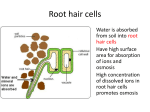
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Blackwell Science, LtdOxford, UKPCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 2002 26 Original Article A. H. de Boer & V. VolkovStructural and functional features of xylem Plant, Cell and Environment (2003) 26, 87–101 Logistics of water and salt transport through the plant: structure and functioning of the xylem A. H. DE BOER1 & V. VOLKOV2,3 1 Vrije Universiteit, Faculty of Earth and Life Sciences, Department of Developmental Genetics, Section Mol. Plant Physiol. & Biophysics, De Boelelaan 1087, 1081 HV Amsterdam, The Netherlands, 2K.A. Timiriazev Institute of Plant Physiology, Root Physiology Laboratory, Botanischeskaya 35, 127276 Moscow, Russia and 3Horticultural Production Chains Group, Wageningen University, Marijkeweg 22, 6709 PG Wageningen, The Netherlands ABSTRACT The xylem is a long-distance transport system that is unique to higher plants. It evolved into a very sophisticated plumbing system ensuring controlled loading/unloading of ions and water and their effective translocation to the required sinks. The focus of this overview will be the intrinsic interrelations between structural and functional features of the xylem. Taken together the xylem is designed to prevent cavitation (entry of air bubbles), induced by negative pressures under transpiration and to repair the cavitated vessels. Half-bordered pits between xylem parenchyma cells and xylem vessels are on the one hand the gates to the vessels but on the other hand a serious ‘bottle-neck’ for transport. Hence it becomes evident that special transport systems exist at the interface between the cells and vessels, which allow intensive fluxes of ions and water to and out of the xylem. The molecular identification and biophysical/ biochemical characterization of these transporters has just started. Paradigms for the sophisticated mechanism of controlled xylem transport under changing environmental conditions are SKOR, a Shaker-like channel involved in K+loading and SOS1, a Na+/H+ antiporter with a proposed dual function in Na+ transport. In view of the importance of plant water relations it is not surprising to find that water channels dominate the gate of access to xylem. Future studies will focus on the mechanism(s) that regulate water channels and ion transporters and on their physiological role in, for example, the repair of embolism. Clearly, progress in this specific field of research will greatly benefit from an integration of molecular and biophysical techniques aimed to understand ‘whole-plant’ behaviour under the everchanging environmental conditions in the daily life of all plants. Key-words: cavitation repair; ion channels; ion transport; ion transporters; potassium; sodium; water transport; xylem. Correspondence: Dr A. H. de Boer. Fax: + 31 20 4447155; e-mail: [email protected] © 2003 Blackwell Publishing Ltd Abbreviations: CNGC, cyclic nucleotide gated channels; PD, electric potential difference; SKOR, stelar potassium outward rectifying channel; TRP, trans-root potential. INTRODUCTION Like most multicellular organisms plants need structures that enable the mass flow of solvents (mainly water) and solutes such as ions, sugars, amino acids, hormones, etc. In analogy to the blood circulation system in animals, plants use tube-like structures called the xylem and phloem. Whereas the study of the blood circulation system is technically relatively easy, xylem and phloem transport pathways are notably difficult to study due to their location deep inside the plant tissue, the presence of reinforced cell walls and the large positive or negative pressures inside the tubes. Despite, or maybe challenged by these hurdles, a large body of research has been devoted to unravelling the mechanisms that sustain the functioning of the plant long-distance transport systems. For recent insights in phloem transport we refer to reviews by Patrick et al. (2001) and Oparka & Cruz (2000). Here we try to link the structure–function relationship of the dead xylem conduits and living adjacent parenchyma cells with a focus on the mechanisms that facilitate and control the flow of ions and water into (loading) and out of the xylem (unloading). Although it was hypothesized in the early days of xylem research that, for example, xylem loading in the root was a passive process due to the lack of oxygen in the core of root tissue (Crafts & Broyer 1938), it is clear now that we are dealing with sophisticated regulation mechanisms aimed to sustain growth and development in highly dynamic environments. Since structure and function are intimately interwoven, the structure of the xylem transport system will be discussed first. Thereafter, current knowledge about a number of essential biophysical parameters namely the electrical, pH and pressure gradients between the vessels and surrounding symplast/apoplast will be summarized. The molecular mechanisms of potassium, sodium, and water transport will be discussed in more detail at the end. 87 88 A. H. de Boer & V. Volkov HOW STRUCTURAL FEATURES OF THE XYLEM MEET THE DEMANDS OF SHORT- AND LONGDISTANCE TRANSPORT The main function of the xylem is to transport large quantities of water and solutes. This requires a continuous system of interconnected tubes with a relatively low resistance to the flow of water. As the driving force for mass flow is a pressure gradient (positive with respect to the atmospheric pressure as during root pressure, or negative during transpiration) the walls of the vessel or tracheary elements must be reinforced. On the one hand leakage of water and solutes out of the tubes must be minimized (so the walls must be as leak-proof as possible), but on the other hand specialized structures in the walls must enable rapid exchange of water and solutes between the vessel interior and surrounding cells. Another feature that is vital for survival of the xylem is to prevent and repair the interruption of the continuous water columns by air bubbles (embolism). a Low resistance allows high water flows The principal conducting cells of the xylem are the tracheary elements of which there are two types: the tracheids and the vessel elements. Both tracheary elements develop normally from proliferating cells from the procambium and vascular cambium. When fully differentiated the end walls of the vessel elements are perforated or removed (what is left is called the perforation plate) and the cells undergo programmed cell death (Roberts & McCann 2000). Thus, files of open interconnected cells are formed. Tracheids also lack protoplasts at maturity, but in contrast to vessels the end walls are not open but contain pits, which are areas lacking the secondary wall. The so-called pit membrane, a modified thin primary cell wall consisting of a dense network of hydrophilic cellulose polymers, is water permeable (Holbrook & Zwieniecki 1999). Water flowing through vessels sometimes has to pass pit membranes as well, as vessels are not continuous over the entire length of the plant and are interconnected through pits or perforation plates (Fig. 1). The principal water-conducting cells in angiosperms are tracheids, whereas vessels are found both in angiosperms and gymnosperms. Important determinants of the hydraulic resistance are the diameter of the vessels and the pit membranes or perforation plates. In xylem vessels, water is assumed to flow according to Poiseuille’s law with a hydraulic conductivity proportional to the fourth power of vessel’s radius. Thus, a small change in the radius will give a large change in resistance. The radius of a vessel may also be related to a function in solute loading. For example, in the first 10–20 cm of roots of monocots only vessels having a small diameter are functional in transport. The reason may be that narrow vessels with a high surface-to-volume ratio are more suitable for controlling the composition of the xylem sap than wide vessels. The large late metaxylem vessels in the apical 10–20 cm of a maize root do not conduct water (St. Aubin, Canny & McCully 1986), because they mature just in the pm b Figure 1. Bordered pit pairs interconnect xylem vessels and halfbordered pits connect the vessels to xylem parenchyma cells. (a) Cryo-planed transverse section through the secondary xylem of a chrysanthemum stem. (b) Bordered pit pair in high magnification (length of pit membrane is 3·7 µm). V, vessel; bp, bordered pit; hp, half-bordered pit; pc, parenchyma cell; lf, libriform fibre; sp, simple pit; pm, pit membrane (Micrograph kindly provided by Dr J. Nijsse, Wageningen University). upper zones of the root. In these upper zones, the wide vessels collect the sap and function as simple conductive elements, whereas narrow xylem vessels function not only as conductive elements but also as xylem sap-controlling elements. The prevailing view that tracheary elements have either a constant or a zero (after embolism) hydraulic conductance has recently been challenged (Van Ieperen, Van Meeteren & van Gelder 2000; Zwieniecki et al. 2001) although the observation was already made by Zimmermann (1978) in a nice example of serendipity. Observing a decreasing flow rate of distilled water through a piece of stem (a problem already described by Huber & Merz 1958), Zimmerman hypothesized that tiny gas bubbles obstructed pit pores. Therefore he replaced the distilled water with tap water (assuming that tap water contained more bubbles than distilled water), but contrary to expectation the flow rate instantaneously increased. He discovered that this effect was due to the presence of ions in the tap water and © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 89 concluded that ‘the phenomenon might be based upon swelling or shrinking of the vessel-to-vessel pit membranes’. Van Ieperen et al. (2000) concluded that the presence of small amounts of cations (even the effect of 10 µM KCl was significant) and not the osmotic strength influenced the conductance. However, their conclusion that swelling and shrinking of vessel-to-vessel pit membranes played no role in this phenomenon was contradicted by Zwieniecki et al. (2001). The latter authors reached the same conclusion as formulated by Zimmermann (1978) namely that the observed change in hydraulic resistance when changing the ionic composition of the xylem sap is due to the pit membranes connecting one vessel to the other. In these ‘membranes’ water flows through microchannels of the modified primary cell wall, made up by cellulose microfibrils, hemicelluloses and pectins. It was suggested that the size of the microchannels increases when the pectin matrix (D-galacturonic acid being a main component) shrinks in response to the binding of ions to the negatively charged matrix. If charge compensation is the mechanism that determines the flow resistance, then it is surprising that no significant effect of pH within the natural pH range of xylem sap (5·8–8·0) was observed (Zwieniecki et al. 2001), although pH affected swelling and shrinking of isolated cell walls (Meychik & Yermakov 