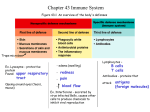
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Innate immune system wikipedia , lookup
DNA vaccination wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Monoclonal antibody wikipedia , lookup
Duffy antigen system wikipedia , lookup
Adaptive immune system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
277 Bioscience Reports 2, 277-287 (1982) Printed in Great Britain T h e p r e d o m i n a n t s u r f a c e g l y c o p r o t e i n s of t h y m o c y t e s and l y m p h o c y t e s Review Alan F, WILLIAMS MRC Cellular Immunology Unit, Sir William Dunn School of Pathology, University of Oxford, Oxford OXI 3RE, U.K. The surfaces of thymocytes and T and B lymphocytes d i f f e r m a r k e d l y in t h e i r p r e d o m i n a n t glycoprotein constituents. Amino acid sequence studies show that a surprising number of t h e cell surface molecules have homologies with immunoglobulins, Analysis :of carbohydrate shows major differences between glycoproteins on the same cell and between the same (or closely related) glycoproteins on the different cell types, Introduction T h e r e are two main types of immunocompetent lymphocytes: B lymphocytes, which differentiate in the bone marrow and give rise to antibody-secreting cells, and T lymphocytes, which differentiate in t h e thymus and are responsible for ceil-mediated immunity. The surface molecules of both B and T lymphocytes have been intensively studied and also those of immature T lymphocytes from the thymus (thymocytes). Under the microscope mature B and T lymphocytes and most thymocytes look very similar, with a large nuclear:cytoplasmic ratio, few mitochondria, and an absence of endoplasmic reticulum. These features are consistent with the f a c t that these are non-dividing cells w~th low levels of metabolism and biosynthetic activity. tn the absence of antigen, lymphocytes have a migratory existence during which they recirculate from blood to lymph. This involves s p e c i f i c r e c o g n i t i o n of s p e c i a l i z e d e n d o t h e l i u m in lymph nodes, movement between endothelial cells, and migration into areas which are different for B and T lymphocytes. The time of passage through the nodes is much shorter for T lymphocytes than for B lymphocytes (l). If foreign antigen is encountered a complex set of recognition events occurs. B lymphocytes can bind antigen directly with their immunoglobulin receptors but require interactions with macrophages and T l y m p h o c y t e s to trigger cell division and differentiation tO produce antibody-secreting cells and memory cells (2). T lymphocytes a p p e a r to r e c o g n i z e antigen in association with histocompatibility antigen presented on macrophages or dendritic cells. The molecular basis of this is not understood at all (3). Given the above functions it seems reasonable to think that the cell surface molecules of B and T lymphocytes will be mainly involved with the recognition phenomena seen in recirculation and cell positioning, and antigen recognition. For thymocytes, surface molecules may be involved in cell positioning and in events leading to committment to a particular specificity of antigen recognition. I would 01982 The Biochemical Society 278 WILLIAMS expect that metabolite transport molecules or ion pumps would be minor components of the membranes of these cells. The situation would presumably be different for the dividing precursor cells of the early stages of lymphoid differentiation or the dividing cells resulting from stimulation of mature lymphocytes with antigen. An example is the transferrin receptor seen on dividing but not quiescent lymphocytes (4). In this article I will discuss the major surface glycoproteins of thymocytes and of mature T and B lymphocytes with regard to three