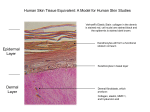
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
Molecular mimicry wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Adaptive immune system wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Life Sciences 67 (2000) 477Ð502 Minireview The cutaneous citadel A holistic view of skin and immunity Brad Spellberg, M.D.* Department of Medicine, Harbor-UCLA Medical Center, Torrance, CA, USA Abstract Human skin has 4 major functions: endogenous homeostasis (e.g. regulation of body temperature and ßuid balance), metabolism (e.g. Vitamin D synthesis), sensory input, and to serve as a barrier to external threats (e.g. infection, mechanical injury, ultraviolet light). It is the latter function which concerns this review, for the skinÕs remarkable success in protecting the human body from the outside world is a major component of our immune system. The eminent pathologist, Virchow, whose work in the mid 19th century revolutionized many aspects of medical understanding, viewed the skin as an effective but inanimate barrier (1). However, recent technologies have elucidated a highly complex, dynamic interplay between the skin and other members of the immune system. © 2000 Elsevier Science Inc. All rights reserved. Keywords: Immunology; Cutaneous inßammation; Dermatology Overview of the skin organ Skin commensal organisms Skin covers approximately 1.5 to 2 m2 of the average humanÕs surface area, and represents by weight the largest organ in the human body. Its absolutely critical role in protecting us from the microbial hordes can be inferred from the alarmingly high infection rate incurred following disruption of the skin barrier, for example by extensive burn destruction (2Ð4), or by placement of percutaneous catheters in patients (5,6). Human skin provides habitation for approximately 1 million bacteria per cm2 (this number varies depending upon the location on the body, e.g. intertriginous counts are much higher, while bacterial density is lower between the scapulae) (7Ð9). The total burden of bacteria co* Corresponding author. 1201 South Catalina Ave. #12, Redondo Beach, CA 90277, USA. Tel.: 310-3161740; fax: 310-541-8634. E-mail address: [email protected] (B. Spellberg) 0024-3205/00/$ Ð see front matter © 2000 Elsevier Science Inc. All rights reserved. PII: S 0 0 2 4 - 3 2 0 5 ( 0 0 )0 0 6 5 3 -6 478 B. Spellberg / Life Sciences 67 (2000) 477Ð502 existing on the average humanÕs skin is thus about 10 billion. These commensal organisms aid in our protection against more virulent microbes. Several mechanisms of this commensalderived protection have been described (10), including pure competition for nutrients and space (11), commensal bacterial secretion of toxins such as lysozyme (12,13) and the bacteriocins (14,15), which eliminate competing microorganisms, and a continued stimulation of the local immune response which keeps the cells of the immune system primed for combat at all times (16Ð18). Gram-positive skin commensals also play a crucial role in maintaining the low pH of the skin microenvironment, which helps inhibit microbial growth. Lipases elaborated by grampositive cocci degrade complex skin triglycerides into free fatty acids which lower the skin pH (19,20) and are also directly toxic to competing microbes (21,22). The importance of the normal commensal component of skin is highlighted by infections which are known to be preceded by disruption of this steady-state balance, for example by extensive use of antibiotics which wipe out normal ßora (23Ð25). Skin ontogeny and histology The skin (Fig. 1) is derived from ectodermal (e.g. keratinocytes, melanocytes, peripheral nerves) and mesodermal (e.g. leukocytes, dermal vasculature, Þbroblasts) precursors (26). In general, ectodermally derived structures populate the superÞcial epidermis, while mesodermally derived components tend to dwell in the deeper dermis. However, during fetal development and throughout life some leukocytes cross the structural barrier between the epidermis and dermis and reside, at least temporarily, in the epidermis. The epidermis Epidermal histology The vast majority of cells found in the epidermis are a distinct type of squamous epithelium which produce a tough, proteinaceous barrier as a bastion against the perils of the external universe. This barrier is comprised of Þbrils of the intermediate Þlament, keratin, hence these specialized epithelial cells are called keratinocytes. Keratin is an effective physical hindrance to penetration by microbes into the deeper compartments of our bodies, and serves as perhaps the single most important contribution of the skin to our immune system. The epidermis can be divided into several substrata based upon the degree of differentiation of the residing keratinocytes. The Stratum Basalis (otherwise known as the Stratum Germinativum or Malpighian Layer) is Þrmly anchored by hemidesmosomes to a basement membrane which separates the superÞcial epidermis from the deeper dermis. The basement membrane serves as a formidable barrier separating these anatomic compartments, and is comprised of type IV collagen, laminin, and the glycosaminoglycan, heparin sulfate, all of which are elaborated by keratinocytes, as well as Þbronectin produced by dermal Þbroblasts. The epidermis, like all epithelial compartments, is entirely avascular, so all leukocytes residing in the epidermis derive from the dermal vasculature. The basement membrane is a hindrance to translocation of leukocytes to the epidermis, and dermal leukocytes, when summoned by epidermal cells to the sites of active infection, must force passage through this sturdy barrier. B. Spellberg / Life Sciences 67 (2000) 477Ð502 479 Fig. 1. Normal histology of skin. Commensal ßora are kept at bay by the tough keratin Þbrils created by the death of the underlying squamous epithelium (keratinocytes). The basement membrane separates the epidermis from the underlying papillary dermis. Keratinocytes, lymphocytes and Langerhans cells are depicted in the epidermis, while macrophages, mast cells, Þbroblasts, and blood vessels are shown in the dermis. The Stratum Basalis is comprised of a self-renewing population of relatively undifferentiated squamous epithelial cells. As these cells divide, they push outward on the more superÞcial epidermal substrata. The cells comprising the Stratum Basalis are tightly linked horizontally by desmosomes. Desmosomes are cell to cell junctions comprised of intracellular actin Þlaments linked to a cell surface molecule which spans the cytoplasmic membrane, called desmoglein (27). Desmoglein is a member of the cadherin family of cell adhesins, whose major characteristic is an extracellular domain which homodimerizes in a calcium dependent manner, serving to link adjacent cells. Thus, due to the desmoglein interactions in the desmosomes, the cells of the Stratum Basalis form a horizontal union which is pushed outward and matures in unison. Cell layers that move more superÞcially differentiate together into substrata of mature keratinocytes. Similarly to desmoglein, E-cadherin is a surface adhesin which homodimerizes in a calcium-dependent manner, but is found in epidermal adherens junctions rather than desmosomes (27). E-cadherin is linked, via a membrane anchoring protein called catenin, to the cytoplasmic actin Þlament network. Thus, E-cadherin, like desmoglein, allows tight cell to cell junctions which are anchored via cytoskeletal structural proteins. Intriguingly, E-cadherin is also capable of binding in a heterotypic fashion to a special in- 480 B. Spellberg / Life Sciences 67 (2000) 477Ð502 tegrin molecule called a«b7 (28,29), which is expressed by epidermal T cells and dendritic cells (30,31). Epidermal T cells and dendritic cells (called Langerhans cells) also express E-cadherin, and thus can bind homotypically (via E-cadherin) or heterotypically (via a«b7) to E-cadherin expressed on keratinocytes (28,32). Thus leukocytes can utilize adhesins expressed by keratinocytes to serve as a scaffolding to support their own movement through the epidermis. As discussed below, this likely serves as a crucial mechanism of leukocyte locomotion in times of need. The maturation of keratinocytes coincides with their increasing production of the tough, Þbrous connective tissue from whence they derive their names. Keratin is a sturdy complex comprised of a union of cytokeratin protein strands with keratohyalin. In the Stratum Granulosum substrata, keratin comes to occupy up to 85% of the cytoplasm of the maturing keratinocytes (33). As the keratinocytes continue to progress towards the most superÞcial layer of the epidermis, their lysosomes begin to rupture. The resulting toxic spillage mortally wounds the cells, and initiates a Þnal spontaneous polymerization of the keratin (33), forming the Stratum Corneum layer. In addition, the lysosomal rupture likely accelerates the commensal bacteria-mediated process of lipid degradation into fatty acids. This liberation of free fatty acids creates a low pH (5Ð6) environment in the skin milieu, which itself is a powerful deterrent to bacterial infection (20,34,35). In the deep Stratum Corneum, the lysosomally stricken cells lose their nuclei and their cytoplasm becomes dominated by organized rows of keratin. However, even as the cells begin to deteriorate and lose their classic structure they maintain their desmosomal junctions, so the horizontal keratinocyte layers remain intact. It is these acellular, highly keratinized layers of skin that form the tough battlement which protects the body from the swarm of microbes living just on the other side of the barrier. Indeed the majority of human commensals actually reside in the superÞcial layers of the Stratum Corneum (36,37), kept at bay only by its 10micron-thick layers of keratin. As the epidermal layers move towards the superÞcial portion of the Stratum Corneum, the desmosomes become degraded and the keratinized cells desquamate. The turnover of the keratinized cell layers is an effective mechanism for preventing any microorganisms from gaining a permanent foothold in the ever-renewing skin. As if walking down an up escalator, bacteria which penetrate into the Stratum Corneum are carried back towards the surface by the rising tide of the substrata below. Epidermal cell components Keratinocytes comprise about 90% of the total number of cells found in the epidermis. Aside from their role in the construction and maintenance of the keratin barrier, keratinocytes are biochemically active cells that help support other cell types in the dermal microenvironment. For example, keratinocytes constitutively secrete interleukin (IL)-7 (38), a growth and survival factor that supports