalinsky, M.H., Kovelman, I., Berens, M.S., & Petitto, L.-A. (2009). Analysis and data collection method for exploring the brain's cognitive functions in babies, children and adults using functional near infrared spectroscopy (fNIRS). Journal of Visualized Experiments, (29), e1268. DOI: 10.3791/1268. 125 Smits-Bandstra, S., & De Nil, L.F. (2007). Sequence skill learning in persons who stutter: Implications for cortico-striato-thalamo-cortical dysfunction. Journal of Fluency Disorders, 32, 251-278. Sommer, M., Koch, M.A., Paulus, W., Weiller, C., & Buchel, C. (2002). Disconnection of speech-relevant brain areas in persistent developmental stuttering. The Lancet, 360, 380-383. Sowman, P.F., Crain, S., Harrison, E., & Johnson, B.W. (2012). Reduced activity of left orbitofrontal cortex precedes blocked vocalization: A magnetoencephalographic study. Journal of Fluency Disorders, 37, 359-365. Sowman, P.F., Crain, S., Harrison, E., & Johnson, B.W. (2014). Lateralization of brain activation in fluent and non-fluent preschool children: A magnetoencephalographic study of picture-naming. Frontiers in Human Neuroscience, 8, 354.doi:10.3389/fnhum.2014.00354. Spielberger, C.D., Gorsuch, R.L., Lushene, R., Vagg, P.R., & Jacobs, G.A. (1983). State Trait Anxiety Inventory (STAI). Consulting Psychologists Press, Inc. Printed by Mind Garden, Inc. Stager, S.V., Jeffries, K.J., & Braun, A.R. (2003). Common features of fluency-evoking conditions studied in stuttering subjects and controls: An h215o pet study. Journal of Fluency Disorders, 28(4), 319-336. Stephan, K.E, Marshall, J.C., Friston, K.J., Rowe, J.B., Ritzl, A., Zilles, K., & Fink, G.R. (2003). Lateralized cognitive processes and lateralized task control in the human brain. Science, 301, 384-386. Stocco, A., & Lebiere, C. (2012). Distinct contributions of the caudate nucleus, rostral prefrontal cortex, and parietal cortex to the execution of instructed tasks. Cognitive Affective Behavioral Neuroscience, 12, 611-628. Strick, P.L., Dum, R.P., & Picard, N. (1995). Macro-organization of the circuits connecting the basal ganglia with the cortical areas. In J.C. Houk, J.L. Davis, & D.G. Beiser (Eds.), Models of information processing in the basal ganglia (pp. 117–130). Cambridge, MA: MIT Press. Sutton, S.K., Ward, R.T., Larson, C.L., Holden, J.E., Perlman, S.B., & Davidson, R.J. (1997). Asymmetry in prefrontal glucose metabolism during appetitive and aversive emotional states: An FDG-PET study. Psychophysiology, 34, S89. 126 Tak, S. & Ye, J.C. (2011). NIRS-SPM: Statistical Parametric Mapping for Near-infrared Spectroscopy. Version 4. Bio Imaging Signal Processing (BISP) Lab. Department of Bio and Brain Engineering, KAIST. Korea. Taniwaki, T., Okayama, A., Yoshiura, T., Togao, O., Nakamura, Y., Yamasaki, T., Ogata, K., Shigeto, H., Ohyagi, Y., Kira, J., & Tobimatsu, S. (2006). Functional network of the basal ganglia and cerebellar motor loops in vivo: Different activation patterns between self-initiated and externally triggered movements. NeuroImage, 31, 745-753. Tanji, J., & Hoshi, E. (2008). Role of the lateral prefrontal cortex in executive behavioral control. Physiological Reviews, 88, 37-57. Tellis, G.M., Mesquita, R.C., & Yodh, A.G. (2011). Use of diffuse correlation spectroscopy to measure brain blood flow differences during speaking and nonspeaking tasks for fluent speakers and persons who stutter. Perspectives on Fluency and Fluency Disorders, 21(3), 96-106. Toyomura, A., Fujii, T., & Kuriki, S. (2011). Effect of external auditory pacing on the neural activity of stuttering speakers. NeuroImage, 57(4), 1507-1516. Van Schouwenburg, M., Aarts, E., & Cools, R. (2010). Dopaminergic