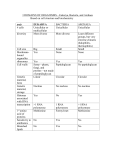
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Research in Microbiology 162 (2011) 77e91 www.elsevier.com/locate/resmic A new fusion hypothesis for the origin of Eukarya: better than previous ones, but probably also wrong Patrick Forterre a,b,* b a Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France Institut de Ge´ne´tique et Microbiologie, Univ. Paris-Sud - CNRS UMR 8621, 91405 Orsay Cedex, France Available online 27 October 2010 Abstract I discuss here the possibility that Eukarya originated from the engulfment of a thaumarchaeon by a PCV (Planctomycetes, Verrucomicrobia, Chlamydiae) bacterium, followed by invasions of NCLDV and retroviruses. The thaumarchaeon provided both informational and operational proteins (actins, ESCRT proteins), including some essential proteins absent in other archaeal phyla (Topo IB), whereas the PVC bacterium provided phospholipids, tubulin and the membrane coat proteins required for the formation of the nucleus. Viral invasions introduced many proteins that are specific to modern Eukarya and produced an arms race that favoured the evolution of Eukarya toward increasing complexity. This scenario is the best possible fusion hypothesis that can be presently proposed. However, it still requires several ad hoc assumptions to explain the origin of the nucleus and the distribution pattern of archaeal and bacterial traits in modern Eukarya. Furthermore, it still fails to explain convincingly the origin of eukaryal viruses and the existence of three distinct lineages of ribosomes. I conclude that Eukarya and their viruses more probably evolved from a specific lineage, according to the three domains scenario originally proposed by Carl Woese. Ó 2010 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. Keywords: Archaea; Thaumarchaea; Planctomycetes; NCLDV; Retrovirus; Origin of eukaryotes 1. Introduction The prokaryote/eukaryote dichotomy, set up in 1962 by Stanier and Van Niel (Stanier and Van Niel, 1962), based on cell biology, has been the dominant paradigm in biology for the last 50 years. It remains today accepted by most biologists despite the fundamental work of Carl Woese and colleagues who demonstrated in 1977, based on molecular biology, the existence of three (and not two) major evolutionary cellular lineages, Archaea, Bacteria and Eukarya (Woese and Fox, 1977; Woese et al., 1990). The procaryote paradigm indeed satisfies the traditional idea that evolution always progresses from simple organisms to complex organisms (see a critique of this idea in Forterre and Philippe, 1999). It is inherent in the terms prokaryote/eukaryote themselves, since « pro » karyotes implies * Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France. Tel.: þ33 169157489. E-mail address: [email protected]. before « eu » karyotes (true nucleus) (Forterre, 1995; Pace, 2009). I suggested in 1992 to replace the terms prokaryote and eukaryote by neutral ones (evolutionary speaking) such as akaryote and synkaryotes (without and with nucleus) to designate the two types of known cellular organization (Forterre, 2002), but this proposal was completely ignored. Another reason explaining the resistance of the prokaryote paradigm is that we (human beings) like dichotomies, so the prokaryote/ eukaryote dichotomy is regularly revived, like the phoenix, even if it is contradicted by facts. In recent years, the prokaryote/eukaryote dichotomy has been on the agenda again under a new appearance, i.e. “fusion” hypotheses in which Eukarya originated from the association of an archaeon and a bacterium (see discussions in Poole and Penny, 2007; Gribaldo et al., 2010; Cavalier-Smith, 2010; Koonin, 2010a,b, Forterre, 2010b). These hypotheses are based on the observation that Eukarya and Archaea share similar informational mechanisms (translation, transcription, replication) very distinct (sometimes non homologous) from 0923-2508/$ - see front matter Ó 2010 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. doi:10.1016/j.resmic.2010.10.005 78 P. Forterre / Research in Microbiology 162 (2011) 77e91 those of Bacteria, whereas Eukarya and Bacteria share a similar type of membrane phospholipid, very distinct from those of Archaea (see below). Bacteria and Eukarya also exclusively share many proteins (often involved in so-called operational mechanisms) whose origin is disputed. Some authors suppose that these proteins have been all introduced in Eukarya either by the alpha-proteobacterium at the origin of mitochondria or by other bacteria used as preys by ancient Eukarya. In contrast, proponents of “fusion” hypotheses argue that these proteins testify for the existence of a specific bacterium at the origin of Eukarya, whose proteome was supplemented by the informational proteins of an archaeon. The fusions scenarios that have been proposed by different authors involve various types of associations, symbioses, syntrophy, etc. often justified by metabolic considerations to explain why the two partners first of all associated. I will put all these types of combinations here under the broad term “fusion” for the sake of simplicity. Proponents of fusion scenarios for the origin of Eukarya usually still use the old name archaebacteria, instead of Archaea, thus perpetuating the confusion between Bacteria and prokaryotes. In that scheme, Bacteria (as a general term encompassing eubacteria and archaebacteria) regain the primitive position that they have occupied in the tree of life from the XIX century (the Haeckel’s monera). In fusion scenarios, the three primary domains of Woese and colleagues are thus replaced by two primary “prokaryotic” domains, and “eukaryotes” therefore emerged once more after prokaryotes, as a derived secondary domain (see discussion in Gribaldo et al., 2010). Proponents of fusion scenarios on the origin of Eukarya often play of the confusion between Eukarya in general (including those which lived before the acquisition of mitochondria) and modern Eukarya. The latter indeed originated after Bacteria, since their last common ancestor already had bona fide mitochondria. However, in the three domains scenario, the primordial eukaryal lineage (urkaryote sensu Woese and Fox, 1977) does not correspond to modern Eukarya, but to a lineage of ancient Eukarya that led to LECA (the Last Eukaryal Common Ancestor) (Fig. 1A). This lineage might be as ancient as those that led to Bacteria and Archaea. In this scenario, FECA (the First Eukaryal Common Ancestor) might have even predated the divergence of the various phyla of modern archaea and bacteria (Fig. 1A). In contrast, in fusion scenarios, FECA only appeared after the diversification of the archaeal and bacterial domains (Fig. 1B). Indeed, with few exceptions, proponents of fusion scenarios for the origin of Eukarya have always proposed that the two partners that associate were close relatives of modern Bacteria and Archaea. Historically, various archaeal and bacterial partners have been proposed by different authors to sell convincingly their particular fusion hypotheses. Lynn Margulis, suggested in the eighties that the archaeal partner was a relative of Thermoplasma acidophilum (Margulis and Bermudes, 1985), a member of the archaeal phylum Euryarchaeota, because T. acidophilum is a wall-less microorganism in which “eukaryotic-like histones” were described (biochemically) in the seventies (Stein and Searcy, 1978). In her hypothesis, the bacterial partner was a Spirochete because these bacteria harbour microtubule-like structures reminiscent of the eukaryal cytoskeleton (Margulis et al., 1979). Later on, Rivera and Lake suggested an archaeon of the phylum Crenarchaeota for the archaeal partner, because Crenarchaeota (called Eocytes by Lake) were sister groups of Eukarya in their phylogenetic analyses (Rivera and Lake, 1992, 2004). More recently, in the “ring of life” scenario, they identified the bacterial partner as a relative of Proteobacteria and/or Cyanobacteria (Rivera and Lake, 2004). In the so-called, “hydrogen hypothesis”, Martin and Muller suggested an unusual version of fusion hypotheses in which the bacterial partner corresponds to the alpha-proteobacterium at the origin of mitochondria (Fig. 1B, arrow b). In their scenario, the “archaebacterium” is an unspecified autotrophic hydrogen-dependent anaerobe (Martin and Müller, 1998). One of the more detailed and original fusion hypothesis is the “syntrophic hypothesis” proposed by Moreira and Lopez- Fig. 1. Two alternative scenarios for the origin and evolution of modern domains. A: Scenario based on the three domains concept (dotted lines symbolize an unrooted tree), B: Fusion scenario based on the prokaryote/eukaryote dichotomy. The green arrows represent the origin of the mitochondrion from an alphaproteobacterium. FECA: First Eukaryotic (specific) Common Ancestor, LECA: Last Eukaryotic Common Ancestor, FME: First Mitochondrial Eukaryote. P. Forterre / Research in Microbiology 162 (2011) 77e91 Garcia (1998). These authors suggested a mesophilic methanogen (more precisely a Methanobacteriale) for the archaeal partner, and a delta-proteobacterium (more precisely an ancestral sulfate-reducing myxobacterium) for the bacterial one, their complementary physiology being supposed to explain their association based on a metabolic symbiosis (interspecies hydrogen transfer). They also justified the choice of a methanogen on molecular basis, i.e. the presence in these mesophilic archaea of eukaryal histone homologues (lacking in Crenarchaea/Eocytes). Similarly, a myxobacterium was proposed as bacterial partner because these social and complex bacteria were supposed to have provided the primordial genes for the social and developmental behaviour of later Eukarya, and because Myxobacteria were supposed to be at that time the only “prokaryotes” encoding serine/threonine protein kinases or small G-proteins (supposed to be eukaryotic hallmaks). Later on, the idea of an association between a methanogenic archaeon and a bacterium was also endorsed by Bell. This author combined the scenarios of Moreira and Lopez-Garcia with the hydrogen hypothesis of Martin and Muller (but removing the myxobacterium) and introduces in the story a giant Pox-like virus infecting the methanogenic archaeon. In this me´nage à trois, the alpha-proteobacterium was transformed into a mitochondrion, whereas the giant virus became the nucleus of modern Eukarya (Bell, 2001). Detailed fusion hypotheses for the origin of Eukarya made during the last years are now open for falsification (or at least criticism) following new discoveries in archaeal and/or bacterial physiology, molecular biology and genomics. These discoveries often make obsolete the reasons that initially justified the choice of a particular partner of the fusion. For instance, the case for a Thermoplasma-like archaeon at the origin of the eukaryotic nucleus became much weaker when it turned out that the “eukaryal-like histones” of Thermoplasmatales are not homologous to eukaryal histones but to the bacterial HU protein (DeLange et al., 1981; Sandman et al. 1990). Similarly, the idea 79 that Eukarya partly derived from methanogens because the presence of eukaryal-like histones in these mesophilic archaea became obsolete when similar histones were discovered in mesophilic thaumarchaea (Cubonová et al., 2005, BrochierArmanet et al., 2008a,b). The case for a spirochete or a deltaproteobacterium at the origin of Eukarya also became weak once the sequencing of their genomes failed to reveal specific