epidermal T cell populations. Keratinocytes also secrete macrophage-colony stimulating factor (M-CSF) and granulocyte/macrophage-CSF (GM-CSF) (39,40), which sustain and activate Langerhans cell populations (41,42). About 1% of the cells found in the epidermis are T lymphocytes (43,44). They are frequently found residing in close proximity to the basement membrane, adjacent to Langerhans cells, in the deepest substrata of the epidermis. The vast majority of these T cells are of memory cell B. Spellberg / Life Sciences 67 (2000) 477Ð502 481 phenotype (CD45RO1) (43), and most express the CD8 receptor (45,46), indicating that they are cytotoxic T lymphocytes (CTLs); however CD41 cells are represented as well. Fifty percent of epidermal T cells express the Cutaneous Lymphocyte Antigen (CLA) (47), a Sialyl Lewis-X glycoprotein which serves as a ligand to the endothelial cell adhesin, E-selectin (48). This proÞle of epidermal T cells stands in stark contrast to the normal 2 to 1 ratio of CD41 to CD81 T cells in peripheral blood, and to the fact that most circulating T cells in peripheral blood and lymph are na•ve (CD45RA1) (49). Thus, it is apparent that T cells do not enter the epidermis by random migration. Rather, epidermal T cells are a special population of memory cells which have been given a speciÞc set of instructions to selectively home to the epidermis. In striking contrast to murine models, in human skin ab T cells outnumber gd T cells by a ratio of 10 to 1 (43,50Ð52), which is identical to the ratio of blood ab to gd T cells. Thus, unlike in rodents, where more gd cells than ab T cells can be found in the skin (53,54), human gd T cells do not preferentially migrate to skin. Both ab and gd T cells in human skin express receptors from a skewed distribution of the entire potential pool of Va/Vb and Vg/Vd gene segments (55Ð57). This suggests that these cells are selected to reside in the epidermis because they respond to a limited number of skin-speciÞc epitopes. As mentioned earlier, epidermal T cells are often found in close proximity to Langerhans cells (epidermal dendritic cells) (43), the dominant antigen presenting cells of human skin. Although Langerhans cells express a high density of class I and class II Major Histocompatibility Complex (MHC) molecules, they are also characterized by a high level of expression of CD1 molecules (58,59). CD1 proteins comprise a family of non-polymorphic surface molecules which are capable of presenting glycolipid and other non-protein antigens to T cells (60Ð62). The skewed population of T cell receptors found in human skin indicates that nonpolymorphic, cutaneous-speciÞc antigens stimulate such T cells. Thus these T cells likely represent a population of lymphocytes selected for by their receptorsÕ speciÞcity for evolutionarily conserved antigens of common, skin commensal organisms. These evolutionarily conserved epitopes can be presented by Langerhans cells in the context of classical class I or II MHC or, if they are glycolipid, lipoprotein, or phospholipids, in the context of nonpolymorphic CD1. Langerhans cells are of myeloid lineage, originally migrating to the skin during the Þrst trimester of fetal life (63), and comprise less than 5% of the population of epidermal cells (64,65). Morphologically Langerhans cells possess numerous long, thin dendritic processes, called veils, which wind between nearby keratinocytes. These veils allow Langerhans cells to simultaneously sample antigenic material from a large volume of epidermis. Like epidermal T cells, Langerhans cells are predominantly found near the external surface of the basement membrane. Interestingly, Langerhans cells are often found adjacent to the free ends of cutaneous nerve Þbers (66). Numerous free nerve endings are found in the epidermis (67), consisting of little more than dendrites branching off of slow conducting nerve Þbers found deeper in the dermis. These nerve Þbers, which are unmyelinated, terminate at the epidermal surface of the basement membrane, which, as mentioned above, places them in an ideal position to monitor, and perhaps communicate with, Langerhans cells and T cells. In addition, nerve endings can be found extending superÞcially between keratinocytes through the Stratum Granulosum. Throughout the lower epidermis, nerves are seen to physically contact keratinocytes (67). Although these free nerve endings are 482 B. Spellberg / Life Sciences 67 (2000) 477Ð502 known to act as afferent sensory receptors, recent studies have demonstrated that they are able to elaborate neuropeptides which modulate local immunologic function (see below) (68,69). The dermis Dermal histology In contrast to the epidermis, the dermis is a relatively acellular layer of skin dominated by connective tissue matrix. The dermis carries out several functions crucial to the integrity of the overlying epidermis. First, the tough mesenchymal matrix which comprises the dermis provides a strong foundation into which the epidermis is anchored. Second, the dermis is a highly vascularized compartment, and nutrients are free to diffuse through its matrix in order to reach the avascular epidermis. Third, the dermis is the port of call for any cell crossing to or from the epidermis. Thus trafÞc of leukocytes to and from the epidermis is coordinated by the dermis. Like the epidermis, the dermis can also be divided into histologic substrata (26). However unlike the multi-stratiÞed, ever changing epidermis, the dermis is comprised of only two substrata, neither of which are in a constant state of developmental ßux. The more superÞcial layer is the papillary dermis. The architectural support for this layer derives from interlacing, Þnely woven type I collagen Þbrils, and intertwined in this loose collagen matrix are Þbroblasts and leukocytes, superÞcial capillaries, and lymph channels. Numerous myelinated and unmyelinated nerve Þbers run through the dermis, heading superÞcially towards the basement membrane. Many of these axons, containing the neuropeptide substance P, entwine the dermal capillaries in a Þne web of Þbers (70). The deeper reticular layer of the dermis is far more sturdy, akin to the bedrock upon which the entire foundation of skin rests. In contrast to the lacy collagen of the papillary layer, the collagen Þbrils of the reticular dermis are extremely thick and intertwined in a Þrm, robust architectural network. Arterioles and venules course through the reticular layer bearing branches which peel off and head superÞcially towards the papillary capillary plexuses. Numerous leukocytes can be found in the reticular layer, many of which are tissue macrophages or mast cells, the latter of whose duty it is to regulate arteriolar and venular tone. Both layers of the dermis contain, in addition to collagen, other structural components. The other major structural protein of the dermis is elastin. Elastin helps give skin its characteristic ßexibility and is responsible for the skinÕs remarkable ability to snap back into place when mechanically displaced. Although collagen and elastin form the backbone of the dermal matrix, it is the mucopolysaccharide, hyaluronic acid, which forms the gel component of the matrix. Hyaluronic acid is a ubiquitous, massive glycosaminoglycan polymer which carries a strong negative charge due to its numerous hydroxyl and carboxyl side groups. Due to this charge, hyaluronic acid is extremely hydrophilic and is a powerful retainer of water molecules, which carry a positive-negative dipole. The hyaluronic acid matrix is thus the main storage site for interstitial ßuid, and the water pressure in the matrix helps support the overlying skin structures. Dermal cell components Fibroblasts are the workhorses of the dermis, laying down all of the type I collagen and elastin Þbers which impose structure upon the glycosaminoglycan matrix. Fibroblasts are found dispersed throughout the dermal mesenchyme, and recently have been shown to be capable of signiÞcant production of cytokines (71,72). In addition, they secrete growth factors B. Spellberg / Life Sciences 67 (2000) 477Ð502 483 necessary for the survival of leukocytes and keratinocytes (73), and they are greatly responsible for the repair of damaged tissue by production of ground matrix and structural proteins. Dermal T lymphocytes comprise 98% of all skin T cells (i.e. only 2% of skin T cells are to be found in the epidermis) (45,52). Dermal T cells, like their epidermal brethren, are mostly memory cells with lineage-restricted T cell receptors, and are typically found grouped in perivascular cuffs, surrounding the small venules and arterioles which course through the dermis. However in contrast to epidermal T cells, dermal CD41 and CD81 cells are found in equivalent ratios (45). The signiÞcance of this difference between epidermal and dermal distributions of CD4 and CD8 T cells is unclear, however it might indicate that CD8 populations in the epidermis are continually being renewed due to low level exposure to cutaneous viruses or other antigens which are presented in the context of class I MHC (typically intracellular antigens) but not class II MHC (typically endocytosed extracellular antigens). Unlike T cells, healthy human skin does not contain B lymphocytes. As well, the human dermis, unlike the epidermis, lacks cells possessing the classic subcellular characteristics of Langerhans cells. However the dermis does possess its own population of specialized dendritic cells (74Ð76). Like Langerhans cells, these dermal dendritic cells express high levels of class II MHC and CD1 molecules, and are extremely efÞcient at antigen presentation. The exact relationship between these dermal dendritic cells and Langerhans cells is not established. Macrophages comprise the bulk of the leukocytes in the papillary dermis (77). These cells, derived from blood monocytes which extravasate across dermal venular walls, are long-lived and end-differentiated (i.e. incapable of cell division). They are often found in close proximity to the dermal side of the basement membrane. These cells scavenge the dermal microenvironment, clearing away debris and participating as effectors when other cells call them to the sites of danger. Mast cells are long-lived tissue leukocytes whose major function is to regulate vascular tone and initiate IgE-mediated allergic or inßammatory responses to recognized antigens. They are typically found surrounding the end arterioles and venules, as well as the capillary beds in between. In addition, they are often in close contact with dermal nerve terminals, some of which are afferent vagal nerve axons (78). From this venue, they can alter local vascular ßow by secreting effector molecules that cause vasodilation or vasoconstriction. In addition, they mediate direct control over vessel permeability, and can regulate the balance of ßuid exchange between the intravascular compartment and the interstitial compartment. Perhaps the key, and for that matter unheralded, regulatory cell type in the skin immune system is the microvascular endothelial cell. These cells line the walls of the dermal venules and arterioles, and form the capillaries running through the papillary dermal compartment. It is across these cells that leukocytes must traverse in order to reach extravascular destinations. Endothelial cells play a crucial role in the transduction of appropriate chemotactic and stimulatory signals from the dermal microenvironment into the bloodstream (see below). Immune functions of the skin Initiation of the innate immune response Keratinocytes are eminently capable of inducing immune responses (Fig. 2), as they store large quantities of IL-1a in their cytoplasm (79,80). Thus, any keratinocyte that is damaged 484 B. Spellberg / Life Sciences 67 (2000) 477Ð502 Fig. 2. Initial cytokine cascade of a skin immune response. Primary cytokines, such as IL-1, IL-6, IL-8, and TNF, are depicted with solid arrows, while secondary cytokines such as IFN-g are depicted with broken arrows. 