Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Signal transduction wikipedia , lookup
Extracellular matrix wikipedia , lookup
Tissue engineering wikipedia , lookup
Cytokinesis wikipedia , lookup
Cell encapsulation wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cell culture wikipedia , lookup
Cell growth wikipedia , lookup
Cellular differentiation wikipedia , lookup
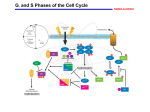
[Cell Cycle 4:4, 618-627; April 2005]; ©2005 Landes Bioscience The Regulation of S Phase Initiation by p27Kip1 in NIH3T3 Cells Report ABSTRACT Most eukaryotic cells become committed to divide once DNA synthesis begins. This critical event is controlled in yeast and Xenopus oocytes by the degradation of cyclin inhibitory proteins, while in mammalian cells over-expression of cyclin E or cyclin D1 promotes rapid entry into DNA synthesis. Curiously, however, this over-expression also lengthens S phase and promotes DNA damage. To directly assess the roles of the cyclin inhibitory protein p27Kip1 (p27) and of cyclin D1 in the regulation of DNA synthesis initiation in mammalian cells, we have utilized a quantitative cytometric approach for the study of cell cycle control in actively proliferating cultures. As evidence that p27 plays a direct role in regulating entry into S phase, we find that its levels fall at the time of DNA synthesis initiation in NIH3T3 cells, and that its suppression shortens G1 phase and shortens the length of the entire cell cycle. In this function, however, the action of p27 appears to be linked with cyclin D1. G1/S phase transition is efficiently blocked by p27 unless excess cyclin D1 is present. These two proteins are coordinately regulated by the cell, and are maintained at a nearly constant ratio throughout an actively proliferating culture. We propose that p27 directly regulates the initiation of DNA synthesis in NIH3T3 cells, and that cyclin D1 serves to modulate this activity. †Present address: Bose Institute; P-1/12 CIT Scheme VII M; Kolkata-700 054, India p27Kip1, cyclin D1, S phase, DNA synthesis, G1 phase, proliferative signaling, cell cycle ACKNOWLEDGEMENTS When environment conditions become conducive to the growth of mammalian cells, cyclin D1 levels are normally induced,1 while p27Kip1 (p27) levels are suppressed.2 These two molecular events then directly influence the activity of key cell cycle regulatory proteins. Cyclin D1 associated with cyclin dependent kinase (CDK) 4 or 6 inactivates the growth inhibitory retinoblastoma protein (Rb).3 This promotes the synthesis of cyclin E and a variety of other molecules required for DNA synthesis.4 P27, on the other hand, inhibits the activity of cyclin E bound to CDK2.5 In either case, proliferative signaling promotes the activation of cyclin E/CDK2, which is generally considered to be central in the control of S phase entry6, since its activity peaks at the G1-S phase transition,6,7 while inhibition of cyclin E function blocks the initiation of DNA synthesis.8 The importance of cyclin E/CDK2 in regulating G1/S phase transition is even more apparent when the mechanism of DNA synthesis initiation is considered.9 Soon after mitosis ORC proteins associate with regions of DNA destined to initiate DNA replication. These proteins then become associated with MCM proteins 2–7 to form the pre-replication complex (pre-RC).10 The pre-RC remains dormant until acted upon by CDC45, which opens up the DNA structure and allows association of proteins involved in DNA synthesis.11,12 Once the first pre-RC becomes activated, mammalian cells are normally committed to rapidly complete replication of the entire genome and pass through mitosis. Thus, the activation of CDC45 and related proteins13 is key to our understanding of the events that control the initiation of DNA synthesis and entry into the cell cycle. Two kinases control the activation of CDC45, cyclin E/CDK2 and CDC7/Dbf1.14,15 Cyclin E/CDK2 is the target of a complex array of regulatory events. It is inhibited by the cyclin inhibitory proteins p21Waf1 and p27, and by phosphorylation on Thr14/Tyr1516,17 (which is accomplished by the mammalian Wee1 analogue). These inhibitory phosphates are removed by the critical cell cycle regulatory protein CDC25A,18,19 whose expression is controlled by cell cycle progression and DNA damage. Activation of cyclin E/CDK2 also requires phosphorylation on Thr 160 by the cyclin H/CDK 7 protein complex;20 which also functions as a core transcription factor,21 and whose kinase activity and substrate specificity are regulated by binding to MAT1,22,23 and by proliferative signaling.24 Finally, the inhibitory capacity of p27 for cyclin E/CDK2 is reportedly reduced upon binding to cyclin D1.25 © 20 05 LA ND ES BIO SC IEN Acknowledgements. We thank M. Hitomi, M. Pagano, and K. Yang for helpful criticisms of this manuscript, and J. Harwalkar, J. Fretthold and A.W. Stacey for assistance throughout. Supported by NIH grant GM-52271. INTRODUCTION .D KEY WORDS CE Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/abstract.php?id=1632 ON OT D Received 02/24/05; Accepted 02/25/05 RIB *Correspondence to: Dennis W. Stacey Ph.D.; Department of Molecular Genetics; The Lerner Research Institute; The Cleveland Clinic Foundation; 9500 Euclid Avenue; Cleveland, Ohio 44195 USA; Tel.: 216.444.0633; Fax 216.444.0512; Email: [email protected] IST Department of Molecular Genetics; The Lerner Research Institute; The Cleveland Clinic Foundation; Cleveland, Ohio USA UT E . Gaurisankar Sa† Yang Guo Dennis W. Stacey* 618 Cell Cycle 2005; Vol. 4 Issue 4 p27Kip1 and the Initiation of DNA Synthesis Each of these regulatory features of cyclin E/CDK2 is potentially a target for the action of signaling pathways involved in cell cycle regulation. The variety of mechanisms of CDC45 control might indicate the number of different mechanisms required for cell cycle regulation during the generation, maintenance, and repair of a vast range of organs and tissues. In yeast and in Xenopus oocytes, G1/S phase transition has been shown to depend upon degradation of the cyclin inhibitory proteins Sic1 or Xic1.26-28,29 Neither system presents the proliferative demands of the mammalian somatic cell, but their behavior suggest the possibility that p27, or another cyclin inhibitory protein, might be involved in regulating the initiation of DNA synthesis in mammalian cells under some circumstances. Accordingly, we devised a series of experiments to determine to what extent, and under what circumstances p27 might be involved in directly regulating the G1/S phase transition in actively proliferating NIH3T3 cells. Our approach was based upon techniques utilized in the study of cyclin D1 expression and function through the cell cycle.30,31 Previous studies were performed in asynchronous cultures to avoid the necessity of interfering with cell cycle progression. The cell cycle position of each actively cycling cell was determined based upon its DNA content, age, and BrdU labeling characteristics. These in turn were assessed with the use of quantitative image analysis, microinjection and time-lapse microscopy. With this approach we have shown that proliferative signaling induces the expression of cyclin D1 primarily during G2 phase.30 Once induced