affinities with Eukarya and after the discovery of protein kinases and Gproteins in many groups of Archaea and Bacteria (Dong et al., 2007; Tyagi et al., 2010). The choice of two specific partners in fusion hypotheses has always been dependant of data available at a given time in terms of microbial diversity, physiology and molecular biology. Interestingly, recent discoveries have revealed novel eukaryal features in members of the bacterial PVC superphylum, that includes Planctomycetes, Verrucomicrobia, Chlamydiae, Lentisphaera and Poribacteria (Wagner and Horn, 2006), and in members of the recently recognized archaeal phylum Thaumarchaeota (whose mesophilic members were formerly assimilated to Crenarchaeota) (Brochier-Armanet et al., 2008a). Proteins similar (probably homologous) to eukaryal membrane coat (MC) proteins have been discovered in PVC bacteria (Santarella et al., 2010), whereas several eukaryal-like features not found in other archaea have been detected in Thaumarchaeota (Brochier-Armanet et al., 2008a,b; Spang et al., 2010). PVC bacteria and Thaumarchaeota have not been previously suggested as potential fusion partners for the origin of Eukarya. Before someone else thus proposes a new fusion scenario involving these two groups, I decided to do it myself. In this new scenario, FECA originated from the engulfment of a thaumarchaeon (the symbiont) by a PVC bacterium (Fig. 2). This fusion was followed by the massive invasion of the FECA descendants by several novel lineages of viruses, such as NCLDV (Nucleo-Cytoplasmic Large DNA Viruses) and retroviruses (Fig. 2). Integration of viral genomes into the genomes of FECA descendants introduced new Fig. 2. The PTV fusion hypothesis based on the engulfment of a thaumarchaeon by a PVC bacterium followed by viral invasions. Bacterial and archaeal membranes, cytoplasmic and nuclear components (including circular chromosome) are in green and purple, respectively. Eukaryal cytoplasmic and nuclear components are in grey to symbolize differences with their archaeal ancestors. Eukaryal chromosomes (linear) are in orange. Abbreviations are as in Fig. 1. ICM: Intracytoplasmic membrane. NE: nuclear envelope. PVC: Planctomycetes, Verrucomicrobia, Chlamydiae superphylum. For simplicity the reticulum endoplasmic membrane deriving from the ICM of the PVC bacterium has not been indicated. 80 P. Forterre / Research in Microbiology 162 (2011) 77e91 proteins (and protein folds) into ancient eukaryal lineages, explaining the presence in modern Eukarya of many proteins without homologues in Archaea and Bacteria. Furthermore, intense competition between viruses and ancient Eukarya produced an arms race that favoured evolution toward increasing complexity from FECA to LECA. For instance, the invasion by retroviruses might have triggered the transformation of circular chromosomes into linear ones and initiated the succession of events at the origin of sex. This new hypothesis will be dubbed the PTV scenario thereafter (for PVC bacteria eThaumarchaea - Virus). I choose a bacterial host because it readily explains why the phospholipids of eukaryal cell membranes (the cytoplasmic membrane, the endoplasmic reticulum and the nuclear membrane) are made of bacterial type phospholipids (esters of sn-glycerol-3-phosphate and fatty acid) (green in Fig. 2). Hypotheses in which the host is an archaeon should explain why archaeal phospholipids (ethers of sn-glycerol-1-phosphate and isoprenol) were replaced by bacterial ones in all these membranes. This is really impossible without spurious ad hoc assumptions. Although unlikely (see below), it seems easier to postulate that the internal membrane of the endosymbiont (the archaeon) disappeared after the fusion, than to assume (if the symbiont was a bacterium) that the bacterial lipids of the symbiont replaced those of the host! I will thereafter explain why, although this new proposal is probably the best possible fusion scenario that one can imagine with our present knowledge, I still don’t believe in it. Finally, I will discuss how some novel aspects of this scenario (in particular the intervention of viruses) could be nevertheless useful for imaging how Eukarya originated and evolved from the Last Universal Cellular Ancestor (LUCA) in the framework of the three primary domains concept of Carl Woese. 2. A new fusion scenario: a PVC bacterium as the host and a thaumarchaeon as the symbiont 2.1. Why a PVC bacterium The PTV scenario posits that the host bacterium at the origin of Eukarya was evolutionarily related to modern bacteria of the superphylum PVC because genes encoding structural analogues of eukaryal MC proteins, such as clathrins and nucleoporins, have been recently detected in the genomes of several Planctomycetes, Verrucomicrobia and Lentisphaera (Santarella et al., 2010). These proteins exhibit a unique structure formed by an amino-terminal beta-propeller domain followed by a carboxy-terminal stacked pairs of alpha-helices. The respective length of the two domains, as well as the respective number of beta strands and alpha helices in each domains are strikingly similar in PVC bacteria and Eukarya, strongly suggesting that MC proteins of PVC bacteria and Eukarya are homologous (5). Similar proteins have never been found in Archaea or in Bacteria other than PVC bacteria. In Eukarya, MC proteins are involved in the formation of the endoplasmic reticulum (e.g. clatherins and related proteins) and of the nuclear pores (e.g. nucleoporins). Strikingly, PVC bacteria encoding eukaryal-like MC proteins exhibit an intracytoplasmic membrane (ICM) mimicking the endoplasmic reticulum of Eukarya (Fuerst, 2005; Lee et al., 2009; Santarella et al., 2010). In Eukarya, the endoplasmic reticulum membrane is recruited to form the double membrane that surrounds the chromosomes (the nuclear envelope). Similarly, in the Planctomycete Gemmata obscuriglobus, a synkaryotic bacterium, the ICM is recruited to form a double membrane that surrounds the bacterial nucleoid (Fuerst, 2005). Recently, it has been shown that G. obscuriglobus can perform endocytosis, a property that was until now thought to be restricted to Eukarya, and one of its MC proteins seems to be involved in that process (Lonhienne et al., 2010). PVC Bacteria thus appear the ideal hosts for a fusion hypothesis, since they could explain the emergence in Eukarya of both the endoplasmic reticulum and the nuclear envelope. Beside the presence of eukaryal-like MC proteins, PVC bacteria exhibit several additional features that favour them as putative hosts in fusion hypotheses for the origin of Eukarya. In particular, synkaryotic PVC bacteria (G. obscuriglobus and Poribacteria) are among the few bacteria in which sterol biosynthesis has been documented (Pearson et al., 2003; Siegl et al., in press). Otherwise, sterol biosynthesis is almost exclusively restricted to Eukarya. Plancomycetes are quite large cells that divide by budding, a feature that reminds us of Eukarya (in fact, Plancomycetes were once confused with fungi, hence their name) (Fuerts, 1995). Similar to Eukarya, some PVC bacteria also lack peptidoglycan and the bacterial cell division protein FtsZ (Pilhofer et al., 2008). This favours PVC bacteria over other bacteria as potential hosts able to engulf small cellular symbionts. All known modern PVC bacteria have cell envelope and cannot perform phagocytosis. However, one can imagine that, building on their ability to perform endocytosis, some ancient or not yet discovered wallless PVC bacteria (L-forms) have used their MC protein to evolved the ability to engulf small preys. Importantly, some Planctomycetales relatives (anammox bacteria) contain an intracellular organelle, the anammoxosome, surrounded by a membrane formed by atypical lipids (ladderanes) (Jetten et al., 2009). The membrane of the anammoxosome contains an FATP synthase, mimicking the cytoplasmic membrane of a bona fide organisms (Van Niftrik et al., 2010). The origin of the anammoxosome is unknown since this organelle lacks DNA. However, in the framework of the present hypothesis, it is tempting to suggest that some ancestor of anammox bacteria have had the capacity to serve as host for other microbes. Finally, closely related homologues of the two eukaryal tubulins (alpha and beta) have been found in bacteria of the genus Prosthecobacter, which are members of Verrucomicrobia (Jenkins et al., 2002). Interestingly, the alpha and beta tubulins of Prosthecobacter do not branch within Eukarya in a tubulin phylogenetic tree, but as outgroups of eukaryal alpha and beta eukaryotic tubulins, respectively, arguing against a lateral gene transfer from a modern Eukarya to this group of PVC bacteria (Jenkins et al., 2002). In the PTV hypothesis, the two tubulin genes originated first in PVC bacteria and were P. Forterre / Research in Microbiology 162 (2011) 77e91 later on transferred to Eukarya via the bacterial partner of the fusion. In summary, in the PTV scenario, the PVC bacterium provided to the future Eukarya not only membrane phospholipids (as in other fusion scenarios) but also the ICM membrane, tubulin and MC proteins (precursors of the cytoskeleton, nucleoproteins and of the nuclear pore complex), as well as the ability to synthesize sterols, a universal component of eukaryal membranes (Table 1). 2.2. Why a thaumarchaeon as the symbiont? In the PTV scenario, the archaeal symbiont was a thaumarchaeon because these archaea exhibit a wider range of eukaryal features than archaea from the two other phyla, Euryarchaeota and Crenarchaeota (Brochier-Armanet et al., 2008a,b; Spang et al., 2010) (Table 1). Hence, whereas the large RNA polymerase A subunit is splitted in Crenarchaea and Euryarchaea, it corresponds to a single polypeptide in Thaumarchaea, as in Eukarya and Bacteria (Kwapisz et al., 2008). Thaumarchaea are also the only Archaea to harbour a type IB DNA topoisomerase, the enzyme responsible in Eukarya to relax positive supeturns that accumulate during DNA replication and transcription (Brochier-Armanet et al., 2008b). They are also the only Archaea that contain the two single-stranded DNA binding proteins (SSB) found in Eukarya, the protein RPA, previously thought to be restricted to Euryarchaea, and the small SSB recently found in human (hSSB), previously thought to be restricted to Crenarchaeota (unpublished observation). Thaumarchaea harbour eukaryal-like histones, otherwise mainly present in Euryarchaeota (Cubonová et al., 2005), and the two eukaryotic-like “cell division” proteins (CdvB/Snf7 Table 1 Molecular features introduced in Eukarya by the thaumarchaeon, the PVC bacterium and viruses, respectively, in the framework of the PTV scenario. Molecular features either present in Thaumarchaea but not in other archaeal phyla or present in some PVC bacteria but not in other bacterial phyla are in bold. Thaumarchaeon PVC bacterium Viruses Translation machinery (ribosomes) Transcription machinery Non split RpoA Transcriptional regulators mRNA capping New basal transcription factors Spliceosomes (?) New DNA polymerases Topo IIA Linear chromosomes Telomers, Centromers Sex Replication machinery, histones RPA D cren SSB Topo IB Bacterial lipids ESCRT proteins Membrane Coat V-ATP ase, Sec system, Proteins SRP Intracytoplasmic membrane (ICM) Sterols Actin Tubulin New proteins to manipulate membranes previously involved in formation of viral factories New cytoskeletal proteins of low-complexity sequence composition 81 and CdvC/Vps4), otherwise mainly present in Crenarchaeota (for review, see