1) Direct trauma, as from a foreign object, disrupts the cell membrane of keratinocytes, spilling pre-existing IL-1 into the local microenvironment, while the break in the keratin battlement exposes the epidermis to microbial invasion. Keratinocytes stimulated by paracrine IL-1 produce more IL-1, as well as IL-6, IL-8, TNF, and other cytokines that diffuse throughout the epidermis. 2) Exposure of Langerhans cells and intraepithelial T cells to invading microbes and primary inßammatory cytokines induce their secretion of secondary cytokines such as IFN-g. 3) Cytokine diffusion and dermal nerve dendrites contacting the epidermis transduce the activating signals into the papillary dermis. Fibroblasts, mast cells, and endothelial cells are all activated in this manner. 4) Cytokine diffusion into the reticular dermis recruits deep macrophages and initiates vascular changes. 5) Endothelial activation transduces the tissue danger signal into the vascular compartment. leaks IL-1 into the surrounding tissue, initiating an inßammatory cascade. IL-1a, therefore, is an instant danger signal released by keratinocytes after the slightest insult (81,82). Neighboring keratinocytes respond to the IL-1 by production of further IL-1a, as well as IL-1b, tumor necrosis factor (TNF)-a and IL-6, amplifying the initial danger signal (83,84). Production of these cytokines can also be directly stimulated by ultraviolet injury or by microbial infection (85,86). In addition, keratinocytes produce chemokines, initially of the CXC family, including IL-8 and members of the GRO family (87,88). The expression of CXC chemokines leads to a predominant early recruitment of granulocytes rather than mononuclear inßux (See Fig. 3). Keratinocytes concurrently secrete growth factors, such as M-CSF, GM-CSF, IL-7, and IL-15, to support the survival and induce the activation of leukocytes responding to the chemokines (38Ð41,89,90). The secreted cytokines diffuse across the basement membrane to reach the dermis (91). Dermal Þbroblasts amplify the epidermal response, as they are stimulated by keratinocyte cytokines to produce IL-1, IL-6, TNF-a, and chemokines of their own (72). Cytokines from both of these sources then diffuse to the dermal vasculature (92,93). In this way, endothelial cells are made aware that an inßammatory response is underway in their vicinity. The chemokines IL-8 and MIP-1a, derived from damaged or activated keratinocytes and Þbroblasts, similarly diffuse, creating a concentration gradient pointing directly back towards the B. Spellberg / Life Sciences 67 (2000) 477Ð502 485 Fig. 3. Reinforcement of the immune response. 1) Activated endothelial cells express E- and P-selectin and secrete CXC chemokines to recruit neutrophils from the vascular compartment. Mast cell degranulation aids this process by inducing vasodilation. 2) Extravasated leukocytes follow the concentration gradient of the diffused cytokines towards the nidus of danger. 3) Macrophages and recruited neutrophils secrete metalloproteinases which degrade the basement membrane, allowing them to pass into the epidermis. Meanwhile Langerhans cells cross the basement membrane in the opposite direction, carrying antigenic material into the lymphatic system to present to na•ve T cells in the lymph nodes. 4) At the site of danger the early responding macrophages stem the tide of microbial invaders, while intraepithelial T cells continue to blare the alarm, informing the immune system that local defenses will not be sufÞcient to stave off the bacterial assault. nidus of danger. The endothelial cells actually pinocytose keratinocyte-derived chemokines on their abluminal surface, translocate the chemokines across their own cytoplasm in protected vesicles, and express the chemokines on their luminal surface in membrane-bound forms (94). The result is a membrane-bound chemokine gradient within the blood vessel, as if a yellow-brick road pointing towards the site of danger. In this way the endothelium transduces an extravascular chemokine gradient into the intravascular compartment where leukocytes can home in on the danger signal. In the meantime, epidermal T cells, the majority of which are resting memory CTLs, are capable of responding to the local disturbance almost immediately. There are several ways for local lymphocyte stimulation to occur. Langerhans cells accumulate new antigen from the necrotic material released during the death of the keratinocytes, and if microorganisms caused the initial insult, the Langerhans cells accumulate their antigens as well. Evidence indicates that some receptor lineage-restricted epidermal T cells are reactive to self-phosphoproteins normally sequestered in the cytosol of host cells but released upon cell damage or tissue necrosis (95). Therefore, along with foreign microbial antigens, host phosphoprotein antigens liberated by keratinocyte destruction can be used to activate immediate local T cell responses. The Langerhans cells, which tend to localize adjacent to T cells, are in an ideal position to present such antigen to the memory T cells, activating them. If sufÞcient cytokines are present in the epidermal environment, this presentation can occur in situ in the epidermis, without requiring 486 B. Spellberg / Life Sciences 67 (2000) 477Ð502 the Langerhans cells to migrate to the draining lymph node (96). Langerhans antigen presentation can be by classical class I or class II MHC molecules or by CD1 presentation of conserved glycolipid or lipoprotein antigens. Finally, epidermal gd T cells, like B cells, are capable of directly binding to small phosphatecontaining antigens with their receptors (97Ð99). These cells do not require antigen presentation in the context of MHC or CD1, and serve as a primitive, immediate response element, recognizing conserved phosphoprotein or phospholipid antigens of microbes and necrotic human tissues. Upon stimulation of local T cells, the cytokine milieu of the epidermis becomes signiÞcantly more complex. It is unlikely that the sparsely numbered epidermal T cells would be able to fend off a signiÞcant threat by themselves. Rather, these cells should be thought of as an immediate response ampliÞer, somewhat akin to an air-raid klaxon. The cytokines produced by these T cells include interferon (IFN)-a, IFN-g, IL-6, and more TNF-a, as well as other pro-inßammatory cytokines such as IL-17, which upregulates keratinocyte secretion of IL-6 and IL-8, and stimulates keratinocyte expression of adhesion molecules such as ICAM (100). The early elaboration of IFN-a and IFN-g are of critical importance to the developing immune response, as they send speciÞc signals directing the adaptive immune response to adopt a pro-inßammatory posture (see below). The overall result of this elaboration of cytokines is to further signal nearby dermal endothelial cells that leukocytes need to be recruited from the vascular compartment as soon as possible. In the meantime, local defenses are quickly augmented by the recruitment of dermal macrophages, like supporting tank divisions, from just beneath the basement membrane (See Fig. 3). Diffusion of lymphocyte- and keratinocyte-derived IFN and growth factors across the basement membrane initiates priming of these macrophages (101,102). As mentioned, the early production of IFN-g by epidermal and dermal T-cells (103Ð105) is of particular importance, because it primes macrophages and dendritic cells for production of IL-12 (106Ð108). SpeciÞc antigenic fractions of microbes, including lipopolysaccharide (LPS), gram-positive cell wall components, and unmethylated CpG-rich prokaryotic DNA also directly stimulate production of IL-12 and other pro-inßammatory cytokines from phagocytes (109Ð112). This is crucial to the development of speciÞc cell-mediated adaptive immunity, as the presence of IL-12 during antigen presentation to T cells induces differentiation to the pro-inßammatory Th1 phenotype (108,113Ð115). Under the inßuence of epidermally-derived cytokines and antigenic stimuli, macrophages mobilize their cytoskeletons to become motile (116) and upregulate their respiratory burst in preparation for combat (101,117). They also secrete metalloproteinases and other degradative enzymes (118,119) which allow them to slice through the collagen and other structural components of the basement membrane so that they can cross into the epidermis (120,121) (See Fig. 3). By utilizing adhesins expressed on the leading edges of their lamellipodia, such as E-cadherin and LFA-1, local macrophages can use ligands expressed on activated keratinocytes, such as E-cadherin and ICAM-1, to pull themselves through the epidermis. In this way, they successfully crawl to the danger site by following the chemokine gradient to its source. Langerhans cells also become activated by the local efßux of cytokines (122), and are spurred to translocate across the basement membrane in the opposite direction from the mac- B. Spellberg / Life Sciences 67 (2000) 477Ð502 487 Fig. 4. Switch to Adaptive Immunity. Twenty-four to 36 hours after the initiation of the innate immune response, continued inßammatory signals induce activation of adaptive immunity. 1) Intense phagocytic activity at the nidus of infection elicits dense secretion of inßammatory cytokines from neutrophils and macrophages. Furthermore, leukocyte eructation of digested microbial fragments and extracellular degradation of microbes exposes host cells to highly stimulatory microbial antigens. Prolonged exposure to such danger signals causes secretion of adaptive cytokines such as IFN-g and CC chemokines (e.g. RANTES). 