during G2 phase, the expression of cyclin D1 remains relatively constant through G1 phase where it is required for the commitment to another round of replication.32 Upon entry into S phase, cyclin D1 levels rapidly decline.31 In the work reported here, this approach was utilized to study the expression of p27 through the cell cycle. We present evidence that p27 regulates the rate of passage through G1 phase, and entry into S phase in NIH3T3 cells. Its action, however, is regulated by the coordinate expression of cyclin D1 within the cell. Digital image analysis. The procedures for staining, image collection and data analysis have been previously described.33 Briefly, cells were fixed with 3% paraformaldehyde/0.05% Triton X100 and then with chilled methanol for 2 min (p27), with chilled methanol alone (cyclin D1 and Skp2) and stained with an indirect antibody stain against the target antigen, and finally stained with DAPI (10 µg/ml). Images of each fluorochrome were taken with a cooled CCD camera (Roper Scientific) with the appropriate filters (Chroma) of the same area of cells. The shading of each image was corrected with a uniformly stained specimen; and the DAPI image was utilized for thresholding with the Metamorph software (Universal Imaging) to identify nuclei of individual cells. The fluorescence in this region was then quantitated for each fluorochrome. The cell cycle related expression of each marker was then presented by plotting the DNA content of each cell vs. the fluorescent intensity of the marker being measured. In the case of cells expressing green fluorescent protein (GFP) the GFP levels were determined directly in living cells just prior to fixation. The average expression level of a marker in a single experiment was determined by grouping all the cells within a narrow DNA range together and determining the mean (±SE) fluorescence intensity. This level was then plotted vs. the average DNA level. Cells were divided into different cell cycle periods based upon DNA content and BrdU labeling. The average of each group was then determined. Injected cells were identified by position on the cover slip as indicated by a circle drawn on the back of the coverslip with a Leitz object marker, or by expression of a fluorescence marker. In most instances comparisons were made between injected cells and neighboring uninjected cells. This was the case even when injections of control protein were performed. This approach ensures that the staining and culture conditions were identical for all the cells compared in a single digital image analysis. This would avoid any complications, which might result from subtle staining differences between separate plates. Thus, when an active reagent and a control protein were being compared, the comparison was first between injected cells and neighboring uninjected cells, and then this result was compared for the active and control injections. In some conditions four fluorescent images were analyzed for a single set of cells. The fluorochromes involved in this analysis included CY2, CY3, CY5 and DAPI staining. Cell culture and microinjection. NIH3T3 cells were maintained in 10% Calf Serum and rendered quiescent by culture for 48 hrs in 0.5% serumcontaining medium. Other cells were cultured in 10% fetal calf serum. Microinjection was performed as previously described31 using back loaded pipets with continuous flow. Injections were performed to be as nearly uniform in size as possible, and to involve all cells in an area defined by a faint circle drawn on the back of a coverslip with a Leitz Object Marker. No adverse effects of any of the injections performed were identified. Injections of plasmid DNA (1.5–10 µg/ml) were nuclear, while injections of siRNA (20 µg/ml) were cytoplasmic. All siRNAs are obtained from Dharmacon Research Inc. SiRNA against human p27 coding sequence is designed to target the sequence AACGCUGGCACUGUGGAGCAG. The targeted sequence for cyclin D1 siRNA is AACUACCUGGACCGCUUCCUG. SiRNA for Skp2 is AAUGGGAUUCCAGCAAGACUU. Plasmid construction. The coding sequence of human cyclin D1 without stop codon was amplified by PCR. The amplified segment was inserted into the mammalian expression vectors pd2EGFP-N1 and pCMV-HA (from Clontech). The human p27 coding sequence without stop codon was cloned and inserted into pd2EGFP-N1 vector as well. Time lapse analysis. For time lapse observations cells were observed at 250X magnification under tungsten illumination. Cells were exposed to light only during collection of images. Images were taken every 20 minutes with a cooled CCD camera, and animated as a stack by NIH Image software. A single fluorescence image of the GFP expression of the same area of cells was taken during the time lapse movie by changing the illumination and filter system. Division was scored as the point at which the cell became most rounded. p27 levels drop upon entry into S phase. To test the possibility that p27 plays a direct role in regulating the initiation of DNA synthesis, we analyzed its expression level through the cell cycle. We assumed that p27 levels should be high enough to block cyclin E/CDK2 activity until DNA synthesis begins, at which time p27 levels would fall dramatically. To test this prediction we first studied p27 expression levels after quiescent NIH3T3 cells were stimulated to re-enter the cell cycle. At various times following serum stimulation these cells were fixed and stained with a fluorescent antibody against p27, while DNA was stained with DAPI. Images taken with a CCD camera of each fluorochrome were then analyzed. The DAPI image was used to generate a mask able to identify the location of each cell within the field. This mask was then used to accurately quantitate the level of each fluorochrome in each cell. As in past studies,31 care was taken to ensure that the p27 fluorescence level within the entire culture was proportional to the absolute p27 protein level (data not shown). When the p27 level of each individual cell was plotted against its DNA content, it was clear that p27 expression in G0/G1 phase was reduced with time following serum addition (Fig. 1A). Only cells with extremely low levels of p27 entered S and G2 phases. This analysis was performed in the opposite way by following cells as they approached confluence. At various times following plating cultures were fixed, stained and analyzed as above. The p27 levels were low in all actively cycling cells, but increased during G1 phase as cultures approached confluence. Critically, even in cultures approaching confluence, the levels of p27 remained low in those cells that continued to pass through S and G2 phases (Fig. 1B). Our next analysis focused upon actively cycling cells. An asynchronous culture of NIH3T3 cells was pulsed with BrdU prior to fixation and stain- MATERIALS AND METHODS www.landesbioscience.com RESULTS Cell Cycle 619 p27Kip1 and the Initiation of DNA Synthesis A Figure 1. Cell cycle regulation of p27 expression in NIH3T3 cells: (A) Quiescent NIH3T3 cells were stimulated to re-enter the cell cycle by serum addition for the times indicated prior to fixation, staining and image analysis. The p27 level of each cell is plotted vs. its DNA content. (B) NIH3T3 cells were allowed to grow to