m a i n a s p e c t s : (a) their patterns of expression and functions, (b) homology of a number of molecules with immunoglobulins, and (c) heterogeneity in their carbohydrate structures. Methods Monoclonat antibodies now provide the primary tools for identifying celt surface proteins (reviewed in refs. 576)~ Purification has been achieved using affinity chromatography with lectin or antibody columns (5~7). The assignment of a molecule as a major surface component rests on a demonstration that it accounts for one of the major entities seen in surface labelling studies and on determination of the number of molecules per cell. For example, on rat thymocytes the three heavily glycosylated molecules given in Table 1 account for the three main bands seen when sialic acid or galactose residues are labelled at the cell surface with 3H borohydride (8). Two of these molecules (L-CA and Thy-1) account for the two main glycoproteins which bind to lentil tectin and are identified by Coomassie Blue staining after sodium dodecyI sulphate polacrylamide-gel electrophoresis (9). LSGP does not bind to lentil tectin, but most cell surface glycoproteins do, as judged by the fact that most of the proteins tabelled with t25I by t h e l a c t o p e r o x i d a s e method are bound by this lectin (g). Taken together these data suggest that there may be only three abundant glycoproteins on thymocytes. Other molecules found at levels of about 10~ to 3 x 10~ molecules per cell have been described and include the markers of T-lymphocyte subsets (5). Patterns o5 E x p r e s s i o n Glycoproteins and Functions o5 t h e Major S u r f a c e The known abundant constituents of lymphoid surfaces are given in Table i, which shows data for rat lymphoid cells, The same patterns seem to apply equally to mouse or human cells~ with the exception that T h y - I antigen is found on lymphocytes as well as thymocytes in the mouse and not on either in humans (reviewed in ref. tO) and that the main human Class-II antigens are homologues of the rodent E type ( t 1). A l l the m o l e c u l e s listed in Table 1 are glycoproteins or multimers of protein and glycoprotein chains, and all but LSGP and Thy-1 are known to traverse the lipid bilayer ( i 2 - 1 4 ) . For LSGP there are at present no data on membrane integration. Thy-1 has all the properties of a molecule which binds to the lipid bilayer, but these cannot be due to the polypeptide and there is evidence for a nonprotein hydrophobic 'tail' (10). LYMPHOCYTE GLYCOPROTEINS 279 c~ ,~ u m S ~g m ao ~4 ~i U 0 ,~ I> ,iJ ~ ~, ~.~ ~,~ U ~I ,,,,,I ,'~ ~ q~ ~ ,~: "~ E ~ ~, ~ ~.~ 1 r,~ e q ~ v v m I-I ~ ,.~_t ~ ~'t '~'I ~'-1 ,--~ C> 1.1 Cu~ ~ .,.4 m Q~ 0 m 0 m 0 ~ ".'4 ~ 2 0 i 1~ N I-~ I>-, ~ :>,, C,,,~ l-~A= Itl ~ E 9to e ~ 280 WILLIAMS Simple inspection of Table 1 shows that the different lymphoid cell types are very different in their major cell surface constituents. The only molecule in Table 1 which may be identical on any two cell types is the Class-I histocompatibility antigen on T and B lymphocytes. Thymocytes and T l y m p h o c y t e s both express the LSGP and all the c e l l s express L-CAs. However, the carbohydrate on these glycoproteins differs on the different cell types (see below). Presumably the differences in surface molecules are related to the functions of the cells. However, for most glycoproteins this is difficult to think a b o u t in a simple way because of their unpredictable expression amongst various cell types, including non-lymphoid ceils (Table 1). Of t h e m o l e c u l e s in Table i, f u n c t i o n s are known only for immunoglobulin and histocompatibility antigens. Immunoglobulin is the antigen receptor of B lymphocytes and is not known to be synthesized by any other cell type (15). This molecule thus fits with the concept of a