2) Newly arrived memory T cells activated in the blood or lymph add to the IFN-g rich milieu. 3) Prolonged exposure of endothelial cells to danger signals induces transcriptional changes. The selectins are downregulated and ICAM/VCAM, which preferentially bind to mononuclear cells, are expressed. As well, endothelial cells switch from secreting CXC chemokines (e.g. IL-8) to CC chemokines, thereby recruiting mononuclear cells at the expense of granulocytes. rophages. The Langerhans cells leave the epidermis and enter the local lymphatic channels in the dermis. Activated Langerhans cells lose their expression of E-cadherin as they leave the epidermis (31), consistent with their need to exit its cadherin-rich environment. Meanwhile Langerhans cells acquire expression of a6 integrins, which mediate adhesion to laminin components of the interstitial matrix, allowing migration through the epidermis, across the basement membrane, and through the dermis to lymph channels (123) (See Fig. 4). Once the Langerhans cells enter the lymph channels, they down-regulate the now useless a6 integrins and upregulate a4 integrins which they utilize to interact with structural motifs in the lymph nodes (123,124). At the same time, in preparation for their need to stimulate na•ve T cells, Langerhans cells undergo signiÞcant upregulation of surface expression of class II MHC and the costimulatory molecules, B7-1 and B7-2 (125,126). The dermal lymph channels carry Langerhans cells to the draining lymph nodes, where they encounter na•ve T cells waiting to be exposed to an antigen/MHC complex which their T cell-receptor recognizes (127). Once na•ve T cells are primed in draining lymph nodes by Langerhans cells, the T cells begin to express the Cutaneous Lymphocyte Antigen (CLA), which allows them to home to skin when they eventually enter systemic circulation (128). 488 B. Spellberg / Life Sciences 67 (2000) 477Ð502 The na•ve T cells will take days to become activated, enter the cell cycle, proliferate, and then home to the site of danger. However, memory cells circulating through the lymphatics which recognize the antigen/MHC complex expressed on the Langerhans cells are capable of full mobilization within 24 hours. These cells are already programmed to return to dermal compartments once they become reactivated. In the meantime, the dermal vasculature will have started the process of calling in systemic reinforcements. Reinforcement of the innate immune response The keys to the arrival of reinforcements to defend the skin are the dermal microvascular endothelial cells (Fig. 3). As the endothelium senses keratinocyte- and Þbroblast-derived IL-1, TNF-a, and IL-6, several actions are simultaneously set in motion. First, the endothelial cells are stimulated to mobilize their Weibel-Palade bodies, resulting in immediate P-selectin expression on their cell surface (129,130). The endothelium also secretes arachidonic acid metabolites, including prostaglandin E and leukotrienes, which result in local vasodilation (131,132). Endothelial cell contraction opens pores in the papillary capillaries and the reticular layer venules, creating leaky vessel walls (133,134). This allows plasma protein to escape from the vasculature, and eases the extravasation of leukocytes into the dermal interstitium. The nervous system aids in this response. The free-ended nerve Þbers in the epidermis are capable of modulating vascular tone in inßammatory reactions via secretion of a variety of factors, including nitric oxide and substance P (135,136). Nociceptive Þbers stimulated by local cytokines send afferent signals to the CNS, informing the brain that an inßammatory response is underway (137,138). Efferent signals returning to the dermis from the central nervous system serve to modulate the immune response, either in a stimulatory (139,140) or inhibitory (141,142) fashion. The duration of afferent signaling is the determining factor in whether the resultant efferent response is stimulatory or inhibitory, with acute signals eliciting a stimulatory efferent response, and prolonged, persistent signaling eliciting an inhibitory efferent effect. Acutely, substance P released from vascular-associated nerve Þbers acts on endothelial cells to induce vasodilation and increased vascular permeability (70,135). As well, substance P and other neuropeptides induce mast cell degranulation (143Ð145), as do complement factors (146,147) which ooze into the interstitium after leaking through the porous vasculature. The resultant liberation of histamine and other mast cell mediators ampliÞes the local vasodilation and vascular leakage, further easing leukocyte efßux into the dermal interstitium (148). Furthermore, mast cell granules contain preformed TNF-a (149) and upon activation mast cells can be induced to secrete chemokines (150). The vasodilation increases local blood ßow, allows leakage of serum into the interstitium, and drops intracapillary and intravenular pressures, allowing blood pooling to occur. The drop in intraluminal pressure diminishes the shear forces associated with blood ßow, allowing easier leukocyte extravasation. Thus, the four cardinal signs of inßammation, rubor (redness), calor (warmth), dolor (pain), and tumor (swelling), are the result of increased vascular permeability secondary to endothelial cell interactions with mast cells, cutaneous axons, and skin leukocytes and keratinocytes. The Þnal result of all of the above is that leukocytes are able to migrate away from the center of the blood vessel lumen and reach the vessel walls, where they can bind to activated endo- B. Spellberg / Life Sciences 67 (2000) 477Ð502 489 thelial cells without being swept away by high shear forces. Endothelial selectin binding to counter ligands on leukocytes recruits the white blood cells to the local areas of danger. The early proÞle of endothelial cell adhesin expression, predominantly P-selectin and then E-selectin within the Þrst 8 hours after endothelial stimulation, favors the binding of granulocytes and memory lymphocytes (128,151Ð154) (See Fig. 3). These adhesins act in concert with the initial expression of IL-8, which favors the activation and recruitment of neutrophils out of the blood and into the dermal tissue (155). From there, the neutrophils migrate along the diffusible chemokine concentration gradient, and, like dermal macrophages, express extracellular proteases to cleave a path through the basement membrane and enter the epidermis (156). As neutrophils migrate through different tissue microenvironments, they undergo significant changes in their shape and cytosolic composition. Calcium-regulated cycling of actin from its Þbrous to globular conformation allows the neutrophil to extend long cellular projections, called lamellipodia or pseudopods (157). Like amoebae, neutrophils elongate these projections to seek out passable tracts along the chemokine gradient ahead of them. By utilizing surface adhesins on these pseudopods, the neutrophils are able to generate traction to pull themselves through the tissue microenvironment. Furthermore, neutrophils alter their intracellular electrolyte concentrations, increasing their osmolality to precipitate osmotic water uptake which markedly expands their cytoplasmic volume (158,159). This accrual of cytoplasm allows the neutrophils to maintain turgor pressure in their extended pseudopods. Neutrophil movement through the dermal interstitium is dependent upon a coordinated series of surface protein ligations on their pseudopods. In order to successfully adhere to ligands in the dermis, the neutrophils must undergo cell surface changes as they exit the vasculature. For example, PECAM, which is crucial to neutrophil extravasation through the vascular wall, is down-regulated as the neutrophils exit the blood vessel and enter the dermis (160), where there are no PECAM ligands. The b2-integrin, Mac-1 (CD11b/CD18), is a crucial adhesin which neutrophils use to bind to dermal Þbroblasts (161). In inßamed tissues, neutrophil LFA-1 also plays an important role, as stimulated Þbroblasts upregulate their expression of ICAM-1, the LFA-1 counterligand (161). Furthermore, the b1-integrins VLA-4, VLA-5, and VLA-6 play a signiÞcant role in neutrophil migration through the dermal tissue (162). By utilizing these surface adhesins, neutrophils are able to use Þbroblasts embedded in the dermis as stepping stones through the dermal Þbrous matrix. However, neutrophil progress is not strictly dependent upon encountering the sparse cells which pepper the dermal matrix. Indeed, neutrophils are able to directly utilize components of the connective tissue matrix as scaffolding for their amebic movement. By extending pseudopods into crevices within the Þbroelastic matrix, neutrophils are able to generate sufÞcient traction to propel themselves forward along the chemokine gradient (163,164). Neutrophil pseudopods express appropriate adhesins on their surfaces to allow Þrm binding to the matrix protein components. Among the matrix components utilized by neutrophils are collagen, Þbronectin and laminin (164), as well as entactin (165). As described above, the histology of different layers of skin are dramatically different, so the components and density of the matrix across which the neutrophils must trek on their way from the deep dermal vasculature to the epidermis is highly variable. However, neutrophils are capable of detecting the 490 B. Spellberg / Life Sciences 67 (2000) 477Ð502 tensile strength of the underlying matrix and adjusting their own force output to maintain their progress despite the Þrmness or inÞrmity of their supporting scaffold (166). Movement inhibits neutrophils from expressing degradative enzymes (167). Thus, neutrophils must be stationary prior to their excretion of the metalloproteinases which they utilize to degrade the basement membrane. This makes teleological sense, as such a system not only prevents neutrophil degradation of endothelium prior to diapedesing across the subendothelial basement membrane, but also prevents excessive dermal destruction prior to the neutrophils reaching the dermal-epidermal border. Once the neutrophils are no longer able to propel themselves through the dermal matrix along the chemokine gradient (i.e. they abut against the basement membrane and therefore cannot move forward), they are triggered to secrete enzymes to degrade the obstacle in their path. Having crossed the defunct basement membrane, neutrophils are able to continue their migration even through the tightly adherent epidermis. They do so