confluence. Cells at day 2 were actively cycling, had begun to enter quiescence at day 3, and by day 4 had almost all become arrested in cell cycle progression. As above, the p27 level of each cell at each time point is plotted vs. its DNA content. B ing of p27 and BrdU with fluorescent antibodies, and for DNA with DAPI. The p27 level of each cell was plotted against its DNA content, with BrdUlabeled, S phase cells noted. While p27 levels were low in all cell cycle phases of these rapidly proliferating cells, there was a definite decline as the cells passed from G1 to S phase (Fig. 2A). To understand the kinetics of this change, NIH3T3 cells were followed in time-lapse for 30 hrs prior to a brief pulse with BrdU, fixation, staining and image analysis. Cells were separated into cell cycle phases based upon DNA and BrdU content. The age of each cell, or the time since passing through mitosis, was then plotted vs. its p27 level for each cell cycle phase (Fig. 2B). It was apparent that p27 levels were low immediately following mitosis, but increased through G1 phase, prior to A falling rapidly upon entry into S phase. This fact was even more apparent when the average p27 levels for each cell cycle stage were plotted vs. age (Fig. 2C). Those cells that remained in G1 phase for longer periods of time displayed continuously increasing average p27 levels. The suppression of p27 upon entry into S phase might be due, at least in part, to phosphorylation by CDK2 and ubiquitination by the SCF complex containing the Skp2 F-box protein.34 The increase during G1 phase is most probably due to the destruction of Skp2 by the APC ubiquitin complex during G1 phase.35,36 Suppression of p27 levels speeds passage through G1 phase. The above results indicate that p27 levels are not only lower in S phase than in G1 phase, it is clear that p27 levels are suppressed near the time of G1/S phase transition. This provides direct support for the notion that p27 levels might play a direct role in regulating the initiation of DNA synthesis in NIH3T3 cells. To gain further evidence for this possibility, the ability of p27 to regulate the rate of passage through G1 phase was next determined. An siRNA able to suppress p27 levels was microinjected into NIH3T3 cells. This siRNA reduced p27 levels in all cell cycle phases at both 6 and 12 hrs following its microinjection (Fig. 3A), but had little effect upon cyclin D1 B C Figure 2. Cell cycle expression of p27: (A) An asynchronous NIH3T3 culture was pulsed with BrdU, fixed, stained and subjected to image analysis. The p27 level of each cell is plotted vs. its DNA content, with BrdU positive cells noted with solid circles. (B) A cycling culture was followed in time-lapse 30 hrs prior to a pulse of BrdU, fixation and image analysis. Age is the time between mitosis and the end of the movie, and is plotted vs. p27 level. The cell cycle phase of each cell was determined from DNA content and BrdU labeling, and is indicated by separate symbols. (C) The same data in the previous analysis was reanalyzed by separating cells according to cell cycle phase and then grouping cells according to age. The average p27 level for each cell cycle phase was determined for each age group, and plotted against age. In each experiment p27 fluorescence levels are represented by arbitrary units which vary between experiments. 620 Cell Cycle 2005; Vol. 4 Issue 4 p27Kip1 and the Initiation of DNA Synthesis A B C D E F Figure 3. Suppression of p27 levels shortens G1 phase. (A-C) NIH3T3 cells were microinjected with siRNA against p27 (20 µg/ml), and at 6 or 12 hrs thereafter were fixed, stained, and analyzed. The p27 expression (A) and cyclin D1 expression (B) of injected cells in each cell cycle phase can be compared to neighboring uninjected cells for the same time period. The proportion of injected and uninjected cells in each cell cycle phase is also presented for each time point (C). (D-F) NIH3T3 cells were injected with siRNA against p27 and followed in time-lapse for 15 hrs prior to a pulse with BrdU, fixation, staining and analysis. Only injected (large, open circles) and uninjected (small solid circles) cells in G1 phase are presented. The age of the cells is plotted vs. p27 level (D), cyclin D1 level (E) or against the ratio of p27 level/cyclin D1 level (F). The data from this experiment is almost identical to that from two other analyses. levels (Fig. 3B). Significantly, the proportion of cells in S phase increased dramatically following siRNA injection (Fig. 3C). To understand this increase in the S phase population, NIH3T3 cells were microinjected with the siRNA and followed in time-lapse for 15 hrs prior to a pulse with BrdU, fixation and staining. S phase and G2 phase cells were identified by DNA and BrdU content, and eliminated from the analysis. Thus, the plot of p27 level vs. age included only cells in G1 phase (Fig 3D–F). As expected, the siRNA-injected cells displayed lower p27 levels than neighboring uninjected cells (or control injected cells, data not shown). Most injected as well as uninjected cells passed through G1 phase and disappeared from this analysis within 6–8 hrs following mitosis. Among the uninjected cells, however, a number displayed high p27 levels and remained in G1 phase for much longer periods of time. The absence of such cells among the siRNA injected population indicated that the reduction in p27 level had promoted rapid passage through G1 phase (Fig. 3D). Consequently, the average G1 length was decreased following siRNA injection in this and two other analogous experiments (average age of G1 phase cells 3.9 ± 0.2 hrs for injected, compared to 5.4 ± 0.4 hrs for uninjected cells). In independent studies the levels of Skp2 in G1 phase were increased by modulating APC activity, resulting in the suppression of G1 phase p27 levels.35,36 Similarly, the expression level of p27 was also suppressed by blocking expression of a polypyrimidine tract-binding protein involved in internal ribosomal initiation.37 In both cases the effect upon p27 expression was indirect, and other molecules might also have been effected, yet the cells responded with shortened G1 length as we observed following direct suppression of p27 levels in this study. The critical finding here is the fact that reduction in p27 levels altered the rate of passage through G1 phase only in that portion of all cells that would have remained in G1 phase longer www.landesbioscience.com than 6–8 hrs. Among the cells with naturally short G1 length there was little noticeable effect. In NIH3T3 cells, therefore, there appeared to be two mechanisms of regulating G1/S phase transition. One was evident in cells with low p27 levels and ensured that cells remain in G1 phase at least 6–8 hrs. Increased p27 levels were able to further delay entry into DNA synthesis beyond this minimal length. In the above siRNA injection experiment, a plot of p27 vs. age showed that p27 levels increased among those cells remaining in G1 phase (Fig. 3D). There was, however, no obvious relationship between age and cyclin D1 levels in the same cells (Fig. 3E). Nevertheless, as predicted by previous studies,38,39 microinjection of a cyclin D1 expression vector (tagged with the green fluorescence protein (GFP) at its carboxyl terminus) also promoted rapid passage through G1 phase (data not shown). The ability of cyclin D1 to reduce G1 phase length might result from the inactivation of the Rb protein, which would otherwise block cyclin E synthesis, and which can also directly interfere with MCM proteins.40 Additionally, cyclin D1 might function indirectly by modulating the activity of p27.25 To gain more information on the cell cycle regulatory activities of p27 compared to cyclin D1, experiments were performed to determine the effect upon the rate of passage through the entire cell cycle of either suppressing p27 or elevating cyclin D1. The siRNA against p27 was microinjected into NIH3T3 cells, which were then followed in time lapse (beginning 6 hrs thereafter) to determine the length of the cell cycle following p27 suppression. The siRNA injected cells displayed cell cycle lengths of 18–22 hrs (Fig. 4A), while among the uninjected (or mock injected cells, data not shown), a high proportion of the culture displayed cell cycle lengths much greater than 22 hrs (Fig. 4C). As above, this indicates that p27 is rate limiting for cell cycle progression, but only among those cells with slow rates of proliferation. Cell Cycle 621 p27Kip1 and the Initiation of DNA Synthesis A B C D Figure 4. Regulation of cell cycle length by p27 and cyclin D1. (A, C) The siRNA against p27 (20 µg/ml) was microinjected into NIH3T3 cells. Injected cells, together with an equal number of uninjected cells, were followed in time lapse beginning 6 hrs thereafter. The time between mitoses for each cell was determined, and the results presented as a histogram representing the number of injected (A) and uninjected (C) cells with each cell cycle length. This is the combination of three separate experiments, each with similar profiles. (B, D) The cyclin D1-GFP expression plasmid (10 µg/ml) was injected into NIH3T3 cells which were followed in time lapse beginning 2 hrs thereafter. At 16 hrs following injection a single fluorescence photograph revealed the level of cyclin D1-GFP expression in each cell. The movie was followed to determine cell cycle lengths for each cell. The cyclin D1-GFP fluorescence is plotted vs. cell cycle length for each cell of an individual injection (B) or for the average results from 4 separate injections (D). Elevation of cyclin D1 levels has been reported to slow passage through the cell cycle, despite its ability to shorten G1 length.41 To verify these findings, a cyclin D1-GFP expression vector was introduced into cells that were followed in time lapse as above. A single fluorescence image 16 hrs following injection indicated the levels of cyclin D1-GFP expression in each cell. The expression of cyclin D1 increased the length of the cell cycle (Fig. 4B and D). The extent of this increase was roughly proportional to the expression level of cyclin D1 within the cells. This was observed in individual injected cells (Fig. 4B), together with the average of all cells in four separate experiments (Fig. 4D). Injections of a GFP expression plasmid alone did not inhibit cell cycle progression (not shown). Inhibition of cell cycle progression by cyclin D1 might relate to its ability to block the activity of PCNA, which is required for DNA synthesis.42,43 It is clear from this result that cyclin D1 does not function on its own in NIH3T3 cells to regulate the rate of cell cycle progression. p27 inhibits cell cycle progression. Evidence was presented above to indicate that p27 is rate limiting for cell cycle progression in only a subset of A B C D 622 NIH3T3 cells, presumably those with higher p27 levels. To determine if p27 could inhibit cell cycle progression even in those cells where it is not normally rate limiting, p27 levels were increased in cells and their proliferative capacity analyzed. A plasmid containing a viral promoter driving expression of a p27-GFP chimeric protein was microinjected into NIH3T3 (Fig. 5A) cells at a concentration known to induce average p27 levels in the range of quiescent cells (1.5 µg/ml), and to remain nuclear (Fig. 6D). Injected cells were followed in time lapse for 15 hrs, pulsed with BrdU, and photographed to determine the level of p27-GFP fluorescence. Of the cells that passed through mitosis during the analysis, those also able to enter S phase were identified by BrdU labeling. Most p27-expressing cells failed to enter S phase (Fig. 5A). Those few that did expressed the lowest detectable levels of p27-GFP. To confirm this result, the p27-GFP expression plasmid was microinjected into NIH3T3 cells that were followed in fluorescence timelapse for 30 hrs. Mitosis was rarely observed, and only in cells with the lowest levels of GFP expression detectable (data now shown). Clearly, even slightly elevated p27 levels inhibit passage through G1 phase in essentially all Figure 5. p27 and cyclin D1 function together in cell cycle control. (A) The p27-GFP expression vector (1.5 µg/ml) was injected alone, or (B) together with a 3-fold excess of a plasmid expressing cyclin D1 tagged with HA. Cells were followed for 15 hrs in time lapse, pulsed with BrdU and photographed to determine p27-associated GFP fluorescence levels. The GFP expression of all cells (that passed through mitosis during the time-lapse analysis) is presented as a histogram, with cells able to enter S phase following the injection indicated in solid. (C) Cells injected as above with the p27 expression vector alone, or in combination with a three-fold excess of the cyclin D1 vector, were followed in time-lapse for an extended time. The number of cells with each cell cycle length (hr) is plotted as a linear histogram for cells following each treatment, or for neighboring uninjected cells. (D) A similar cell cycle analysis is presented except that the cyclin D1 vector alone (10 µg/ml), or in combination with the p27-GFP vector (1.5 µg/ml), was injected and the cell cycle length determined. In this case, the average cell cycle length of injected cells is presented as a proportion of the cell cycle length observed in neighboring uninjected cells. Injection of a control vector had no influence upon cell cycle length (data not shown). Cell Cycle 2005; Vol. 4 Issue 4 p27Kip1 and the Initiation of DNA Synthesis Figure 6. Coordinate regulation of p27 and cyclin D1. (A-B) The p27-GFP expression vector was injected into NIH3T3 cells and 6 hrs later the cells were analyzed for p27 (A) and cyclin D1 (B) expression. For this analysis, cells were grouped according to DNA content and BrdU labeling, and the average staining level for each group plotted vs. DNA content. The profile of injected and uninjected neighboring cells is presented for each analysis. (C) In a similar analysis, a cyclin D1-GFP expression vector (5 µg/ml) was injected and 12 hrs later the cyclin D1-associated GFP fluorescence was photographed, and the cells fixed and stained for endogenous p27. The average GFP fluorescence and p27 levels for each cell cycle phase are presented for injected and neighboring uninjected cells. (D) The p27-GFP expression vector at 1.5 µg/ml was injected into NIH3T3 cells, which were photographed 10 hrs later in phase contrast or fluorescence optics to reveal GFP-fluorescence. Mock injections are