uniquely expressed molecule mediating a particular function of a differentiated cell. Histocompatibility antigens function as controlling elements in T-lymphocyte responses (2,3). T lymphocytes appear to recognize foreign antigen only at the surface of a presenting cell and t h i s s o m e h o w involves the s i m u l t a n e o u s r e c o g n i t i o n of h i s t o c o m p a t i b i l i t y antigens. The part of the histocompatibility antigen which i s recognized is that which shows polymorphism within the species. Class-I histocompatibiiity antigens seem to be mainly involved in recognition by cytotoxic T ceils and may be particularly relevant to the killing of ceils infected with virus, while Ctass-II antigens are recognized by helper T cells and are thus important in the initiation of i m m u n e responses controlled by these ceils. The wide tissue d i s t r i b u t i o n of Class-I antigens thus seems to fit well with their function, since most cell types might by infected by virus. HoweveG the distribution of Class-II antigens at present seems bizarre, since only macrophages and/or dendritic ceils are known to be involved in the presentation of antigen to helper T cells. The expression of LSGP and Thy-I antigen on various cell types is as unusual as t h a t of Class-II antigens. These antigens are all d i s c r e t e l y e x p r e s s e d on some, but not all cell types without any obvious functional correlation. This expression cannot be dismissed as a serological a r t i f a c t since Class-II antigens (16), Thy-1 (17), and LSGP (18) have all been characterized from different celt types and the results show that the shared antigenicity reflects the expression of a s i m i l a r molecule (probably identical in the polypeptide) on the different cell types. The phenomenon of unusual patterns of antigen expression is clearly of major biological significance, since it applies to other membrane molecules (5) as well as those shown in Table l. The results of Structural studies on the major glycoproteins provide some basic data for thinking about the functions of these molecules. Homology with Immunoglobulins Fig. l shows cell s u r f a c e m o l e c u l e s with known amino acid sequences found on lymphoid cells. These include histocompatibility antigens, Thy-1 antigen, and immunoglobulins. These molecules have all been studied for completely unrelated reasons yet they all turn out to show sequence homology and are all likely to contain segments homologous to the Ig-fold. LYMPHOCYTE GLYCOPROTEINS 281 Immunoglobulin-Related Molecules Found At Cell Surfaces IgM V-domains MHC Antigens "~r-~ ", ~,,o ClassI Class][ C/~"rj~<~l.>=~) C~,~ <,al "-'t TTTT ITTTTTTTI TTTTTTTTTT' ,IJTTI Cytoplasm Fig. 1, Tmmunoglobulin-related molecules found at lymphoid cell surfaces. Ig domains and their homologous regions in the other molecules are represented by circles. Intra-chain disulphide bonds are shown by the ~ symbols and interchain bonds in IgM by dashed lines. N-linked carbohydrate structures are shown by ~ and n and c identify the N-terminus and C-terminus of the polypeptides with the exception of ~2-microglobulin (~2-m). The polymorphic determinants of the major histocompatibility (MHC) antigens are found mainly in the heavy chains of Class-I antigens and the 13 chains of Class-ll antigens. The diagrams are based on references 10,19,34 (Thy-l); 42 (Class-I antigens); ii,12~22-24~46 (Class-ll antigens); 14,47 (IgM). The h e a v y and light chains of immunoglobulins can be subdivided i n t o d o m a i n s of a b o u t 110 a m i n o acids which show s e q u e n c e and structural homology (18). This is i l l u s t r a t e d for an IgM m o l e c u l e in Fig. i. In all c a s e s (including V - d o m a i n s ) t h e s e domains a r e c h a r a c t e r i z e d by a c o n s e r v e