by extending their pseudopods in between the gap junctions which join neighboring keratinocytes (167). As the neutrophils slide in between the epithelial cells, their own surface adhesins are capable of displacing those of the neighboring epithelium. As if undoing a zipper, the neutrophils squeeze in between the keratinocytes, adhering to both of the cells which had previously been joined. As the neutrophils move forward, the keratinocytes merely rejoin, or re-zip, along the trailing neutrophil edge (167). With the entry of neutrophils into the area of danger, the battle is begun in earnest. The rich cytokine milieu at the danger site is enough to prime the neutrophils for combat. The IL-1, TNF-a, IL-6, GM-CSF, and IFN activate the respiratory burst (168,169) and upregulate surface markers for phagocytosis, such as the complement receptor 3 (CR3) and Fcg receptors (170Ð173). Serum protein efßuent from the leaky dermal vasculature diffuses into the dermis, allowing complement to become active against any prokaryotic cell wall components present (174). Furthermore, keratinocytes actually produce some complement components, and their secretion of these components is sufÞcient to allow C3 activation to occur along the alternative pathway even if serum-derived complement factors are not present (175). The deposition of complement on microbial cell surfaces opsonizes the particles for ease of neutrophil phagocytosis. Any preformed antibodies circulating in the serum also escape the leaky vasculature, further aiding opsonization. As well, both the complement degradation products, C3a and C5a (176), and phagocytosed microbial particles (177) induce neutrophils to secrete IL-8 as a signal that further granulocyte reinforcements are needed. With the convening of phagocytes, cytokines, and opsonins at the danger site, enemy microbes are cleared at a rapid pace. Each activated neutrophil can phagocytose 1 bacteria per minute, and can ingest a total of 50 gram-positive or gram-negative organisms (178). The thousands of neutrophils arriving from leaky dermal vessels are therefore capable of clearing away much of the initial inßammatory stimulus. The residue of that initial stimulus, however, remains in the epidermis. This includes the plethora of pro-inßammatory cytokines elaborated by the keratinocytes, Þbroblasts, dermal T cells, macrophages, and neutrophils, as well as the detritus of destroyed microbes. At this point the neutrophils and the supporting cells must decide whether further upregulation of inßammation is warranted or if the innate response has been successful and should be shut down. B. Spellberg / Life Sciences 67 (2000) 477Ð502 491 Terminate inßammation or call in the lymphocytes? Keratinocytes constitutively secrete the dominant suppressive cytokine IL-10 (179), and they upregulate their secretion of this cytokine concomitantly with their initial expression of IL-1 (180,181). In addition, the keratinocytes also secrete IL-1 receptor antagonist (IL-1ra) (181,182), which abrogates the effects of the IL-1 present in the epidermis. Along these lines, both keratinocytes and cutaneous nerves express a-melanocyte stimulating hormone (a-MSH) (183Ð186). a-MSH is a potent anti-inßammatory molecule which directly antagonizes IL-1b by blocking its binding to the IL-1 receptor (187,188), and markedly upregulates secretion of IL-10 secretion by phagocytes (189). Furthermore, likely via its induction of IL-10, a-MSH inhibits phagocyte production of IL-12 and IFN-a and lymphocyte production of IFN-g (190,191), and suppresses phagocyte expression of class I MHC and the B7 costimulatory molecules (192). Finally, a-MSH abrogates neutrophil chemotaxis in response to IL-8 concentration gradients (193). As expression of a-MSH is induced by IL-1, TNF-a, or trauma (184,186), a-MSH likely represents a safeguard for keratinocytes in protecting themselves against Òfriendly ÞreÓ from an overactive cell-mediated immune response. Resolution of inßammation also depends upon local production of transforming growth factor (TGF)-b by Þbroblasts, macrophages, and lymphocytes (194). TGF-b is a potent terminator of inßammatory responses, mediating such diverse effects as inhibiting expression of co-stimulatory molecules on antigen-presenting cells (195), suppressing secretion of inßammatory cytokines (196) and survival factors for inßammatory cells (197), and inducing secretion of IL-1ra (198). As well, the presence of TGF-b during T lymphocyte activation induces antigen-speciÞc T cell anergy (199Ð201). Furthermore, in order to protect the stromal matrix around the site of inßammation, TGF-b suppresses secretion of tissue proteases by migrating phagocytes (202) while stimulating Þbroblast secretion of tissue inhibitors of metalloproteinases (TIMPs) (203,204). TGF-b also suppresses chemokine production and adhesion molecule expression by endothelial cells (205) and inhibits inßammatory cell proliferation (197,206Ð208). Thus TGF-b directly suppresses accumulation of inßammatory cells, allowing their spontaneous apoptosis to clear them from the extravascular compartment. On the other hand, TGF-b stimulates proliferation of Þbroblasts (209,210) and stromal supporting cells (211), and induces mesenchymal cells and Þbroblasts to lay down new collagen and structural proteins to repair the damage wrought by local inßammation (204,209,210). As the invaders are cleared by the innate immune cells, the microbial stimulus for continued inßammation wanes and neutrophils and macrophages down-regulate their production of pro-inßammatory cytokines while maintaining production of IL-10. Newly arriving T-cells Þnding a low IFN-g environment are more likely to differentiate into Th2 cells secreting IL-4 and IL-10 (212), which are suppressive to inßammation. Furthermore, the presence of IL-4 and IL-10, combined with a dearth of IFN-g, induces differentiation of T-cells to the socalled Th3 type, secreting high levels of TGF-b (213). Thus keratinocyte- and leukocytederived IL-4, IL-10, TGF-b, and IL-1ra tilt the balance towards shutting down inßammation. On the other hand, if the danger stimulus is too robust to be cleared by phagocytes within a short period of time, innate immune cells continue to produce pro-inßammatory cytokines, overwhelming the suppressive effects of IL-10, TGF-b, and IL-1ra. After approximately 12Ð 24 hours, the threshold for activation of the adaptive immune response is crossed, ushering in the next phase of inßammation. 492 B. Spellberg / Life Sciences 67 (2000) 477Ð502 Initiation of the adaptive immune response An inßammatory state that continues beyond 24 to 36 hours induces the onset of an adaptive immune response (Fig. 4). Due to alterations in transcriptional patterns, the proÞle of both chemokine and cell surface adhesin expression in the local area of danger changes. The presence of IFN-g in the epidermis stimulates leukocytes and Þbroblasts to express CC chemokines (214), particularly MCP-1 (215), MIP-1 (216), and RANTES (217,218), while suppressing the production of CXC chemokines such as IL-8 (216,217). CC chemokines tend to summon mononuclear cells rather than neutrophils, but there is some variation on the theme. In fact MIP-1a is capable of inducing chemotaxis of both neutrophils and lymphocytes (219,220). This indicates that while MIP-1a is structurally a member of the CC family, it should be considered a ÒhermaphroditicÓ chemokine which serves to bridge the gap during the switch from CXC- to CC-mediated responses. While MCP-1 is chemotactic for monocytes and lymphocytes, RANTES is signiÞcantly more selective. In combination with preferential binding to endothelial adhesins (see below), RANTES is capable of selectively summoning pro-inßammatory Th1 lymphocytes, and not inßammatory-suppressive Th2 cells, to areas of inßammation (221,222). This is likely due to the preferential expression by Th1 cells of RANTES receptors such as CCR5 (223). In a feed-forward loop, Th1 cells produce IFN-g which drives RANTES production by lymphocytes and macrophages (224), which then selectively summons additional Th1 cells to the area of danger. Thus the elaboration of IFN-g, a Th1 cytokine product, indicates to the immune system that additional Th1 support is needed. As mentioned, endothelial cell adhesins are also critical to the selection of leukocyte extravasation. Indeed, several hours after stimulation, endothelial cells lower their expression of E-selectin, upregulate their ICAM-1 expression, and begin expressing VCAM (225), resulting in diminished neutrophil diapedesis across the vascular lumen. While neutrophil accumulation wanes, mononuclear cells, including lymphocytes and monocytes, are recruited into the area of danger. Thus the early, predominantly neutrophilic response converts into a subacute/chronic mononuclear cell inßux (See Fig. 4). Clinical pathologists have, for decades, determined the temporal nature of inßammatory lesions based upon this switch in leukocyte migration (226), whose mechanism is only recently understood. In addition to its role in leukocyte recruitment, exposure of the endothelium to IFN-g from the site of danger causes the endothelium to express class II MHC (227Ð229). Endothelial cells are capable of phagocytosing any antigen which migrates into the dermis from the site of danger, and presenting this antigen in an MHC-restricted fashion to T cells (230,231). The upregulated ICAM and LFA molecules on the endothelium provide costimulation via binding to LFA and CD2 on T cells (232,233). Thus in the presence of IFN-g, microvascular endothelial cells are capable of acting as professional antigen presenting cells to circulating T cells. Importantly, there remain several safeguards to shut down the immune response even as the mononuclear inßux begins. For example, like endothelium, keratinocytes are induced to express class II MHC molecules by IFN-g, and are capable of presenting antigen to incoming ab T cells (234). In distinct contrast to endothelium, however, keratinocytes are unable to express sufÞcient costimulatory molecules to drive T cell activation, and the result of keratinocyte presentation of antigen to T cells is T cell anergy or deletion (235). Therefore if the B. Spellberg / Life Sciences 67 (2000) 477Ð502 493 degree of the innate inßammation is insufÞcient to warrant activation of the adaptive immune response, keratinocytes are able to abort the adaptive response by competing with phagocytes for presentation of antigen to T cells. If the threshold of adaptive immune response activation is crossed, a second safeguard remains. This involves the differentiation of newly arriving T lymphocytes into either the pro-inßammatory Th1 subtype or the anti-inßammatory