Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Nicotinamide adenine dinucleotide wikipedia , lookup
Mitochondrial replacement therapy wikipedia , lookup
Photosynthesis wikipedia , lookup
Metalloprotein wikipedia , lookup
Biochemistry wikipedia , lookup
Mitochondrion wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Microbial metabolism wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Citric acid cycle wikipedia , lookup
Photosynthetic reaction centre wikipedia , lookup
Light-dependent reactions wikipedia , lookup
NADH:ubiquinone oxidoreductase (H+-translocating) wikipedia , lookup
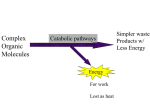
Mitochondrial Electron Transport • The cheetah, whose capacity for aerobic metabolism makes it one of the fastest animals Fatty Acids Acetyl Co A Pyruvate Glucose Citric acid cycle supplies NADH and FADH2 to the electron transport chain Amino Acids Reduced coenzymes NADH and FADH2 are formed in matrix from: (1) Oxidative decarboxilation of pyruvate to acetyl CoA (2) Aerobic oxidation of acetyl CoA by the citric acid cycle (3) Oxidation of fatty acids and amino acids The NADH and FADH2 are energy-rich molecules because each contains a pair of electrons having a high transfer potential. The reduced and oxidized forms of NAD The reduced and oxidized forms of FAD Electrons of NADH or FADH2 are used to reduce molecular oxygen to water. A large amount of free energy is liberated. The electrons from NADH and FADH2 are not transported directly to O2 but are transferred through series of electron carriers that undergo reversible reduction and oxidation. The flow of electrons through carriers leads to the pumping of protons out of the mitochondrial matrix. The resulting distribution of protons generates a pH gradient and a transmembrane electrical potential that creates a protonmotive force. ATP is synthesized when protons flow back to the mitochondrial matrix through an enzyme complex ATP synthase. The oxidation of fuels and the phosphorylation of ADP are coupled by a proton gradient across the inner mitochondrial membrane. Oxidative phosphorylation is the process in which ATP is formed as a result of the transfer of electrons from NADH or FADH2 to O2 by a series of electron carriers. OXIDATIVE PHOSPHORYLATION IN EUKARYOTES TAKES PLACE IN MITOCHONDRIA Two membranes: outer membrane inner membrane (folded into cristae) Two compartments: (1) the intermembrane space (2) the matrix The outer membrane is permeable to small molecules and ions because it contains pore-forming protein (porin). The inner membrane is impermeable to ions and polar molecules. Contains transporters (translocases). Location of mitochondrial complexes • Inner mitochondrial membrane: Electron transport chain ATP synthase • Mitochondrial matrix: Pyruvate dehydrogenase complex Citric acid cycle Fatty acid oxidation THE ELECTRON TRANSPORT CHAIN Series of enzyme complexes (electron carriers) embedded in the inner mitochondrial membrane, which oxidize NADH2 and FADH2 and transport electrons to oxygen is called respiratory electron-transport chain (ETC). The sequence of electron carriers in ETC NADH FMN Fe-S succinate FAD Fe-S Co-Q Fe-S cyt b cyt c1 cyt c cyt a cyt a3 O2 High-Energy Electrons: Redox Potentials and Free-Energy Changes In oxidative phosphorylation, the electron transfer potential of NADH or FADH2 is converted into the phosphoryl transfer potential of ATP. Phosphoryl transfer potential is G°' (energy released during the hydrolysis of activated phosphate compound). G°' for ATP = -7.3 kcal mol-1 Electron transfer potential is expressed as E'o, the (also called redox potential, reduction potential, or oxidation-reduction potential). E'o (reduction potential) is a measure of how easily a compound can be reduced (how easily it can accept electron). All compounds are compared to reduction potential of hydrogen wich is 0.0 V. The larger the value of E'o of a carrier in ETC the better it functions as an electron acceptor (oxidizing factor). Electrons flow through the ETC components spontaneously in the direction of increasing reduction potentials. E'o of NADH = -0.32 volts (strong reducing agent) E'o of O2 = +0.82 volts (strong oxidizing agent) NADH FMN Fe-S succinate FAD Fe-S Co-Q Fe-S cyt b cyt c1 cyt c cyt