indistinguishable from uninjected cells in these and other experiments. A B C D NIH3T3 cells. On the other hand, suppression of cyclin D1 to less than half the normal level following microinjection of an siRNA against cyclin D1 had little effect upon the length of the cell cycle in NIH3T3 cells (data not shown). Cyclin D1 reduces the inhibitory potential of p27. Notwithstanding their differing abilities to regulate the rate of passage through the cell cycle, there is a close physical and functional relationship between p27 and cyclin D1. The cyclin D1/CDK4 complex can bind and neutralize the inhibitory activity of p27,44 while p27 can promote the association of the cyclin D1/CDK4 complex.45,46 It is therefore, possible that cyclin D1 might have been able to speed passage through G1 phase indirectly by neutralizing the activity of p27. The ability of cyclin D1 to neutralize the inhibitory activity of p27 was, therefore, tested by injecting the p27-GFP expression plasmid combined with a 3-fold excess (4.5 µg/ml) of a plasmid expressing the cyclin D1 protein (tagged with hemagglutinin antigen (HA) at its amino terminus). Cells were followed 15 hr in time-lapse, photographed, and analyzed for passage into S phase as above. The co-injected cyclin D1 partially overcame the inhibitory influence of p27 and allowed entry into S phase for cells that expressed sufficient p27-GFP to have otherwise blocked DNA synthesis (Fig. 5B). The ability of cyclin D1 expression to overcome the inhibitory influence of p27 was further investigated by determining the length of the cell cycle following these injections. The p27 alone induced a dramatic increase in cell cycle length that was partially reversed by co-injected cyclin D1 (Fig. 5C). When the concentration of cyclin D1 was increased to 10 µg/ml, the inhibition of p27 (1.5 µg/ml) was nearly eliminated, but at this concentration the inhibitory effects of cyclin D1 alone upon the cell cycle became apparent (Fig. 5D). A careful analysis of the above data indicates that the effect of cyclin D1 upon p27 extends beyond simple neutralization of inhibitory potential. When the cyclin D1-HA expression plasmid was co-injected with the p27GFP expression plasmid, the expression of p27-GFP itself was stimulated (Fig. 5A and B). If cyclin D1 functioned simply to neutralize the inhibitory effects of p27, it would not be expected to induce an increase in p27 expression. Further studies were, therefore, performed to determine if elevated cyclin D1 expression was able to have a similar stimulatory effect upon endogenous p27 expression. The cyclin D1-GFP expression plasmid (5 µg/ml) was microinjected into NIH3T3 cells that were photographed 12 hrs later to determine the level of cyclin D1-GFP expression, followed by staining for endogenous p27 levels. The endogenous p27 had increased dramatically in the cyclin D1 plasmid-injected cells (Fig. 6C). Likewise, the p27-GFP expression plasmid was introduced into NIH3T3 cells, which were fixed 6 hrs later and stained for both p27 and endogenous cyclin D1. The levels of both had increased dramatically (Fig. 6A and B). Similar results have been reported.46,47 It is not clear how this mutual regulation might be achieved, although it was observed with p27-GFP expressed from www.landesbioscience.com a plasmid with viral regulatory elements, and therefore is likely to involve protein stability. As additional evidence for the importance of the interaction between p27 and cyclin D1 in regulating passage through G1 phase, the data obtained following the siRNA injection described above was re-evaluated (See Fig. 3). Recall that p27 levels increased progressively in uninjected cells that remained longest in G1 phase (Fig. 3D). When the ratio of p27 expression to cyclin D1 expression was plotted vs. age, however, the profile was different (Fig. 3F). Rather than progressively increasing with the length of G1 phase, the p27/cyclin D1 ratio exhibited a wide variation among siRNA injected and uninjected cells early in G1 phase. With time, cells with lower ratios apparently entered S phase and disappeared from the analysis, leaving only those cells with highest p27/cyclin D1 ratios in late G1 phase. While further study will be required to fully understand this phenomenon, it is possible that the eventual length of G1 phase was determined early in this cell cycle period by the ratio of p27 to cyclin D1. The ratio of p27 to cyclin D1 is constant in actively cycling cells. To further emphasize the importance of the p27/cyclin D1 ratio in growth regulation, the p27 levels (Fig. 7A) and cyclin D1 levels (Fig. 7B) were determined in the same actively cycling NIH3T3 cells, and separately plotted against DNA level. In past studies it has been difficult to explain why cells with similar proliferative rates could express such wide variations in either cyclin D1 or p27 levels during G1 phase. As a potential explanation for this observation, we found that those cells with high cyclin D1 levels were the same cells with high p27 expression levels, and vice versa (Fig. 7C). Perhaps it is not so much the absolute levels of either protein that determines the proliferative properties of a cell, but their relationship to each other. To test this possibility, p27 levels were plotted against cyclin D1 levels in the same cells (Fig. 7C). The resulting profile indicated that the ratio between these two cell cycle regulatory molecules was remarkably uniform throughout the actively cycling culture (Fig. 7C). This result was repeated numerous times with another established mouse line (Balb3T3), and in two human diploid fibroblast lines (MRC5 and IMR90). The uniform expression of p27 and cyclin D1 within the same cells might be due in part to the overall variations in basal transcription activity within individual cells of the culture.48,49 Because protein-protein interactions are apparently involved (see above), however, we consider it more likely that the constant ratio of these two proteins results from their interaction together and mutual protection against proteolytic degradation. Cell Cycle 623 p27Kip1 and the Initiation of DNA Synthesis A B C D E F Figure 7. Coordinated expression of p27 and cyclin D1: Cycling NIH3T3 cells were stained for both cyclin D1 and p27. The cyclin D1 (A) and p27 (B) levels of each cell were plotted vs. DNA level; or (C) the p27 level for each cell was plotted vs. its cyclin D1 level. (D–F) The p27 expression level was then plotted vs. cyclin D1 expression as above, but at the indicated times following serum removal from NIH3T3 cells. The overall ratio of p27 to cyclin D1 was maintained in the culture until the growth environment of the cells changed, at which time the ratio was altered. Thus, when actively cycling cells (Fig. 7D) were deprived of serum for 12 hrs (Fig. 7E) or 18 hrs (Fig. 7F) the ratio of p27 to cyclin D1 expression was dramatically increased. Similar results were observed as cells approached confluence (data not shown). We conclude that cyclin D1 levels play a major role in determining the inhibitory capacity of p27 upon G1/S phase transition. As a direct test of this hypothesis, we microinjected siRNA against the protein responsible for degradation of p27 during S and G2 phases, Skp2.34 