modulation of cognitive control: distinct roles for the prefrontal cortex and the basal ganglia. Current Pharmaceutical Design, 16, 2026-2032. Veale, J.F. (2013). Edinburgh Handedness Inventory- Short Form: A revised version based on confirmatory factor analysis. Laterality: Asymmetries of Body, Brain and Cognition. doi:10.1080/1357650X.2013.783045. Wagner, A.D., Maril, A., Bjork, R.A., & Schacter, D.L. (2001). Prefrontal contributions to executive control: fMRI evidence for functional distinctions within lateral prefrontal cortex. NeuroImage, 14, 1337-1347. Warburton, E., Wise, R.J., Price, C.J., Weiller, C., Hadar, U., Ramsay, S., & Frackowiak, R.S.J. (1996). Noun and verb retrieval by normal subjects: studies with PET. Brain, 119, 159–79. Watanabe, E., Maki, A., Kawaguchi, F., Takashiro, K., Yamashita, Y., Koizumi, H., & Mayanagi, Y. (1998). Non-invasive assessment of language dominance with near infrared spectroscopic mapping. Neuroscience Letters, 256(1), 49-52. Watson, B.C., Freeman, F.J., Devous Sr., M.D., Chapman, S.B., Finitzo, T., & Pool, K.D. (1994). Linguistic performance and regional cerebral blood flow in persons who stutter. Journal of Speech and Hearing Research, 37, 1221-1228. 127 Weber-Fox, C., Wray, A.H., & Arnold, H. (2013). Early childhood stuttering and electrophysiological indices of language processing. Journal of Fluency Disorders, 38(2), 206-221. Webster, W.G. (1998). Brain models and the clinical management of stuttering. Journal of Speech-Language Pathology & Audiology, 22, 220–230. Webster, W.G. (2004). From hand to mouth: Contributions of theory to evidence-based practice. In A. K. Bothe (Ed.), Evidence-based treatment of stuttering (pp. 17-26). Mahwah, NJ: Lawrence Erlbaum. Wiener, M., Turkeltaub, P., & Coslett, H.B. (2010). The image of time: A voxel-wise meta-analysis. NeuroImage, 49(2), 1728-1740. Wood, A.G., Saling, M.M., Abbott, D.F., & Jackson, G.D. (2001). A neurocognitive account of frontal lobe involvement in orthographic lexical retrieval: An fMRI study. NeuroImage, 14, 162-169. Wood, J.N., & Grafman, J. (2003). Human prefrontal cortex function: processing and representational perspectives. Nature Reviews Neuroscience, 4, 139–147. Wu, J.C., Maguire, G., Riley, G., Fallon, J., LaCasse, L., Chin, S., Klein, E., Tang, C., Cadwell, S., & Lottenberg, S. (1995). A positron emission tomography [18F] deoxyglucose study of developmental stuttering. Neuroreport, 6, 501–505. Wu, T., Wang, L., Hallett, M., Chen, Y., Li, K., & Chan, P. (2011). Effective connectivity of brain networks during self-initiated movement in Parkinson’s disease. Neuroimage, 55, 204–15. Xu, J., Kemeny, S., Park, G., Frattali, C., & Braun, A. (2005). Language in context: emergent features of word, sentence, and narrative comprehension. Neuroimage, 25(3), 1002-1015. Yairi, E., & Ambrose, N. (1999). Early childhood stuttering I: Persistency and recovery rates. Journal of Speech, Language, and Hearing Research, 42, 1097-1112. Yaruss, J.S., & Quesal, R. (2006). Overall Assessment of the Speaker’s Experience of Stuttering (OASES): Documenting multiple outcomes in stuttering treatment. Journal of Fluency Disorders, 31, 90-115. Yaruss, J.S., & Quesal, R. (2008). Overall Assessment of the Speaker’s Experience of Stuttering (OASES). Pearson Education, Inc. 128 Yu, G. (2012). Diffuse correlation spectroscopy (DCS): A diagnostic tool for assessing tissue blood flow in vascular-related diseases and therapies. Current Medical Imaging Reviews, 8, 194-201. Zelanic, H. N., Smith, A., Franz, E. A., & Ho, M. (1997). Differences in bimanual coordination associated with stuttering. Acta Psychologica, 96, 229–243. 129