a cyt a3 O2 Important characteristic of ETC is the amount of energy released upon electron transfer from one carrier to another. This energy can be calculated using the formula: Go’=-nFE’o n – number of electrons transferred from one carrier to another; F – the Faraday constant (23.06 kcal/volt mol); E’o – the difference in reduction potential between two carriers. When two electrons pass from NADH to O2 : Go’=-2*96,5*(+0,82-(-0,32)) = -52.6 kcal/mol THE RESPIRATORY CHAIN CONSISTS OF FOUR COMPLEXES Components of electrontransport chain are arranged in the inner membrane of mitochondria in packages called respiratory assemblies (complexes). FMN II III IV III I NADH I Fe-S II succinate FAD Fe-S Co-Q Fe-S cyt b IV cyt c1 cyt c cyt a cyt a3 O2 The energy is released not in a single step of electron transfer but in incremental amount at each complex. 26.8 Energy released at three specific steps in the chain is collected in form of transmembrane proton gradient and used to drive the synthesis of ATP. Complexes I-IV • Mobile coenzymes: ubiquinone (Q) and cytochrome c serve as links between ETC complexes • Complex IV reduces O2 to water Complex I (NADH-ubiquinone oxidoreductase) Transfers electrons from NADH to Co Q (ubiquinone) Consist of: - enzyme NADH dehydrogenase (FMN - prosthetic group) - iron-sulfur clusters. NADH reduces FMN to FMNH2. Electrons from FMNH2 pass to a Fe-S clusters. Fe-S proteins convey electrons to ubiquinone. QH2 is formed. The flow of two electrons from NADH to coenzym Q leads to the pumping of four hydrogen ions out of the matrix. NADH FMN matrix Fe-S Iron-sulfur clusters contains two or four iron ions and two or four inorganic sulfides. Clusters are coordinated by four cysteine residues. Iron ions in Fe-S complexes cycle between Fe2+ or Fe3+ states. NADH-Q oxidoreductase - an enormous enzyme consisting of 34 polypeptide chains. L-shaped (horizontal arm lying in the membrane and a vertical arm that projects into the matrix). Complex II (succinate-ubiquinon oxidoreductase) Transfers electrons from succinate to Co Q. Form 1 consist of: - enzyme succinate dehydrogenase (FAD – prosthetic group) - iron-sulfur clusters. Succinate reduces FAD to FADH2. Then electrons pass to Fe-S proteins which reduce Q to QH2 Form 2 and 3 contains enzymes acyl-CoA dehydrogenase (oxidation of fatty acids) and glycerol phosphate dehydrogenase (oxidation of glycerol) which direct the transfer of electrons from acyl CoA to Fe-S proteins. Complex II does not contribute to proton gradient. All electrons must pass through the ubiquinone (Q)ubiquinole (QH2) pair. Ubiquinone Q: - lipid soluble molecule, - smallest and most hydrophobic of all the carriers - diffuses within the lipid bilayer - accepts electrons from I and II complexes and passes them to complex III. Complex III (ubiquinol-cytochrome c oxidoreductase) Transfers electrons from ubiquinol to cytochrome c. Consist of: cytochrome b, Fe-S clusters and cytochrome c1. Cytochromes – electron transferring proteins containing a heme prosthetic group (Fe2+ Fe3+). Oxidation of one QH2 is accompanied by the translocation of 4 H+ across the inner mitochondrial membrane. Two H+ are from the matrix, two from QH2 Q-cytochrome c oxidoreductase is a dimer. Each monomer contains 11 subunits. Q-cytochrome c oxidoreductase contains three hemes: two b-type hemes within cytochrome b, and one c-type heme within cytochrome c1. Enzyme also contains an iron-sulfur protein with an 2Fe-2S center. Q cycle two molecules of QH2 are oxidized to form two molecules of Q, one molecule of Q is reduced to QH2, two molecules of cytochrome c are reduced, four protons are released on the cytoplasmic side, two protons are removed from the mitochondrial matrix Complex IV (cytochrome c oxidase) Transfers electrons from cytochrome c to O2. Composed of: cytochromes a and a3. Catalyzes a four-electron reduction of molecular oxygen (O2) to water (H2O): O2 + 4e- + 4H+ 2H2O Translocates 2H+ into the intermembrane space Cytochrome c oxidase consists of 13 subunits and contains two hemes (two iron atom) and three copper ions, arranged as two copper centers. The Catalytic Cycle of Cytochrome c Oxidise The four protons used for the production of two molecules of water come from the matrix. The consumption of these four protons contributes to the proton gradient. Cytochrome c oxidase pumps four additional protons from the matrix to the cytoplasmic side of the membrane in the course of each reaction cycle (mechanism under study). Totally eight protons are removed from the matrix in one reaction cycle (4 electrons) Cellular Defense Against Reactive Oxygen Species If oxygen accepts four electrons - two molecules of H2O are produced single electron - superoxide anion (O2.