Makarova et al., 2010). In Eukarya, the proteins CdvB/Snf7 and CdvC/Vps4 belong to the ESCRT complex (Endosomal Sorting Complex Required for Transport) that is involved in vesicles formation and cell division. Importantly, the archaeal proteins seem to be involved in similar mechanisms (Samson and Bell, 2009; Ettema and Bernander, 2009). In the PTV scenario, I will suppose that the thaumarchaeon at the origin of Eukarya also contained the archaeal actin protein (Arch-actin) recently detected in Thermoproteales (members of the phylum Crenarchaeaota) and in the only Korarchaeon whose genome has been sequenced, "Candidatus Korarchaeum cryptofilum" (Yutin et al., 2009). The Arch-actin is much more similar to eukaryal actin than other actin homologues previously found in Bacteria or Archaea, MreB and FtsA, respectively (for review, see Makarova et al., 2010). Although the three presently Thaumarchaeota whose genomes have been sequenced lack the Arch-actin gene, genome context analysis suggests that this gene was present in the Last Archaeal Common Ancestor (LACA) and was later on lost in the branch leading to the three Thaumarchaea whose genomes have been sequenced (Makarova et al., 2010). In the PTV scenario, a major consequence of the fusion was thus to bring for the first time actin (from the thaumarchaeon) and tubulin (from the PVC bacterium) in the same organism and later on (once the archaeal membrane has disappeared) in the same cytoplasm. In previously proposed fusion hypotheses, one has to assume a dramatic acceleration of the rate of protein evolution after the fusion to explain why are tubulin and actin now so different in Eukarya from their distant homologues in Bacteria and Archaea, FtsZ and MreB proteins, respectively (Doolittle, 1995; Erickson, 2007). This is especially difficult to admit if the bacterial partner was the alpha-proteobacterium at the origin of the mitochondrion (as in the hypothesis of Martin and Muller). Why did the FtsZ of the alpha-proteobacterium evolved beyond recognition (at the sequence level) from its bacterial ancestor, whereas many other proteins from the same alpha-proteobacterium remain so similar in Bacteria and mitochondria. To take a single example, the mitochodrial protein Qri7 branches as sister group of the alpha-proteobacterial YgjD (its ancestor) in phylogenetic trees, and the mitochondrial protein can complement a YgjD bacterial mutant (Oberto et al., 2009). The dramatic transformation of FtsZ and MreB proteins into tubulin and actin was therefore a major drawback of previous fusion scenarios. In contrast, in the PTV scenario, eukaryal tubulin and actin did not originate from bacterial FtsZ and MreB proteins, but directly from archaeal actin and bacterial tubulin, with sequences already very similar to those of modern Eukarya. In addition to exhibit a unique combination of eukaryoticlike features not found in other archaeal phylums, Thaumarchaea are also good candidates to be the archaeal symbiont in a fusion scenario because they are abundant in marine, lacustre and terrestrial mesophilic environments where they coexist with bacteria, including PVC bacteria. For instance, Cenarchaeum symbiosum (a thaumarchaeon) and Poribacteria (PVC bacteria) live as intracellular symbionts within sponges. Of course, one cannot argue that a bacterium and an archaeon 82 P. Forterre / Research in Microbiology 162 (2011) 77e91 meet in a sponge to produce the ancestor of sponges! Nevertheless, many possibilities remain open. I will not try to propose here a specific type of metabolism complementation to justify the PTV scenario. Indeed, both Thaumarchaea and PVC bacteria are very diverse but poorly studied groups that probably include many types of metabolisms. For instance, the capacity of some thaumarchaea to oxidize ammonium has only been discovered five years ago (Treusch et al., 2005). I thus assume that Thaumarchaea and PVC Bacteria with complementary metabolisms will be discovered soon or later to justify their association in the PTV scenario. It will then be possible to imagine the specific type of “fusion” suitable to associate these complementary partners and the type of environment in which they meet each other. In summary, in the PTV scenario, the thaumarchaeon provided to the future Eukarya all proteins and machineries that remain highly similar today between Archaea and modern Eukarya. These proteins and mechanisms (e.g. ribosomes, DNA replication and transcription apparatus) are typically bacterial in PVC bacteria, indicating that Eukarya cannot be derived directly from a PVC bacterium, even if the latter is a synkaryote. Proteins provided by the thaumarchaeon included not only informational proteins but also operational proteins, such as some precursors of the modern eukaryal cytoskeleton (actin) and vesicle trafficking systems (ESCRT proteins) (Table 1). 2.3. Origins of mitochondria In the PTV scenario, the endosymbiotic event that produced mitochondria occurred obviously after the fusion event that produced the FECA (Fig. 2). At first sight, fusion hypotheses in which the bacterial partner became the mitochondrion (arrow b in Fig. 1B) seems more parsimonious because they have no need to postulate the existence of ancient Eukarya without mitochondria (Archezoa, sensu Cavalier-Smith, 1989). For some authors, this explains the failure to discover such Archezoa today (living fossils). However, it is crystal clear that ancient Eukarya should have existed, whatever the scenario we prefer for the origin of Eukarya. For instance, reconstruction of the mitochondrion present in the LECA clearly indicates that mitochondria have evolved a long way between the alphaproteobacterium present in the first mitochondrial Eukarya (FME) and the mitochondrion present in LECA. Indeed, phylogenomic analysis shown that the mitochondrial ribosome of LECA has already gained 19 novel proteins that were not present in its bacterial ancestor (Desmond et al., 2011). Therefore, even if the FECA and the FME were the same organism, one has to postulate the existence of many intermediate forms of ancient Eukarya which had mitochondrial ribosomes with protein compositions intermediate between those of the FECA/FME and the LECA (Fig. 1B). Furthermore, in any scenario, the emergence of specific eukaryal features already present in LECA such as the nucleus, the cytoskeleton, spliceosomes, or the mitotic apparatus, should have spanned a long evolutionary period (Gribaldo et al., 2010). For example, it is quite clear that meiosis evolved from mitosis through a complex evolutionary pathway involving several successive steps (Wilkins and Holliday, 2009). Ancient Eukarya (urkaryotes, sensu Woese and Fox, 1977) have thus certainly existed for a long period of time (co-existing with Archaea and Bacteria) Fig. 3. The three possible (but unlikely) hypotheses for the origin of the eukaryal nuclear envelope in the PTV scenario. A: the eukaryal nuclear envelope derived directly from the nucleus of a synkaryotic PVC bacterium, such as Gemmata obscuriglobus. B, the eukaryal nuclear envelope derived from the ICM of an akaryotic PVC bacterium. C: the eukaryal nuclear envelope derived directly from the membrane of the thaumarchaeon. Membranes with bacterial type and archaeal type phospholipids are in green and purple, respectively. Bacterial and archaeal cytoplasmic components and chromosomes are in green and purple, respectively. Colour shift from green to purple symbolizes the replacement of bacterial components (e.g. ribosomes) by archaeal ones. In the three scenarios, FECA is transformed into a proto-eukarya before the mitochondrial endosymbiosis, and not yet remodelled by extensive viral invasion. For simplicity the reticulum endoplasmic membrane deriving from the ICM of the PVC bacterium (A, B) or from the modified envelope of the thaumarchaeon (C) has not been indicated. Abbreviations are as in Figs. 1 and 2. P. Forterre / Research in Microbiology 162 (2011) 77e91 and have diversified in many lineages that have all disappeared today (much like pre-Homo sapiens during Homo evolution) (Fig. 3). The existence of extinct lineages is indeed a consequence of biological evolution already discussed by Darwin in the “origin of species”. 3. The viral hypothesis for the origin of eukaryal complexity The existence of numerous complex and specific molecular features in Eukarya (either specific protein families or specific molecular mechanisms) has always been a major challenge to proponents of fusion hypotheses (Koonin, 2010). In the most recent reconstitution of the putative proteome of LECA (performed after the sequencing of the protist Naegleria gruberi) Fritz-Laylin and colleagues concluded that at least 40% of proteins in LECA e 1420 - were “eukaryotic inventions” (they have no detectable archaeal or bacterial homologues) (FritzLaylin et al., 2010). In the PTV scenario, some eukaryal specific proteins might correspond to thaumarchaeal or PVC bacterial proteins that have been lost in the few genomes of PVC bacteria or Thaumarchaea presently available. As a matter of fact, the archaeal orthologues of actin and Topo IB have been only recently detected in modern archaeal genomes. Further archaeal or bacterial homologues of eukaryal specific proteins will be also probably detected in the future at the structural level, as it was recently the case for MC proteins of PVC bacteria. However, it seems unlikely that “prokaryotic” homologues will be finally found for all eukaryal specific protein families and protein folds. Indeed, even before the sequencing of N. gruberi, Kurland and colleagues identified in the universe of protein folds 244 fold superfamilies that are specific to Eukarya, versus 16 and 49 that are specific for either Archaea or Bacteria, respectively (Kurland et al., 2007). Furthermore, it is unlikely that we will find in the future eukaryotic specific mechanisms, such as mRNA capping and splicing, telomers, meiotic sex, mid-bodies and so on, in some PVC bacteria or Thaumarchaeota. The creation of all these eukaryal novelties has been a major puzzle for all fusion hypotheses. The only possibility to bypass this problem is to suggest that the fusion triggered an explosion of creativity, leading to a dramatic increase in complexity of the resulting entity. However, there is no known example in current biology of such situation. Usually, a combination of organisms led to the reductive evolution of one of them, but never to an explosion of creativity and drastic modifications in the molecular make up of the other. For instance, the “fusion” (endosymbiosis) between Eukarya and Cyanobacteria did not change the basic molecular biology of Eukarya, although it triggered the emergence of a new eukaryal division (Viridiplantae). In the framework of the PTV scenario, I suggest here a new hypothesis that could help to solve this conundrum. I propose that eukaryal specific protein families and mechanisms originated as the consequence of several massive viral invasions that pecifically shaped the evolution of the eukaryal lineage from FECA to LECA. In this hypothesis, multiple rounds of integration of viral genomes into the genomes of ancient 83 Eukarya introduced new proteins (and protein folds) into the descendants of FECA, whereas the intense competition between viruses and ancient Eukarya produced an arms race that favoured evolution toward increasing complexity from FECA to LECA. What makes plausible the idea that repeated viral invasions introduced in ancestral Eukarya new protein families