2001). Relevant of course is the question whether these ion-mediated changes in hydraulic resistance do have a physiological role. The K+ concentrations having the strongest effect (0– 20 mM; Zwieniecki et al. 2001), are within the range measured in transpiring intact plants (Herdel et al. 2001). However, it remains to be seen whether these K+ variations in a constant background of, for example, divalent cations still have an effect on the hydraulic properties. Reinforcement and waterproofing to withstand negative pressures and retain water Tracheary elements not only have to withstand the compressive forces from surrounding cells, but especially the large negative pressures that develop in actively transpiring plants (see below). Therefore, secondary walls composed of cellulose and xylan are deposited on top of the primary cell wall (Fukuda 1996). Early-formed protoxylem elements of the primary xylem have ringlike (annular) or spiral (helical) thickenings that still allow a certain degree of cell elongation. In the late-formed metaxylem elements and elements of the secondary xylem the secondary cell walls cover the entire primary walls, except at pit membranes shared with adjoining tracheids/vessels or xylem parenchyma cells and contact cells. Subsequently, the secondary cell walls are lignified: a network is formed that strengthens and waterproofs the wall by the polymerization of aromatic compounds called monolignols (McCann 1997). Not surprisingly, inhibitors of lignification result in a plant phenotype with collapsed vessels (Smart & Amrhein 1985). Primary cell wall polymers are also important for the subsequent lignification and strengthening of a vessel, and mutants in cellulose deposition in the cell wall with col- lapsed xylem vessels were isolated as well (Turner & Somerville 1997). In order to prevent ‘leakage’ of water and solutes from the vessels it seems logical that the lignified secondary walls are relatively impermeable to these compounds. One of the few studies on xylem cell wall permeability is the one by Wisniewski, Ashwort & Schaffer (1987) who studied deep supercooling in woody plants and used lanthanum nitrate as an apoplastic tracer. Lanthanum penetrates capillaries as small as 2 nm, so a cell wall without lanthanum staining is relatively impermeable. They found that the primary cell wall of cortical tissues was very permeable to the lanthanum solution, but primary (except parts associated with the pit membranes) and secondary walls of vessel elements were impermeable. Interestingly, the pit membranes composed of the middle lamella, primary walls of the adjoining cells and the protective layer (in xylem parenchyma cells and contact cells) were highly permeable. These results are in line with the conclusion that after occlusion of pit membranes, vessel elements can assume a closed-cell structure and may have a permeability approaching zero (Siau 1984). These permeability properties of the xylem walls are relevant to the question whether water and solutes can directly enter the vessels from the stelar apoplast. If not, then they have to be taken up by stelar cells into the symplast and loaded into the vessel lumen after passing the membrane of a xylem parenchyma cell underlying a pit membrane. For example, the stelar potassium outward rectifying channel (SKOR; activated at depolarizing voltages) is thought to have a function in loading K+ into the xylem (Wegner & Raschke 1994; Wegner & de Boer 1997a; Gaymard et al. 1998). Besides expression in xylem parenchyma cells, clear GUS-staining of root pericycle cells was observed in plants expressing the GUS reporter gene under the control of the SKOR promoter region (Gaymard et al. 1998). Note also that pericycle cells of barley roots stained strongly for the plasma membrane ATPase (Samuels, Fernando & Glass 1992). The pericycle cells do not directly border a xylem vessel. Therefore, the question is how K+ ions released from the pericycle cells into the stelar apoplast do reach the xylem lumen if the xylem vessel walls are as impermeable to ions and water as shown by Wisniewski et al. (1987). Pit structures facilitate water-/solute (un)loading and embolism repair Pits constitute the gates between vessels and from vessels to adjacent xylem parenchyma cells (Fig. 1). The total area of pit fields, their shape and pattern of lignification are of large variation between species. For example, maize pit fields constitute about 15% of the vessel area, with pit membranes having a diameter of about 2 µm with a hydraulic conductivity about 10 times over that of the vessel wall (Peterson & Steudle 1993). Typically, between vessels bordered pits are found that are characterized by overarching walls that form a bowl-shaped chamber (Fig. 1b). It has long been known that these pits are vital structural ele- © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 90 A. H. de Boer & V. Volkov ments in the plant’s defence against cavitation (rupture of water columns) and subsequent embolism (filling of vessels and tracheids with air) (Zimmermann 1983; Tyree & Sperry 1989). This is because the modified primary cell wall that constitutes the pit membrane is permeable to water, but does not allow the passage of even the tiniest air bubbles. For this reason tracheids, wherein all elements are linked through pits are thought to be safer for plants than vessels although the latter elements are more efficient conductors of water. In some monocots, such as rye, in the root–shoot junction tracheids separate the vessels in the root from those in the shoot to protect the vessels of the roots from embolism originating in the shoot (Aloni & Griffith 1992). However, pits are not only important structural elements that prevent spreading of air bubbles through the waterconducting elements of plants. They are instrumental in repairing the ‘broken water pipes’ as well (Tyree et al. 1999; Holbrook & Zwieniecki 1999; Zwieniecki & Holbrook 2000). Repair means removal of the air bubble; how can this be achieved? As we have seen above, an air bubble cannot be simply ‘flushed’ out of a conduit. The existing paradigm suggests that an air bubble is removed through gas dissolution in the surrounding water (see Table 1). An important structural demand is that in order to contain this pressurization the perimeter of the embolized vessels must be sealed. As we have seen above, the secondary wall meets this requirement since the thickness and lignification make it waterproof. However, what about the pits in which the water permeability of the pit membrane is high (see below)? Let us first consider the hypothesis that a plant has to generate positive (above atmospheric) pressures in the vessels. The best understood mechanism that generates positive pressures is root pressure; namely pressure that is generated in the root during the night or in the shoot in early spring when transpiration is low through osmotic water flow into the vessels. There is good experimental evidence that this mechanism operates in a range of Table 1. Threshold xylem pressure Removal of air from an embolized vessel must occur by dissolution of the gas in the surrounding water. The solubility of a gas in water is proportional to the partial pressure of the gas species adjacent to the water (Henry’s law). Water in plants is saturated with air at atmospheric pressure and therefore the pressure of the gas in the bubble must be above atmospheric for the gas to dissolve in the surrounding water. What determines the pressure of the gas in the bubble (Pg)? (1) The pressure inside the xylem element, Px; and (2) the pressure inside the gas bubble that develops due to the surface tension at the air–water interface, 2τ/r, where r is the radius of the curvature of the air–water interface and τ is the surface tension of the water in the vessel. Thus the pressure of the gas in the bubble equals: Pg = Px + 2τ/r. Therefore when the xylem pressure is greater than −2τ/r (the threshold xylem pressure), the gas in an embolism will exceed atmospheric pressure and start to dissolve. For a narrow vessel of 10 µm diameter this threshold pressure is around −30 kPa (−0·3 bar). See Tyree et al. (1999) for further details. angiosperms and gymnosperms under certain conditions (Tyree et al. 1986; Sperry et al. 1987; Tyree & Yang 1992; Cochard, Ewers & Tyree 1994). However, not all plants seem to develop root pressure (20 out of 109 species studied had no root pressure, Fisher et al. 1997) and it must be kept in mind that root pressure is dissipated by gravity at a rate of 10 kPa per m height. Therefore, tall trees are unlikely to develop positive pressures in the shoot as a consequence of pressure built-up in the root. So, how do plants dissolve embolism without root pressure? This question is even more relevant in the light of recent studies reporting embolism repair under conditions when the threshold xylem pressure (Px) (see Table 1) is much less than −2τ/r (where τ is the surface tension of water in the vessel and r is the radius of curvature of the air–water interface) (Salleo et al. 1996; Tyree et al. 1999). The only possible explanation for this repair mechanism seems to be that locally (i.e. in the embolized vessel or tracheid element) the xylem pressure is higher than −2τ/r. There are a number of intriguing problems with this model: 1 The data suggest that this mechanism of embolism removal is concurrent with transpiration (Salleo et al. 1996; Canny 1997b; Tyree et al. 1999). If so, how can xylem pressure increase in the embolized element while the adjacent vessels are still under tension? 