This siRNA suppressed Skp2 expression rapidly and for an A extended period of time (Fig. 8A). Yet, neither the length of the cell cycle (not shown) nor the proportion of cells in S phase (Fig. 8C) was changed in these injected cells. As expected, injection of siRNA against Skp2 increased p27 levels in all cell cycle phases, but it also increased cyclin D1 levels (Fig. 8B). These two events had apparently compensated, yielding a consistent p27/cyclin D1 ratio and normal proliferation. B C Figure 8. Microinjection of siRNA against Skp2. (A) siRNA against Skp2 was injected into NIH3T3 cells which were stained for Skp2 40 hrs later. An area of the plate was photographed in phase contrast and fluorescence optics so as to reveal both injected and uninjected areas. (B) siRNA vs. Skp2 was injected into NIH3T3 cells that were fixed and stained 12 hrs later. Injected and neighboring uninjected cells were grouped according to cell cycle phase, and the average p27 levels or cyclin D1 levels for each group are presented. (C) siRNA vs. Skp2 was injected into six separate plates in two experiments. The proportion of cells in each cell cycle phase for injected and neighboring uninjected cells determined. The ratio of injected/uninjected cells for each cell cycle period in all 6 experiments is presented (±S.E.). 624 Cell Cycle 2005; Vol. 4 Issue 4 p27Kip1 and the Initiation of DNA Synthesis DISCUSSION From these results we conclude that p27 functions to regulate the timing of DNA synthesis initiation in NIH3T3 cells. This conclusion is based upon the fact that the length of G1 phase and the overall length of the cell cycle can be sensitively regulated by the levels of p27, both positively and negatively. It is further supported by the observation that p27 levels invariably fall upon entry into S phase, regardless of the growth conditions. Moreover, this decline in p27 levels takes Figure 9. Model for p27 regulation of G /S phase transition. The following illusplace at the time of entry into S phase. In this way, cell cycle tration is presented to explain the regulation1 and function of p27 and cyclin D1 from progression is governed in mammalian cells similarly to yeast, mitosis through S phase. Cyclin D1 levels are elevated during G2 phase and remain where degradation of the cyclin inhibitory protein Sic1 regu- relatively constant, until they fall after entry into S phase. We assume p27 synthesis lates entry into S phase.27,28 In Xenopus oocytes, degradation is relatively constant throughout the cell cycle, and that its levels are regulated priof a similar protein, Xic1 is required for the initiation of marily by protein stability. In mitosis, p27 levels are suppressed by Skp2 and CDK2 DNA synthesis, although this degradation is not dependent phosphorylation. At anaphase Skp2 and S phase cyclins are destroyed, allowing upon phosphorylation by CDK2 as is the case for p27 in the increase in p27 levels. These levels increase through early G1 phase until they reach the level able to be protected by binding to cyclin D1. Any increase beyond mammalian cells.50,51 Despite these similarities, the prolifer- this level in late G phase will be suppressed by a proliferative signaling-directed, 1 ative requirements in yeast and oocytes are obviously differ- p27 degradation mechanism (that does not involve Skp2). This will maintain p27 ent than in mammalian somatic cells. This is reflected in the levels in rough proportion to cyclin D1 levels, and below those required to inhibit fact that we find evidence for a mechanism to control DNA cyclin E/CDK2. In such cells, p27 will not influence cell cycle progression, and entry synthesis initiation that can be observed even with low p27 into S phase will take place without delay when other cellular requirements are met. levels. This mechanism apparently determines the minimal On the other hand, if proliferative signaling is weak, p27 levels will be allowed to increase beyond the protective capacity of cyclin D1. This extra p27 will be availlength of G1 phase, presumably to allow time to attain the able to bind and inhibit cyclin E/CDK2 and thereby retard or block entry into S size and mass required for initiation of another round of cell phase. Upon entry into S phase both p27 and cyclin D1 levels fall. In this way, procycle progression. The effect of higher p27 levels is to extend liferative signaling during G1 phase determines if p27 is able to inhibit entry into S G1 phase beyond this minimal length through its inhibitory phase. activity. In those cells where conditions are optimal for cell cycle progression, p27 has apparently a diminished role in cell cycle these two molecules is nearly constant, so long as the proliferative regulation. In such cells p27 levels are apparently low enough that environment of the cell remains optimal. It is the lack of proliferacyclin E/CDK2 can become activated and promote firing of the pre- tive signaling that leads to an imbalance in this ratio, in favor of p27. RC as soon as the metabolic requirements for cell cycle progression The need for this pattern of mutual regulation might depend upon are met. the profound inhibitory capacity of p27. We find even the lowest The influence of p27 upon the initiation of DNA synthesis can detectable levels of exogenous p27 efficiently block entry into S be compared to that of cyclin E and cyclin D1, which have sepa- phase. The normal co-regulation of cyclin D1 might be a means for rately been postulated to regulate progression through G1 the cell to temper the inhibitory influence of p27. Thus, changes in phase.38,52-54 In this and other studies both molecules hasten passage p27 expression are modulated by the presence of cyclin D1, resultthrough G1 phase when overexpressed. Moreover, both are absolute- ing in gradual alterations in the overall inhibitory capacity of p27 as ly required in most somatic cells for progression through G1 its levels vary. In this way, proliferative signaling is able to use a phase8,30 (despite the fact that their functions can be taken over by potent inhibitor to effect marginal positive and negative changes in other molecules in germ-line knockouts55-61). The effects of these the rate of proliferation, while maintaining the ability to completely two cyclins might result indirectly from their ability to influence p27 block proliferation when conditions dictate. These observations also levels. On the other hand, while over-expression of cyclin E or cyclin help explain why the levels of cyclin D1 and of p27 vary so dramatD1 shortens G1 phase, it also lengthens the cell cycle and promotes ically between individual cells in an actively proliferating culture. It DNA damage.41,62,63 This negative effect is possibly due to the pre- is apparently the ratio of p27 to cyclin D1, or a similar relationship, mature activation of the pre-replication complex.64 which is critical to cell cycle progression. It is most likely, therefore, that under normal circumstances As evidence that it is the relationship of p27 to cyclin D1, rather cyclin D1 influences passage through G1 phase first by neutralizing than the absolute level of either that determines cell cycle progresthe inhibitory influences of Rb, and second by modulating the sion, p27 levels