without archaeal or bacterial homologues is that most proteins encoded by viral genomes today have no cellular homologues in protein databases (up to 100% in the case of some archaeal viruses) (Prangishvili et al., 2006b; Yin and Fischer, 2008; Boyer et al., 2010) and that most viral proteins whose structure has been solved harbour unique folds (Bamford et al., 2005; Saini and Fisher, 2007, for a recent case studies, see Keller et al., 2009). Genes are indeed created continuously de novo in viral genomes within the infected cells (virocells, sensu Forterre, 2010b) by the same processes that create new genes in cellular genomes (gene duplication, gene slippage, overprinting and so on) (Rancurel et al., 2009; Forterre, 2010b). In the PTV scenario. I suggest that ancestors of modern NCLDV most likely played a major role in the introduction of new protein families in Eukarya. Ogata and Claverie indeed concluded from an analysis of proteins families specific for various lineages of NCLDV that these viruses with large genomes (between 150 and 1200 kb) act indeed as “an invention factory for new genes” (Ogata and Claverie, 2007). These viruses are very ancients, as the diverse NCLDV families probably diverged even before LECA (Koonin and Yutin, 2010). Recently, large portions of an NCLDV genome were found integrated in the genome of the Brown algae Ectocarpus siliculosus, indicating that NCLDV specific genes can become incorporated into eukaryal genomes, becoming “eukaryal specific genes” (Cock et al., 2010). The genomes of modern NCLDV carry a relatively high percentage of genes with eukaryal homologues (10e20%) that are usually supposed to be derived from eukaryal genomes (Moreira and López-Garcı́a, 2009). In fact, many of them have probably originated first in NCLDV and were later on recruited by Eukarya (discussed in Forterre, 2010a). Interestingly, Ogata and Claverie noticed that NCLDV family-specific proteins are enriched in lowcomplexity sequences, resembling eukaryal proteins involved in nucleic acid-interactions and in the architecture of the cytoskeleton (Ogata and Claverie, 2007). Proteins with lowcomplexity sequences could originate more frequently in viruses since mistakes such as replication slippage or abnormal recombination probably occur more frequently in viral genomes than in cellular genomes. I thus suggested previously that many eukaryal proteins with low-complexity sequences involved in interactions with nucleic acids or in cytoskeleton architecture first originated in NCLDV and were later on introduced in Eukarya (Forterre, 2010a). Beside NCLDV, other viruses such as Retroviridae, Adenoviridaea, Herpesviridae, Baculoviridaea or still other not yet discovered viral lineages (for instance among viruses infecting protists that have been very poorly screened) could have been contributed to the introduction of new proteins in Eukarya. 84 P. Forterre / Research in Microbiology 162 (2011) 77e91 In the PTV scenario, viral invasions not only provide new protein families to ancient Eukarya but also induced a continuous arms race between viruses and eukaryal cells that dramatically altered the outcome of eukaryal evolution; this arms race leading to the emergence of more and more complex organisms, with both the creation of entirely novel molecular mechanisms (e.g. mRNA capping, see below) and improvement of ancient already existing ones (e.g. transcription). As an example, let’s see how the viral invasion might have triggered the formation of the odd cap structures that tag in 50 eukaryotic messenger RNA (unknown in Archaea and Bacteria). NCLDVencode homologues of the eukaryal enzymes responsible for the formation of this structure (Shuman, 1995). Imagine that an ancient NCLDV first introduced a simple tag, “A”, into the 50 end of its mRNA in order to degrade selectively cellular mRNA (the tag providing protection of mRNA of the virus against its own nuclease). As a result, cells will be later on selected that have aquired the ability to label their own mRNA with the viral tag A. In response, viral mutants producing tag of increasing complexity (AB) will then be selected. This will later favour mutant cells having aquired this “AB” tag, leading to the appearance of viruses with even more complex tag (ABC) and so on. A strong argument in favour of such viral scenario for the origin of mRNA tags is that two different types of complex mRNA tags have now been documented in viruses (Koonin and Moss, 2010; Ogino et al., 2010), whereas only one of those exists in cellular mRNA. The type of scenario suggested here to explain the origin of the complex eukaryal mRNA capping system can be readily applied to other systems, explaining why many molecular mechanisms become so sophisticated in Eukarya. The spliceosome and the various layers (mediator and so on) of the eukaryal transcription apparatus, including its regulation, are hence prime players in cellvirus molecular interactions. If the PVC bacterium partner in the fusion was an akaryore (with no nucleus, but simply an ICM), viruses might have also play a major role in the emergence of the eukaryal nucleus. Indeed, many DNA and RNA viruses that replicate in the cytoplasm of eukaryal cells recruit the reticulum endoplasmic membrane of their hosts (victims) to build their viral factories, much like Eukarya recruit the endoplasmic reticulum membrane to form their nucleus. Among them, Mimivirus (an NCLDV) produces viral factories that are as big as the nucleus of an Amoeba (Suzan-Monti et al., 2007). One can therefore imagine that proteins used by one such virus to produce viral factories in the PVC bacterium were recruited by an ancient eukaryal cell to build its nucleus from the PVC ICM. The viral factory is built as to protect the viral genome against the defences of the host. Similarly, the nucleus might have been first built to protect the cellular genome against invading viruses (Forterre and Prangishvili, 2009b; Bandea, 2009). A major feature that distinguishes modern Eukarya from Archaea and Bacteria is the structure of their chromosomes. In the PTV scenario, retroviruses have probably played a crucial role in transforming the circular chromosome(s) of FECA into the linear chromosome of modern eukaryal cells. Indeed, both telomeres and centromeres appear to be related to retroviruses. Telomerase is homologous to reverse transcriptase (Arkhipova, 2006) and it is intriguing that Penelope retroelements containing telomeric-like sequences directly integrate at chromosome ends (Gladyshev and Arkhipova, 2007), whereas centromeres are rich in repeated sequences ultimately of retroviral origin (Chueh et al., 2009). Retroviruses and derived elements might have also been major players in the origin of sex in Eukarya, and, more specifically, in the evolution from mitosis to meiosis. Wilkins and Holliday (2009) have recently suggested that the first step in the evolution of meiosis was the emergence of a mechanism to pair homologous chromosomes. In their hypothesis, this mechanism was selected to prevent potentially damaging recombinations between repeated regions present in non homologous chromosomes. If the chromosomes of ancient Eukarya have been invaded by retroviruses, integrated retroviruses have been most likely a major source of repeated sequences in their chromosomes. In that case, retroviruses and retroelements might have provided the selection pressure for the evolution of meiosis from mitosis by the mechanism proposed by Wilkins and Holliday. The introduction of viruses in the PTV scenario could a priori solve another major problem (at least partly see below) that plague all fusion hypotheses: why eukaryal proteins of archaeal origin experienced such a dramatic acceleration of their evolutionary rate after the fusion, such that most of them do not branch within Archaea in phylogenetic tree, but as sister group of Archaea (Yutin et al., 2008, see discussion in Gribaldo et al., 2010). Proponents of fusion hypotheses usually argue that this acceleration was induced by the fusion itself that led to a very peculiar chimera. However, as already discussed in the case of the mirochondrial protein Qri7 and its bacterial homologue YgjD, actual fusion between Eukarya and Bacteria did not induce such acceleration of the rate of protein evolution. For another example, the DNA gyrase, of bacterial origin, which is today encoded in the nuclear genome of plants and is targeted to both chloroplats and mitochondria, still branches within Bacteria, as sister group of Cyanobacteria in the phylogenetic tree of type II DNA topoisomerases (Forterre et al., 2007). This protein has therefore evolved very slowly after the endosymbiosis event that led to the chloroplast, despite a complete modification of its cellular environment and likely modification and diversification of its function. Thus, evolutionary rate acceleration is not an inevitable outcome of endosymbiotic fusion events. In the PTV scenario, one can imagine that most eukaryal genes have shuttled (after the fusion) between the genomes of ancient Eukarya and those of their invading viruses. During their passage in viral genomes, these genes would have evolved at a more rapid pace than their archaeal sisters that remained most of the time confined in cellular genomes. We have recently observed such a phenomenon in studying the evolution of the replicative helicase MCM in Archaea (Krupovic et al., 2010). An MCM protein of cellular origin that had transited by viral genomes in Methanococci evolved much more rapidly than its orthologues in other archaeal lineages. A similar scenario could explain, for example, why some eukaryal DNA replication P. Forterre / Research in Microbiology 162 (2011) 77e91 proteins diverged so much from their archaeal ancestors, since DNA replication proteins appear to be frequently captured by viruses. This is for instance the case of some eukaryal cellular DNA polymerases that are much more related to DNA polymerases of NCLDV than to archaeal DNA polymerases (Villarreal and DeFilippis, 2000; Filée et al., 2002). The importance of viruses in the PTV scenario is in line with the recent recognition of their major role in the evolution of life on our planet (Forterre, 2005, 2006; Comeau and Krish, 2005; Forterre and Prangishvili, 2009a,b, Brüssow, 2009; Bandea, 2009; Cortez et al., 2009; Villarreal and Witzany, 2010). In the PTV scenario, the evolution of Eukarya toward complexity from FECA to LECA occurred by combining the power of viral creativity (import of new proteins) and selection pressure (arms race type of evolution) (Comeau and Krish, 2005; Forterre and Prangishvili, 2009b). Importantly, the role of viruses in this scenario satisfies the principle of continuity, being consistent with their roles in recent eukaryotic evolution. Viruses have been indeed essential in the creation of novelties such as the unique replication and transcription system of mitochondria (Filée and Forterre, 2005) or else the formation of the placenta in Mammals (Rote et al., 2004). 