2 What is the mechanism that generates locally a xylem pressure > −2τ/r? The first problem was distinguished by Tyree et al. (1999) and Holbrook & Zwieniecki (1999) came up with a hypothesis as to why in two adjacent vessels (one embolized, the other filled and under tension) connected by pits, a pressure difference can be maintained until the embolized vessels are water filled again (Holbrook & Zwieniecki 1999; Zwieniecki & Holbrook 2000). Without going into much detail, the idea is that the structure of the bordered pits ensures that the embolized element remains hydraulically isolated from the neighbouring vessels under tension until the moment that all air has been dissolved. Zwieniecki & Holbrook (2000) showed that the contact angle of water and the angle of the bordered pit chamber walls are such that a convex gas–water interface forms in the pit chamber. The surface tension of this convex curvature opposes the hydrostatic pressure within the filling lumen as long as the pressure in the filling element does not exceed the force due to the surface tension. Clearly, this model deserves further experimental testing. The second problem seems rather easy to solve. Namely, xylem conduits are generally in contact with numerous living cells, the xylem parenchyma cells and contact cells (Zimmermann 1983; Lachaud & Maurousset 1996). These cells could be triggered by embolism to load osmotically active ions such as K+ and Cl– across the pit membrane into the vessel lumen. Such a mechanism was proposed for the first time by Grace (1993) who studied embolism repair in tracheids from Pinus sylvestris. Water would then be driven by osmosis into the vessel lumen. This idea is consistent with the observation that convex drops protrude through © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 91 Figure 2. Part of an embolized late metaxylem vessel in maize root. Note the pits between the vessel and bordering xylem parchyma cells. Arrows indicate droplets of liquid exuding through pits into the lumen (× 1580) [Reproduced from McCully et al. (1998), with permission of Blackwell Publishing, Oxford, UK]. the pits into the lumen of embolized vessels (see Fig. 2) from sunflower petioles (Canny 1997b), roots of maize (McCully, Huang & Ling 1998) and Laurel nobilis branches (Tyree et al. 1999). However, thus far there is one major problem: attempts to measure high concentrations of salts in the water that enters an embolized vessel have failed (Canny 1997b; McCully et al. 1998; McCully 1999; Tyree et al. 1999). All of the methods used to estimate the xylem sap ion concentration, rely on the technique of X-ray microanalysis except one (Tyree et al. 1999). In the latter study ‘native’ xylem sap from well-watered and cavitated twigs was collected by perfusion. Treatments wherein cavitation recovery was high also showed a strong increase (up to four-fold) in K+ concentration. This positive correlation does suggest that the increase in ion concentration has something to do with the refilling of embolized elements. If so, then the increase in concentration of solutes in the refilling vessels may be much higher since the sap recovered from the perfused stems will be diluted by water from the full vessels. So, the discussion on the mechanism of refilling embolized vessels seems to hinge on the question whether droplets entering an embolized element through the pits do contain high enough levels of solutes. If X-ray microanalysis gives a reliable estimate of the xylem sap ion concentration, then we need a new paradigm of how positive xylem pressures build-up in embolized vessels and how root pressure in general is generated (Enns, Canny & McCully 2000). Clearly, there is a need for reliable methods for sampling and analysis of ‘native’ xylem sap. In the literature many methods have been described, as summarized by Schurr (1998), none of which is without its own specific problems. The methods range from the collection of xylem sap under root pressure to such an unusual method as using xylem feeding Philaenus spumarius (spittlebugs) to get the sap sample (Watson, Pritchard & Malone 2001). As a general rule, it is important to define the question to be addressed and then define the requirements a sampling method must meet. Ideally, the method of analysis does not disturb the plant in general and the conditions of transpiration in particular. Non-invasive techniques such as nuclear magnetic imaging (NMRi), nuclear magnetic resonance (NMR) spectrometry and magnetic resonance imaging (MRI) do in principle meet these requirements (Scheenen et al. 2000; Peuke et al. 2001; Holbrook et al. 2001). Spatial and temporal resolution was initially an important disadvantage of these methods, but recent methods have decreased measurement time drastically (Rokitta et al. 1999; Scheenen et al. 2000; Peuke et al. 2001). If the refilling of embolized vessels is not driven osmotically by salts loaded into the vessel lumen by the surrounding xylem parenchyma cells, then maybe reverse osmosis is a feasible mechanism (McCully 1999; Tyree et al. 1999) as originally proposed by Canny (1997a, b). Reverse osmosis could occur if cells surrounding the xylem decrease their osmotic potential (through ion uptake or conversion of starch to sugars), built up a higher turgor pressure, pressurize xylem parenchyma cells that then squeeze water (without ions) into the embolized vessel element. An auxininduced loading of solutes in the phloem may be part of this mechanism (Salleo et al. 1996; Tyree et al. 1999). However, Tyree et al. (1999) have pointed out the problems that come with this mechanism. One important problem to solve is how to prevent water from flowing in all directions from the ‘squeezed’ parenchyma cells. In other words, how to generate a direction in the flow of water (a problem solved in the ‘osmosis’ hypothesis by specific loading of ions into the embolized element across the pit membrane). One possibility is that water channels in the patch of membrane underlying the pit membrane are activated, thus creating a path of low resistance to waterflow into the vessel element (see section on water channels). Vessel-associated cells are equipped for a task in (un)loading Living vessel-associated cells, the xylem parenchyma cells and contact cells, are the cells that have direct access to the © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 92 A. H. de Boer & V. Volkov vessel lumen. Therefore, they play a key role in the loading and unloading of the xylem. In line with the energy demand of these (un)loading processes, structural features of cells indicate high metabolic activities that could fuel massive transmembrane transport. These features are a dense cytoplasm, well-developed endoplasmic reticulum, numerous ribosomes, mitochondria and peroxisomes (Läuchli et al. 1974). The specialized transport function of these cells is evidenced by the development of cell wall invaginations in the area underlying the pit membrane bordering the vessels (Kramer et al. 1977; Yeo et al. 1977; Kuo et al. 1980; Mueller & Beckman 1984). This folding pattern increases the membrane surface area and is characteristic for transfer cells (Pate & Gunning 1972; Gunning 1977) and the development is stimulated by salt stress (Kramer et al. 1977; Yeo et al. 1977). Relatively little is known thus far about the distribution of ion transporters in the membrane of vesselassociated cells. Sauter, Iten & Zimmermann (1973) observed high ATP hydrolysing activity at the large pits of contact cells facing the vessels and suggested a role for this enzymatic activity in sugar loading of the xylem. This activity was only observed during starch mobilization in the rays cells in spring or when starch mobilization was artifically induced by defoliation. Xylem parenchyma cells of barley and oat roots were strongly labelled by antibodies against the plasma membrane ATPase, but the resolution was insufficient to discern differences in subcellular distribution of the protein (i.e. patches on the vessel-facing side) (Parets-Soler, Pardo & Serrano 1990; Samuels et al. 1992). These results are in line with the localization of ATPase activity in barley xylem parenchyma cells as determined with a cytochemical lead precipitation method (WinterSluiter, Läuchli & Kramer 1977). The external cytoplasmic layer of xylem parenchyma cells has specific features at the point of contact pits (pits between xylem parenchyma cells and xylem vessels). There, the network of actin filaments contains fewer cortical microtubules in comparison with other parts of the cytoplasmic layer where an intensive cortical microtubule cytoskeleton is present (Chaffey, Barlow & Barnett 2000; Chaffey & Barlow 2001). The function of these structural changes of the cortical cell layer may be two-fold: to prevent the deposition of a thick secondary wall, a process which depends on cortical microtubules, and the regulation of ion and water channel activity. Actin involvement in ion channel regulation was shown for guard cells (Hwang et al. 1997). THE PROTON MOTIVE FORCE AT THE XYLEM/ SYMPLAST INTERFACE: DRIVING FORCE FOR XYLEM (UN)LOADING The energy necessary to drive transport of ions, amino acids, sugars, etc. across the membrane of vessel-associated cells into or out of the vessel lumen, is derived from the electrical gradient across this membrane, the pH gradient or both. Therefore, it is important to know the value of these two parameters and the dynamic regulation thereof. Xylem electrical potential The initial hypothesis postulated by Crafts & Broyer (1938) that primary active transport (e.g. an ATP driven H+ATPase) at the symplast/vessel boundary was precluded due to lack of oxygen, proved not to be correct (Okamoto, Katou & Ichino 1979; De Boer et al. 1983; Clarkson, Williams & Hanson 1984). In roots of Plantago and hypocotyls of Vigna unguiculata the electric potential difference (PD) between vessel and parenchyma cells contains an electrogenic component, which in both roots and hypocotyls can amount to 70 mV. The primary H+-ATPase is thought to be responsible for this electrogenic component because it disappears in the presence of inhibitors of the oxidative phosphorylation (anoxia and azide) and increases with auxin and fusicoccin, the activator of the plasma membrane H+ATPase (De Boer et al. 1985; De Boer 1997). It must be kept in mind that the electrical potential of the cortical and xylem (stelar) apoplast are not necessarily the same. An insulating layer formed by the ring of endodermis cells with their Casparian strips, can maintain a PD between the two apoplasts. Because the cortical, endodermal and stelar cells are connected through plasmodesmata (forming the symplast) the apoplast PD is in fact the difference of two membrane potentials in series: (PDsymplast/cortical apoplast) minus (PDsymplast/xylem apoplast) (Okamoto et al. 1979; De Boer et al. 1983; De Boer et al. 1985). This PD is also called trans-root potential (TRP), but the same principle holds for stems and other tissues. All measurements mentioned above were carried out on excised plants or perfused root and stem segments. This approach can be criticized because in excised roots the turgor and intracellular osmotic pressure gradients collapse (Rygol et al. 1993). Therefore, measurements on intact plants are necessary. Dunlop (1982) succeeded for the first time to measure directly with a microelectrode in the vessels of intact Trifolium repens plants. The electrical potential of the vessels was 90 mV negative with respect to the cortical apoplast. Because xylem parenchyma cells had a membrane potential of −168 mV it can be calculated that the potential difference across the symplast/xylem boundary was around 80 mV (xylem positive). These values are close to the values measured in excised Plantago roots where the electrogenic component of the symplast/xylem PD was zero (De Boer et al. 1983). Recently, a new method was introduced, using a modified pressure probe, enabling the simultaneous measurement of the trans-root potential and the xylem pressure in intact transpiring plants (Wegner & Zimmermann 1998). One of the conclusions of these measurements was that the roots of intact transpiring plants have electrical properties similar to those of excised roots (Wegner et al. 1999). These measurements also demonstrated that the regulation of the trans-root electrical potential is a dynamic process. Namely, a switch to a higher light intensity induced strong oscillations in TRP that preceded oscillations in the negative xylem pressure. Interestingly, the depolarization of TRP