-) two electrons – peroxide (O22-). O2.-, O22- and, particularly, their reaction products are harmful to cell components - reactive oxygen species or ROS. DEFENSE superoxide dismutase (manganese-containing version in mitochondria and a copper-zinc-dependent in cytosol) O2.- + O2.- + 2H+ = H2O2 + O2 catalase H2O2 + H2O2 = O2 + 2 H2O antioxidant vitamins: vitamins E and C reduced glutathione This complex mechanical art fo can be viewed as a metaphor for t molecular apparatus underly electron transport and ATP synthe by oxidative phosphorylatio OXIDATIVE PHOSPHORYLATION A PROTON GRADIENT POWERS THE SYNTHESIS OF ATP The transport of electrons from NADH or FADH2 to O2 via the electron-transport chain is exergonic process: NADH + ½O2 + H+ H2O + NAD+ FADH2 + ½O2 H2O + FAD+ Go’ = -52.6 kcal/mol for NADH -36.3 kcal/mol for FADH2 How this process is coupled to the synthesis of ATP (endergonic process)? ADP + Pi ATP + H2O Go’=+7.3 kcal/mol The Chemiosmotic Theory • Proposed by Peter Mitchell in the 1960’s (Nobel Prize, 1978) • Chemiosmotic theory: electron transport and ATP synthesis are coupled by a proton gradient across the inner mitochondrial membrane Mitchell’s postulates for chemiosmotic theory 1. Intact inner mitochondrial membrane is required 2. Electron transport through the ETC generates a proton gradient 3. ATP synthase catalyzes the phosphorylation of ADP in a reaction driven by movement of H+ across the inner membrane into the matrix Overview of oxidative phosphorylation + + + - - - + + + - As electrons flow through complexes of ETC, protons are translocated from matrix into the intermembrane space. The free energy stored in the proton concentration gradient is tapped as protons reenter the matrix via ATP synthase. As result ATP is formed from ADP and Pi. An artificial system demonstrating the basic principle of the chemiosmotic hypothesis Synthetic vesicles contains bacteriorhodopsin and mitochondrial ATP synthase. Bacteriorhodopsin protein that pumps protons when illuminated. When the vesicle is exposed to light, ATP is formed. ATP Synthase Two units, Fo and F1 (“knob-andstalk”; “ball on a stick”) F1 contains the catalytic subunits where ADP and Pi are brought together for combination. F0 spans the membrane and serves as a proton channel. Energy released by collapse of proton gradient is transmitted to the ATP synthesis. • F1 contains 5 types of polypeptide chains a3b3gde • Fo - a1b2c10-14 (c subunits form cylindrical, membranebound base) • Fo and F1 are connected by a ge stalk and by exterior column (a1b2 and d) • The proton channel – between c ring and a subunit. • there are 3 active sites, one in each b subunit • c-e-g unit forms a “rotor” • a-b-d-a3b3 unit is the “stator” • passage of protons through the Fo channel causes the rotor to spin • rotation of the g subunit inside the a3b3 hexamer causes domain movements in the bsubunits, opening and closing the active sites Each b subunit contains the catalytic site. At any given time, each site is in different conformation: open (O), loose (L) or tight (T). O conformation binds ADP and Pi The affinity for ATP of T conformation is so high that it converts ADP and Pi into ATP. Binding-Change Mechanism of ATP Synthase 1. ADP and Pi bind to an open site 2. Passage of protons causes each of three sites to change conformation. 