4. Why the best possible fusion hypothesis is probably wrong The PTV scenario solves several questions raised by fusion hypotheses in general, and proposes much more convincing “prokaryotic” partners than previous hypotheses. However, I don’t think that this scenario can finally save fusion hypotheses from being fundamentally flawed, and I will discuss now why. Firstly, despite the presence of a nucleus in some PVC bacteria, it remains difficult to explain the origin of the eukaryal nucleus, since all molecular mechanisms working today within the eukaryal nucleus are much more similar to their archaeal than to their bacterial countrparts. In the PTV scenario, the eukaryal nucleus might be either a direct descendant of a nucleus already present in the host PVC bacterium (a synkaryote) or it might have originated after the fusion, from the ICM of the PVC bacterium (an akaryote) (Fig. 3A). The first hypothesis seems a priori more parsimonious since it implies that the “invention” of the nucleus occurred only once in life history (in PVC bacteria) and that the fusion directly produced a FECA with a nucleus (Fig. 3A). However, this hypothesis is difficult to reconcile with the fact that the DNA replication and transcription machineries, as well as the chromatin structure (histones) of modern Eukarya should have originated from the archaeal symbiont. If the eukaryal nucleus derived directly from the nucleus of the PVC bacterium host, one should thus assume that all informational genes of bacterial origins were replaced in situ (in the bacterial chromosome) by archaeal genes encoding similar function recruited from the chromosome of the thaumarchaeon. There is no obvious selection pressure to explain this highly unusual event. Furthermore, the thaumarchaeon (especially its membrane) should have completely disappeared in the process (Fig. 3). Massive transfer 85 of bacterial genes into the eukaryal nucleus have occurred after the endosymbiosis event at the origin of mitochondria, but they have not displaced their eukaryal functional analogues. Remnants of the bacterial genome remain in most mitochondria and Remnants of the bacterial membrane is still present in all Eukarya (either as mitochondrial or hydrogenosome membranes). The second possibility is to imagine that the eukaryal nucleus originated from the ICM of an akaryotic PVC bacterium after the formation of FECA (Fig. 3B). Again, this involves a complex and unlikely series of events. In a first step, the archaeal symbiont should lost its membrane, whereas in a second step, the archaeal chromosome became enclosed by a new (double) membrane formed via invagination of the ICM. During that process, most of the genome of the PVC bacterium should have been lost, except for some genes that were transferred to the newly formed nuclear genome of archaeal origin. The latter should have included in particular the genes for bacterial lipid biosynthesis and those for tubulin and MC proteins. In both scenarios, the disappearance of the thaumarchaeal membrane is especially difficult to imagine. The complete disappearance of the membrane of the symbiont has never been observed in actual symbiosis (for instance mitochondria and chloroplasts have retained the two membranes of their bacterial ancestors) and cannot be justified easily (except by the fact that we have to explain the present situation!). The disappearance of the archaeal membrane is especially strange considering that Eukarya have only conserved the archaeal version of all universal proteins working at the membrane, such as the V-ATPase, the Sec core proteins, or else the signal recognition ribonucleo-particle that target to the membrane ribosomes synthesizing membrane proteins. The Eukarya also retain archaeal specific machinery, such as the ESCRT proteins, to work now with a bacterial membrane. A third possibility is to imagine that the eukaryal chromosome evolved directly from the archaeal one that remained in the thaumarchaeon (Fig. 3C). This implies that the archaeal membrane was transformed in situ into a bacterial one. In that case, one should imagine that the archaeal genes for lipid biosynthesis were specifically replaced in situ (in the archaeal chromosome) by genes for bacterial lipid biosynthesis coming from the PVC bacterium. Again, there was no obvious selection pressure for this replacement. For that very reason, I previously refuted in the introduction the possibility that the thaumarchaeon was the host in the fusion. Furthermore, in that third scenario, one has now to explain the disappearance of the bacterial chromosome. Of course, considering the role of viruses in our hypothesis, one can still imagine that a virulent virus originally infecting a PVC bacterium find its way into a descendant of FECA and specifically destroy the bacterial chromosome (the archaeal one being protected by the archaeal membrane or the proto-nuclear envelope or else by specific chemical modifications). One should then assume that this destruction only occurred after some critical bacterial genes had already been transferred to the archaeal nucleus (i.e. genes encoding CM proteins and proteins for the biosynthesis of bacterial phospholipids). This again sounds as an ad hoc 86 P. Forterre / Research in Microbiology 162 (2011) 77e91 assumption. Two more problems plague this third scenario, firstly, the endoplasmic reticulum originated now after the nucleus, as an extension of the nuclear envelope. This is in contradiction with sound logic, the nuclear envelope being instead clearly an extension of the endoplasmic reticulum (Jékely, 2003, 2007; DeDuve, 2007). Secondly, the ICM of the PVC bacterium has no more any role in the origin of the eukaryal nucleus, which is in contradiction with the original idea behind the PTV scenario. In summary, the transformation of a FECA born from a fusion event into a reasonable ancestor of modern eukarya remains difficult to imagine in a convincing way in the PTV scenario as in all other fusion scenarios. Another major problem of all fusion hypotheses (rarely discussed) is the origin of eukaryal viruses. This problem is of course central in the PTV scenario since the descendants of FECA are supposed to be invaded by specific lineages of eukaryal viruses. Where did they come from? In any fusion scenario, eukaryal viruses should have originated from a mixture of archaeal and bacterial viruses (Koonin et al., 2006), in the PTV scenario, eukaryal viruses should have specifically evolved mostly from bacterial ones, since FECA originally only contained receptors for bacterial viruses at its cell surface. The only archaeal viruses that might have contributed were those that were integrated as “proviruses” in the thaumarchaeal symbiont. Some eukaryal and bacterial viruses harbour indeed major capsid proteins (MCPs) with similar folds. This is the case for Herpes viruses and “head and tails” bacterioviruses whose MCPs harbour the Hong-Kong fold, or else for NCLDV, Adenovirus and Tectiviridae (bacterioviruses) whose MCPs harbour the double jelly roll fold (Bamford, 2003; Bamford et al., 2007). The replicases (RNA dependent RNA polymerase) of some eukaryal and bacterial RNA viruses or else the proteinprimed DNA polymerases encoded by different viruses infecting either Bacteria or Eukarya are also homologous between the two domains (Filée et al., 2002,Kidmose et al., 2010). However, despite these homologous molecular features, eukaryal viruses in general differ dramatically of their bacterial counterparts (and also of their archaeal ones) both in terms of virion architecture and genome content. Since viruses infecting modern Eukarya were probably already there at the time of LECA, one really wonders why the fusion of an archaeon and a bacterium would have so rapidly triggered the evolution of “head and tailed” viruses into Herpes viruses, or else the evolution of Tectiviridaea into NCLDV? More generally, considering the high number and great diversity of eukaryal virus families, how can we imagine that the PVC bacterium and the Thaumarchaeon at the origin of the Eukarya were bona fide targets for ancestors of all these viral families? Indeed, in the PTV scenario, one should imagine that ancestors of all modern eukaryal viruses were able to infect PVC bacteria (they should have used as hosts the early descendants of FECA) or were integrated as proviruses into the genome of the Thaumarchaeon. If this is the case, the descendants of these viruses should be still there today infecting modern PVC bacteria and Thaumarchaea. Of course, one can postulate the existence of many unknown viruses infecting PCV Bacteria and/or Thaumarchaea that resemble NCLDV, retroviruses (not retroelements, but bona fide retroviruses with gag, pol, env genes) and all other viruses specific for Eukarya. However, their presence has not yet been detected in the genomes of PCV Bacteria and Thaumarchaea that are already available. I predict that these eukaryal-like viruses will not be found, and that the difficulty to imagine the transmutation of archaeal and bacterial viruses into eukaryotic ones will remain a very weak point of all fusion hypotheses. Finally, and this is the critical point, one has to explain why they are today three and not two versions of ribosomes on our planet. Fusion hypotheses in fact tend to roll history backward by fundamentally denying the critical advance in our understanding of life evolution that has been brought about by the development of molecular biology in the middle of the last century. The discovery of three distinct deep evolutionary lineages of organisms based on rRNAwas the key discovery that led to formulate the three domains concept (Woese and Fox, 1977). This observation remains the major challenge to any kind of fusion hypotheses. In the present scenario, it is indeed hard to explain why the ribosomes of the bacterial host completely disappeared after the fusion and why the archaeal ribosomes of the symbiont experienced such an extreme acceleration of their evolutionary rate in the eukaryotic lineage, becoming sufficiently divergent from those of their archaeal ancestor to be recognized as a different version (sensu Woese). I have suggested previously in this paper that protein shuttle between the genomes of ancient Eukarya and viruses infecting them could have explained the accelerated evolution of some proteins in Eukarya. However, this cannot be applied in the case of ribosomal proteins since, by definition, viral genomes do not encode ribosomes (Raoult and Forterre, 2008). The nature of the eukaryal ribosome thus remains central to discuss the validity of fusion hypotheses. Proponents of fusion hypotheses will argue that the rate of evolution of ribosomal proteins in Eukarya has been dramatically increased by the addition of new proteins and rRNA segments during the transition from the FECA to the LECA. However, addition of new proteins and rRNA segments should have instead introduce more constraints (from proteineprotein and protein-RNA interactions) to the evolution of eukaryotic ribosomal RNAs and proteins, slowing down their rate of divergence instead to produce an acceleration. A particular aspect of all fusion hypotheses that make me suspicious is that they all invoke some kind of miracle that occurred only once in life evolution (Koonin, 2007), although the condition for its appearance have been there for the last 3 billion years (the coexistence of some Archaea and Bacteria in the same biotope). If such fusion was possible once, it is not clear why many eukaryal-like lineages did not emerge continuously during the history of our planet leading to the emergence of multiple independent (polyphyletic) eukaryal (or eukaryal-like) groups. One cannot argue that these lineages have been all eliminated by the descendants of LECA, because new eukaryal-like lineages should still appeared today. The only possibility to solve this problem is to argue that, for some reason, the fusion was only possible in environmental conditions that do not exist