correlated with the most negative xylem pressure. Since the membrane potential of the © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 93 cortical cells showed no oscillations, the oscillations in TRP must be the result of a change in ion transport activity across the symplast/xylem boundary or as suggested by Wegner et al. (1999) by a change in streaming potential due to variations in the transpiration flow. However, thus far it is not clear whether streaming potentials contribute significantly to the potential differences within a xylem vessel or between the xylem and cortical apoplast. Although we now know that xylem transport processes are not simply passive, the early suggestion that the supply of oxygen to the xylem might be a limiting factor (Crafts & Broyer 1938; De Boer & Prins 1984) deserves renewed attention in the light of recent analysis of the oxygen supply to sapwood using a novel optical method for oxygen detection (Gansert, Burgdorf & Lösch 2001). Xylem sap pH The pH of the xylem sap is relevant for (un)loading processes for two reasons: (i) as driving force for antiport and symport processes (see below) and (ii) as regulator of ion transporters or signalling molecules. There are no measurements of the xylem sap pH in intact plants as far as we are aware. Measurements on xylem sap collected from root exudates or by means of centrifugation agree on a slightly acidic pH in the range 5·5–6·5 (Schurr & Schulze 1995; Wilkinson & Davies 1997; López-Millán et al. 2000). Somehow the xylem has a high capacity to maintain its pH to a set-value (Clarkson et al. 1984; Senden et al. 1992). However, in response to changes in environmental conditions the xylem sap pH does fluctuate. Under drought stress the xylem sap pH from Commelina communis increased from 6·1 to 6·7 (Wilkinson & Davies 1997) and iron stress conditions reduced the pH from sugar beet xylem sap from 6·0 to 5·7 (López-Millán et al. 2000). What is the mechanism that on the one hand buffers and on the other hand changes the xylem pH? Buffering is certainly a consequence of the cation exchange capacity of the xylem cell walls due to the presence of negatively charged groups (Ferguson & Bollard 1976; Senden et al. 1992) and the presence of organic acids such as succinate and malate (Schurr & Schulze 1995). But the physiological changes may be due to regulation of the activity of the H+-ATPase and/or H+-exchange systems in the plasma membrane of vessel associated cells. If for example the H+-ATPase activator fusicoccin is perfused through xylem vessels the pH of the emerging sap decreases within minutes after application, reaching values as low as 4·2 (Fig. 3). This effect is explained by the uptake of fusicoccin by xylem parenchyma cells around the vessels. Fusicoccin then activates the plasma membrane H+ATPase (De Boer 1997), which results in a strong efflux of protons into the xylem sap. Further evidence for the involvement of metabolically active cells is the observation that the pH rapidly increases when roots are subjected to anoxia (De Boer & Prins 1985; Lacan & Durand 1996). Several roles for changes in xylem sap pH have been proposed. Thus, the increase in pH initiated by drought stress, is thought to increase the efficiency of abscisic acid in stim- Figure 3. Effect of fusicoccin, activator of the plasma membrane H+-ATPase, on the ion concentration and ion fluxes into/out of the xylem as measured by means of a xylem perfusion technique. (a) Perfusion of taproots from Plantago maritima and (b) of nodal roots from barley. In both roots, fusicoccin (10 µM) strongly acidifies the xylem sap, reduces the flow of K+ into the xylem from the surrounding cells and stimulates the release of Na+ into the xylem stream. [(a), De Boer unpubl. res.; (b), Volkov and De Boer unpubl. res.]. ulating the closure of stomata (Wilkinson & Davies 1997). Moreover, K+ loading into the root xylem may be a pHsensitive process through modulation of the activity of SKOR, the outward rectifying K+ channel involved in K+ release into the xylem (Lacombe et al. 2000 and see below). MOLECULAR MECHANISM OF SOLUTE AND WATER (UN)LOADING Potassium Xylem loading of K+ is regulated separately from K+ uptake from the external solution (Cram & Pitman 1972; Engels & Marschner 1992). Studies at the cellular and molecular level in recent years have shown two principal types of K+ channels in the root (Gaymard et al. 1998; Spalding et al. 1999): AKT1 for K+ uptake from the medium and SKOR for K+ loading into the xylem. The in planta properties of SKOR [formerly KORC in barley (Wegner & De Boer 1997a)] were first characterized through patch-clamp studies on xylem parenchyma protoplasts isolated from barley and maize roots (Wegner & Raschke 1994; Roberts & Tester © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 94 A. H. de Boer & V. Volkov 1995; Wegner & De Boer 1997a; Roberts 1998). These outward rectifying channels activate in a time-dependent manner at membrane potentials that shift with and remain slightly positive of the reversal potential of K+ (EK). They are selective for K+ but do facilitate the passage of Ca++ as well. They cannot be responsible for Na+ loading of the xylem as they are virtually impermeable to Na+. Under physiological conditions a depolarization of the membrane potential to values positive of EK will result in a K+ efflux and an influx of Ca++. Therefore, a non-functional SKOR may result in a reduction of the amount of K+ and an increase in the amount of Ca++ in the shoot (see below). In addition to its sensitivity to membrane voltage and apoplastic K+ concentration, the activity of SKOR is regulated by cytosolic Ca++, the stress hormone abscisic acid (ABA) and pH. A rise in the cytosolic Ca++ concentration from 150 nM to 5 µM reduced the frequency at which SKOR was recorded in barley xylem parenchyma cells (whole-cell configuration) from 90 to 10% (Wegner & De Boer 1997a). As this effect was absent when inside-out patches were used, the involvement of a cytosolic intermediate was suggested. ABA has a dual effect on SKOR activity: a shortterm effect measurable within minutes after addition of ABA and a long-term effect (Roberts 1998; Gaymard et al. 1998). The short-term effect was observed when ABA was added to protoplasts during a patch-clamp experiment: within minutes it reduced the outward current in 55% of the cells; the inward current was unaffected. Drought, or overnight pre-treatment of intact plants with ABA also resulted in a strong reduction of the outward K+ current. In line with earlier experiments (Cram & Pitman 1972; Pitman 1977) ABA pre-treatment had no effect on in- and outward-currents in cortical root cells (Roberts 1998). It remains to be shown if Ca++ plays a role as second messenger in the xylem parenchyma ABA signalling pathway, as it does in guard cells (Allen et al. 2000). An indication maybe that both Ca++ and ABA affect SKOR in a similar fashion. The gene encoding the channel responsible for K+ loading in the xylem, called SKOR, belongs to the Shaker superfamily (Gaymard et al. 1998). SKOR expressed in Xenopus oocytes, exhibited most if not all of the characteristics of xylem parenchyma K+ channel characterized in barley and maize: sigmoidal activation, shift in activation potential with EK, sensitivity to blockers, Ca++ permeability and increased magnitude of the outward current with increasing external K+ concentration. Northern blot analysis showed that SKOR is expressed exclusively in roots and there the expression is limited to pericycle cells and cells surrounding the xylem vessels; hence the name stelar K+ outward rectifier. Treatment of intact plants with the stress hormone abscisic acid resulted in a rapid decline in SKOR mRNA abundance (Gaymard et al. 1998), thus shedding light on ABA inhibition of xylem K+ loading in intact barley plants (Cram & Pitman 1972) and the activity of K+-outward rectifying channels in maize stelar protoplasts as measured with the patch-clamp technique (Roberts 1998). The conclusion that xylem loading of K+ is regulated separately from K+ uptake from the external solution (Cram & Pitman 1972; Pitman & Wellfare 1978; Engels & Marschner 1992) is underlined by the fact that the K+ channel (activity and expression) responsible for K+ uptake into the root symplast, AKT1 is not affected by ABA (Roberts 1998; Gaymard et al. 1998). Roberts (1998) also observed a short-term effect of ABA upon addition to the bath medium during a patch-clamp measurement: within 5 min after ABA addition the outward K+ current in maize stelar protoplasts declined by about 50%. This effect might be mediated by changes in cytosolic pH since a decrease in cytosolic pH from 7·4 to 7·2 induced a strong (80%) voltage-independent decrease of the macroscopic SKOR current in a heterologous expression system (oocytes) (Lacombe et al. 2000). However, both in dark-grown coleoptiles and in barley aleurone protoplasts ABA induced an alkalinization of 0·1–0·2 units (Gehring, Irving & Parish 1990; Van der Veen, Heimovaraa-Dijkstra & Wang 1992). If the same were true for xylem parenchyma cells, then an ABA-induced alkalinization of the cytoplasm would increase the outward K+ current. It remains necessary to determine the pH sensitivity of SKOR in planta as well as the effect of ABA on the pH of xylem parenchyma cells. The high level of expression of SKOR in the pericycle cells raises some questions. As these cells are not in direct contact with the xylem vessels, K+ ions released through SKOR will end up in the stelar apoplast. If the ion permeability of the secondary walls of the xylem vessels is low (see above) then loading of these ions into the xylem might be preceded by uptake into the xylem parenchyma cells followed by release into the vessels. This could explain the relatively large number of inward rectifying K+ channels (i.e. channels that activate at hyperpolarizing voltages) in barley xylem parenchyma cells, two of which are G-protein regulated (Wegner & de Boer 1997b). The explanation for such a complex mechanism of loading is not immediately obvious, but in some plants sugar loading into the phloem also includes an apoplast unloading step followed by uptake into the companion cells and symplast loading into the sieve elements (Van Bel et al. 1992). These xylem parenchyma K+ inward rectifying channels may also have a function in the apoplast resorption of K+ ions released by the phloem in the root stelar apoplast. Retranslocation of K+ from the shoot to the root in the phloem is a well-known phenomenon and it has been speculated that the amount of K+ returning from the shoot acts as a signal for root K+ uptake from the soil (White 1997; Jeschke & Hartung 2000). Xylem perfusion experiments, using tetraethylammonium as blocker of SKOR, showed the in vivo function of SKOR in loading the xylem with K+ (Wegner & De Boer 1997a). The phenotype of the skor-1 knockout mutant (Gaymard et al. 1998) indeed showed a reduced transport (50%) of K+ to the shoot of skor-1. Electrophysiological characterization of the selectivity properties of SKOR in barley