3. The open conformation (containing the newly bound ADP and Pi) becomes a loose site. The loose site filled with ADP and Pi becomes a tight site. The ATP containing tight site becomes an open site. 4. ATP released from open site, ADP and Pi form ATP in the tight site Experimental observation of ATP synthase rotation • Fluorescent protein arm (actin) attached to g subunits • a3b3 subunits bound to a glass plate • Arm seen rotating when ATP added (observed by microscopy) OXIDATIVE PHOSPHORYLATION Oxidative phosphorylation creates ATP which is the universal currency of free energy in biological systems MOVEMENT ACROSS THE MITOCHONDRIAL MEMBRANES Electrons from Cytosolic NADH Enter Mitochondria by Shuttles NADH is generated in the cytosol in glycolysis. The inner mitochondrial membrane is impermeable to NADH and NAD+. Electrons from NADH, but not NADH itself, are carried across the mitochondrial membrane. Two shuttles move electrons: glycerol 3-phosphate shuttle and malate-aspartate shuttle Glycerol 3-phosphate shuttle Active in skeletal muscles and brain. Electrons enter the electrontransport chain via complex II. Therefore only 1.5 molecules of ATP are produced. Malate-aspartate shuttle Active in heart and liver. 2.5 molecules of ATP are produced. Active Transport of ATP, ADP and Pi Across the Inner Mitochondrial Membrane • ATP must be transported to the cytosol, and ADP and Pi must enter the matrix • ADP/ATP carrier, adenine nucleotide translocase, exchanges mitochondrial ATP4- for cytosolic ADP3• The exchange causes a net loss of -1 in the matrix (draws some energy from the H+ gradient) • Phosphate (H2PO4-) is transported into matrix in symport with H+. Phosphate carrier draws on pH. • Both transporters consume proton-motive force Mechanism of ATP and ADP Transport ATP-ADP translocase is abundant in the inner mitochondrial membrane (about 14% of the protein) The entry of ADP into the matrix is coupled to the exit of ATP. Mitochondrial Transporters ATP-ADP translocase – antiport of ATP and ADP Phosphate carrier – antiport of H2PO4- and OH- (symport of H2PO4- and H+) Dicarboxylate carrier – antiport of malate, succinate, or fumarate and H2PO4Tricarboxylate carrier – antiport of citrate and H+ and malate Pyruvate carrier – antiport of pyruvate and OH- (symport of pyruvate and H+) REGULATION OF OXIDATIVE PHOSPHORYLATION Coupling of Electron Transport with ATP Synthesis Electron transport is tightly coupled to phosphorylation. ATP can not be synthesized by oxidative phosphorylation unless there is energy from electron transport. Electrons do not flow through the electron-transport chain to O2 unless ADP is phosphorylated to ATP. Important substrates: NADH, O2, ADP Intramitochondrial ratio ATP/ADP is a control mechanism High ratio inhibits oxidative phosphorylation as ATP allosterically binds to a subunit of Complex IV Respiratory control The most important factor in determining the rate of oxidative phosphorylation is the level of ADP. The regulation of the rate of oxidative phosphorylation by the ADP level is called respiratory control Uncoupling of Electron Transport with ATP Synthesis Uncoupling of oxidative phosphorylation generates heat to maintain body temperature in hibernating animals, in newborns, and in mammals adapted to cold. Brown adipose tissues is specialized for thermogenesis. Inner mitochondrial membrane contains uncoupling protein (UCP), or thermogenin. UCP forms a pathway for the flow of protons from the cytosol to the matrix. Uncouplers • Uncouplers are lipid-soluble aromatic weak acids • Uncouplers deplete proton gradient by transporting protons across the membrane 2,4-Dinitrophenol: an uncoupler • Because the negative charge is delocalized over the ring, both the acid and base forms of DNP are hydrophobic enough to dissolve in the membrane. Specific inhibitors of electron transport chain and ATP-synthase Specific inhibitors of electron transport are invaluable in revealing the sequence of electron carriers. Rotenone and amytal block electron transfer in Complex I. Antimycin A interferes with electron flow thhrough Complex III. Cyanide, azide, and carbon monoxide block electron flow in Complex IV. ATP synthase is inhibited by oligomycin which prevent the influx of protons through ATP synthase. ATP Yield Ten protons are pumped out of the matrix during the two electrons flowing from NADH to O2 (Complex I, III and IV). 3 Six protons are pumped out of the matrix during the two electrons flowing from FADH2 to O2 4 (Complex 4III and IV). 2 Translocation of 3H+ required by ATP synthase for each ATP produced 1 H+ needed for transport of Pi. Net: 4 H+ transported for each ATP synthesized For NADH: 10 H+/ 4H+) = 2.5 ATP For FADH2: 6 H+/ 4 H+ = 1.5 ATP