anymore on our planet. However, P. Forterre / Research in Microbiology 162 (2011) 77e91 in that case, one should abandon any hope to find the actual partners and, as a consequence, to understand the reasons which selected for the fusion event itself. Again, it seems that we want Archaea and Bacteria to associate only because we need an explanation for our own presence in this world. Nevertheless, a miraculous event at the origin of this association does not seem to be a good starting point from a scientific point of view. 5. Some aspects of this novel fusion scenario that are relevant for hypotheses based on the three domains concept This paper can be read as an “exercice de style” to demonstrate the failure of the best possible fusion hypothesis to solve all problems that plague this type of evolutionary scenario. Another objective was to consider some aspects of the origin of Eukarya with new angles, trying to refute as much as possible the usual critiques put forward against fusion hypotheses in general, and, during this process, to get new ideas that could be generally relevant for scenarios describing ancient cellular evolution. I am indeed tempted to conclude from this exercise that, focusing on Thaumarchaea and PVC bacteria on one side, and viruses on the other, bring new ideas for more realistic scenarios for the origin of Eukarya. The discovery of previously ignored “eukaryal-like” features in PVC bacteria and Thaumarchaea suggests that the Last Bacterial Common Ancestor (LBCA) and the Last Archaea Common Ancestor (LACA) were probably both more complex than previously thought. Some of these features might have been even present in LUCA (see below). It is thus now easier than before to derive modern Eukarya from LUCA and/or from the last common ancestor of Archaea and Bacteria. The detection of eukaryal-like MC proteins in PVC bacteria is an especially important milestone. Phylogenetic analysis of PVC MC proteins suggests that the PVC ancestor was probably compartmentalized, harbouring at least four genes encoding MC proteins (Santarella et al., 2010). Also presently restricted to a subgroup of PVC bacteria, the presence of eukaryal-like tubulins in some members of this superphylum might be also significant. If PVC bacteria occupy a basal position in bacterial phylogeny, as suggested by the analysis of slowly evolving positions in rRNA genes (Brochier and Philippe, 2002), one can imagine that the LBCA was itself compartmentalized, already containing ancestors of MC proteins and tubulin. Some observations suggest indeed a tendency to reductive evolution in Bacteria. In particular, phylogenomic analysis suggests that the LBCA had a complete operon for genes encoding the typical bacterial cell division and peptidoglycan synthesis apparatus, whereas many modern bacteria (especially in the PVC superphylum) have only retained a subset of these genes (Pilhofer et al., 2008). A tendency to reductive evolution is also apparent in Archaea. The distribution of actin, FtsZ and ESCRT proteins in this domain suggests that LACA had a machinery for cell division and vesicle production more complex than those of modern archaea (Makarova et al., 2010). Similarly, 87 phylogenomic analysis indicates that the ribosome of LACA contained more ribosomal proteins than the ribosomes of modern archaea (Lecompte et al., 2002). Recently, using a probabilistic model that takes into account various modes of evolution, Csurös and Miklós, (2009) also concluded that archaeal evolution occurred mainly via streamlining of a large ancestral genome. Archaea and Bacteria have thus both probably experienced reductive evolution during the divergence of their various phyla. If this tendency was already present in the lineages leading to the LACA and the LBCA, it might well be that these two ancestors were themselves already less complex than LUCA. In particular, CM proteins from PVC bacteria and Eukarya being probably homologous, they might have been already present in LUCA, and later on lost in Archaea and most Bacteria. The frequent loss of these proteins that occurred during PVC diversification supports such a scenario (Santarella et al., 2010; Forterre and Gribaldo, 2010). The phylogeny of true tubulins, with tubulins of PVC bacteria branching as an outgroup of eukaryal tubulins (Jenkins et al., 2002) can be also interpreted as supporting the presence of tubulin in the common ancestor of Eukarya and Bacteria. LUCA might have thus been a rather complex compartmentalized cell (Forterre and Gribaldo, 2010). There is presently a debate about the nature of the genome of LUCA (either RNA or DNA). At this point, one should realize that a compartmentalized LUCA with an ancestral cytoskeleton does not refute the hypothesis of a LUCA with an RNA genome. Indeed, as previously argued by Poole and Logan (2005) RNA genomes can be replicated and repaired faithfully. Furthermore, modern RNA viruses encode proteins as sophisticated as those encoded by DNA viruses and some of these proteins are very efficent in the manipulation of both the eukaryal cytoskeleton and endomembrane system to produce viral factories (Novoa et al., 2005; Netherton et al., 2007). If LUCA was already a sophisticated cell with some features of Eukarya (and probably “bacterial type” lipids) the path from LUCA to LECA and modern Eukarya becomes much easier to imagine without the need to invoke dubious fusion scenarios. In the framework of the three domains concept, modern Eukarya did not evolved from Archaea (or from Bacteria, as in Cavalier-Smith’s scenarios, see CavalierSmith, 1989, 2010) but from an ancestor that they share with Archaea and/or Bacteria (Fig. 4). This explains why Eukarya exhibit a mixture of archaeal and bacterial features (we don’t need to invoke fusion to explain this observation). Eukarya have inherited these features either from LUCA (they were lost in Archaea or Bacteria) or from the specific common ancestor they share with either Archaea or Bacteria (Fig. 4). The main advantage of the three domains scenarios is to readily explain the existence of eukaryal specific protein families and molecular mechanisms. These eukaryal specific features correspond either to ancient features that have been lost in both Archaea and Bacteria or to features that have appeared in the eukaryal lineage after its divergence from the two others. Unfortunately, it is not easy to decide between these alternatives. For instance, a major question mark that 88 P. Forterre / Research in Microbiology 162 (2011) 77e91 Fig. 4. Update evolutionary scenario based on the three domains concept. Eukarya can exhibit a mixture of features that appear today as bacterial-like (green circles) or archaeal-like (red circles) if these features are ancient traits that were already present in LUCA and were later on selectively lost in either Archaea or Bacteria. In addition, Eukarya can exhibit features in common with their sister group in the universal tree of life (orange circles). In this drawing, the root has been put in the bacterial branch as currently assumed. In the viral hypothesis for the origin of eukaryal complexity, many specific eukaryal features were introduced by viruses in the eukaryal lineage (violet circles). remains open is the existence (or not) of spliceosomes in LUCA. Phylogenomic analyses have shown that LECA already has a full fledge spliceosome and that its genome was probably full of introns (Collins and Penny, 2005; Koonin, 2009). The spliceosome might thus have been either already present in LUCA or emerged between LUCA and LECA. The spliceosome is often view as a recent features that only originated in the eukaryal lineage, however, the recent finding of a nucleomorph (relic nuclei of algal endosymbionts of nonphotosynthetic Eukarya) that have lost all introns and genes encoding spliceosomal components (Lane et al., 2007) testifies that one can easily imagine that the spliceosome was present in LUCA and later on lost in Archaea and Bacteria. At that point, it still remains to understand why Bacteria and Archaea evolved according to a reduction scenario, whereas Eukarya evolved according to a scenario whose hallmark is sophistication (at least from FECA to LUCA, since many eukaryal lineages have experienced reductive evolution after LECA). I suggested 15 years ago that Bacteria and Archaea evolved toward simplification because they experienced passage through a thermophilic period (Forterre, 1995). In that “thermoreduction hypothesis”, their “prokaryotic” characters (small size, high macromolecular turnover, coupling of traduction and translation) were selected in response to adaptation to high temperatures (see also Poole et al., 1999; Glansdorff et al., 2008). A major problem for life at high temperature is indeed the chemical thermodegradation of informational macromolecules, especially mRNA and proteins. Thermoreduction might have been stronger in Archaea, with, in particular, the invention of “archaeal lipids” that, together with those of reverse gyrase (Forterre, 2002), allowed some of them to conquer biotopes above 90 C. The thermoreduction hypothesis has recently received a strong experimental support from the work of Boussau and co-workers who concluded from in silico analyses of rRNA and universal proteins evolution that LUCA was probably a mesophile, but that the LACA and LBCA were both thermophiles (Boussau et al., 2008). In that case, Eukarya might be the descendants of those ancient cells that failed to adapt to high temperature. Most of them were possibly wiped out by the descendants of the LACA and LBCA that adapted later on to low temperature, except those with large size that succeeded to make a living by eating mesophilic archaea and bacteria. Another possibility (non-exclusive) is that viruses play a role in the orientation of each domain toward different evolutionary pathways. In that case, we can reintroduce here our proposals on the role of viruses in the PTV scenario for the origin of Eukarya, but this time in the framework of the three domains concept (Fig. 4). One can still assume that massive invasions of ancient Eukarya by specific viral families bring indeed new proteins in the eukaryal lineage and induced an arms race that favoured their evolution toward increasing complexity. The ideas that many eukaryal specific proteins originated first in NCLDV or other viral lineage unique to Eukarya and that retroviruses played a critical role in shaping the eukaryal chromosomes and/or in the origin of meiotic sex can be easily recycled in scenarios based on the three domains concept. Furthermore, whereas in fusion hypotheses eukaryal viruses are supposed to have evolved from bacterial or archaeal viruses, something difficult to accept, one can suggest now that specific eukaryal viruses, such as retroviruses and NCLDV, originated from ancient viruses that were already thriving at the time of LUCA (Prangishvili et al., 2006a). As previously mentioned, a striking conclusion from studies of viral diversity is that members of different domains are infected by different viral families (for review, see Prangishvili et al., 2006a). In particular, at least in the state of our present knowledge, Eukarya are infected by a much greater diversity of viral families than Bacteria and Archaea. For instance, RNA viruses are very abundant and diverse in Eukarya (singlestranded, double stranded, segmented, non segmented) whereas they are quite rare in Bacteria and presently unknown in Archaea. In the framework of the thermoreduction