yielded a PCa++/PK+ ratio of 0·68 (Wegner & De Boer 1997a). This outcome suggested a role for SKOR in Ca++ © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 95 XP V V XP Figure 4. Conspicuous wall ingrowths of transfer cells lining the xylem of the cotelydonary traces of a seedling of Jacob’s ladder (Polemonium caeruleum) (× 600). V, xylem vessel; XP, xylem parenchyma cell. [Reproduced from Gunning et al. (1970), with permission of Springer-Verlag Austria]. root (a) 700 600 shoot (b) 0 mM NaCl 1 mM NaCl 25 mM NaCl 500 –1 Na (me.kg d.m.) 400 300 + unloading of the xylem in roots, which was confirmed by the higher Ca++ content of the skor-1 leaves (Gaymard et al. 1998). On the way up through the stem into the leaves K+ ions will be taken up into the symplast by cells in contact with the vessels and tracheids. The uptake of K+ in the elongation zone of the hypocotyl of Vigna unguiculata is stimulated by the growth-promoting hormone auxin and fusicoccin (De Boer et al. 1985). The actual mechanism (ion channel or H+/K+-symporter) of uptake is not known. Keunecke et al. (1997) used a patch-clamp approach to unravel the molecular mechanism of xylem unloading. In protoplasts isolated from xylem contact cells in leaves of Vicia faba and maize two different types of K+ channels were identified, but a role in K+ unloading could not be assigned. The small veins of leaves are often surrounded by a layer of compactly arranged cells, the bundle sheath. These cells isolate the veins from the intercellular spaces (the apoplast) of the leaf. Keunecke et al. (2001) showed that uptake of K+ from the vessels of small veins in maize leaves occurs at the xylem/bundle sheath cell interface. An intriguing question is whether the flow of K+ (and other solutes) to the different parts of the shoot is simply determined by the transpiration stream or whether there is at certain points a ‘special filter’ diverting more K+ to, for example, rapidly growing buds/leaves that still have a relatively low transpiration rate? Gunning, Pate & Green (1970) showed that xylem transfer cells in stems of many plants were almost exclusively associated with departing foliar traces (Fig. 4). They estimated that in these regions the membrane surface area of cells surrounding the vessels may be amplified by as much as 20-fold. This, suggests a potential for very intensive lateral exchange of solutes. Furthermore, xylem–phloem exchange is a very important 200 100 0 1 P.media 2 P.maritima 1 P.media 2 P.maritima Figure 5. Different strategies of Plantago media (glycophyte) and P. maritima (halophyte) towards the accumulation of Na+ ions in the root and shoot. The glycophyte has a reduced translocation of Na+ from the root to the shoot, whereas the halophyte preferentially accumulates Na+ in the leaves. Plants were grown on sand supplied with nutrient solution with Na+ concentrations as indicated [from De Boer (1985), where further details can be obtained]. transport step in nutrient distribution and the C/N economy of the plant (Van Bel 1990; White 1997; Jeschke & Hartung 2000). On the other hand, it seems contra-productive to transport large amounts of, for example, K+ to rapidly transpiring mature leaves which are not in need of K+. It will be interesting to study whether the flow of K+ into the petiole of these leaves is restricted by active resorption from the transpiration stream entering that leaf. Sodium Salt-sensitive (glycophytes) and salt-tolerant (halophytes) plants use different strategies when exposed to high Na+ levels in their root medium. In general, glycophytes try to reduce net Na+ uptake from the soil and translocation of Na+ to the shoot, whereas halophytes rapidly absorb and translocate Na+ and sequester it in the vacuole where it is used as ‘cheap’ osmoticum (see Fig. 5a). Especially in halophytes the accumulation of Na+ is at the expense of K+ (Fig. 5b). Therefore it is to be expected that different transport mechanisms for Na+ are present at the symplast/xylem boundary in the root where xylem loading takes place. Certain glycophytes (such as bean, maize and reed) have developed a xylem Na+ unloading mechanism in the basal part of the root and shoot where Na+ ions are taken up from the transpiration stream (Jacoby 1965; Johanson & Cheeseman 1983; Lacan & Durand 1996; Ehwald 2001). Although the capacity of this system is limited, it may play a role in avoiding salt damage in the young growing parts of the shoot at moderate salt stress. Sodium entry into the root symplast is probably mediated by weakly voltage-dependent non-selective cation channels (Roberts & Tester 1995; Tyerman & Skerrett 1999; Davenport & Tester 2000; Demidchik, Davenport & Tester © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 96 A. H. de Boer & V. Volkov 2002; Demidchik & Tester 2002). The properties of these channels resemble those of cyclic nucleotide-gated channels (CNGC) (Leng et al. 1999). Evidence for a role of CNGCs in Na+ uptake is two-fold: (i) the cngc1 mutant shows enhanced Na+ tolerance when grown on moderate external Na+ (Talke et al. 2001); and (ii) membrane-permeable cAMP/cGMP analogues reduced unidirectional Na+ influx and overall Na+ accumulation in NaCl grown Arabiodopsis (Maathuis & Sanders 2001). In the plasma membrane of barley xylem parenchyma cells a non-selective ion channel, called NORC, has been characterized but in contrast to the CNGCs it is non-selective for both cations and anions and it is voltage dependent (Wegner & Raschke 1994; Wegner & De Boer 1997a). Although NORC is equally permeable to K+ and Na+, it remains to be shown whether, under physiological conditions, NORC has a function in loading of Na+ into the transpiration stream. NORC also passes glutamate (Wegner & de Boer 1997a). In this respect it is noteworthy that a glutamate receptor gene, AtGluR2, is expressed in cells adjacent to the conducting vessels in Arabidopsis leaves (Kim et al. 2001). AtGluR2 is homologous to mammalian ionotropic glutamate receptors that are non-selective cation channels permeable to Ca++, K+ and Na+, but not Mg++ (Lam et al. 1998). The primary physiological function of these channels is thought to be Ca++ influx into cells (Dennison & Spalding 2000). Over-expression of AtGluR2 resulted in Ca++-deficiency symptoms and according to Kim et al. (2001) this might be due to increased Ca++ unloading from the transpiration stream in the root and stem causing Ca++ deficiency in the young, growing parts of the plant. The transgenic plants also exhibited hypersensitivity to Na+ and K+ ionic stress, which might be due to reduced Ca++ uptake in the presence of excess Na+ or K+ in combination with increased Ca++ unloading on the way up to the top of the plant. Na+ loading into the xylem of barley and maize roots is not mediated by the outward rectifying K+ channel of xylem parenchyma cells since this channel is virtually impermeable to Na+ (Roberts & Tester 1995; Wegner & De Boer 1997a). Heterologous expression of SKOR in Xenopus oocytes corroborates this conclusion (Gaymard et al. 1998). A prominent candidate gene encoding a Na+ transporter involved in Na+ transport across the symplast/xylem boundary is SOS1 (Ding & Zhu 1997; Shi et al. 2000, 2002). SOS1 was identified as a NaCl-hypersensitive Arabidopsis mutant and initially described as a mutant defective in high-affinity potassium uptake (Wu, Ding & Zhu 1996). The 22Na-uptake and efflux measurements indicated that the sos1 mutants were impaired both in high-affinity K+-and low-affinity Na+uptake (Ding & Zhu 1997). However, in these experiments the whole seedling was exposed to the uptake/efflux solution and this makes interpretation of the results in terms of the SOS1 function in uptake and translocation in planta hazardous. Subsequent sequencing of the locus showed that sos1 encodes a 127 kDa protein with 12 transmembrane spanning domains and with significant sequence similarity to plasma membrane Na+/H+ antiporters from bacteria and fungi (Shi et al. 2000; see also Hasegawa, Bressan & Pardo 2000). In the same article it was reported that SOS1 gene expression is concentrated in the cells surrounding the xylem, suggesting that SOS1 may function in loading Na+ into the xylem for long-distance transport. Recently, the suggestion that SOS1 encodes a Na+/H+ antiporter was experimentally substantiated using purified plasma membrane vesicles from salt-stressed Arabidopsis thaliana leaves (Qiu et al. 2001). Salt-treated sos1 mutant plants showed a 60% lower Na+/H+ exchange activity in comparison with wild-type. SOS1 expression is up-regulated by NaCl stress and this up-regulation is abolished in plants with a mutation in the SOS3 gene encoding a phosphatase with significant sequence similarity with the calcineurin B subunit from yeast (Liu & Zhu 1998). In the presence of Ca++ SOS3 interacts with SOS2 (a protein kinase) and activates the kinase activity of SOS2 (Halfter, Ishitani & Zhu 2000). Apparently, SOS2 directly modulates SOS1 since in vitro addition of SOS2 to plasma membrane vesicles isolated from NaCl stressed plants resulted in a two-fold increase in the Na+/H+ exchange activity (Qiu et al. 2001). During SOS1-facilitated loading of Na+ into the xylem (Shi et al. 2002), one proton flowing from the xylem to the symplast (a ‘down-hill’ process energetically speaking) drives the transport of one Na+ ion into the xylem vessels. Na+ loading into the xylem is unlike K+ loading probably not a channel-mediated process. Much remains to be learned from a comparison of K+ and Na+ exchange across the symplast/xylem boundary in experiments in which the xylem of root or shoot segments is perfused with defined solutions. As discussed above, the high K+ concentration in the xylem parenchyma cells (Drew, Webb & Saker 1990) allows K+ loading down the electrochemical K+ gradient facilitated by the K+-outward channel SKOR (Wegner & de Boer 1997a; Gaymard et al. 1998). In line with the properties of SKOR (depolarization activated and inactivated by external acidification; Lacombe et al. 2000), xylem perfusion of the H+-ATPase activator fusicoccin stops the K+ loading process in roots of the halophyte Plantago maritima (Fig. 3a). Interestingly, the response of Na+ is very different from that of K+: in the first 45 min after the start of fusicoccin perfusion the Na+ concentration in the xylem decreases (i.e. Na+ unloading of the xylem sap is stimulated). This increase in uptake is unlikely to be caused by a pH change of the xylem sap (because this is still small), but rather due to the rapid hyperpolarization of the membrane potential induced by fusicoccin (De Boer & Prins 1985; De Boer et al. 1985). This uptake may be a channel-mediated process. However, when the xylem sap starts to acidify there is a turning point resulting in Na+ loading into the xylem when fusicoccin has acidified the xylem sap pH with more than 1 unit (Fig. 3a). The large pH gradient between xylem sap and the cytoplasm of adjacent parenchyma cells may now facilitate a Na+/H+ antiport system similar to SOS1 (Shi et al. 2002). Similar fusicoccin-induced responses of xylem