d disulphide bond and a c h a r a c t e r i s t i c folding p a t t e r n r e f e r r e d to as t h e I g - f o l d (18). This consists of a n t i - p a r a l l e l B - s t r a n d s f o r m i n g t w o B - s h e e t s w h i c h are held t o g e t h e r by the disulphide bond and by hydrophobic i n t e r a c t i o n s b e t w e e n the sheets. S e q u e n c e homologies b e t w e e n the I g - d o m a i n s a r e seen m o s t l y around t h e c o n s e r v e d c y s t e i n y l residues and in s t r e t c h e s of s e q u e n c e which f o r m the B-strands. A p a t t e r n for V- or C - d o m a i n s can be distinguished by p a r t s of the s e q u e n c e which are c o n s e r v e d within but not b e t w e e n t h e s e domains, and by h o m o l o g i e s in the middle of the domain w h e r e V - d o m a i n s h a v e e x t r a s e q u e n c e c o m p a r e d with C - d o m a i n s . Thus 282 WILLIAMS in looking for homologies with Ig the features to be noted are: the c h a r a c t e r i s t i c disulphide bond, B - s h e e t s t r u c t u r e , and sequence homologies. B2-microglobulin (B2-m) and T h y - i antigen have all of these features and both are homologous to a single immunoglobulin domain. Thy-1 ~ t s best with V-domains (19) while B2-m is clearly like a C-domain (20). Parts of the Class-I heavy chain (2i) and Class-II a (22) and B (23,24) chains also have homology with Ig domains in terms of the conserved disulphide bond and sequence homology. In all cases these are clearly like Ig C-domains rather than V-domains. Thus all of the domain-like segments of histocompatibility antigens are h o m o l o g o u s to C - d o m a i n s , and Thy-1 antigen is the only non-Ig molecule so far identified which has a V-domain pattern. Carbohydrate Heterogeneity The nature oi the carbohydrate of lymphoid glycoproteins has been studied by analysis of their composition and by measuring their binding to l e c t i n s . The r e s u l t s show that there are striking differences b e t w e e n m o l e c u l e s on t h e same cell and between the same (or related) molecules on different cells. Cell surface glycoproteins can be labeIled with 3H at sialic acid or galactose residues by reducing with 3H borohydride after oxidation with p e r i o d i c acid, or g a l a c t o s e oxidase subsequent to neuraminidase digestion to expose galactose residues (25). When this is done only a few heavily labelled glycoproteins are seen, namely L-CA, LSGP, and Thy-1 on thymocytes, L-CA and LSGP on T tymphocytes, and L-CA only on B lymphocytes (8). The thymocyte molecules have all been purified (26) and their carbohydrate compositions show that the LSGP molecule has very different carbohydrate structures from L-CA and Thy-i (Table 2). The composition and size of the LSGP carbohydrate structures suggest that one amino acid in five has attached to it an O-linked structure of the type: sialic acid c~2-~ Gal 81-3~ GaINAc ta2-6 sialic acid Ser/Thr. The molecule is thus highly acidic and is likely to have extended c o n f o r m a t i o n , since t h e a t t a c h m e n t of such a large number of carbohydrate structures might be expected to inhibit protein folding. S t u d i e s on binding of glycoproteins to lectins have established differences in their carbohydrates between cells. For Thy-1 antigen, differences were seen in binding to lentil lectin of the brain and thymus forms of the antigen (17), and more recently peanut lectin ( P N L ) and soybean lectin (SBL) have been particularly useful in a n a l y s i n g c a r b o h y d r a t e h e t e r o g e n e i t y b e t w e e n different types of lymphoid cells (27). PNL is specific for GalB1-3GalNAc disaccharide, which is the asialo form of the O-linked structure given above. PNL binds in large amounts to thymocytes compared with T lymphocytes