Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Deoxyribozyme wikipedia , lookup
Gene desert wikipedia , lookup
Gene expression profiling wikipedia , lookup
Gene regulatory network wikipedia , lookup
Non-coding DNA wikipedia , lookup
Transcriptional regulation wikipedia , lookup
Gene expression wikipedia , lookup
Plant breeding wikipedia , lookup
Molecular cloning wikipedia , lookup
Genome evolution wikipedia , lookup
Molecular evolution wikipedia , lookup
Genetic engineering wikipedia , lookup
Real-time polymerase chain reaction wikipedia , lookup
Promoter (genetics) wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Community fingerprinting wikipedia , lookup
Silencer (genetics) wikipedia , lookup
Genomic library wikipedia , lookup
Expression vector wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
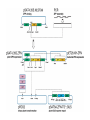
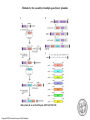
Gene Targeting ● ● GT是指在基因组的原位实现对基因的定点突变、定 点整合、基因置换及基因修复等。对基因功能的研 究提供了极大的方便,比基因的反义抑制和RNAi更 直观可靠,尤其在鉴定基因家族成员的功能时更是 如此。 Gene targeting (GT) refers to the alteration of a specific DNA sequence in an endogenous gene at its original locus in the genome by homologous recombination (HR) and, often, to the conversion of the endogenous gene into a designed sequence, i.e. base changes or gene disruptions through gene replacements. Two approaches for GT: 1. Homologous recombination-dependent gene targeting, e.g. tobacco(ALS乙酰乳酸合成酶 ) , Arbidopsis(PPO, protoporphyrinogen oxidase原卟啉原氧化酶 ) and rice(Waxy); 2. Chimeric RNA/DNA oligonucleotide-directed targeted point mutations, e.g. AHAS(乙酰羟酸合成酶) in maize and ALS in tobacco and rice. ● 同源重组介导的GT在细菌、真菌、低等藻类和 动物中已成功广泛的应用,而在高等植物的应 用研究进展缓慢,重组率很低(<1%),仅为 非同源重组的万分之一。 GT在植物中的进展 ● ● ● ● Paszkowski等1988年首次报道在烟草中作的基因打 靶的尝试。 Kammerer和Cove(1996)首次在小立碗藓中实现了基 因的同源重组,打靶效率达到90%,近似于酵母的95 % 。 Terada 等(2002;2007)利用高效正负向筛选标记 筛选同源重组子的策略,白喉毒素基因DT-A作负向筛 选标记,成功的在水稻Waxy或Adh2中整合进一个潮霉 素抗性基因htp,实现对靶基因的敲除。打靶效率达 到1%。 打靶的效率与DNA断裂双链的修复过程中倾向于同源 重组还是非同源重组有很大的关系。 Integration events of a transgene associated with homology-dependent GT with positive-negative selection Terada, R., et al. Plant Physiol. 2007;144:846-856 Copyright ©2007 American Society of Plant Biologists Strategy for the modification of the Adh locus Terada, R., et al. Plant Physiol. 2007;144:846-856 Copyright ©2007 American Society of Plant Biologists 提高打靶效率的方法 ● ● ● 通过表达内源或外源同源重组相关的酶提高同 源重组率,如RAD、RecQ和RecA等。 引入重组酶体系Cre/lox。 锌指核酶ZFNs(zinc finger nucleases)引 入定点双链断裂。ZFN由一个DNA识别域和一个 非特异性核酸内切酶构成,具有很强的特异性 和可塑性。 Zinc-finger nucleases. (a) Zinc fingers are depicted that recognize nucleotide triplets of a target sequence. Multiple fingers can be joined together to create zinc-finger proteins that recognize extended sequence patterns. (b) Zinc-finger domains are fused to a type II restriction endonuclease such as FokI. (c) When the FokI monomers are brought into proximity by DNA binding, a functional nuclease is created that cleaves the target sequence. ZFN homodimer binding to DNA. Shown is a three-finger zinc finger linked to the Fn domain through a flexible peptide linker. At the N-terminus of each ZFN resides a nuclear localization signal (NLS). The Fn domain is linked to the C-terminal finger (in this case finger 3) of the zinc finger domain. For most efficient cleavage there is no amino acid linker between the zinc finger domain and the Fn domain. The binding sites are arranged in an inverted orientation so that one ZFN is making most of its major contacts with one strand of DNA, whereas the other ZFN is making most of its major contacts with the other strand of DNA. Between the two binding sites is a nucleotide spacer (NNN...), the sequence of which does not seem to be important. This figure is a modification of Figure 2c from Jantz et al.47. Nature Biotechnology 23, 967 - 973 (2005) Molecular reagents for measuring homologous recombination. (a) pDW1273 encodes a functional GUS:NPTII reporter gene. AI denotes the artificial intron within the GUS coding sequence. The ColE1 replicon and Ampr gene are for recovery of the integrated construct by plasmid rescue. The GUS:NPTII coding sequence in pDW1363 has a 600 bp deletion that includes GUS and NPTII coding sequences critical for function (mutant forms of GUS and/or NPTII are indicated by lower-case letters). A Zif268 recognition site (depicted in Figure 1) is inserted at the site of the deletion. The Hygr marker functions in plants and can be used to select cells carrying the reporter construct. Filled triangles depict the left and right borders of the T-DNA. Open arrows indicate the primers used for the PCR reactions in Figure 4. The donor DNA, pDW1269, lacks sequences 5' of the artificial intron and is used to repair the GUS:NPTII reporter in pDW1363 by homologous recombination. The donor DNA has a diagnostic XhoI restriction site. (b) The target gene and donor DNA are shown undergoing recombination. Numbers adjacent to the open arrows indicate the size of expected PCR products. The length of homology between donor and target is given below the donor DNA. Schematic of the target reporter and zinc finger recognition sequences. a Target reporter construct with tandem, overlapping, partial gfp gene fragments, a 3′ pat selectable marker gene fragment, left and right homologous sequences and zinc finger binding sites. b Recognition sequences for ZFN-1 and ZFN-2. Outline of zinc finger nuclease (ZFN) functional assays and their vector systems. The step-by-step comprehensive analysis of novel ZFNs is composed of four distinct assays (left panel), each based on monitoring ZFN activity using a defined set of vectors (right panel). The in vitro digestion assay tests the digestion activity of an Escherichia coli-expressed ZFN on its recognition site cloned on a target vector. The TDNA repair assay requires the assembly of a dual-expression cassette on an Agrobacterium tumefaciens binary vector and tests the ability of a constitutively expressed ZFN to digest and repair a mutated GUS reporter gene cloned on the T-DNA region of that vector. The transgene repair assay calls for separating the ZFN expression cassette from its target site based on the activation of a mutated GUS reporter gene in transgenic calli, while the whole-plant repair assay activates the mutated GUS reporter gene in seedling or mature plant tissues upon specific activation of the ZFN. TS, ZFN target site; KAN, kanamycin; hsp, heat shock promoter. ● Structural features of the zinc finger nuclease (ZFN) assembly and expression vector systems. A zinc finger protein (ZFP) coding sequence can be assembled by Klenow/PCR using a combination of overlapping backbone and sequence-dependent oligonucleotides fused to the FokI endonuclease domain in pSAT4.35SP.NLSFokI, producing the plant expression vector pSAT4.35SP.ZFN. The entire ZFN coding sequence can be transferred onto a pET28.XH-based vector producing a pET28.XH-ZFN vector, suitable for ZFN expression in bacterial cells and for in vivo digestion assays. The pSAT4.35SP.ZFN can be modified by replacing the 35S constitutive promoter with a heat-shock-inducible promoter, producing a plasmid that is useful for the whole-plant DNA repair assay. A plant-selectable marker, a ZFN and a mutated GUS reporter expression cassette can be mounted onto a pRCS2-based binary plant transformation vector using a combination of rare-cutting restriction enzymes and can then be used for various in planta assays. 35SP, 35S promoter; 35ST, 35S terminator; hsp, heat shock promoter. Delivery of Multiple Transgenes to Plant Cells ● The growing interest in dissecting and analyzing complex metabolic pathways and the need to exploit the full potential of multigene traits for plant biotechnology mandate the development of new methods and tools for the delivery and stacking of multiple genes in plant cells. Three approaches for the production of transgenic plants carrying multiple new traits ● ● ● retransformation the stacking of several transgenes by successive delivery of single genes into transgenic plants; Cotransformation the combined delivery of several transgenes in a single transformation experiment; sexual crosses between transgenic plants carrying different transgenes. ● For all the benefits and simplicity of combining cotransformation, retransformation, and crosses while using single-gene vectors for the delivery of multiple genes into plant species, these methods suffer from several drawbacks. These include the undesirable incorporation of a complex T-DNA integration pattern, often observed during integration of T-DNA molecules from multiple sources (De Neve et al., 1997 ; De Buck et al., 1999 ), and the time needed for retransformation or crosses between transgenic plants. More importantly, transgenes derived from different sources typically integrate at different locations in the plant genome, which may lead to various expression patterns and possible segregation of the transgenes in the offspring. ● Methods for the assembly of multiple-gene binary plasmids. A, The Cre/loxPmediated multigene assembly process. i, Cre/loxP recombination of the pYLVSA plasmid into the pYLTAC747 acceptor binary plasmid. ii, Release of the pYLVS backbone by I-SceI digestion and ligation with a LS linker. This ligation abolishes the I-SceI site from pYLTAC747-A. iii, Cre/loxP recombination of the pYLSV-B plasmid into the pYLTAC747-A and release of the pYLSV backbone by PI-SceI digestion and ligation with a LV linker (iv). B, The MultiRound Gateway assembly process. i, Gateway recombination between attL1 and attR1 and between attL1 and attR1 sites by LR clonase and conversion of the ccdBbased binary pDEST vector into a sacB-based Destination vector. ii, Gateway recombination between attL3 and attR3 and between attL4 and attR4 sites by LR clonase and reconversion of the destination binary vector into a ccdB-based plasmid. C, The homing endonucleases pRCS/pAUX vector system. Assembly of a multigene binary plasmid is achieved by successive cloning of various gene expression cassettes using rare-cutting homing endonucleases. Methods for the assembly of multiple-gene binary plasmids Dafny-Yelin, M., et al. Plant Physiol. 2007;145:1118-1128 Copyright ©2007 American Society of Plant Biologists The general structure of a pSAT-based plant expression vector Dafny-Yelin, M., et al. Plant Physiol. 2007;145:1118-1128 Copyright ©2007 American Society of Plant Biologists