sap pH and K+/Na+ (un)loading were observed in xylem perfusion experiments using nodal roots of barley (Fig. 3b). © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 97 Shi et al. (2002) proposed a model where SOS1 plays a dual role in Na+ loading and reabsorption from the xylem sap, depending on salinity and pH difference between symplast and xylem, in analogy to a model proposed by Lacan & Durand (1996). The latter authors observed an increase in Na+ unloading and an increase in K+ loading of the xylem sap when the initial pH of the perfusion solution was more acidic. They conclude that Na+ is taken up from the xylem through a Na+/H+ antiport mechanism, but now with H+ driven ‘uphill’ by a ‘down-hill’ Na+ gradient. However, a mechanistic interpretation of their results is difficult since it is not clear how the xylem pH changes in view of the strong pH-stat properties of the xylem (Clarkson et al. 1984). Moreover, it is also not clear how the different initial pH values of the perfusion solution affect the membrane potential. The observed positive correlation between K+ loading and acidity of perfusion solution as reported by Lacan & Durand (1996) is not in line with a SKORmediated loading mechanism since SKOR activity is very low at acidic external pH (Lacombe et al. 2000). It must be kept in mind that perfusion of an acidic buffer will result in an exchange of protons with other cations such as K+ at the negatively charged cell walls of the xylem and that this may cause an apparent increase in K+ loading (Senden et al. 1992). The reabsorption of Na+ from the upgoing transpiration stream in the basal parts of the root and shoot has been well documented both in intact plants and in perfused segments (Jacoby 1965; Johanson & Cheeseman 1983; Lacan & Durand 1996; Ehwald 2001). However, the storage capacity of the stelar cells is limited and therefore a retranslocation of Na+ by the phloem to the root and extrusion into the medium has been suggested as a mechanism to increase the detoxification capacity. Although Van Bel et al. 1992) has already pointed to the importance of xylem– phloem exchange processes nothing is yet known about the mechanism of Na+ loading into the phloem and unloading in the root. Water On its route to the root xylem, water flow is restricted by a number of morphological structures such as the epidermis, outer cortex layer (exodermis), several layers of cortical cells, endodermis, pericycle, parenchyma cells of central cylinder, and cell walls of xylem vessels (Steudle & Peterson 1998). Under conditions of high transpiration, water flow is passively dragged across the roots at a high rate, whereas under high humidity and absence of transpiration the ascending water flow to the shoot is generated by root pressure with low flow rates. In the permanent fluctuating river of water, the aim of a plant is to control ion fluxes to the xylem and to keep most of them relatively independent of water flow. In fact, transpiration may not be required for the supply of minerals to the shoot (Tanner & Beevers 2001). The latter authors provided evidence that xylem flow brought about by root pressure and Münch’s phloem counterflow, might be sufficient for long-distance mineral supply. The plant can discriminate well between water and ions because water molecules have no charge, whereas ions do. Water can move via the apoplast (within cell walls), symplast (moving via cells without leaving them) or via a cellto-cell route (moving from one cell to another, but crossing the membranes, including the tonoplast, of the cells) (Steudle 2000). Generations of physiologists before the 1980s had the firm opinion that water flow occurs mainly in the apoplast apart from the endodermis, where water has to cross the endodermal barrier due to the suberized cell walls of endodermis, known as the Casparian strip (Schreiber 1996). Modelling and hydration kinetic experiments, experiments with cell and root pressure probes and NMR studies showed that water probably moves mainly via the transcellular pathway (= a combination of symplastic and cell-tocell pathways) and therefore via membranes. The arguments in favour of a transcellular route were provided by Newman (1976). In fact, the cross-section of apoplast is small compared with that of the symplast, and water, having a fast equilibrium, would enter the cells even under relatively high flow rates in the apoplast. Much experimental support for the transcellular pathway came from pressure probe measurements, in which the membrane resistance of one cell was compared with the resistance of the whole root. The data demonstrated that the results for a root agreed well with those, when modelling a root as a series of membranes (Jones et al. 1988; Steudle & Peterson 1998). The idea that water molecules not just diffuse through the lipid bilayer of a membrane, but that proteins facilitate rapid exchange of water was corroborated 10 years ago by the cloning of genes encoding water channels or aquaporins (for reviews see Maurel 1997; Kaldenhoff & Eckert 1999; Johansson et al. 2000; Maurel & Chrispeels 2001). Because the water permeability of the lignified secondary walls of the xylem vessels is low, water very likely enters the vessels across the pit membranes separating the xylem parenchyma cells and the vessels. At this point water has to pass the xylem parenchyma plasma membrane underlying the pit membranes. Rough calculations show that water flow across the xylem parenchyma/xylem interface increases one-hundred-fold compared with the flow into the epidermis cells (the vessel radius is about 50 times less than the root radius, with pit fields occupying about 10–20% of the vessel’s surface). So, entry into the xylem is clearly a ‘bottleneck’ in the plants plumbing system and it is to be expected that the density of water channels in these membrane patches is high in order to create a pathway of low resistance to water flow. As early as 1991 it was demonstrated that a putative plasma membrane aquaporin gene (TobRB27) is highly expressed in the central cylinder of roots (Yamamoto et al. 1991). Since the cell-to-cell route of water transport includes passage through the vacuolar membranes (Steudle 2000) it is not surprising that the Arabidopis aquaporin γTIP was found to be highly expressed in the vascular bundles of roots and leaves (Ludevid et al. 1992). In roots of the common ice plant (Mesembryanthemum crystallinum) the plasma membrane aquaporins MIP A and MIP B are expressed in the youngest portions of the xylem and in © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 98 A. H. de Boer & V. Volkov xylem parenchyma cells, respectively (Yamada et al. 1995, 1997). Kirch et al. (2000) corroborated by means of immunolocalization that the MIP-B protein is present in xylem parenchyma cells of Mesembryanthemum roots. An unexpected observation was the presence of MIP-B between xylem vessels, namely in places not expected to contain membranes. Moreover, in these areas and between vessels and xylem parenchyma cells MIP-B was found in small areas in a patchy fashion. Recently, the tobacco plasma membrane aquaporin NtAQP1 was found to be expressed close to the xylem vessels in roots, stems and petioles (Otto & Kaldenhoff 2000). Electron microscopy analysis of a root xylem cross-section showed an uneven distribution of the NtAQP1 protein resembling pitch-like structures (Siefritz et al. 2001). This could indicate that this aquaporin is concentrated at the contact pits between xylem parenchyma cells and xylem vessels ensuring a polarity in the hydraulic resistance of cells adjacent to the vessels. Such a polar distribution of aquaporins could lend support to the hypothesis put forward by Canny (1997a, b) describing the filling of embolized vessels by means of reverse osmosis. Pressurization of cells bordering an embolized vessel could then direct the flow of water along the pathway of least resistance, namely across the pit membrane into the vessel. Although there are still other problems with this model, as indicated by Tyree et al. (1999), it seems worthwhile to study the behaviour of aquaporins in cells bordering an embolized vessel. Fine-tuned control over the fluxes of water into, inside and out of the plant is vital for their struggle to survive in ever-changing environmental conditions. Intense research in the field of aquaporins during the last 10 years has shown that regulation of membrane water permeability by differential expression of a large number of aquaporin genes in combination with post-translational regulation are instrumental for this fine-tuned control (Maurel 1997; Clarkson et al. 2000; Maurel & Chrispeels 2001). A very nice example of such a dynamic regulation is the recently described change in osmotic water permeability of leaf cells in response to the magnitude of the transpiration stream (Morillon & Chrispeels 2001). ACKNOWLEDGMENTS We acknowledge support from the Commission of the European Union via contract FMRX-CT96-0007 (RYPLOS) to A.H. de B. V.V acknowledges support by the Foundation for Single Cell Research and by an IAC fellowship. REFERENCES Allen G.J., Chu S.P., Schumacher K., et al. (2000) Alteration of stimulus-specific guard cell calcium oscillations and stomatal closing in Arabidopsis det3 mutant. Science 289, 2338–2342. Aloni R. & Griffith M. (1992) Functional xylem anatomy in rootshoot junctions of 6 cereal species. Planta 184, 123–129. Canny M.J. (1997a) Vessel contents of leaves after excision – a test of Scholander’s assumption. American Journal of Botany 84, 1217–1222. Canny M.J. (1997b) Vessel contents during transpiration- embolisms and refilling. American Journal of Botany 84, 1223–1230. Chaffey N.J. & Barlow P.W. (2001) The cytoskeleton facilitates a three-dimensional symplasmic continuum in the long-lived ray and axial parenchyma cells of angiosperm trees. Planta 213, 811– 823. Chaffey N.J., Barlow P.W. & Barnett J.R. (2000) A cytoskeletal basis for wood formation in angiosperm trees: the involvement of microfilaments. Planta 210, 890–896. Clarkson D.T., Carvajal M., Henzler T., Waterhouse R.N., Smyth A., Cooke D.T. & Steudle E. (2000) Root hydraulic conductance: diurnal aquaporin expression and the effects of nutrient stress. Journal of Experimental Botany. 