and has been used to fractionate ceils from the thymus (28). The LSGP is clearly LYMPHOCYTE Table 2. GLYCOPROTEINS 293 Carbohydrate compositions of the three major rat-thymocyte glycoproteins L-CA Thy-i LSGP % Carbohydrate 25 32 60 Estimated structures per i00 amino acids N-linked 1.5 3 nil O-linked few nil 20 Residues per I00 amino acids Fucose 0.9 1 0 Mannose 4.4 10.6 0 Galactose 3.6 5.5 22.3 Glucosamine 5.7 9.4 0 Galactosamine 1.1 0 20.1 Sialic acid 2.9 2.1 27.2 Data from refs. 17 and 26. a candidate for interaction with PNL, and as expect ed the whole of this glycoprotein from t h y m o c y t e s and T lymphocytes binds to a PNL affinity column if terminal sialic acid is removed with neuraminidase. With undigested cells part of the LSGP from t hym ocyt es binds to the PNL column but no binding of the T - l y m p h o c y t e LSGP occurs (27). This suggests that the differential binding of PNL is due simply to the f a c t that t h y m o c y t e carbohydrate structures partially lack terminal sialic acid while those of T lymphocytes are fully sialylated. Previous studies in the mouse have shown that the same phenomenon occurs for r h y - 1 antigen ( 2 9 ) . T h y m o c y t e Thy-1 antigen partially lacks sialic acid while T - l y m p h o c y t e Thy-l is fully sialylated. Thus it seems t hat th e c a r b o h y d r a t e structures of t h y m o c y t e s may in general lack sialic a c i d and t h a t in t h e d i f f e r e n t i a t i o n to mature lymphocytes the glycosylation changes to give structures which have a full complement of terminal sialic acid. SBL is s p e c i f i c for terminal galactosamine and binds in large amounts to 15 lymphocytes but virtually not at all to T lymphocytes. In a f f i n i t y chromatography no glycoproteins from t h y m o c y t e s or T lymphoctyes bound t o SBL, but a significant fraction of B-lymphocyte L - C A was bound ( 2 7 ) . A f t e r t h e r e m o v a l of sialic acid with neuraminidase all of the B-lymphocyte L-CA bound to the SBL column but still virtually no t h y m o c y t e or T - l y m p h o c y t e L-CA was bound. In af f in ity chromatography with the PNL column, p a r t of the asialo L-CA f r o m T l y m p h o c y t e s and B lymphocytes was bound but t h y m o c y t e L-CA did not bind even in the asialo form. Taken t o g e t h e r these data establish t hat L-CA differs in its c a r b o h y d r a t e st ruct ures on all t h r e e 28• WILLIAMS cell types, and this conclusion is supported by the report that L-CA f r o m r a t s p l e e n ( f r o m B and T l y m p h o c y t e s ) expresses the 'i' car b o h y d r at e antigenic determinant while the t h y m o c y t e form does not (30). L C - A shows e x t e n s i v e s e r o l o g i c a l c r o s s - r e a c t i o n between the d i f f e r e n t cell types and it seems likely that the di fferent forms share the same polypeptide with differing carbohydrate structures. However, t h e r e are antibodies which distinguish B-lymphocyte L-CA(31,32) from the other forms, and sequence studies will be essential to establish whether the polypeptides are identical or whether the different L-CAs are coded by a closely related but non-identical set of genes. Discussion A t the outset I suggested that the major molecules at the surface of lymphocytes should be mainly involved in the recognition functions which occur in antigen recognition or lymphocyte recirculation and p o s i t i o n i n g in l y m p h o i d t i s s u e s . For immunoglobulins and histocompatibility antigens this is so, with both sets of molecules being involved in different ways in antigen recognition. A relationship in the functions of these molecules is now strongly supported by the findings of homology in the sequences. The functions of immunoglobulins are in part understood at the level of the