were increased in NIH3T3 cells by ablation of Skp2 inhibitory activity of p27. It has been proposed that cyclin by siRNA. Despite the fact that p27 levels increased in these cells, D1/CDK4 binds p27 and thereby titrates it away from cyclin they continued to proliferate normally, presumably because cyclin E/CDK2; resulting in activation of cyclin E/CDK4, which then pro- D1 levels were also elevated, and were able to modify the inhibitory motes entry into and passage through S phase.25,65 Based upon our influence of p27. A different result has been reported in HeLa cells, studies, we view this relationship from a slightly different prospec- where suppression of Skp2 lengthened the cell cycle.35 HeLa cells, tive. If the elevation of cyclin D1 were designed simply to suppress however, do not require cyclin D1 activity for growth due to the p27 activity, the two would not be expected to be coordinately reg- neutralization of Rb function by a viral protein. In these cells it is ulated by proliferative signaling. This, however, is an unmistakable likely that suppression of Skp2 resulted in the elevation of p27 levconclusion from our studies. The elevation of cyclin D1 induces an els without a corresponding effect upon cyclin D1 activity, resulting elevation of p27 levels, and vise versa (see also refs. 46 and 47). in cell cycle inhibition. Moreover, in all the normal cell lines we have tested, the ratio of www.landesbioscience.com Cell Cycle 625 p27Kip1 and the Initiation of DNA Synthesis We present the following model to explain these and other results relating to the role of p27 and cyclin D1 expression in the control of DNA synthesis initiation. It is known that p27 levels during S, G2, and early M phase are suppressed by the action of CDK2 phosphorylation and Skp2 ubiquitination.34 Beginning at anaphase and continuing through G1 phase, CDK2 activity and Skp2 protein are eliminated from the cell,35,36 resulting in an increase in p27 levels through G1 phase as seen here (see Fig. 2B and 2C). In separate studies33 we found that p27 levels are suppressed during G1 phase by a signaling pathway involving Ras, MEK, and potentially phosphatidylinositol-3 kinase (see refs. 66, 67 and 68). This might involve the recently identified ubiquitin ligase targeted to p27 following stimulation of quiescent cells.69 This mechanism does not apparently block the increase in p27 levels during G1 phase because of the protective influence of cyclin D1, which shields p27 from degradation. However, when the protective capacity of cyclin D1 binding is reached, further production of p27 is subject to inhibition by this proliferative signaling-directed degradation mechanism. In optimal growth conditions, most cells suppress p27 to the level able to be protected by cyclin D1, resulting in a constant ratio of the two proteins. Such cells are able to enter S phase without interference from p27, since binding to cyclin D1 not only protects p27 from degradation, it also restrains p27 from associating with and inhibiting the activity of cyclin E/CDK2. If the growth conditions are not optimal, however, the Ras-MEK pathway of p27 degradation does not operate, p27 accumulates beyond the protective capacity of cyclin D1, and p27 levels are able to block the activity of cyclin E/CDK2 and therefore the activation of the pre-RC. The action of growth factors would then be required to suppress p27 levels sufficiently to allow cyclin E/CDK2 to become activated if such a cell is to subsequently enter the cell cycle. Upon entry into S phase, cyclin E/CDK2 becomes active, and Skp2 levels increase.70 This results in a rapid suppression of overall p27 levels within the cell, the global activation of CDK2, and prepares the way for the firing of all preRCs within the cell. The entire genome is replicated, and p27 levels remain suppressed until the next anaphase. References 1. Sherr CJ. D-Type Cyclins. Trends Biochem Science 1995; 20:187-90. 2. Aktas H, Cai H, Cooper GM. Ras links growth factor signaling to the cell cycle machinery via regulation of cyclin D1 and the cdk inhibitor p27kip1. Mol Cell Biol 1997; 17:38507. 3. Sherr CJ. Cancer cell cycles. Science 1996; 274:1672-7. 4. Lukas J, et al. Cyclin E-induced S phase without activation of the pRb/E2F pathway. Genes Dev 1997; 11:1479-92. 5. Polyak K, et al. Cloning of p27Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell 1994; 78:59-66. 6. Reed SI. Control of the G1/S transition. Cancer Surveys 1997; 29:7-23. 7. Dulic V, Lees E, Reed SI. Association of human cyclin E with a periodic G1-S phase protein kinase. Science 1992; 257:1958-61. 8. Ohtsubo M, Theodoras AM, Schumacher J, Roberts JM, Pagano M. Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol Cell Biol 1995; 15:2612-24. 9. Stillman B. Initiation of eukaryotic DNA replication in vitro. Ann Rev Cell Biol 1989; 5:197-245. 10. Stillman B. Cell cycle control of DNA replication. Science 1996; 274:1659-64. 11. Mimura S, Takisawa H. Xenopus Cdc45-dependent loading of DNA polymerase alpha onto chromatin under the control of S-phase Cdk. EMBO J 1998; 17:5699-707. 12. Pacek M, Walter JC. A requirement for MCM7 and Cdc45 in chromosome unwinding during eukaryotic DNA replication. EMBO J 2004; 23, 3667-3676. 13. Wohlschlegel JA, Dhar SK, Prokhorova TA, Dutta A, Walter JC. Xenopus Mcm10 binds to origins of DNA replication after Mcm2-7 and stimulates origin binding of Cdc45. Mol Cell 2002; 9:233-40. 14. Mimura S, Masuda T, Matsui T, Takisawa H. Central role for cdc45 in establishing an initiation complex of DNA replication in Xenopus egg extracts. Genes to Cells 2000; 5:439-52. 15. Jares P, Donaldson A, Blow JJ. The Cdc7/Dbf4 protein kinase: target of the S phase checkpoint? EMBO Reports 1, 319-22 (2000). 626 16. Gu Y, Rosenblatt J, Morgan DO. Cell cycle regulation of CDK2 activity by phosphorylation of Thr160 and Tyr15. EMBO J 1992; 11:3995-4005. 17. Sebastian B, Kakizuka A, Hunter T. Cdc25M2 activation of cyclin-dependent kinases by dephosphorylation of threonine-14 and tyrosine-15. Proceedings of the National Academy of Sciences of the United States of America 1993; 90:3521-4. 18. Jinno S, et al. Cdc25A is a novel phosphatase functioning early in the cell cycle. EMBO J 1994; 13:1549-56. 19. Coulonval K, Bockstaele L, Paternot S, Roger PP. Phosphorylations of cyclin-dependent kinase 2 revisited using two-dimensional gel electrophoresis. Journal of Biological Chemistry 2003; 278:52052-60. 20. Devault A, et al. MAT1 ('menage a trois') a new RING finger protein subunit stabilizing cyclin H-cdk7 complexes in starfish and Xenopus CAK. EMBO J 1995; 14:5027-36. 21. Kaldis P, Solomon MJ. Analysis of CAK activities from human cells. Eur J Biochem 267, 4213-21 (2000). 22. Yee A, et al. Molecular cloning of CDK7-associated human MAT1, a cyclin-dependent kinase-activating kinase (CAK) assembly factor. Cancer Research 1995; 55:6058-62. 23. Yankulov KY, Bentley DL. Regulation of CDK7 substrate specificity by MAT1 and TFIIH. EMBO J 1997; 16:1638-46. 24. Lents NH, Keenan SM, Bellone C, Baldassare JJ. Stimulation of the Raf/MEK/ERK cascade is necessary and sufficient for activation and Thr-160 phosphorylation of a nucleartargeted CDK2. J Biol Chem 2002; 277:47469-75. 