hypotheses, it is possible that RNA viruses (relics of the RNA-protein world, see Forterre, 2005) were unable, with few exceptions, to follow their hosts in hot environments, considering the high instability of RNA at high temperature (Forterre, 1995). In particular, retroviruses and most retroelements, that are so abundant in eukaryal genomes, might have been eliminated during the period of reductive evolution that led from LUCA to LBCA and LACA. P. Forterre / Research in Microbiology 162 (2011) 77e91 I suggest here that the type of viruses that were “selected” by a given lineage at the onset of the formation of the three domains of life (Prangishvili et al., 2006a) was determinant in the mode of evolution of this domain. Hence, the type of viruses present in ancient Eukarya was possibly a major factor in the evolution of this domain toward more and more sophistication. In particular, the loss of retroviruses in Archaea and Bacteria and their conservation in Eukarya might have been a major factor (possibly in addition to the thermoreduction story) that pushed the evolution of Archaea and Bacteria toward reduction and those of Eukarya toward complexification. The absence of retroviruses in yeast could explain, for instance, why these microbial Eukarya, which have evolved by reduction from multicellular fungi, seem now “blocked” in a kind of “prokaryotic-like situation” (small size, small and compact genomes, few introns, unicellular). All models for the origin of Eukarya have generally ignored the possible role of viruses in that process, except for the origin of the nucleus (Takemura, 2001; Bell, 2001). However, comparative genomics and common sense tell us that, in order to understand the history of life on our planet, one has to study not only the evolutionary relationships between the three cellular domains, but also those between the three viral “domains”, and, above all, the consequences of their co-evolution. For that reason, I am really convinced that the viral hypothesis for the origin of complexity in Eukarya is worth further exploration. 6. Perspective Hypotheses on the origin of Eukarya will be always based on our knowledge of modern biodiversity and molecular mechanisms. The discovery of novel eukaryal features in groups of Archaea and Bacteria only recently recognized and extensively studied should encourage us to continue actively the exploration of the microbial world. The recent discovery of giant multicellular species in Thaumarchaea illustrate well the possibility of further surprising discoveries in these phyla (Muller et al., 2010). The discovery of mimivirus was another breakthrough that helped to remind biologists of the importance to consider viruses in evolutionary scenarios (Raoult et al., 2004; Claverie and Abergel., 2010, Forterre, 2010a). In-depth exploration of the cellular and viral biodiversity, especially in those microbial phyla for which there is presently no cultivable representatives, should be now on the top priority list of biologists looking for exciting scientific adventures in the XXI century (and of course on the top priority list of funding agencies and Heads of Institutes). We don’t know if we will have one day the last word on the origin of Eukarya, but, if coupled to further exploration of terrestrial life, this quest will certainly not be vain. In any case, one can only wonder how many surprises remain to be unveiled in the vast microbial world. Acknowledgments I am grateful to the two reviewers for especially important remarks and suggestions. 89 References Arkhipova, 2006. Distribution and phylogeny of Penelope-like elements in eukaryotes. Syst. Biol. 55, 875e885. Bamford, D.H., 2003. Do viruses form lineages across different domains of life? Res. Microbiol. 154, 231e236. Bamford, D.H., Grimes, J.M., Stuart, D.I., 2005. What does structure tell us about virus evolution? Curr. Opin. Struct. Biol. 15, 655e663. Bandea, C.I., 2009. The origin and evolution of viruses as molecular organisms. http://hdl.handle.net/10101/npre.2009.3886.1 Available from Nature Preceeding. Bell, P.J., 2001. Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus? J. Mol. Evol. 53, 251e256. Boussau, B., Blanquart, S., Necsulea, A., Lartillot, N., Gouy, M., 2008. Parallel adaptations to high temperatures in the Archaean eon. Nature 456, 942e945. Boyer, M., Gimenez, G., Suzan-Monti, M., Raoult, D., 2010. Classification and determination of possible origins of ORFans through analysis of nucleocytoplasmic large DNA viruses. Intervirology 53, 310e320. Brüssow, H., 2009. The not so universal tree of life or the place of viruses in the living world. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 364, 2263e2274. Brochier, C., Philippe, H., 2002. Phylogeny: a non-hyperthermophilic ancestor for bacteria. Nature 417, 244. Brochier-Armanet, C., Boussau, B., Gribaldo, S., Forterre, P., 2008a. Mesophilic Crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 6, 245e252. Brochier-Armanet, C., Gribaldo, S., Forterre, P., 2008b. A DNA topoisomerase IB in Thaumarchaeota testifies for the presence of this enzyme in the last common ancestor of Archaea and Eucarya. Biol. Direct 3, 54. Cavalier-Smith, T., 1989. Molecular phylogeny. Archaebacteria and Archezoa. Nature 339, 100e101. Cavalier-Smith, T., 2010. Origin of the cell nucleus, mitosis and sex: roles of intracellular coevolution. Biol. Direct 5, 7. Chueh, A.C., Northrop, E.L., Brettingham-Moore, K.H., Choo, K.H., Wong, L.H. , 2009. LINE retrotransposon RNA is an essential structural and functional epigenetic component of a core neocentromeric chromatin. PLoS Genet. 5, e1000354. Cock, J.M., Sterck, L., Rouzé, P., Scornet, D., Allen, A.E., Amoutzias, G., Anthouard, V., Artiguenave, F., Aury, J.M., Badger, J.H., Beszteri, B., Billiau, K., Bonnet, E., Bothwell, J.H., Bowler, C., Boyen, C., Brownlee, C., Carrano, C.J., Charrier, B., Cho, G.Y., Coelho, S.M., Collén, J., Corre, E., Da Silva, C., Delage, L., Delaroque, N., Dittami, S.M., Doulbeau, S., Elias, M., Farnham, G., Gachon, C.M., Gschloessl, B., Heesch, S., Jabbari, K., Jubin, C., Kawai, H., Kimura, K., Kloareg, B., Küpper, F.C., Lang, D., Le Bail, A., Leblanc, C., Lerouge, P., Lohr, M., Lopez, P.J., Martens, C., Maumus, F., Michel, G., Miranda-Saavedra, D., Morales, J., Moreau, H., Motomura, T., Nagasato, C., Napoli, C.A., Nelson, D.R., Nyvall-Collén, P., Peters, A.F., Pommier, C., Potin, P., Poulain, J., Quesneville, H., Read, B., Rensing, S.A., Ritter, A., Rousvoal, S., Samanta, M., Samson, G., Schroeder, D.C., Ségurens, B., Strittmatter, M., Tonon, T., Tregear, J.W., Valentin, K., von Dassow, P., Yamagishi, T., Van de Peer, Y., Wincker, P., 2010. The Ectocarpus genome and the independent evolution of multicellularity in brown algae. Nature 465, 617e621. Collins, L., Penny, D., 2005. Complex spliceosomal organization ancestral to extant eukaryotes. Mol. Biol. Evol., 1053e1066. Comeau, A.M., Krisch, H.M., 2005. War is peaceedispatches from the bacterial and phage killing fields. Curr. Opin. Microbiol. 8, 488e494. Cortez, D., Forterre, P., Gribaldo, S., 2009. A hidden reservoir of integrative elements is the major source of recently acquired foreign genes and ORFans in archaeal and bacterial genomes. Genome Biol. 10, R65. Csurös, M., Miklós, I., 2009. Streamlining and large ancestral genomes in Archaea inferred with a phylogenetic birth-and-death model. Mol. Biol. Evol. 26, 2087e2095. Cubonová, L., Sandman, K., Hallam, S.J., Delong, E.F., Reeve, J.N., 2005. Histone in crenarchaea. J. Bacteriol. 187, 5482e5485. 90 P. Forterre / Research in Microbiology 162 (2011) 77e91 DeDuve, C., 2007. The origin of eukaryotes: a reappraisal. Nat. Rev. Genet. 8, 395e403. DeLange, R.J., Williams, L.C., Searcy, D.G., 1981. A histone-like protein (HTa) from Thermoplasma acidophilum. II. Complete amino acid sequence. J. Biol. Chem. 256, 905e911. Desmond, E., Brochier-Armanet, C., Forterre, P., Gribaldo, S., 2010. On the last common ancestor and early evolution of eukaryotes: reconstructing the history of mitochondrial ribosomes. Res. Microbiol. 162, 53e70. Dong, J.H., Wen, J.F., Tian, H.F., 2007. Homologs of eukaryotic Ras superfamily proteins in prokaryotes and their novel phylogenetic correlation with their eukaryotic analogs. Gene 396, 116e124. Doolittle, R.F., 1995. The origins and evolution of eukaryotic proteins. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 349, 235e240. Erickson, H.P., 2007. Evolution of the cytoskeleton. Bioessays 29, 668e677. Ettema, T.J., Bernander, R., 2009. Cell division and the ESCRT complex: a surprise from the archaea. Commun. Integr. Biol. 2, 86e88. Filée, J., Forterre, P., 2005. Viral proteins functioning in organelles: a cryptic origin? Trends Microbiol. 13, 510e513. Filée, J., Forterre, P., Sen-Lin, T., Laurent, J., 2002. Evolution of DNA polymerase families: evidences for multiple gene exchange between cellular and viral proteins. J. Mol. Evol. 54, 763e773. Forterre, P., 1995. Thermoreduction, a hypothesis for the origin of prokaryotes. CR. Acad. Sci. 318, 415e422. Forterre, P., 2002. A hot story from comparative genomics: reverse gyrase is the only hyperthermophile-specific protein. Trends Genet. 18, 236e237. Forterre, P., 2005. The two ages of the RNA world, and the transition to the DNA world, a story of viruses and cells. Biochimie 87, 793e803. Forterre, P., 2006. Three RNA cells for ribosomal lineages and three DNA viruses to replicate their genomes: a hypothesis for the origin of cellular domain. Proc. Natl. Acad. Sci. USA 103, 3669e3674. Forterre, P., 2010a. Giant viruses: conflicts in revisiting the virus concept. Intervirology 53, 362e378. Forterre, P., 2010b. Manipulation of cellular syntheses and the nature of viruses: the virocell concept. C.R. Acad. Sci. Chimie in press, doi:10.1016/j.crci. 2010.06.007. Forterre, P. 2010c The universal tree of life and the Last Universal Cellular Ancestor (LUCA): revolution and counter-revolutions. In: CaetanoAnollés (Ed.), Evolutionary Genomics and Systems Biology. Forterre, P., Gribaldo, S., 2010. Bacteria with a eukaryotic touch: a glimpse on ancient evolution? Proc. Natl. Acad. Sci. USA 107, 12739e12740. Forterre, P., Philippe, H., 1999. Where is the root of the tree of life. Bioessays 21, 871e879. Forterre, P., Prangishvili, D., 2009a. The great billion-year war between ribosome- and capsid-encoding organisms (cells and viruses) as the major source of evolutionary novelties. Ann. N. Y. Acad. Sci. 1178, 65e77. Forterre, P., Prangishvili, D., 2009b. The origin of viruses. Res. Microbiol. 160, 466e472. Forterre, P., Gribaldo, S., Gadelle, D., Serre, M.C., 2007. Origin and evolution of DNA topoisomerases. Biochimie 89, 427e446. Fritz-Laylin, L.K., Prochnik, S.E., Ginger, M.L., Dacks, J.B., Carpenter, M.L., Field, M.C., Kuo, A., Paredez, A., Chapman, J., Pham, J., Shu, S., Neupane, R., Cipriano, M., Mancuso, J., Tu, H., Salamov, A., Lindquist, E., Shapiro, H., Lucas, S., Grigoriev, I.V., Cande, W.Z., Fulton, C., Rokhsar, D.S., Dawson, S. C., 2010. The genome of Naegleria gruberi illuminates early eukaryotic versatility. Cell 140, 631e642. Fuerst, J.A., 2005. Intracellular compartmentation in planctomycetes. Annu. Rev. Microbiol. 59, 299e328. Fuerts, J.A., 1995. The planctomycetes :emerging models for microbial ecology, evolution and cell biology. Microbiology 141, 1493e1506. Gladyshev, E.A., Arkhipova, I.R., 2007. Telomere-associated endonucleasedeficient Penelope-like retroelements in diverse eukaryotes. Proc. Natl. Acad. Sci. USA 104, 9352e9357. Glansdorff, N., Xu, Y., Labedan, B., 2008. The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner. Biol. Direct 9, 29. Gribaldo, S., Daubin, V., Forterre, P. Poole, A., 2010. Brochier-Armanet, C. The origin of eukaryotes and their evolutionary relationship with Archaea: have we reached a phylogenomic impasse? Nat. Rev. Microbiol. 