individual domains wh ile for h i s t o c o m p a t i b i l i t y antigens this is not so. For histoc o m p a t i b i l i t y a n t i g e n s all t h a t is known is that somehow their polymorphic parts must be recognized at the same time that foreign antigen is recognized by T lymphocytes at a cell surface. T h e f u n c t i o n s of immunoglobulins can be divided into antigen r e c o g n i t i o n by V - d o m a i n s and mediation of e f f e c t o r functions by C-domains. Both sorts of domains have a similar folding pat t ern for their polypeptide chain, and the different functions are mediated by sequences which do not contribute to the structural stability of the Ig-fold. These occur in the bends connecting the anti-parallel strands and also possibly on the exposed surfaces of the B-strands (33). For both V- and C-domains it could be argued that the essence of the f u n c t i o n is t h a t t h e Ig-fold provides a stable platform on which variations of sequence can occur and mediate di fferent recognition functions. This idea is clearly compatible with the function seen for histocompatibility antigens. Can the argument be extended to Thy-1 antigen? The structural homologies would argue for this, and it has b e e n s u g g e s t e d t h a t Thy-1 antigen functions as a ligand in cell i n t e r a c t i o n s r e s p o n s i b l e for tissue formation (34). A key point regarding Thy-1 antigen is that it breaks the link between the Ig-fold and immunological recognition functions. This is so since expression of Thy-1 is conserved in different species in neuronal ceils and fibroblasts but not on lymphoid cells (10). If Thy-1 is involved in recognition functions then the distribution of the antigen would suggest that it can undertake a triggering role on more than one cell type. The carbohydrate het er oge ne i t y of the surface glycoproteins is also of interest in thinking about recognition phenomena. The hypothesis th at carbohydrates mediate cellular interactions by acting as ligands for recognizers (glycosyl transferases or lectins) on other ceils has long been discussed (35,36). There is no direct evidence for this LYMPHOCYTE GLYCOPROTEINS 285 hypothesis but clearly heterogeneity of the sort seen in the carbohydrates of lymphoid glycoproteins could be a basis for such interactions. It is established that the carbohydrates of enzymes (37) and serum glycoproteins (3g) can play an informational role in the uptake of t h e s e m o l e c u l e s by cells, and it is easy to imagine that the carbohydrate of LSGP, L-CA, or Thy-I might be presented as ligands for mediating cell interactions. If this view is taken then one can argue that the same antigen on different cell types could provide a cell-specific ligand in that the carbohydrate may be cell-type-specific (I7). Alternatively one could argue that the carbohydrate is important for structural reasons. This seems to be the case for the N-linked carbohydrate structure of IgG (39), and for Thy-l antigen correct glycosylation is essential for the molecule to be expressed at the cell surface (40). Structural aspects could include determining t h e charge characteristics of the cell surface, and this may be the main role of the LSGP. P o s s i b l y t h e s u r f a c e carbohydrates will turn out to have both i n f o r m a t i o n a l and structural roles. Acknowledgements t am grateful to Mrs. Chris Scott for assistance with the manuscript. References I. Ford W~ 9 Smith ME & Andrews P (1978) in Cell-Cell Recognition (Curtis ASG, ed) 9 pp 359-3929 Cambridge University Press 9 Cambridge. 2. Katz DH (1980) in Advances in Immunology 29 (Kunkel HG & Dixon FJ, eds), pp 137-207, Academic Press, New York and London. 3. Klein J, Jureti~ A, Baxevanis CN & Nagy ZA (1981) Nature 291, 455-460. 