Th2 subtype. As discussed earlier, early response T cells are prone to the secretion of high levels of IFN-g and TNF-a (103Ð 105). The presence in the skin of IFN-g, IFN-a, and TNF-a, as well as microbial particles such as LPS, gram-positive cell wall fractions, and prokaryotic DNA, induces macrophage and dendritic cell production of IL-12 (106,108,109). The IL-12 induces newly activated T-cells to differentiate into the Th1 phenotype, resulting in increased local secretion of IFN-g. In a powerful feed-forward loop, this burst in local IFN-g production further serves to stimulate local phagocyte activity, as well as inducing local expression of RANTES and E-selectin, preferentially summoning additional Th1 cells to the danger area. However, there are 2 potential categories of danger signals which favor the outcome of a Th2 response in the skin. If the stimulus initiating the danger response is too bulky to be easily phagocytosed (e.g. hyphal fungi, helminths, large foreign bodies, etc.) the leukocytes will initiate the Òfrustrated phagocyte complexÓ of activities (236Ð238). This induces their production of IL-10 preferentially over IL-12 (239), particularly if stimulatory microbial components such as prokaryotic DNA, LPS, and gram-positive cell wall fractions are not present. Entering T-cells will Þnd a low IFN-g, higher IL-10 environment, favoring antibody-based Th2 outcomes. As well, danger signals which preferentially engage mast cells without stimulating phagocytes (typically IgE-mediated reactions, such as allergens or metazoal parasites), induce liberation of large amounts of IL-4 from dermal mast cells (150,240). The presence of IL-4 and dearth of IL-12 will induce incoming T cells to differentiate to a Th2 phenotype. Induction of Th2 cytokine proÞles when danger stimuli are too large to be removed by phagocytosis is teleologically consistent. Antibody production stimulated by the Th2 response favors destruction of non-phagocytosable targets such as metazoa and Þlamentous fungi because it allows leukocytes to damage large organisms via antibody-dependent cell-mediated cytotoxicity (ADCC) (241). ADCC is essentially antibody-targeted Òfrustrated phagocytosisÓ; it involves leukocyte exocytosis of toxic cytoplasmic granules, often containing small cationic molecules (242Ð245), directly onto invading pathogens. This granule-exocytosis is stimulated by the inability of the leukocyte to ingest its target (246Ð248). Eosinophils (241), neutrophils (249), Natural Killer (NK) cells (250,251), and monocyte/macrophages (252Ð 254), are all able to utilize antibody bound to Fc receptors on their own cell surfaces to target their excretion of these toxic granules directly onto the surface of the non-phagocytosable stimulant (241,248,255). This allows leukocytes to damage or destroy invading pathogens or inert foreign bodies even when they are too large to be internalized and digested. Resolution of the cutaneous immune response Resolution of the inßammatory response occurs via several mechanisms. First, the continued presence of phagocytes begins to eliminate more and more of the invading microbes, diminishing host cell exposure to their innately stimulatory antigenic fractions. The resultant 494 B. Spellberg / Life Sciences 67 (2000) 477Ð502 waning secretion of pro-inßammatory cytokines allows the persistent IL-10 and other suppressive molecules (e.g. IL-1ra, a-MSH and TGF-b) to begin to tip the balance in favor of completely shutting off inßammation. Second, as the cytokine milieu changes from one of a pro-inßammatory nature to one of IL10 and TGF-b dominance, T cell differentiation will be directed towards the anti-inßammatory Th2 or Th3 subtype (213). The Th2 pattern of high levels of secreted IL-4 and IL-10 favors resolution of cell-mediated immune functions by suppressing pro-inßammatory cytokine production (256), antigen presentation (126,257), and lymphocyte chemotaxis and adhesion molecule expression (258), and by inducing functional anergy of pro-inßammatory cells (259Ð261). As discussed above, TGF-b also inhibits inßammatory activities, ranging from cytokine production to lymphocyte stimulation. These mechanisms are generally sufÞcient to appropriately quell inßammatory responses as the danger is eliminated by the immune system. Finally, if the danger stimulus cannot be cleared within days to weeks, prolonged inßammation results in abscess formation and scarring. Macrophages, particularly under the inßuence of Th2 cytokines such as IL-4 (262) and IL-10 (263), begin to fuse into giant cells, and a ÒgranulomatousÓ response is initiated. Under the stimulus of TGF-b (264), giant histiocytes and recruited Þbroblasts begin to wall off the site of inßammation by laying down thick strands of connective tissue. Eventually the inßammatory site is completely enclosed in a sphere of tough, Þbroproteinaceous material, and an abscess is formed. As well, persistent elaboration of TGF-b by chronically stimulated mesenchymal cells and leukocytes induces local Þbrosis to scar-down tissue damaged during the inßammatory response. Conclusions Serving at the hinterlands of the human host, the skin is a constantly embattled citadel comprised of a variety of cell types, housing both a permanent garrison of immune cells as well as temporary reinforcements in times of need. Immunologists tend to view the immune system with a ÒleukocentricÓ bias. However, when immunity is examined at the organ level, it becomes clear that leukocytes cannot function without constant support from parenchymal tissues. The skin is an excellent example of this ÒholisticÓ model of immunity; local immune responses could not be initiated, developed, sustained, or turned off without input from nonleukocytic cell types. When the immune system is considered as a functional entity, therefore, these non-leukocytic components become as important as their more renowned leukocytic relatives. References 1. R. VIRCHOW, Cellular Pathology, John Churchill, London (1860). 2. A.T. MCMANUS, A.D. MASON, JR., W.F. MCMANUS and B.A. PRUITT, JR., Arch. Surg. 129 1306Ð9 (1994). 3. N. COOK, Burns 24 91Ð8 (1998). 4. B.A. PRUITT, JR., A.T. MCMANUS, S.H. KIM and C.W. GOODWIN, World J. Surg. 22 135Ð45 (1998). 5. J. LI„ARES, A. SITGES-SERRA, J. GARAU, J.L. PƒREZ and R. MARTêN, J. Clin. Microbiol. 21 357Ð60 (1985). 6. S. HURAIB, A. ASKAR, H. ABU-AISHA and J. AL-WAKEEL, Angiology 45 1047Ð51 (1994). B. Spellberg / Life Sciences 67 (2000) 477Ð502 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. 46. 47. 48. 495 M.J. MARPLES, Br. J. Dermatol. 81 Suppl 1:2Ð13 (1969). S.M. PUHVEL, R.M. REISNER and D.A. AMIRIAN, J. Invest. Dermatol. 65 525Ð31 (1975). R. ALY, H.I. MAIBACH and E. BLOOM, Acta Derm. Venereol. 58 501Ð4 (1978). P.A. MACKOWIAK, New Engl. J. Med. 307 83Ð93 (1982). W.C. DUNCAN, M.E. MCBRIDE and J.M. KNOX, J. Invest. Dermatol. 52 479Ð84 (1969). R.J. HOLT, J. Med. Microbiol. 4 375Ð9 (1971). N. OCHI, M. AZEGAMI and S.I. ISHII, J. Biochem. 83 727Ð36 (1978). P. REEVES, Bacterol. Rev. 29 24 (1965). S. SELWYN, Br. J. Dermatol. 93 487Ð493 (1975). A. TRIPPESTAD and T. MIDTVEDT, Clin. Exp. Immunol. 8 639Ð646 (1971). Y. ITO, Y. NISIYAMA, K. SHIMOKATA, Y. KIMURA, I. NAGATA and A. KUNII, Infect. Immun. 13 332Ð336 (1976). J.W. SCHEFFEL and Y.B. KIM, Infect. Immun. 26 202Ð210 (1979). L.G. SCHEIMANN, G. KNOX, D. SHER and R. S., J. Invest. Dermatol. 34 171Ð177 (1960). W.C. NOBLE, Br. J. Dermatol. 80 279Ð81 (1968). R.K. FREINKEL and Y. SHEN, J. Invest. Dermatol. 53 422Ð7 (1969). R.R. MARPLES, A.M. KLIGMAN, L.R. LANTIS and D.T. DOWNING, J. Invest. Dermatol. 55 173Ð8 (1970). K. SPRUNT, G.A. LEIDY and W. REDMAN, J. Infect. Dis. 123 1Ð10 (1971). R.A. AMONETTE and E.W. ROSENBERG, Arch. Dermatol. 107 71Ð3 (1973). R.C. STEWART, J. Am. Podiatry Assoc. 63 639Ð646 (1973). H.G. BURKITT, B. YOUNG and J.W. HEATH, WheaterÕs Functional Histology (3rd Ed), Churchill Livingstone, London (1993). M. AMAGAI, J. Invest. Dermatol. 104 146Ð52 (1995). K.L. CEPEK, S.K. SHAW, C.M. PARKER, G.J. RUSSELL, J.S. MORROW, D.L. RIMM and M.B. BRENNER, Nature 372 190Ð3 (1994). P.I. KARECLA, S.J. BOWDEN, S.J. GREEN and P.J. KILSHAW, Eur. J. Immunol. 25 852Ð6 (1995). S. AIBA, S. NAKAGAWA, H. OZAWA and H. TAGAMI, J. Invest. Dermatol. 105 379Ð82 (1995). K. SCHWARZENBERGER and M.C. UDEY, J. Invest. Dermatol. 106 553Ð8 (1996). A. TANG, M. AMAGAI, L.G. GRANGER, J.R. STANLEY and M.C. UDEY, Nature 361 82Ð5 (1993). E. FUCHS, Annu. Rev. Cell Dev. Biol. 11 123Ð53 (1995). D.M. PILLSBURY and G. REBELL, J. Invest. Dermatol. 18 173Ð180 (1952). D.A. EMMATTY and R.J. GREEN, JR., Can. J. Microbiol. 13 635Ð42 (1967). L.F. MONTES and W.H. WILBORN, Br. J. Dermatol. 81 Suppl 1 S23ÐS32 (1969). L.F. MONTES and W.H. WILBORN, Arch. Dermatol. 101 145Ð59 (1970). C. HEUFLER, G. TOPAR, A. GRASSEGER, U. STANZL, F. KOCH, N. ROMANI, A.E. NAMEN and G. SCHULER, J. Exp. Med. 178 1109Ð14 (1993). J.A. CHODAKEWITZ, T.S. KUPPER and D.L. COLEMAN, J. Immunol. 140 832Ð6 (1988). J.A. CHODAKEWITZ, J. LACY, S.E. EDWARDS, N. BIRCHALL and D.L. COLEMAN, J. Immunol. 144 2190Ð6 (1990). C. HEUFLER, F. KOCH and G. SCHULER, J. Exp. Med. 167 700Ð5 (1988). S. XU, K. ARIIZUMI, D. EDELBAUM, P.R. BERGSTRESSER and A. TAKASHIMA, Eur. J. Immunol. 25 1018Ð24 (1995). C.A. FOSTER, H. YOKOZEKI, K. RAPPERSBERGER, F. KONING, B. VOLC-PLATZER, A. RIEGER, J.E. COLIGAN, K. WOLFF and G. STINGL, J. Exp. Med. 171 997Ð1013 (1990). A. ELBE, C.A. FOSTER and G. STINGL, Semin. Immunol. 8 341Ð9 (1996). J.D. BOS, I. ZONNEVELD, P.K. DAS, S.R. KRIEG, C.M. VAN DER LOOS and M.L. KAPSENBERG, J. Invest. Dermatol. 88 569Ð73 (1987). A.L. DAVIS, J.L. MCKENZIE and D.N. HART, Immunology 65 573Ð81 (1988). J.D. BOS, O.J. DE BOER, E. TIBOSCH, P.K. DAS and S.T. PALS, Arch. Dermatol. Res. 285 179Ð83 (1993). E.L. BERG, J. MAGNANI, R.A. WARNOCK, M.K. ROBINSON and E.C. BUTCHER, Biochem. Biophys. Res. Commun. 184 1048Ð55 (1992). 496 B. Spellberg / Life Sciences 67 (2000) 477Ð502 49. C.R. MACKAY, W.L. MARSTON and L. DUDLER, J. Exp. Med. 171 801Ð17 (1990). 50. R.P. BUCY, C.L. CHEN and M.D. COOPER, J. Immunol. 142 3045Ð9 (1989). 51. P. DUPUY, M. HESLAN, S. FRAITAG, T. HERCEND, L. DUBERTRET and M. BAGOT, J. Invest. Dermatol. 94 764Ð8 (1990). 52. J.D. BOS, M.B. TEUNISSEN, I. CAIRO, S.R. KRIEG, M.L. KAPSENBERG, P.K. DAS and J. BORST, J. Invest. Dermatol. 94 37Ð42 (1990). 53. G. STEINER, F. KONING, A. ELBE, E. TSCHACHLER, W.M. YOKOYAMA, E.M. SHEVACH, G. STINGL and J.E. COLIGAN, Eur. J. Immunol. 18 1323Ð8 (1988). 54. M. ALAIBAC, J. MORRIS, R. YU and A.C. CHU, Br. J. Dermatol. 127 458Ð62 (1992). 55. K. UYEMURA, R.J. DEANS, H. BAND, J. OHMEN, G. PANCHAMOORTHY, C.T. MORITA, T.H. REA and R.L. MODLIN, J. Exp. Med. 174 683Ð92 (1991). 56. D.A. DUNN, A.S. GADENNE, S. SIMHA, E.A. LERNER, M. BIGBY and P.A. BLEICHER, Proc. Natl. Acad. Sci. USA 90 1267Ð71 (1993). 