25. Cheng M, Sexl V, Sherr CJ, Roussel MF. Assembly of cyclin D-dependent kinase and titration of p27Kip1 regulated by mitogen-activated protein kinase kinase (MEK1). Proc Natl Acad Sci USA 1998; 95:1091-6. 26. Deshaies RJ. Phosphorylation and proteolysis: partners in the regulation of cell division in budding yeast.[erratum appears in Curr Opin Genet Dev 1997 Jun;7(3):424]. Curr Opin Gen Dev 1997; 7:7-16. 27. Schwob E, Bohm T, Mendenhall MD, Nasmyth K. The B-type cyclin kinase inhibitor p40SIC1 controls the G1 to S transition in S. cerevisiae. [erratum appears in Cell 1996 Jan 12;84(1):following 174]. Cell 1994; 79:233-44. 28. Verma R, et al. Phosphorylation of Sic1p by G1 Cdk required for its degradation and entry into S phase. Science 1997; 278:455-60. 29. Swanson C, Ross J, Jackson PK. Nuclear accumulation of cyclin E/Cdk2 triggers a concentration-dependent switch for the destruction of p27Xic1. Proc Natl Acad Sci USA 2000; 97:7796-801. 30. Hitomi M, Stacey WD. Cellular ras and cyclin D1 are required during different cell cycle periods in cycling NIH3T3 cells. Mol Cell Biol 1999; 19:4423-32. 31. Hitomi M, Stacey DW. Cyclin D1 production in cycling cells depends on ras in a cell-cyclespecific manner. Curr Biol 1999; 9:1075-84. 32. Stacey DW. Cyclin D1 serves as a cell cycle regulatory switch in actively proliferating cells. Curr Opin Cell Biol 2003; 15:158-63. 33. Sa G, Stacey DW. p27 expression is controlled by multiple, cell cycle specific pathways. Exp Cell Res 2004; 300:427-39. 34. Carrano AC, Eytan E, Hershko A, Pagano M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nature Cell Biol 1999; 1:193-9. 35. Bashir T, Dorrello NV, Amador V, Guardavaccaro D, Pagano M. Control of the SCF(Skp2Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature 2004; 428:190-3. 36. Wei W, et al. Degradation of the SCF component Skp2 in cell-cycle phase G1 by the anaphase-promoting complex. Nature 2004; 428:194-8. 37. Cho S, Kim HK, Jang SK. Polypriidine tract-binding protein enhances the internal robosomal entry site-dependent translation of p27Kip1 mRNA and modulates translation from G1 to S phase. Mol Cell Biol 25, 1283-1297 (2005). 38. Quelle DE, et al. Overexpression of mouse D-type cyclins accelerates G1 phase in rodent fibroblasts. 1993; 7. 39. Musgrove EA, Lee CS, Buckley MF, Sutherland RL. Cyclin D1 induction in breast cancer cells shortens G1 and is sufficient for cells arrested in G1 to complete the cell cycle. Proc Natl Acad Sci USA 1994; 91:8022-6. 40. Gladden AB, Diehl JA. The cyclin D1-dependent kinase associates with the pre-replication complex and modulates RB.MCM7 binding. J Biol Chem 2003; 278:9754-60. 41. Han EK, et al. Stable overexpression of cyclin D1 in a human mammary epithelial cell line prolongs the S-phase and inhibits growth. Oncogene 1995; 10:953-61. 42. Pagano M, Theodoras AM, Tam SW, Draetta GF, Chen J. Cyclin D1-mediated inhibition of repair and replicative DNA synthesis in human fibroblasts A 39 amino acid fragment of the cell cycle regulator p21 is sufficient to bind PCNA and partially inhibit DNA replication in vivo. Genes Dev 1994; 8:1627-39. 43. Fukami-Kobayashi J, Mitsui Y. Cyclin D1 inhibits cell proliferation through binding to PCNA and cdk2. Exp Cell Res 1999; 246:338-47. 44. Sherr CJ, Roberts JM. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev 1999; 13:1501-12. 45. Cheng M, et al. The p21(Cip1) and p27(Kip1) CDK 'inhibitors' are essential activators of cyclin D-dependent kinases in murine fibroblasts. EMBO J 1999; 18:1571-83. 46. Bagui TK, Mohapatra S, Haura E, Pledger WJ. P27Kip1 and p21Cip1 are not required for the formation of active D cyclin-cdk4 complexes. Mol Cell Biol 2003; 23:7285-90. 47. Han EK, et al. Increased expression of cyclin D1 in a murine mammary epithelial cell line induces p27kip1, inhibits growth, and enhances apoptosis. Cell Growth Diff 1996; 7:699-710. Cell Cycle 2005; Vol. 4 Issue 4 p27Kip1 and the Initiation of DNA Synthesis 48. Steuer R. Effects of stochasticity in models of the cell cycle: from quantized cycle times to noise-induced oscillations. J Theoretical Biol 2004; 228:293-301. 49. Raser JM, O'Shea EK. Control of Stochasticity in Eukaryotic Gene Expression. Science 2004; 304:1811-14. 50. You Z, Harvey K, Kong L, Newport J. Xic1 degradation in Xenopus egg extracts is coupled to initiation of DNA replication. Genes Dev 2002; 16:1182-94. 51. Furstenthal L, Swanson C, Kaiser BK, Eldridge AG, Jackson PK. Triggering ubiquitination of a CDK inhibitor at origins of DNA replication. Nat Cell Biol 2001; 3:715-22. 52. Ohtsubo M, Roberts JM. Cyclin-dependent regulation of G1 in mammalian fibroblasts. Science 1993; 259:1908-12. 53. Connell-Crowley L, Elledge SJ, Harper JW. G1 cyclin-dependent kinases are sufficient to initiate DNA synthesis in quiescent human fibroblasts. Curr Biol 1998; 8:65-8. 54. Resnitzky D, Reed SI. Different roles for cyclins D1 and E in regulation of the G1-to-S transition. Mol Cell Biol 1995; 15:3463-9. 55. Kozar K, et al. Mouse development and cell proliferation in the absence of D-cyclins. Cell 2004; 118:477-91. 56. Aleem E, Berthet C, Kaldis P. Cdk2 as a master of S phase entry: fact or fake? Cell Cycle 2004; 3:35-7. 57. Berthet C, Aleem E, Coppola V, Tessarollo L, Kaldis P. Cdk2 knockout mice are viable. Curr Biol 2003; 13:1775-85. 58. Ortega S, et al. Cyclin-dependent kinase 2 is essential for meiosis but not for mitotic cell division in mice. Nature Gen 2003; 35:25-31. 59. Tetsu O, McCormick F. Proliferation of cancer cells despite CDK2 inhibition. Cancer Cell 2003; 3:233-45. 60. Geng Y, et al. Resue of cyclin D1 deficiency by knockin cyclin E. Cell 1999; 97:767-77. 61. Parisi T, et al. Cyclins E1 and E2 are required for endoreplication in placental trophoblast giant cells. EMBO J 2003; 22:4794-803. 62. Spruck CH, Won KA, Reed SI. Deregulated cyclin E induces chromosome instability. Nature 1999; 401:297-300. 63. Zhou P, Jiang W, Weghorst CM, Weinstein IB. Overexpression of cyclin D1 enhances gene amplification. Cancer Res 1996; 56:36-9. 64. Tanaka S, Diffley JF. Deregulated G1-cyclin expression induces genomic instability by preventing efficient pre-RC formation. Genes Dev 2002; 16:2639-49. 65. Poon RYC, Toyoshima H, Hunter T. Redistribution of the CDK inhibitor p27 between different cyclin-CDK complexes in the mouse fibroblast cell cycle and in cells arrested with lovastatin or ultraviolet irradiation. Mol Biol Cell 1995; 6:1197-1213. 66. Malek NP, et al. A mouse knock-in model exposes sequential proteolytic pathways that regulate p27Kip1 in G1 and S phase.[erratum appears in Nature 2001 413:652]. Nature 413, 323-7 (2001). 67. Boehm M, et al. A growth factor-dependent nuclear kinase phosphorylates p27(Kip1) and regulates cell cycle progression. EMBO J 2002; 21:3390-401. 68. Chiariello M, Gomez E, Gutkind JS. Regulation of cyclin-dependent kinase (Cdk) 2 Thr160 phosphorylation and activity by mitogen-activated protein kinase in late G1 phase. Biochem J 2000; 349:869-76. 69. Kamura T, et al. Cytoplasmic ubiquitin ligase KPC regulates proteolysis of p27kip1 at G1 phase. Nature Cell Biol 2004; 6:1229-35. 70. Guo Y, et al. Phosphorylation of cyclin D1 at Thr 286 during S phase leads to proteasomal degradation and allows efficient DNA synthesis. Oncogene 2005; 24:1032-42. www.landesbioscience.com Cell Cycle 627