51, 61–70. Clarkson D.T., Williams L. & Hanson J.B. (1984) Perfusion of onion root xylem vessels: a method and some evidence of control of the pH of the xylem sap. Planta 162, 361–369. Cochard H., Ewers F.W. & Tyree M.T. (1994) Water relations of a tropical vine-like bamboo (Rhipidocladum racemiflorum): root pressures, vulnerability to cavitation and seasonal changes in embolism. Journal of Experimental Botany 45, 1085–1089. Crafts A.S. & Broyer T.C. (1938) Migration of salts and water into xylem of the roots of higher plants. American Journal of Botany 25, 529–535. Cram W.J. & Pitman M.G. (1972) The action of abscisic acid on ion uptake and water flow in plant roots. Australian Journal of Biological Sciences 25, 1125–1132. Davenport R.J. & Tester M. (2000) A weakly voltage-dependent, non-selective cation channel mediates toxic sodium influx in wheat. Plant Physiology 122, 823–834. De Boer A.H. (1985) Xylem/symplast ion exchange: mechanism and function in salt-tolerance and growth. PhD Thesis, University of Groningen, The Netherlands. De Boer A.H. (1997) Fusicoccin- a key to multiple 14–3−3 locks? Trends in Plant Science 2, 60–66. De Boer A.H. & Prins H.B.A. (1984) Trans-root potential in roots of Plantago media L. as affected by hydrostatic pressure: the induction of an O2 deficient root core. Plant and Cell Physiology 25, 643–655. De Boer A.H. & Prins H.B.A. (1985) Xylem perfusion of tap root segments of Plantago maritima: the physiological significance of electrogenic xylem pumps. Plant, Cell and Environment 8, 587– 594. De Boer A.H., Katou K., Mizuno A., Kojima H. & Okamoto H. (1985) The role of electrogenic xylem pumps in K+ absorption from the xylem of Vigna unguiculata: the effects of auxin and fusicoccin. Plant, Cell and Environment 8, 579–586. De Boer A.H., Prins H.B.A. & Zanstra P.E. (1983) Biphasic composition of trans-root potential of Plantago species: involvement of spatially separated electrogenic pumps. Planta 157, 259–266. Demidchik V. & Tester M. (2002) Sodium fluxes through nonselective cation channels in the plasma membrane of protoplasts from Arabidopsis roots. Plant Physiology 128, 379–387. Demidchik V., Davenport R.J. & Tester M. (2002) Nonselective cation channels in plants. Annual Review of Plant Physiology and Plant Molecular Biology 53, 67–107. Dennison K.L. & Spalding E.P. (2000) Glutamate-gated calcium fluxes in Arabidopsis. Plant Physiology 124, 1511–1514. Ding L. & Zhu J.-K. (1997) Reduced Na+ uptake in the NaClhypersensitive sos1 mutant of Arabidopsis thaliana. Plant Physiology 113, 795–799. Drew M.C., Webb J. & Saker L.R. (1990) Regulation of K+ uptake and transport to the xylem in barley roots: K+ distribution determined by electron probe X-ray microanalysis of frozen-hydrated cells. Journal of Experimental Botany 41, 815–825. © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 99 Dunlop J. (1982) Membrane potentials in the xylem in roots of intact plants. Journal of Experimental Botany 33, 910–918. Ehwald R. (2001) Salt and water stress of common reed growing in wind-protected flat brackish locations. In International Symposium ‘Plants Under Environmental Stress’, Moscow, October, 23–28, pp. 66–67. Timiriazev Institute, Moscow, Russia. Engels C. & Marschner H. (1992) Adaptation of potassium translocation into the shoot of maize (Zea mays) to shoot demand: evidence for xylem loading as a regulating step. Physiologia Plantarum 86, 263–268. Enns L.C., Canny M.J. & McCully M.E. (2000) An investigation of the role of solutes in the xylem sap and in the xylem parenchyma as the source of root pressure. Protoplasma 211, 183–197. Ferguson I.B. & Bollard E.G. (1976) The movement of calcium in woody stems. Annals of Botany 40, 1057–1065. Fisher J.B., Angeles G., Ewers F.W. & Lopez-Portillo J. (1997) Survey of root pressure in tropical vines and woody species. International Journal of Plant Science 158, 44–50. Fukuda H. (1996) Xylogenesis: initiation, progression, and cell death. Annual Review of Plant Physiology and Plant Molecular Biology 47, 299–325. Gansert D., Burgdorf M. & Lösch R. (2001) A novel approach to the in situ measurement of oxygen concentrations in the sapwood of woody plants. Plant, Cell and Environment 24, 1055– 1064. Gaymard F., Pilot G., Lacombe B., Bouchez D., Bruneau D., Boucherez J., Michaux-Ferrière N., Thibaud J.-B. & Sentenac H. (1998) Identification and disruption of a plant Shaker-like outward channel involved in K+ release into the xylem sap. Cell 94, 647–655. Gehring C.A., Irving H.R. & Parish R.W. (1990) Effects of auxin and abscisic acid on cytosolic calcium and pH in plant cells. Proceedings of the National Academy of Sciences USA 87, 9645– 9649. Grace J. (1993) Refilling of embolized xylem. In Water Transport in Plants Under Climatic Stress (eds M. Borghetti, J. Grace, & A. Raschi), pp. 52–62. Cambridge University Press, Cambridge, UK. Gunning B.E.S. (1977) Transfer cells and their roles in transport of solutes in plants. Scientific Progress, Oxford 64, 539–568. Gunning B.E.S., Pate J.S. & Green L.W. (1970) Transfer cells in the vascular system of stems: taxonomy, association with nodes, and structure. Protoplasma 71, 147–171. Halfter U., Ishitani M. & Zhu J.-K. (2000) The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proceedings of the National Academy of Sciences USA 97, 3735–3740. Hasegawa M., Bressan R. & Pardo J.M. (2000) The dawn of plant salt tolerance genetics. Trends in Plant Science 8, 317–319. Herdel K., Schmidt P., Feil R., Mohr A. & Schurr U. (2001) Dynamic of concentrations and nutrient fluxes in the xylem of Ricinus communis – diurnal course, impact of nutrient availability and nutrient uptake. Plant, Cell and Environment 24, 41–52. Holbrook N.M. & Zwieniecki M.A. (1999) Embolism repair and xylem tension: do we need a miracle? Plant Physiology 120, 7– 10. Holbrook N.M., Ahrens E.T., Burns M.J. & Zwieniecki M.A. (2001) In vivo observation of cavitation and embolism repair using magnetic resonance imaging. Plant Physiology 126, 27–31. Huber B. & Merz W. (1958) Über die Bedeutung des Hoftüpfelverschlusses für die axiale Wasserleitfähigkeit von Nadelhölzern. Planta 51, 645–672. Hwang J.-U., Suh S., Kim J. & Lee Y. (1997) Actin filaments modulate both stomatal opening and inward K+-channel activities in guard cells of Vicia faba L. Plant Physiology 115, 335–342. Jacoby B. (1965) Sodium retention in excised bean stems. Physiologia Plantarum 18, 730–739. Jeschke W.D. & Hartung W. (2000) Root–shoot interactions in mineral nutrition. Plant and Soil 226, 57–69. Johanson J.G. & Cheeseman J.M. (1983) Uptake and distribution of sodium and potassium by corn seedlings. Role of the mesocotyl in sodium exclusion. Plant Physiology 73, 153–158. Johansson I., Karlsson M., Johanson U., Larsson C. & Kjellbom P. (2000) The role of aquaporins in cellular and whole plant water balance. Biochimica Biophysica Acta 1465, 324–342. Jones H., Leigh R.A., Wyn Jones R.G. & Tomos A.D. (1988) The integration of whole-root and cellular hydraulic conductivities in cereal roots. Planta 174, 1–7. Kaldenhoff R. & Eckert M. (1999) Features and function of plant aquaporins. Journal of Photochemistry and Photobiology B 52, 1–6. Keunecke M., Lindner B., Seydel U., Schulz A. & Hansen U.P. (2001) Bundle sheath cells of small veins in maize leaves are the loction of uptake from the xylem. Journal of Experimental Botany 52, 709–714. Keunecke M., Sutter J.U., Sattelmacher B. & Hansen U.P. (1997) Isolation and patch clamp measurements of xylem contact cells for the study of their role in the exchange between apoplast and symplast of leaves. Plant and Soil 196, 239–244. Kim S.A., Kwak J.M., Jae S.-K., Wang M.-H. & Nam H.G. (2001) Overexpression of the AtGluR2 gene encoding an Arabidopsis homolog of mammalian glutamate receptors impairs calcium utilization and sensitivity to ionic stress in transgenic plants. Plant and Cell Physiology 42, 74–84. Kirch H.-H., Vera-Estrella R., Golldack D., Quigley F., Michalowski C.B., Barkla B.J. & Bohnert H.J. (2000) Expression of water channel proteins in Mesembryanthemum crystallinum. Plant Physiology 123, 111–124. Kramer D., Läuchli A., Yeo A.R. & Gullasch J. (1977) Transfer cells in roots of Phaseolus coccineus: ultrastructure and possible function in exclusion of sodium from the shoot. Annals of Botany 41, 1031–1040. Kuo J., Pate J.S., Rainbird R.M. & Atkins C.A. (1980) Internodes of grain legumes – new location for xylem parenchyma transfer cells. Protoplasma 104, 181–185. Lacan D. & Durand M. (1996) Na+-K+ exchange at the xylem/ symplast boundary. Plant Physiology 110, 705–711. Lachaud S. & Maurousset L. (1996) Occurrence of plasmodesmata between differentiating vessels and other cells in Sorbus torminalis L. Crantz and their fate during xylem maturation. Protoplasma 191, 220–226. Lacombe B., Pilot G., Gaymard F., Sentenac H. & Thibaud J.-B. (2000) pH control of the plant outward-rectifying potassium channel SKOR. FEBS Letters 466, 351–354. Lam H.-M., Chiu J., Hieh M.-H., Meisel L., Oliveira I.C., Shin M. & Coruzzi G. (1998) Glutamate-receptor genes in plants. Nature 396, 125–126. Läuchli A., Kramer D., Pitman M.G. & Lüttge U. (1974) Ultrastructure of xylem parenchyma cells of barley roots in relation to ion transport to the xylem. Planta 119, 85–99. Leng Q., Mercier R.W., Yao W. & Berkowitz G.A. (1999) Cloning and functional characterization of a plant nucleotide-gated cation channel. Plant Physiology 121, 753–761. Liu J. & Zhu J.-K. (1998) A calcium sensor homolog required for plant salt tolerance. Science 280, 1943–1945. López-Millán A.F., Morales F., Abadía A. & Abadía J. (2000) Effects of iron deficiency on the composition of the leaf apoplastic fluid and xylem sap in sugar beet. Implications for iron and carbon transport. Plant Physiology 124, 873–884. Ludevid D., Höfte H., Himmelblau E. & Chrispeels M.J. (1992) The expression pattern of the tonoplast intrinsic γ-TIP in Ara- © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 100 A. H. de Boer & V. Volkov bidopsis thaliana is correlated with cell enlargement. Plant Physiology 100, 1633–1639. Maathuis F.J.M. & Sanders D. (2001) Sodium uptake in Arabidopsis roots is regulated by cyclic nucleotides. Plant Physiology 127, 1617–1625. Maurel C. (1997) Aquaporins and water permeability of plant membranes. Annual Review of Plant Physiology and Plant Molecular Biology 48, 399–429. Maurel C. & Chrispeels M.J. (2001) Aquaporins. A molecular entry into plant water relations. Plant Physiology 125, 135– 138. McCann M.C. (1997) Tracheary element formation: building up to a dead end. Trends in Plant Science 2, 333–338. McCully M.E. (1999) Root xylem embolisms and refilling. Relation to water potentials of soil, roots, and leaves, and osmotic potentials of root xylem sap. Plant Physiology 119, 1001–1008. McCully M.E., Huang C.X. & Ling L.E.C. (1998) Daily embolism and refilling of xylem vessels in the roots of field-grown maize. New Phytologists 138, 327–342. Meychik N.R. & Yermakov I.P. (2001) Swelling of root cell walls as an indicator of their functional state. Biochemistry (Moscow) 66, 178–187. Morillon R. & Chrispeels M.J. (2001) The role of ABA and the transpiration stream in the regulation of the osmotic water permeability of leaf cells. Proc. Natl. Acad. Sci. USA 98, 14138– 14143. Mueller W.C. & Beckman C.H. (1984) Ultrastructure of the cell wall of vessel contact cells in the xylem of tomato stems. Annals of Botany 53, 107–114. Newman E.I. (1976) Water movement through root systems. Philosophical Transactions of Royal Society London. Series B. 273, 463–478. Okamoto H., Katou K. & Ichino K. (1979) Distribution of electrical potential and ion transport in the hypocotyl of Vigna sesquipedalis. VI. The dual structure of radial electrogenic activity. Plant and Cell Physiology 20, 103–114. Oparka K.J. & Cruz S.S. (2000) The great escape: phloem transport and unloading of macromolecules. Annual Review of Plant Physiology and Plant Molecular Biology 51, 323–347. Otto B. & Kaldenhoff R. (2000) Cell-specific expression of the mercury-insensitive plasma-membrane aquaporin NtAQP1 from Nicotiana tabacum. Planta 211, 167–172. Parets-Soler A., Pardo J.M. & Serrano R. (1990) Immunolocalization of plasma membrane H+-ATPase. Plant Physiology 93, 1654–1658. Pate J.S. & Gunning B.E.S. (1972) Transfer cells. Annual Review of Plant Physiology 23, 173–196. Patrick J.W., Zhang W.H., Tyerman S.D., Offler C.E. & Walker N.A. (2001) Role of membrane transport in phloem translocation of assimilates and water. Australian Journal of Plant Physiology 13, 695–707. Peterson C.A. & Steudle E. (1993) Lateral hydraulic conductivity of early metaxylem vessels in Zea mays L. roots. Planta 189, 288– 297. Peuke A.D., Rokitta M., Zimmermann U., Schreiber L. & Haase A. (2001) Simultaneous measurement of water flow velocity and solute transport in xylem and phloem of adult plants of Ricinus communis over a daily time course by nuclear magnetic resonance spectrometry. Plant, Cell and Evironment 24, 491– 503. Pitman M.G. (1977) Ion transport into the xylem. Annual Review of Plant Physiology 28, 71–88. Pitman M.G. & Wellfare D. (1978) Inhibition of ion transport in excised barley roots by abscisic acid: relation to water permeability of the roots. Journal of Experimental Botany 29, 1125– 1138. Qiu Q.