57. M. ALAIBAC and A.C. CHU, Mol. Immunol. 33 1035Ð8 (1996). 58. M. FURUE, M. NINDL, K. KAWABE, K. NAKAMURA, Y. ISHIBASHI and K. SAGAWA, J. Am. Acad. Dermatol. 27 419Ð26 (1992). 59. S.A. PORCELLI, Adv. Immunol. 59 1Ð98 (1995). 60. E.M. BECKMAN, S.A. PORCELLI, C.T. MORITA, S.M. BEHAR, S.T. FURLONG and M.B. BRENNER, Nature 372 691Ð4 (1994). 61. S.A. PORCELLI and R.L. MODLIN, J. Immunol. 155 3709Ð10 (1995). 62. P.A. SIELING, D. CHATTERJEE, S.A. PORCELLI, T.I. PRIGOZY, R.J. MAZZACCARO, T. SORIANO, B.R. BLOOM, M.B. BRENNER, M. KRONENBERG, P.J. BRENNAN and et al., Science 269 227Ð30 (1995). 63. S.I. KATZ, K. TAMAKI and D.H. SACHS, Nature 282 324Ð6 (1979). 64. K. WOLFF, J. Dermatol. 10 401Ð10 (1983). 65. K. WOLFF and G. STINGL, J. Invest. Dermatol. 80 Suppl 17sÐ21s (1983). 66. A.D. HOGAN and A.W. BURKS, Ann. Allergy Asthma Immunol. 75 5Ð10 (1995). 67. M. HILLIGES, L. WANG and O. JOHANSSON, J. Invest. Dermatol. 104 134Ð7 (1995). 68. J. HOSOI, G.F. MURPHY, C.L. EGAN, E.A. LERNER, S. GRABBE, A. ASAHINA and R.D. GRANSTEIN, Nature 363 159Ð63 (1993). 69. T. LOTTI, G. HAUTMANN and E. PANCONESI, J. Am. Acad. Dermatol. 33 482Ð96 (1995). 70. C.A. DUNNICK, N.S. GIBRAN and D.M. HEIMBACH, J. Burn Care Rehabil. 17 390Ð6 (1996). 71. I. BOXMAN, C. L…WIK, L. AARDEN and M. PONEC, J. Invest. Dermatol. 101 316Ð24 (1993). 72. G. PANG, L. COUCH, R. BATEY, R. CLANCY and A. CRIPPS, Clin. Exp. Immunol. 96 437Ð43 (1994). 73. S. WERNER, K.G. PETERS, M.T. LONGAKER, F. FULLER-PACE, M.J. BANDA and L.T. WILLIAMS, Proc. Natl. Acad. Sci. USA 89 6896Ð900 (1992). 74. L. MEUNIER, A. GONZALEZ-RAMOS and K.D. COOPER, J. Immunol. 151 4067Ð80 (1993). 75. A. LENZ, M. HEINE, G. SCHULER AND N. ROMANI, J. CLIN. INVEST. 92 2587Ð96 (1993). 76. F.O. NESTLE, X.G. ZHENG, C.B. THOMPSON, L.A. TURKA and B.J. NICKOLOFF, J. Immunol. 151 6535Ð6545 (1993). 77. M. ALLG…WER, G.A. SCHOENENBERGER and B.G. SPARKES, Burns 21 Suppl 1 S7Ð47 (1995). 78. R.M. WILLIAMS, H.R. BERTHOUD and R.H. STEAD, Neuroimmunomodulation 4 266Ð70 (1997). 79. T.S. KUPPER, D.W. BALLARD, A.O. CHUA, J.S. MCGUIRE, P.M. FLOOD, M.C. HOROWITZ, R. LANGDON, L. LIGHTFOOT and U. GUBLER, J. Exp. Med. 164 2095Ð100 (1986). 80. T.S. KUPPER, J. Clin. Invest. 86 1783Ð9 (1990). 81. L.C. WOOD, S.M. JACKSON, P.M. ELIAS, C. GRUNFELD and K.R. FEINGOLD, J. Clin. Invest. 90 482Ð7 (1992). 82. B.J. NICKOLOFF and Y. NAIDU, J. Am. Acad. Dermatol. 30 535Ð46 (1994). 83. T.S. KUPPER, J. Invest. Dermatol. 94 146SÐ150S (1990). 84. J. ANSEL, P. PERRY, J. BROWN, D. DAMM, T. PHAN, C. HART, T. LUGER and S. HEFENEIDER, J. Invest. Dermatol. 94 101SÐ107S (1990). 85. A. K…CK, T. SCHWARZ, R. KIRNBAUER, A. URBANSKI, P. PERRY, J.C. ANSEL and T.A. LUGER, J. Exp. Med. 172 1609Ð14 (1990). B. Spellberg / Life Sciences 67 (2000) 477Ð502 86. 87. 88. 89. 90. 91. 92. 93. 94. 95. 96. 97. 98. 99. 100. 101. 102. 103. 104. 105. 106. 107. 108. 109. 110. 111. 112. 113. 114. 115. 116. 117. 118. 497 M.M. HOBBS, T.R. PAUL, P.B. WYRICK and T.H. KAWULA, Infect. Immun. 66 2914Ð21 (1998). J.L. WILMER and M.I. LUSTER, Cell Biol. Toxicol. 11 37Ð50 (1995). M. BICKEL, S.M. N…THEN, K. FREIBURGHAUS and D. SHIRE, J. Dental Res. 75 1827Ð34 (1996). M.D. WITMER-PACK, W. OLIVIER, J. VALINSKY, G. SCHULER and R.M. STEINMAN, J. Exp. Med. 166 1484Ð98 (1987). D. EDELBAUM, M. MOHAMADZADEH, P.R. BERGSTRESSER, K. SUGAMURA and A. TAKASHIMA, J. Invest. Dermatol. 105 837Ð43 (1995). S. KONDO, F. KOOSHESH and D.N. SAUDER, J. Cell. Physiol. 171 190Ð5 (1997). R.W. GROVES, M.H. ALLEN, E.L. ROSS, J.N. BARKER and D.M. MACDONALD, Br. J. Dermatol. 132 345Ð52 (1995). S. GRABBE and T. SCHWARZ, Am. J. Contact Dermat. 7 238Ð46 (1996). J. MIDDLETON, S. NEIL, J. WINTLE, I. CLARK-LEWIS, H. MOORE, C. LAM, M. AUER, E. HUB and A. ROT, Cell 91 385Ð95 (1997). G. DE LIBERO, Immunol. Today 18 22Ð6 (1997). G. STINGLE, E. TSCHACHLER, V. GROH and K. WOLFF, The immune functions of epidermal cells, D.A. NORRIS (Ed), 3Ð72, Marcel Degger, New York (1989). P. CONSTANT, F. DAVODEAU, M.A. PEYRAT, Y. POQUET, G. PUZO, M. BONNEVILLE and J.J. FOURNIƒ, Science 264 267Ð70 (1994). Y. TANAKA, S. SANO, E. NIEVES, G. DE LIBERO, D. ROSA, R.L. MODLIN, M.B. BRENNER, B.R. BLOOM and C.T. MORITA, Proc. Natl. Acad. Sci. USA 91 8175Ð9 (1994). B. SCHOEL, S. SPRENGER and S.H. KAUFMANN, Eur. J. Immunol. 24 1886Ð92 (1994). M.B. TEUNISSEN, C.W. KOOMEN, R. DE WAAL MALEFYT, E.A. WIERENGA and J.D. BOS, J. Invest. Dermatol. 111 645Ð9 (1998). H.L. COLLINS and G.J. BANCROFT, Eur. J. Immunol. 22 1447Ð54 (1992). D.V. JOVANOVIC, J.A. DI BATTISTA, J. MARTEL-PELLETIER, F.C. JOLICOEUR, Y. HE, M. ZHANG, F. MINEAU and J.P. PELLETIER, J. Immunol. 160 3513Ð21 (1998). G.A. FOLLOWS, M.E. MUNK, A.J. GATRILL, P. CONRADT and S.H. KAUFMANN, Infect. Immun. 60 1229Ð31 (1992). A.H. ENK and S.I. KATZ, Proc. Natl. Acad. Sci. USA 89 1398Ð402 (1992). G. SCHUHMACHERS, K. ARIIZUMI, P.A. MATHEW, M. BENNETT, V. KUMAR and A. TAKASHIMA, Eur. J. Immunol. 25 1117Ð20 (1995). I.E. FLESCH, J.H. HESS, S. HUANG, M. AGUET, J. ROTHE, H. BLUETHMANN and S.H. KAUFMANN, J. Exp. Med. 181 1615Ð21 (1995). D.H. LIBRATY, L.E. AIRAN, K. UYEMURA, D. JULLIEN, B. SPELLBERG, T.H. REA and R.L. MODLIN, J. Clin. Invest. 99 336Ð41 (1997). C.A. WENNER, M.L. G†LER, S.E. MACATONIA, A. OÕGARRA and K.M. MURPHY, J. Immunol. 156 1442Ð7 (1996). A. DÕANDREA, M. RENGARAJU, N.M. VALIANTE, J. CHEHIMI, M. KUBIN, M. ASTE, S.H. CHAN, M. KOBAYASHI, D. YOUNG, E. NICKBARG and et al., J. Exp. Med. 176 1387Ð98 (1992). M.J. SKEEN, M.A. MILLER, T.M. SHINNICK and H.K. ZIEGLER, J. Immunol. 156 1196Ð206 (1996). F. SONG, G. MATSUZAKI, M. MITSUYAMA and K. NOMOTO, J. Immunol. 156 2979Ð84 (1996). A.M. KRIEG, L. LOVE-HOMAN, A.K. YI and J.T. HARTY, J. Immunol. 161 2428Ð34 (1998). T. GERMANN, M.K. GATELY, D.S. SCHOENHAUT, M. LOHOFF, F. MATTNER, S. FISCHER, S.C. JIN, E. SCHMITT and E. R†DE, Eur. J. Immunol. 23 1762Ð70 (1993). R. MANETTI, P. PARRONCHI, M.G. GIUDIZI, M.P. PICCINNI, E. MAGGI, G. TRINCHIERI and S. ROMAGNANI, J. Exp. Med. 177 1199Ð204 (1993). C.Y. WU, C. DEMEURE, M. KINIWA, M. GATELY and G. DELESPESSE, J. Immunol. 151 1938Ð49 (1993). G.E. JONES, W.E. ALLEN and A.J. RIDLEY, Cell Adhes. Commun. 6 237Ð45 (1998). E. BLASI, S. FARINELLI, L. VARESIO and F. BISTONI, Infect. Immun. 58 1073Ð7 (1990). J.M. SHIPLEY, R.L. WESSELSCHMIDT, D.K. KOBAYASHI, T.J. LEY and S.D. SHAPIRO, Proc. Natl. Acad. Sci. USA 93 3942Ð6 (1996). 498 B. Spellberg / Life Sciences 67 (2000) 477Ð502 119. Y. ZHANG, K. MCCLUSKEY, K. FUJII and L.M. WAHL, J. Immunol. 161 3071Ð6 (1998). 120. T.J. GRONSKI, JR., R.L. MARTIN, D.K. KOBAYASHI, B.C. WALSH, M.C. HOLMAN, M. HUBER, H.E. VAN WART and S.D. SHAPIRO, J. Biol. Chem. 272 12189Ð94 (1997). 121. M. VAALAMO, A.L. KARINIEMI, S.D. SHAPIRO and U. SAARIALHO-KERE, J. Invest. Dermatol. 112 499Ð505 (1999). 122. M. CUMBERBATCH, R.J. DEARMAN and I. KIMBER, Immunology 92 388Ð95 (1997). 123. A.A. PRICE, M. CUMBERBATCH, I. KIMBER and A. AGER, J. Exp. Med. 186 1725Ð35 (1997). 124. S. AIBA, S. NAKAGAWA, H. OZAWA, K. MIYAKE, H. YAGITA and H. TAGAMI, J. Invest. Dermatol. 100 143Ð7 (1993). 125. C.P. LARSEN, S.C. RITCHIE, R. HENDRIX, P.S. LINSLEY, K.S. HATHCOCK, R.J. HODES, R.P. LOWRY and T.C. PEARSON, J. Immunol. 152 5208Ð19 (1994). 126. T. KAWAMURA and M. FURUE, Eur. J. Immunol. 25 1913Ð7 (1995). 127. G.F. MURPHY, E. FONFERKO, T. FLOTTE and A.K. BHAN, Arch. Dermatol. Res. 278 126Ð32 (1985). 128. L.J. PICKER, T.K. KISHIMOTO, C.W. SMITH, R.A. WARNOCK and E.C. BUTCHER, Nature 349 796Ð9 (1991). 129. R.P. MCEVER, J.H. BECKSTEAD, K.L. MOORE, L. MARSHALL-CARLSON and D.F. BAINTON, J. Clin. Invest. 84 92Ð9 (1989). 130. R. BONFANTI, B.C. FURIE, B. FURIE and D.D. WAGNER, Blood 73 1109Ð12 (1989). 131. R.G. SCHAUB, C.J. DUNN, M.R. DEIBEL, A.E. BERGER, D. WUNDERLICH and W.E. FLEMING, Agents Actions 31 127Ð34 (1990). 132. H.A. BULL, M.H. RUSTIN, J. SPAULL, J. COHEN, E. WILSON-JONES and P.M. DOWD, Br. J. Dermatol. 122 153Ð64 (1990). 133. I.G. COLDITZ, Immunol. Cell Biol. 68 397Ð403 (1990). 134. S.A. VINORES, H. SEN and P.A. CAMPOCHIARO, Invest. Ophthalmol. Vis. Sci. 33 1870Ð8 (1992). 135. G. JANCSî, F. OBçL, JR., I. TîTH-KçSA, M. KATONA and S. HUSZ, Int. J. Tissue React. 7 449Ð57 (1985). 136. C.H. HOYLE, R.W. STONES, T. ROBSON, K. WHITLEY and G. BURNSTOCK, J. Anat. 188 633Ð44 (1996). 137. S. BEVAN, Prog. Brain Res. 113 201Ð13 (1996). 138. L.S. SORKIN, W.H. XIAO, R. WAGNER and R.R. MYERS, Neuroscience 81 255Ð62 (1997). 139. D.M. MCDONALD, J.J. BOWDEN, P. BALUK and N.W. BUNNETT, Adv. Exp. Med. Biol. 410 453Ð62 (1996). 140. M. DUX, G. JANCSî, H. SANN and F.K. PIERAU, Inßamm. Res. 45 10Ð3 (1996). 141. M.J. DOMEK, E.I. BLACKMAN, J. KAO, X.Y. ZHANG, F. IWATA, K. SENO and F.W. LEUNG, J. Gastroenterol. Hepatol. 12 698Ð702 (1997). 142. J. GOLDHILL, P. PICHAT, N. ROOME, I. ANGEL and S. ARBILLA, Arzneimittalforschung 48 179Ð84 (1998). 143. J.M. EBERTZ, C.A. HIRSHMAN, N.S. KETTELKAMP, H. UNO and J.M. HANIFIN, J. Invest. Dermatol. 88 682Ð5 (1987). 144. H. YANO, B.K. WERSHIL, N. ARIZONO and S.J. GALLI, J. Clin. Invest. 84 1276Ð86 (1989). 145. M.D. THARP, J. Invest. Dermatol. 93 107SÐ112S (1989). 146. B.F. RAMOS, Y. ZHANG and B.A. JAKSCHIK, J. Immunol. 152 1380Ð4 (1994). 147. A.P. PRODEUS, X. ZHOU, M. MAURER, S.J. GALLI and M.C. CARROLL, Nature 390 172Ð5 (1997). 148. H. MATSUDA, K. KAWAKITA, Y. KISO, T. NAKANO and Y. KITAMURA, J. Immunol. 142 927Ð31 (1989). 149. J.R. GORDON and S.J. GALLI, Nature 346 274Ð6 (1990). 150. M. PLAUT, J.H. PIERCE, C.J. WATSON, J. HANLEY-HYDE, R.P. NORDAN and W.E. PAUL, Nature 339 64Ð7 (1989). 151. L.J. PICKER, R.A. WARNOCK, A.R. BURNS, C.M. DOERSCHUK, E.L. BERG and E.C. BUTCHER, Cell 66 921Ð33 (1991). 152. D. ROHDE, W. SCHL†TER-WIGGER, V. MIELKE, P. VON DEN DRIESCH, B. VON GAUDECKER and W. STERRY, J. Invest. Dermatol. 98 794Ð9 (1992). B. Spellberg / Life Sciences 67 (2000) 477Ð502 499 153. K.L. MOORE and L.F. THOMPSON, Biochem. Biophys. Res. Commun. 186 173Ð81 (1992). 154. K.L. MOORE, N.L. STULTS, S. DIAZ, D.F. SMITH, R.D. CUMMINGS, A. VARKI and R.P. MCEVER, J. Cell Biol. 118 445Ð56 (1992). 155. I. STRICKLAND, L.E. RHODES, B.F. FLANAGAN and P.S. FRIEDMANN, J. Invest. Dermatol. 108 763Ð8 (1997). 156. C. DELCLAUX, C. DELACOURT, M.P. DÕORTHO, V. BOYER, C. LAFUMA and A. HARF, Am. J. Respir. Cell Mol. Biol. 14 288Ð95 (1996). 157. M.U. EHRENGRUBER, D.A. DERANLEAU and T.D. COATES, J. Exp. Biol. 199 741Ð7 (1996). 158. S. ROSENGREN, P.M. HENSON and G.S. WORTHEN, Am. J. Physiol. 267 1623Ð32 (1994). 159. G.S. WORTHEN, P.M. HENSON, S. ROSENGREN, G.P. DOWNEY and D.M. HYDE, Am. J. Respir. Cell Mol. Biol. 10 1Ð7 (1994). 160. M. CHRISTOFIDOU-SOLOMIDOU, M.T. NAKADA, J. WILLIAMS, W.A. MULLER and H.M. DELISSER, J. Immunol. 158 4872Ð8 (1997). 161. J.X. GAO and A.C. ISSEKUTZ, Immunology 88 463Ð70 (1996). 162. J.X. GAO and A.C. ISSEKUTZ, Immunology 90 448Ð54 (1997). 