4. Trowbridge IS & Omary MB (1981) Proc. Natl. Acad. Sci. U.S.A. 78, 3039-3043. 5. Williams AF (1980) Biochem. Soc. Symp. 459 27-50. 6. Milstein C & Lennox E (1980) in Current Topics in Developmental Biology 14 (Friedlander M~ ed), pp 1-32, Academic Press, New York and London. 7. Crumpton MJ & Snary D (1979) in Defence and Recognition IIB Structural Aspects(Lennox ES, ed), International Review of Biochemistry 23, pp 127-176, University Park Press, Baltimore. 8. Standring R, M c M a s t e r W R 9 Sunderland CA & Williams AF (1978) Eur. J. Immunol. 8, 832-839. 9. Standring R & Williams AF (1978) Bioehim. Biophys. Acta 508, 85-96. i0. Campbell DG, Gagnon J, Reid KBM & Williams AF (1981) Biochem. J. 195~ 15-30. ii. Cook RG, Siegelman M H, Capra JD~ Uhr JR & Vitetta ES (1979) J. Immunol. 122, 2232-2236. 286 WILLIAMS 12. Owen MJ, Knott JCA & Crumpton MJ (1980) Biochemistry 19, 3092-3099. 13. Omary MB & Trowbridge IS (1980) J. Biol. Chem. 255, 16621669. 14. Kehry M, Ewald S) Douglas R, Sibley C~ Raschke W, Fambrough D & Hood L (1981) Cell 21, 393-406. 15. Goding JW) Scott DW & Layton JE (1977) Immunol. Rev. 37, 152-186. 16. Klareskog L, Forsum U & Peterson PA (1980) Eur. J. Immunol. 10) 958-963. 17. Barclay AN, Letarte-Muirhead M) Williams AF & Faulkes R (1976) Nature 263, 563-567. 18. Amzel LM & Poljak RJ (1979) Ann. Rev. Biochem. 48, 961-997. 19. Williams AF & Gagnon J (1982) Science 216, 696-703. 20. Peterson PA, Cunningham BA, BerggSrd I & Edelman GM (1972) Proc. Natl. Acad. Sci. U.S.A. 69, 1697-1701. 21. Orr HT, Lancet D, Robb RJ, Lopez de Castro JA & Strominger JL (1979) Nature 282, 266-270. 22. Lee JS) Trowsdale J, Travers PJ, Carey J) Jenkins J & Bodmer WF, manuscript in preparation. 23. Kratzin H, Yang C-Y) GStze H, Pauly E, K~ibel S, Egert G) Thinnes FP) Wernet P) Altevogt P & Hilschmann N (1981) Hoppe-Seyler's Z. Physiol. Chem. 362, 1665-1669. 24. Larhammar D, Wiman K) Schenning L, Claesson L, Gustafsson K, Peterson PA & Rask L (1981) Scand. J. Immunol. 14, 617-622. 25. Gahmberg CG, H~yry P & Andersson LC (1976) J. Cell Biol. 68, 642-653. 26. Brown WRA, Barclay AN) Sunderland CA & Williams AF (1981) Nature 289, 456-460. 27. Brown WRA & Williams AF, in press, Immunology 46. 28. Sharon N (1980) in Progress in Immunology IV (Fougereau M & Dausset J) eds)) pp 254-278, Academic Press, New York and London. 29. Hoessli D) Bron C & Pink JRL (1980) Nature 283, 576-578. 30. Childs RA & Feizi T (1981) Biochem. Biophys. Res. Commun. 102, 1158-1164. 31. Dalchau R & Fabre JW (1981) J. Exp. Med. 153) 753-765. 32. Coffman RL & Weissman IL (1981) Nature 289, 681-683. 33. Burton DR) Boyd J) Brampton AD) Easterbrook-Smith SB) Emanuel EJ) Novotn~ J, Rademaeher TW) van Schravendijk MR) Sternberg MJE & Dwek RA (1980) Nature 288, 338-344. 34. Cohen FE) Novotn~ J) Sternberg MJE) Campbell DG & Williams AF (1981) Biochem. J. 195) 31-40. 35. Hughes RC (1975) Essays in Biochemistry II) 1-36. 36. Williams AF (1978) Biochem. Soc. Trans. 6, 490-494. 37. Home LH, Weissmann B & Neufeld E (1979) Proc. Natl. Aead. Sci. U.S.A. 76, 2331-2334. 38. Tanabe T) Pricer WE & Ashwell G (1979) J. Biol. Chem. 254) 1038-1043. 39. Huber R) Diesenhofer J) Colman PM) Matsushima M & Palm W (1976) Nature 264) 415-420. 40. Trowbridge IS, Hyman R & Mazauskas C (1978) Cell 14) 21-32. 41. Williams AF (1975) Eur. J. Immunol. 5) 883-885. LYMPHOCYTE GLYCOPROTEINS 287 42. Ploegh HL, Orr HT & Strominger JL (1981) Cell 24, 287-299. 43. BjDrck L, ~kerstrDm B & BerggSrd I (1979) Eur. J. Immunol. 9, 486-490. 44. Fukumoto T, McMaster WR & Williams AF (1982) Eur. J. Immunol. 12~ 237-243. 45. Barclay AN (1981) Immunology 44, 727-736. 46. Owen MJ, Kissonerghis A-M~ Lodish HF & Crumpton MJ (1981) J. Biol. Chem. 256, 8987-8993. 47. Putnam FW, Florent G, Paul C, Shinoda T & Shimizu A (1973) Science 182, 287-291. 48. Trowbridge IS (1978) J. Exp. Med. 148, 313-323.