-S., Guo Y., Dietrich M.A., Schumaker K.S. & Zhu J.-K. (2001) Characterization of the plasma membrane Na+/H+ exchanger in Arabidopsis thaliana. In The 12th International Workshop Plant Membrane Biology, Madison, p. 235. University of Wisconsin, Madison, WI, USA. Roberts S.K. (1998) Regulation of K+ channels in maize roots by water stress and abscisic acid. Plant Physiology 116, 145–153. Roberts K. & McCann M.C. (2000) Xylogenesis: the birth of a corpse. Current Opinion in Plant Biology 3, 517–522. Roberts S.K. & Tester M. (1995) Inward and outward K+-selective currents in the plasma membrane of protoplasts from maize root cortex and stele. Plant Journal 8, 811–825. Rokitta M., Peuke A.D., Zimmermann U. & Haase A. (1999) Dynamic studies of phloem and xylem flow in fully differentiated plants by fast nuclear-magnetic-resonance microimaging. Protoplasma 209, 126–131. Rygol J., Pritchard J., Zhu J.J. & Zimmermann U. (1993) Transpiration induces radial turgor pressure gradients in wheat and maize roots. Plant Physiology 103, 493–500. Salleo S., Lo Gullo M.A., De Paoli D. & Zippo M. (1996) Xylem recovery from cavitation-induced embolism in young plants of Laurus nobilis: a possible mechanism. New Phytologist 132, 47– 56. Samuels A.L., Fernando M. & Glass A.D.M. (1992) Immunofluorescent localization of plasma membrane H+-ATPase in barley roots and effects of K nutrition. Plant Physiology 99, 1509– 1514. Sauter J.J., Iten W. & Zimmermann M.H. (1973) Studies on the release of sugar into the vessels of sugar maple (Acer saccharum). Canadian Journal of Botany 51, 1–8. Scheenen T.W.J., Van Dusschoten D., De Jager P.A. & Van As H. (2000) Quantification of water transport in plants with NMR imaging. Journal of Experimental Botany 51, 1751–1759. Schreiber L. (1996) Chemical composition of Casparian strips isolated from Clivia miniata Reg. roots: evidence for lignin. Planta 199, 596–601. Schurr U. (1998) Xylem sap sampling – new approaches to an old topic. Trends in Plant Science 3, 293–298. Schurr U. & Schulze E.-D. (1995) The concentration of xylem sap constituents in root exudate, and in sap from intact, transpiring castor bean plants (Ricinus communis L.). Plant, Cell and Environment 18, 409–420. Senden M.H.M.N., Van Paassen F.J.M., van der Meer A.J.G.M. & Wolterbeek H.Th. (1992) Cadmium – citric acids – xylem cell wall interactions in tomato plants. Plant, Cell and Environment 15, 71–79. Shi H., Ishitani M., Kim C. & Zhu J.-K. (2000) The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proceedings of the National Academy of Sciences USA 97, 6896–6901. Shi H., Quintero F.J., Pardo J.M. & Zhu J.-K. (2002) The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell. 14, 465–477. Siau J.F. (1984) Transport Processes in Wood. Springer, Berlin. Siefritz F., Biela A., Eckert M., Otto B., Uhlein N. & Kaldenhoff R. (2001) The tobacco plasma membrane aquaporin NtAQP1. Journal of Experimental Botany 52, 1953–1957. Smart C.C. & Amrhein N. (1985) The influence of lignification on the development of vascular tissue in Vigna radiata L. Protoplasma 124, 87–95. Spalding E.P., Hirsch R.E., Lewis D.R., Qi Z., Sussman M.R. & Lewis B.D. (1999) Potassium uptake supporting plant growth in the absence of AKT1 channel activity. Inhibition by ammonium and stimulation by sodium. Journal of General Physiology 113, 909–918. Sperry J.S., Holbrook N.M., Zimmermann M.H. & Tyree M.T. © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101 Structural and functional features of xylem 101 (1987) Spring filling of xylem vessels in wild grapevine. Plant Physiology 83, 414–417. St. Aubin G., Canny M.J. & McCully M.E. (1986) Living vessel elements in the late metaxylem of sheathed maize roots. Annals of Botany 58, 577–588. Steudle E. (2000) Water uptake by plant roots: an integration of views. Plant and Soil 226, 45–56. Steudle E. & Peterson C.A. (1998) How does water get through roots? Journal of Experimental Botany 49, 775–788. Talke I.N., Bouché N., Bouchez D., Fromm H., Sanders D. & Maathuis F.J.M. (2001) Characterization of CNGC1, a putative cyclic nucleotide-gated channel from Arabidopsis thaliana. In The 12th International Workshop Plant Membrane Biology, Madison, p. 69. University of Wisconsin, Madison, WI, USA. Tanner W. & Beevers H. (2001) Transpiration, a prerequisite for long-distance transport of minerals in plants? Proceedings of the National Academy of Sciences USA 98, 9443–9447. Turner S.R. & Somerville C.R. (1997) Collapsed xylem phenotype of Arabidopsis identifies mutants deficient in cellulose deposition in the secondary cell wall. Plant Cell 9, 689–701. Tyerman S.D. & Skerrett I.M. (1999) Root ion channels and salinity. Scientia Horticulturae 78, 175–235. Tyree M.T. & Sperry J.S. (1989) Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Physiology and Plant Molecular Biology 40, 19–38. Tyree M.T. & Yang S. (1992) Hydraulic conductivity recovery versus water pressure in xylem of Acer saccharum. Plant Physiology 100, 669–676. Tyree M.T., Fiscus S.D., Wullschleger M.A. & Dixon M.A. (1986) Detection of xylem cavitation in corn under field conditions. Plant Physiology 82, 597–599. Tyree M.T., Salleo S., Nardini A., Lo Gullo M.A. & Mosca R. (1999) Refilling of embolized vessels in young stems of Laurel. Do we need a new paradigm? Plant Physiology 120, 11–21. Van Bel A.J.E. (1990) Xylem-phloem exchange via the rays: the undervalued route of transport. Journal of Experimental Botany 41, 631–644. Van Bel A.J.E., Gamalei Y.V., Ammerlaan A. & Bik L.P.M. (1992) Dissimilar phloem loading in leaves with symplastic or apoplastic minor-vein configurations. Planta 186, 518–525. Van der Veen R., Heimovaraa-Dijkstra S. & Wang M. (1992) Cytosolic alkalinization mediated by abscisic acid is necessary, but not sufficient, for abscisic acid-induced gene expression in barley aleurone protoplasts. Plant Physiology 100, 699–705. Van Ieperen W., Van Meeteren U. & van Gelder H. (2000) Fluid composition influences hydraulic conductance of xylem conduits. Journal of Experimental Botany 51, 769–776. Watson R., Pritchard J. & Malone M. (2001) Direct measurement of sodium and potassium in the transpiration stream of saltexcluding and non-excluding varieties of wheat. Journal of Experimental Botany 52, 1873–1881. Wegner L.H. & De Boer A.H. (1997a) Properties of two outwardrectifying channels in root xylem parenchyma cells suggest a role in K+ homeostasis and long-distance signaling. Plant Physiology 115, 1707–1719. Wegner L.H. & De Boer A.H. (1997b) Two inward K+ channels in the xylem parenchyma cells of barley roots are regulated by Gprotein modulators through a membrane-delimited pathway. Planta 203, 506–516. Wegner L.H. & Raschke K. (1994) Ion channels in the xylem parenchyma of barley roots. Plant Physiology 105, 799– 813. Wegner L.H. & Zimmermann U. (1998) Simultaneous recording of xylem pressure and trans-root potential in roots of intact glycophytes using a novel xylem pressure probe technique. Plant, Cell and Environment 2, 849–865. Wegner L.H., Sattelmacher B., Läuchli A. & Zimmermann U. (1999) Trans-root potential, xylem pressure, and root cortical membrane potential of ‘low-salt’ maize plants as influenced by nitrate and ammonium. Plant, Cell and Environment 22, 1549– 1558. White P.J. (1997) The regulation of K+ influx into roots of rye (Secale cereale L.) seedlings by negative feedback via the K+ flux from the shoot to root in the phloem. Journal of Experimental Botany 48, 2063–2073. Wilkinson S. & Davies W.J. (1997) Xylem sap pH increase: a drought signal received at the apoplastic face of the guard cell that involves the suppression of saturable abscisic acid uptake by the epidermal symplast. Plant Physiology 113, 559–573. Winter-Sluiter E., Läuchli A. & Kramer D. (1977) Cytochemical localization of K+-stimulated adenosine-triphosphate activity in xylem parenchyma of barley roots. Plant Physiology 60, 923– 927. Wisniewski M., Ashworth E. & Schaffer K. (1987) The use of lanthanum to characterize cell wall permeability in relation to depp supercooling and extracellular freezing in woody plants. I. Intergeneric comparisons between Prunus, Cornus and Salix. Protoplasma 139, 105–116. Wu S.-J., Ding L. & Zhu J.-K. (1996) SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8, 617–627. Yamada S., Katsuhara M., Kelly W., Michalowski C. & Bohnert H. (1995) A family of transcripts encoding the water channel proteins: tissue specific expression in the common ice plant. Plant Cell 7, 1129–1142. Yamada S., Nelson D.E., Ley E., Marquez S. & Bohnert H.J. (1997) The expression of an aquaporin promoter from Mesembryanthemum crystallinum in tobacco. Plant and Cell Physiology 38, 1326–1332. Yamamoto Y.T., Taylor C.G., Acedo G.N., Cheng C.L. & Conkling M.A. (1991) Characterization of cis-acting sequences regulating root-specific gene expression in tobacco. Plant Cell 3, 371–382. Yeo A.R., Kramer D., Läuchli. A. & Gullasch J. (1977) Ion distribution in salt-stressed mature Zea mays roots in relation to ultrastructure and retention of sodium. Journal of Experimental Botany 28, 17–29. Zimmermann M.H. (1978) Hydraulic architecture of some diffuseporous trees. Canadian Journal of Botany 56, 2286–2295. Zimmermann M.H. (1983) Xylem Structure and the Ascent of Sap. Berlin: Springer-Verlag. Zwieniecki M.A. & Holbrook N.M. (2000) Bordered pit structure and vessel wall surface properties. Implications for embolism repair. Plant Physiology 123, 1015–1020. Zwieniecki M.A., Melcher P.J. & Holbrook N.M. (2001) Hydrogel control of xylem hydraulic resistance in plants. Science 291, 1059–1062. Received 11 December 2001; received in revised form 15 May 2002; accepted for publication 20 May 2002 © 2003 Blackwell Publishing Ltd, Plant, Cell and Environment, 26, 87–101