163. J.T. MANDEVILLE, M.A. LAWSON and F.R. MAXFIELD, J. Leukoc. Biol. 61 188Ð200 (1997). 164. R.M. KUNTZ and W.M. SALTZMAN, Biophys. J. 72 1472Ð80 (1997). 165. H.D. GRESHAM, I.L. GRAHAM, G.L. GRIFFIN, J.C. HSIEH, L.J. DONG, A.E. CHUNG and R.M. SENIOR, J. Biol. Chem. 271 30587Ð94 (1996). 166. D. CHOQUET, D.P. FELSENFELD and M.P. SHEETZ, Cell 88 39Ð48 (1997). 167. R. STEADMAN, P.L. ST JOHN, R.A. EVANS, G.J. THOMAS, M. DAVIES, L.W. HECK and D.R. ABRAHAMSON, Int. J. Biochem. Cell Biol. 29 993Ð1004 (1997). 168. A. FERRANTE, M. NANDOSKAR, A. WALZ, D.H. GOH and I.C. KOWANKO, Int. Arch. Allergy Appl. Immunol. 86 82Ð91 (1988). 169. D.H. LIVINGSTON, S.H. APPEL, G. SONNENFELD and M.A. MALANGONI, J. Surg. Res. 46 322Ð6 (1989). 170. H.D. GRESHAM, L.T. CLEMENT, J.E. LEHMEYER, F.M. GRIFFIN, JR. and J.E. VOLANAKIS, J. Immunol. 137 868Ð75 (1986). 171. A.M. BUCKLE and N. HOGG, J. Immunol. 143 2295Ð301 (1989). 172. R. STEADMAN, M.M. PETERSEN, N. TOPLEY, D. WILLIAMS, N. MATTHEWS, B. SPUR and J.D. WILLIAMS, J. Immunol. 144 2712Ð8 (1990). 173. P.A. DETMERS, D.E. POWELL, A. WALZ, I. CLARK-LEWIS, M. BAGGIOLINI and Z.A. COHN, J. Immunol. 147 4211Ð7 (1991). 174. T. TERUI, T. KATO and H. TAGAMI, J. Invest. Dermatol. 92 593Ð7 (1989). 175. N. DOVEZENSKI, R. BILLETTA and I. GIGLI, J. Clin. Invest. 90 2000Ð12 (1992). 176. A. VECCHIARELLI, C. RETINI, A. CASADEVALL, C. MONARI, D. PIETRELLA and T.R. KOZEL, Infect. Immun. 66 4324Ð30 (1998). 177. M. HACHICHA, P. RATHANASWAMI, P.H. NACCACHE and S.R. MCCOLL, J. Immunol. 160 449Ð54 (1998). 178. P.C. LEIJH, M.T. VAN DEN BARSELAAR, T.L. VAN ZWET, I. DUBBELDEMAN-REMPT and R. VAN FURTH, Immunology 37 453Ð65 (1979). 179. C.D. ENK, D. SREDNI, A. BLAUVELT and S.I. KATZ, J. Immunol. 154 4851Ð6 (1995). 180. T.A. FERGUSON, P. DUBE and T.S. GRIFFITH, J. Exp. Med. 179 1597Ð604 (1994). 181. T.S. KUPPER and R.W. GROVES, J. Invest. Dermatol. 105 62SÐ66S (1995). 182. R.W. GROVES, J. GIRI, J. SIMS, S.K. DOWER and T.S. KUPPER, J. Immunol. 154 4065Ð72 (1995). 183. E. SCHAUER, F. TRAUTINGER, A. K…CK, A. SCHWARZ, R. BHARDWAJ, M. SIMON, J.C. ANSEL, T. SCHWARZ and T.A. LUGER, J. Clin. Invest. 93 2258Ð62 (1994). 184. A. CHAKRABORTY, A. SLOMINSKI, G. ERMAK, J. HWANG and J. PAWELEK, J. Invest. Dermatol. 105 655Ð9 (1995). 185. A. SLOMINSKI, G. ERMAK, J. HWANG, A. CHAKRABORTY, J.E. MAZURKIEWICZ and M. MIHM, FEBS Lett. 374 113Ð6 (1995). 186. M. WINTZEN, M. YAAR, J.P. BURBACH and B.A. GILCHREST, J. Invest. Dermatol. 106 673Ð8 (1996). 500 B. Spellberg / Life Sciences 67 (2000) 477Ð502 187. J.G. CANNON, J.B. TATRO, S. REICHLIN and C.A. DINARELLO, J. Immunol. 137 2232Ð6 (1986). 188. K.G. MUGRIDGE, M. PERRETTI, P. GHIARA and L. PARENTE, Eur. J. Pharm. 197 151Ð5 (1991). 189. R.S. BHARDWAJ, A. SCHWARZ, E. BECHER, K. MAHNKE, Y. ARAGANE, T. SCHWARZ and T.A. LUGER, J. Immunol. 156 2517Ð21 (1996). 190. A.W. TAYLOR, J.W. STREILEIN and S.W. COUSINS, Neuroimmunomodulation 1 188Ð94 (1994). 191. T.A. LUGER, T. SCHOLZEN, T. BRZOSKA, E. BECHER, A. SLOMINSKI and R. PAUS, Ann. N. Y. Acad. Sci. 840 381Ð94 (1998). 192. R. BHARDWAJ, E. BECHER, K. MAHNKE, M. HARTMEYER, T. SCHWARZ, T. SCHOLZEN and T.A. LUGER, J. Immunol. 158 3378Ð84 (1997). 193. A. CATANIA, N. RAJORA, F. CAPSONI, F. MINONZIO, R.A. STAR and J.M. LIPTON, Peptides 17 675Ð9 (1996). 194. T. KRAKAUER, J. VILCEK and J.J. OPPENHEIM, Transforming Growth Factor Beta (4th Ed), W.E. Paul (Ed), 800Ð803, Lippincott-Raven, New York (1999). 195. V.T. NGUYEN, W.S. WALKER and E.N. BENVENISTE, Eur. J. Immunol. 28 2537Ð48 (1998). 196. J.J. BRIGHT and S. SRIRAM, J. Immunol. 161 1772Ð7 (1998). 197. J. TANG, B.L. NUCCIE, I. RITTERMAN, J.L. LIESVELD, C.N. ABBOUD and D.H. RYAN, J. Immunol. 159 117Ð25 (1997). 198. S.M. WAHL, G.L. COSTA, M. CORCORAN, L.M. WAHL and A.E. BERGER, J. Immunol. 150 3553Ð60 (1993). 199. K.M. GILBERT, M. THOMAN, K. BAUCHE, T. PHAM and W.O. WEIGLE, Immunol. Invest. 26 459Ð72 (1997). 200. R. JOSIEN, P. DOUILLARD, C. GUILLOT, M. M†SCHEN, I. ANEGON, J. CHETRITT, S. MENORET, C. VIGNES, J.P. SOULILLOU and M.C. CUTURI, J. Clin. Invest. 102 1920Ð6 (1998). 201. C. PARDOUX, X. MA, S. GOBERT, S. PELLEGRINI, P. MAYEUX, F. GAY, G. TRINCHIERI and S. CHOUAIB, Blood 93 1448Ð55 (1999). 202. K. PAVELIC, N. DESPOT, S. LEVANAT and T. CASL, Biol. Chem. 371 687Ð92 (1990). 203. J.K. WRIGHT, T.E. CAWSTON and B.L. HAZLEMAN, Biochim. Biophys. Acta 1094 207Ð10 (1991). 204. J. SLAVIN, E. UNEMORI, T.K. HUNT and E. AMENTO, Growth Factors 11 205Ð13 (1994). 205. W.B. SMITH, L. NOACK, Y. KHEW-GOODALL, S. ISENMANN, M.A. VADAS and J.R. GAMBLE, J. Immunol. 157 360Ð8 (1996). 206. F.E. FOX, H.C. FORD, R. DOUGLAS, S. CHERIAN and P.C. NOWELL, Cell. Immunol. 150 45Ð58 (1993). 207. I. MUROHASHI, K. ENDHO, S. NISHIDA, S. YOSHIDA, I. JINNAI, M. BESSHO and K. HIRASHIMA, Exp. Hematol. 23 970Ð7 (1995). 208. F. LARDON, H.W. SNOECK, L. HAENEN, M. LENJOU, G. NIJS, S.F. WEEKX, P.C. VAN RANST, Z.N. BERNEMAN and D.R. VAN BOCKSTAELE, Leukemia 10 1937Ð43 (1996). 209. D.A. BETTINGER, D.R. YAGER, R.F. DIEGELMANN and I.K. COHEN, Plastic Reconstruct. Surg. 98 827Ð33 (1996). 210. J.K. JUTLEY, E.J. WOOD and W.J. CUNLIFFE, Matrix 13 235Ð41 (1993). 211. E.J. BATTEGAY, E.W. RAINES, R.A. SEIFERT, D.F. BOWEN-POPE and R. ROSS, Cell 63 515Ð24 (1990). 212. R.A. SEDER and W.E. PAUL, Annu. Rev. Immunol. 12 635Ð73 (1994). 213. R.A. SEDER, T. MARTH, M.C. SIEVE, W. STROBER, J.J. LETTERIO, A.B. ROBERTS and B. KELSALL, J. Immunol. 160 5719Ð28 (1998). 214. L.M. TERAN, M. MOCHIZUKI, J. BARTELS, E.L. VALENCIA, T. NAKAJIMA, K. HIRAI and J.M. SCHR…DER, Am. J. Respir. Cell Mol. Biol. 20 777Ð86 (1999). 215. R.L. SCHMOUDER, R.M. STRIETER and S.L. KUNKEL, Kidney Int. 44 43Ð9 (1993). 216. C.A. MARTIN and M.E. DORF, Cell. Immunol. 135 245Ð58 (1991). 217. P. RATHANASWAMI, M. HACHICHA, M. SADICK, T.J. SCHALL and S.R. MCCOLL, J. Biol. Chem. 268 5834Ð9 (1993). 218. M. GOEBELER, T. YOSHIMURA, A. TOKSOY, U. RITTER, E.B. BR…CKER and R. GILLITZER, J. Invest. Dermatol. 108 445Ð51 (1997). B. Spellberg / Life Sciences 67 (2000) 477Ð502 501 219. S.R. MCCOLL, M. HACHICHA, S. LEVASSEUR, K. NEOTE and T.J. SCHALL, J. Immunol. 150 4550Ð 60 (1993). 220. P.A. TESSIER, P.H. NACCACHE, I. CLARK-LEWIS, R.P. GLADUE, K.S. NEOTE and S.R. MCCOLL, J. Immunol. 159 3595Ð602 (1997). 221. F. AUSTRUP, D. VESTWEBER, E. BORGES, M. L…HNING, R. BR€UER, U. HERZ, H. RENZ, R. HALLMANN, A. SCHEFFOLD, A. RADBRUCH and A. HAMANN, Nature 385 81Ð3 (1997). 222. J.T. SIVEKE and A. HAMANN, J. Immunol. 160 550Ð4 (1998). 223. A. OÕGARRA, L.M. MCEVOY and A. ZLOTNIK, Curr. Biol. 8 R646Ð9 (1998). 224. S. SCHRUM, P. PROBST, B. FLEISCHER and P.F. ZIPFEL, J. Immunol. 157 3598Ð604 (1996). 225. A. SILBER, W. NEWMAN, K.A. REIMANN, E. HENDRICKS, D. WALSH and D.J. RINGLER, Lab. Invest. 70 163Ð75 (1994). 226. Inßammation and Repair (5th Ed), R.S. COTRAN, V. KUMAR, S.L. ROBBINS and F.J. SCHOEN (Eds), 51Ð59, W.B. Saunders (1994). 227. T.D. GEPPERT and P.E. LIPSKY, J. Immunol. 135 3750Ð62 (1985). 228. J.F. WEDGWOOD, L. HATAM and V.R. BONAGURA, Cell. Immunol. 111 1Ð9 (1988). 229. D.R. JOHNSON and J.S. POBER, Proc. Natl. Acad. Sci. USA 87 5183Ð7 (1990). 230. M. VORA, H. YSSEL, J.E. DE VRIES and M.A. KARASEK, J. Immunol. 152 5734Ð41 (1994). 231. C.O. SAVAGE, C.J. BROOKS, G.C. HARCOURT, J.K. PICARD, W. KING, D.M. SANSOM and N. WILLCOX, Int. Immunol. 7 471Ð9 (1995). 232. M.C. CID, J. ESPARZA, M. JUAN, A. MIRALLES, J. ORDI, R. VILELLA, A. URBANO-MçRQUEZ, A. GAYË, J. VIVES and J. YAG†E, Eur. J. Immunol. 24 1377Ð82 (1994). 233. W. MA and J.S. POBER, J. Immunol. 161 2158Ð67 (1998). 234. B.J. NICKOLOFF and L.A. TURKA, Immunol. Today 15 464Ð9 (1994). 235. F.M. MARELLI-BERG, A. WEETMAN, L. FRASCA, S.J. DEACOCK, N. IMAMI, G. LOMBARDI and R.I. LECHLER, J. Immunol. 159 5853Ð61 (1997). 236. R. TAKEMURA, P.E. STENBERG, D.F. BAINTON and Z. WERB, J. Cell Biol. 102 55Ð69 (1986). 237. B.A. KRUSKAL and F.R. MAXFIELD, J. Cell Biol. 105 2685Ð93 (1987). 238. D.F. BAINTON, R. TAKEMURA, P.E. STENBERG and Z. WERB, Am. J. Pathol. 134 15Ð26 (1989). 239. L. ROMANI, A. MENCACCI, E. CENCI, G. DEL SERO, F. BISTONI and P. PUCCETTI, J. Immunol. 158 2356Ð62 (1997). 240. M. WANG, A. SAXON and D. DIAZ-SANCHEZ, Clin Immunol 90 47Ð54 (1999). 241. R.F. GRAZIANO, R.J. LOONEY, L. SHEN and M.W. FANGER, J. Immunol. 142 230Ð5 (1989). 242. F. KIERSZENBAUM, S.J. ACKERMAN and G.J. GLEICH, Am. J. Tropical Med. Hygiene 30 775Ð9 (1981). 243. J. BUYS, R. WEVER, R. VAN STIGT and E.J. RUITENBERG, Eur. J. Immunol. 11 843Ð5 (1981). 244. S.J. ACKERMAN, G.J. GLEICH, D.A. LOEGERING, B.A. RICHARDSON and A.E. BUTTERWORTH, Am. J. Tropical Med. Hygiene 34 735Ð45 (1985). 245. H.A. MOLINA, F. KIERSZENBAUM, K.J. HAMANN and G.J. GLEICH, Am. J. Tropical Med. Hygiene 38 327Ð34 (1988). 246. F. LISZT, K. SCHNITTKER-SCHULZE, H.W. STUHLSATZ and H. GREILING, European Journal of Clinical Chemistry and Clinical Biochemistry 29 123Ð30 (1991). 247. E.E. GARDINER, S.S. MOK, A. SRIRATANA, H.C. ROBINSON, B.J. VEITCH, D.A. LOWTHER and C.J. HANDLEY, Eur. J. Biochem. 221 871Ð9 (1994). 248. A. EGESTEN, M. BLOM, J. CALAFAT, H. JANSSEN and E.F. KNOL, Int. Arch. Allergy Immunol. 115 121Ð8 (1998). 249. M.A. VADAS, N.A. NICOLA and D. METCALF, J. Immunol. 130 795Ð9 (1983). 250. N. NABAVI and J.W. MURPHY, Infect. Immun. 51 556Ð62 (1986). 251. A.M. SHENOY and Z. BRAHMI, Nat. Immun. Cell Growth Regulation 8 338Ð48 (1989). 252. E.W. FUSON, M.W. SHAW, R.A. HUBBARD, D.G. SUGANTHARAJ and E.W. LAMON, Exp. Hematol. 9 197Ð206 (1981). 253. M. RICHTER and S. SKLAR, Clin. Invest. Med. 6 19Ð24 (1983). 254. R.A. MUFSON, J. AGHAJANIAN, G. WONG, C. WOODHOUSE and A.C. MORGAN, Cell. Immunol. 119 182Ð92 (1989). 502 B. Spellberg / Life Sciences 67 (2000) 477Ð502 255. J. KHALIFE, D.W. DUNNE, B.A. RICHARDSON, G. MAZZA, K.J. THORNE, A. CAPRON and A.E. BUTTERWORTH, J. Immunol. 142 4422Ð7 (1989). 256. S. ITO, P. ANSARI, M. SAKATSUME, H. DICKENSHEETS, N. VAZQUEZ, R.P. DONNELLY, A.C. LARNER and D.S. FINBLOOM, Blood 93 1456Ð63 (1999). 257. K. STEINBRINK, M. W…LFL, H. JONULEIT, J. KNOP and A.H. ENK, J. Immunol. 159 4772Ð80 (1997). 258. Y.C. LIM, L. HENAULT, A.J. WAGERS, G.S. KANSAS, F.W. LUSCINSKAS and A.H. LICHTMAN, J. Immunol. 162 3193Ð201 (1999). 259. D.J. BERG, M.W. LEACH, R. K†HN, K. RAJEWSKY, W. M†LLER, N.J. DAVIDSON and D. RENNICK, J. Exp. Med. 182 99Ð108 (1995). 260. M.F. ROMANO, A. LAMBERTI, A. PETRELLA, R. BISOGNI, P.F. TASSONE, S. FORMISANO, S. VENUTA and M.C. TURCO, J. Immunol. 156 2119Ð23 (1996). 261. B.K. HALAK, H.C. MAGUIRE, JR. and E.C. LATTIME, Cancer Res. 59 911Ð7 (1999). 262. C. DUGAST, A. GAUDIN and L. TOUJAS, J. Leukoc. Biol. 61 517Ð21 (1997). 263. T.A. WYNN, R. MORAWETZ, T. SCHARTON-KERSTEN, S. HIENY, H.C. MORSE, 3RD, R. K†HN, W. M†LLER, A.W. CHEEVER and A. SHER, J. Immunol. 159 5014Ð23 (1997). 264. C.L. MANTHEY, J.B. ALLEN, L.R. ELLINGSWORTH and S.M. WAHL, Growth Factors 4 17Ð26 (1990).