8, 743e752. Jékely, G., 2003. Small GTPases and the evolution of the eukaryotic cell. Bioessays 5, 1129e1138. Jékely, G., 2007. Origin of eukaryotic endomembranes: a critical evaluation of different model scenarios. Adv. Exp. Med. Biol., 38e51. Jenkins, C., Samudrala, R., Anderson, I., Hedlund, B.P., Petroni, G., Michailova, N., Pinel, N., Overbeek, R., Rosati, G., Staley, J.T., 2002. Genes for the cytoskeletal protein tubulin in the bacterial genus Prosthecobacter. Proc. Natl. Acad. Sci. USA 99, 17049e17054. Jetten, M.S., Niftrik, L., Strous, M., Kartal, B., Keltjens, J.T., Op den Camp, H.J., 2009. Biochemistry and molecular biology of anammox bacteria. Crit. Rev. Biochem. Mol. Biol. 44, 65e84. Keller, J., Leulliot, N., Soler, N., Collinet, B., Vincentelli, R., Forterre, P., van Tilbeurgh, H., 2009. A protein encoded by a new family of mobile elements from Euryarchaea exhibits three domains with novel folds. Protein Sci. 18, 850e855. Kidmose, R.T., Vasiliev, N.N., Chetverin, A.B., Andersen, G.R., Knudsen, C.R., 2010. Structure of the Qbeta replicase, an RNA-dependent RNA polymerase consisting of viral and host proteins. Proc. Natl. Acad. Sci. U S A 107, 10884e10889. Koonin, E.V., 2007. The Biological Big Bang model for the major transitions in evolution. Biol. Direct 2, 21. Koonin, E.V., 2009. Intron-dominated genomes of early ancestors of eukaryotes. J. Hered. 100, 618e623. Koonin, E.V., 2010a. The origin and early evolution of eukaryotes in the light of phylogenomics. Genome Biol. 11, 209. Koonin, E.V., 2010b. The incredible expanding ancestor of eukaryotes. Cell 5 (140), 606e608. Koonin, E.V., Moss, B., 2010. Viruses know more than one way to don a cap. Proc. Natl. Acad. Sci. USA 107, 3283e3284. Koonin, E.V., Yutin, N., 2010. Origin and evolution of eukaryotic large nucleocytoplasmic DNA viruses. Intervirology 53, 284e292. Koonin, E.V., Senkevich, T.G., Dolja, V.V., 2006. The ancient Virus World and evolution of cells. Biol. Direct 9, 1e29. Krupovic, M., Gribaldo, S., Bamford, D., Forterre, P., 2010. The evolutionary history of archaeal MCM helicases: a case study of vertical evolution combined with hitch-hiking of mobile genetic elements. Mol. Biol. Evol. doi: 10.1093/molbev/msq161. Kurland, C.G., Canbäck, B., Berg, O.G., 2007. The origins of modern proteomes. Biochimie 89, 1454e1463. Kwapisz, M., Beckouët, F., Thuriaux, P., 2008. Evolution of Eukaryotic RNA dependent RNA polymerases. Trends Genet. 24, 211e215. Lane, C.E., van den Heuvel, K., Kozera, C., Curtis, B.A., Parsons, B.J., Bowman, S., Archibald, J.M., 2007. Nucleomorph genome of Hemiselmis andersenii reveals complete intron loss and compaction as a driver of protein structure and function. Proc. Natl. Acad. Sci. USA 104, 19908e19913. Lecompte, O., Ripp, R., Thierry, J.C., Moras, D., Poch, O., 2002. Comparative analysis of ribosomal proteins in complete genomes: an example of reductive evolution at the domain scale. Nucleic Acids Res. 30, 5382e53890. Lee, K.C., Webb, R.I., Janssen, P.H., Sangwan, P., Romeo, T., Staley, J.T., Fuerst, J.A., 2009. Phylum Verrucomicrobia representatives share a compartmentalized cell plan with members of bacterial phylum Planctomycetes. BMC Microbiol. 9, 5. Lonhienne, T.G., Sagulenko, E., Webb, R.I., Lee, K.C., Franke, J., Devos, D., Nouwens, A., Carroll, B.J., Fuerst, J., 2010. Endocytosis-like protein uptake in the bacterium Gemmata obscuriglobus. Proc. Natl. Acad. Sci. USA 107, 12883e12888. Makarova, K.S., Yutin, N., Bell, S.D., Koonin, E.V., 2010. Evolution of diverse cell division and vesicle formation systems in Archaea. Nat. Rev. Microbio, 1e11. Margulis, L., Bermudes, D., 1985. Symbiosis as a mechanism of evolution: status of cell symbiosis theory. Symbiosis 1, 101e124. Margulis, L., Chase, D., To, L.P., 1979. Possible evolutionary significance of spirochaetes. Proc. R. Soc. Lond. B. Biol. Sci. 204, 189e198. Martin, W., Müller, M., 1998. The hydrogen hypothesis for the first eukaryote. Nature 392, 37e41. P. Forterre / Research in Microbiology 162 (2011) 77e91 Moreira, D., López-Garcı́a, P., 2009. Ten reasons to exclude viruses from the tree of life. Nat. Rev. Microbiol. 7, 306e311. Moreira, D., Lopez-Garcia, P., 1998. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. J. Mol. Evol. 47, 517e530. Muller, F., Brissac, T., Le Bris, N., Felbeck, H., Gros, O., 2010. First description of giant Archaea (Thaumarchaeota) associated with putative bacterial ectosymbionts in a sulfidic marine habitat. Environ. Microbiol. 12, 2371e2383. Netherton, C., Moffat, K., Brooks, E., Wileman, T., 2007. A guide to viral inclusions, membrane rearrangements, factories, and viroplasm produced during virus replication. Adv. Virus Res. 70, 101e182. Novoa, R.R., Calderita, G., Arranz, R., Fontana, J., Granzow, H., Risco, C., 2005. Virus factories: associations of cell organelles for viral replication and morphogenesis. Biol. Cell. 97, 147e172. Oberto, J., Breuil, N., Hecker, A., Farina, F., Brochier-Armanet, C., Culetto, E., Forterre, P., 2009. Qri7/OSGEPL, the mitochondrial version of the universal Kae1/YgjD protein, is essential for mitochondrial genome maintenance. Nucleic Acids Res. 37, 5343e5352. Ogata, H., Claverie, J.M., 2007. Unique genes in giant viruses: regular substitution pattern and anomalous short size. Genome Res. 17, 1353e1361. Ogino, T., Yadav, S.P., Banerjee, A.K., 2010. Histidine-mediated RNA transfer to GDP for unique mRNA capping vesicular stomatitis virus RNA polymerase. Proc. Natl. Acad. Sci. USA 107, 3463e3468. Pace, N.R., 2009. Problems with « procaryote ». J. Bacteriol. 191, 2008e2010. Pearson, A., Budin, M., Brocks, J.J., 2003. Phylogenetic and biochemical evidence for sterol synthesis in the bacterium Gemmata obscuriglobus. Proc. Natl. Acad. Sci. U S A 100, 15352e15357. Pilhofer, M., Rappl, K., Eckl, C., Bauer, A.P., Ludwig, W., Schleifer, K.H., Petroni, G., 2008. Characterization and evolution of cell division and cell wall synthesis genes in the bacterial phyla Verrucomicrobia, Lentisphaerae, Chlamydiae, and Planctomycetes and phylogenetic comparison with rRNA genes. J. Bacteriol. 190, 3192e3202. Poole, A.M., Logan, D.T., 2005. Modern mRNA proofreading and repair: clues that the last universal common ancestor possessed an RNA genome? Mol. Biol. Evol. 22, 1444e1455. Poole, A.M., Penny, D., 2007. Evaluating hypotheses for the origin of eukaryotes. Bioessays 29, 74e84. Poole, A., Jeffares, D., Penny, D., 1999. Early evolution: prokaryotes, the new kids on the block. Bioessays 21, 880e889. Prangishvili, D., Forterre, P., Garrett, R.A., 2006a. Viruses of the Archaea: a unifying view. Nat. Rev. Microbiol. 4, 837e848. Prangishvili, D., Garrett, R.A., Koonin, E.V., 2006b. Evolutionary genomics of archaeal viruses: unique viral genomes in the third domain of life. Virus Res. 117, 52e67. Rancurel, C., Khosravi, M., Dunker, A.K., Romero, P.R., Karlin, D., 2009. Overlapping genes produce proteins with unusual sequence properties and offer insight into de novo protein creation. J. Virol. 83, 10719e10736. Raoult, D., Forterre, P., 2008. Redefining viruses: lessons from Mimivirus. Nat. Rev. Microbiol. 6, 315e319. Raoult, D., Audic, S., Robert, C., Abergel, C., Renesto, P., Ogata, H., La Scola, B., Suzan, M., Claverie, J.M., 2004. The 1.2-megabase genome sequence of Mimivirus. Science 306, 1344e1350. Rivera, M.C., Lake, J.A., 1992. Evidence that eukaryotes and eocyte prokaryotes are immediate relatives. Science 257, 74e76. Rivera, M.C., Lake, J.A., 2004. The ring of life provides evidence for a genome fusion origin of eukaryotes. Natur 431, 152e155. Rote, N.S., Chakrabarti, S., Stetzer, B.P., 2004. The role of human endogenous retroviruses in trophoblast differentiation and placental development. Placenta 25, 673e683. Saini, H.K., Fischer, D., 2007. Structural and functional insights into Mimivirus ORFans. BMC Genomics. 8, 115. 91 Samson, R.Y., Bell, S.D., 2009. Ancient ESCRTs and the evolution of binary fission. Trends Microbiol. 17, 507e513. Sandman, K., Krzycki, J.A., Dobrinski, B., Lurz, R., Reeve, J.N., 1990. HMf, a DNA-binding protein isolated from the hyperthermophilic archaeon Methanothermus fervidus, is most closely related to histones. Proc. Natl. Acad. Sci. U S A 87, 5788e5791. Santarella, R., Franke, J., Jaedicke, A., Gorjanacz, M., Bauer, U., Budd, A., Mattaj, I.W., Devos, D.P., 2010. The compartimentalized bacteria of the Planctomycetes-Verrucomicrobia-Chlamydiae superphylum have membrane coat-like proteins. Plos Biol. 8, e1000028. Shuman, 1995. Capping enzyme in eukaryotic mRNA synthesis. Prog. Nucl. Acid Res. Mol. Biol. 50, 101e129. Siegl, A., Kamke, J., Hochmuth, T., Piel, J., Richter, M., Liang, C., Dandekar, T., Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J in press. Spang, A., Hatzenpichler, R., Brochier-Armanet, C., Rattei, T., Tischler, P., Spieck, E., Streit, W., Stahl, D.A., Wagner, M., Schleper, C., 2010. Distinct gene set in two different lineages of ammonia-oxidizing archaea supports the phylum Thaumarchaeota. Trends Microbiol. 18, 331e340. Stanier, R.Y., Van Niel, C.B., 1962. The concept of a bacterium. Arch. Mikrobiol. 42, 17e35. Stein, D.B., Searcy, D.G., 1978. Physiologically important stabilization of DNA by a prokaryotic histone-like protein. Science 202, 219e221. Suzan-Monti, M., La Scola, B., Barrassi, L., Espinosa, L., SuzanMonti, Raoult D., 2007. Ultrastructural characterization of the giant volcano-like virus factory of Acanthamoeba polyphaga Mimivirus. PLoS One. 2, e328. Takemura, M., 2001. Poxviruses and the origin of the eukaryotic nucleus. J. Mol. Evol. 52, 419e425. Treusch, A.H., Leininger, S., Kletzin, A., Schuster, S.C., Klenk, H.P., Schleper, C., 2005. Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling. Environ. Microbiol. 7, 1985e1995. Tyagi, N., Anamika, K., Srinivasan, N., 2010. A framework for classification of prokaryotic protein kinases. PLoS One 5, e10608. Van Niftrik, L., van Helden, M., Kirchen, S., van Donselaar, E.G., Harhangi, H.R., Webb, R.I., Fuerst, J.A., Op den Camp, H.J., Jetten, M.S., Strous, M., 2010. Intracellular localization of membrane-bound ATPases in the compartmentalized anammox bacterium ’Candidatus Kuenenia stuttgartiensis’. Mol. Microbiol. 77, 701e715. Villarreal, L.P., DeFilippis, V.R., 2000. A hypothesis for DNA viruses as the origin of eukaryotic replication proteins. J. Virol. 74, 7079e7084. Villarreal, L.P., Witzany, G., 2010. Viruses are essential agents within the roots and stem of the tree of life. J. Theor. Biol. 262, 698e710. Wagner, M., Horn, M., 2006. The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol. 17, 241e249. Wilkins, A.S., Holliday, R., 2009. evolution meiosis Mitosis.Genetics 181, 3e12. Woese, C.R., Fox, G.E., 1977. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. USA 74, 5088e5090. Woese, C.R., Kandler, O., Wheelis, M.L., 1990. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. U S A 87, 4576e4579. Yin, Y., Fischer, D., 2008. Identification and investigation of ORFans in the viral world. BMC Genomics 9, 24. Yutin, N., Makarova, K.S., Mekhedov, S.L., Wolf, Y.I., Koonin, E.V., 2008. The deep archaeal roots of eukaryotes. Mol. Biol. Evol. 25, 1619e1630. Yutin, N., Wolf, M.Y., Wolf, Y.I., Koonin, E.V., 2009. The origins of phagocytosis and eukaryogenesis. Biol. Direct 4, 9.