Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Transformation (genetics) wikipedia , lookup
Magnesium transporter wikipedia , lookup
Gene regulatory network wikipedia , lookup
Light-dependent reactions wikipedia , lookup
Biochemistry wikipedia , lookup
Paracrine signalling wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Biochemical cascade wikipedia , lookup
Community fingerprinting wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Signal transduction wikipedia , lookup
Microbial metabolism wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
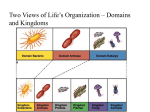
Chapter 17 Archaea I. Phylogeny and General Metabolism 17.1 Phylogenetic Overview of Archaea 17.2 Energy Conservation and Autotrophy in Archaea 17.1 Phylogenetic Overview of Archaea Archaea share many characteristics with both Bacteria and Eukarya Archaea are split into two major groups Crenarchaeota Euryarchaeota Detailed Phylogenetic Tree of the Archaea Figure 17.1 17.2 Energy Conservation and Autotrophy in Archaea Chemoorganotrophy and chemolithotrophy ▪ With the exception of methanogenesis, bioenergetics and intermediary metabolism of Archaea are similar to those found in Bacteria - Glucose metabolism : EMP or slightly modified Entner-Doudoroff pathway - Oxidation of acetate to CO2 : TCA cycle or some slight variations of TCA cycle : Reverse route of acetyl-CoA pathway - Electron transport chains with a, b, and c-type cytochromes present in some Archaea : use O2, S0, or some other electron acceptor (nitrate or fumarate) : establish proton motive force : ATP synthesis through membrane-bound ATPase - Chemolithotrophy : H2 as a common electron donor and energy source is well established ▪ Autotrophy via several different pathways is widespread in Archaea ▪ acetyl-CoA pathway in methanogens and most hyperthermophiles ▪ Reverse TCA cycle in some hyperthermophiles ▪ Calvin cycle in Methanococcus jannaschii and a Pyrococcus species (both are hyperthermophiles) II. Euryarchaeota 17.3 Extremely Halophilic Archaea 17.4 Methane-Producing Archaea: Methanogens 17.5 Thermoplasmatales 17.6 Thermococcales and Methanopyrus 17.7 Archaeoglobales 17.8 Nanoarchaeum and Aciduliprofundum II. Euryarchaeota Euryarchaeota Physiologically diverse group of Archaea Many inhabit extreme environments E.g., high temperature, high salt, high acid 17.3 Extremely Halophilic Archaea Haloarchaea Extremely halophilic Archaea Have a requirement for high salt concentrations Typically require at least 1.5 M (~9%) NaCl for growth Found in solar salt evaporation ponds, salt lakes, and artificial saline habitats (i.e., salted foods) Hypersaline Habitats for Halophilic Archaea Great Salt Lake, Utah Figure 17.2a Seawater Evaporating Ponds Near San Francisco Bay, California Figure 17.2b Pigmented Haloalkaliphiles Growing in pH 10 Soda Lake in Egypt Figure 17.2c SEM of Halophilic Bacteria Figure 17.2d 17.3 Extremely Halophilic Archaea Extremely hypersaline environments are rare Most found in hot, dry areas of world Salt lakes can vary in ionic composition, selecting for different microbes Great Salt Lake similar to concentrated seawater Soda lakes are highly alkaline hypersaline environments Ionic Composition of Some Highly Saline Environments Haloarchaea Reproduce by binary fission Do not form resting stages or spores Most are nonmotile Most are obligate aerobes Possess adaptations to life in highly ionic environments Some Genera of Extremely Halophilic Archaea Some Genera of Extremely Halophilic Archaea Micrographs of the Halophile Halobacterium salinarum Dividing cell Glycoprotein subunit structure on the cell wall Figure 17.3 Water balance in extreme halophiles Halophiles need to maintain osmotic balance This is usually achieved by accumulation or synthesis of organic compatible solutes Halobacterium species instead pump large amounts of K+ into the cell from the environment Intracellular K+ concentration exceeds extracellular Na+ concentration and positive water balance is maintained Concentration of Ions in Cells of Halobacterium salinarum Proteins of halophiles Highly acidic Contain fewer hydrophobic amino acids and lysine residues Some haloarchaea are capable of light-driven synthesis of ATP Bacteriorhodopsin Cytoplasmic membrane proteins that can absorb light energy and pump protons across the membrane Model for the Mechanism of Bacteriorhodopsin Figure 17.4 Other rhodopsins can be present in Archaea Halorhodpsin Light-driven pump that pumps Cl- into cell as an anion for K+ Sensory rhodopsins Control phototaxis 17.4 Methane-Producing Archaea: Methanogens Methanogens Microbes that produce CH4 Found in many diverse environments Taxonomy based on phenotypic and phylogenetic features Process of methanogensis first demonstrated over 200 years ago by Alessandro Volta The Volta Experiment Figure 17.5 Habitats of Methanogens Micrographs of Cells of Methanogenic Archaea Methanobrevibacter ruminantium Figure 17.6a Micrographs of Cells of Methanogenic Archaea Methanobrevibacter arboriphilus Figure 17.6b Micrographs of Cells of Methanogenic Archaea Methanospirillum hungatei Figure 17.6c Micrographs of Cells of Methanogenic Archaea Methanosarcina barkeri Figure 17.6d Characteristics of Some Methanogenic Archaea Characteristics of Some Methanogenic Archaea Diversity of Methanogens Demonstrate diversity of cell wall chemistries Pseudomurein (e.g., Methanobacterium, Methanobrevibacter) Methanochondroitin (e.g., Methanosarcina) - (N-acetylgalactosamine + glucuronic acid)n Protein or glycoprotein (e.g., Methanocaldococcus) S-layers (e.g., Methanospirillium) Micrographs of Thin Sections of Methanogenic Archaea Methanobrevibacter ruminantium Methanosarcina barkeri Figure 17.7a Hyperthermophilic and Thermophilic Methanogens Methanocaldococcus jannaschii Figure 17.8a Substrates for Methanogens Obligate anaerobes 11 substrates, divided into 3 classes, can be converted to CH4 by pure cultures of methanogens Other compounds (e.g., glucose) can be converted to methane, but only in cooperative reactions between methanogens and other anaerobic bacteria (syntrophic metabolism) Substrates Converted to Methane by Methanogens 17.5 Thermoplasmatales Methanocaldococcus jannaschii as a model methanogen Contains 1.66 mB circular genome with about 1,700 genes Genes for central metabolic pathways and cell division - Similar to those in Bacteria Genes encoding transcription and translation - More closely resemble those of Eukarya Over 50% of genes - Have no counter parts in known genes from Bacteria and Eukarya Thermoplasmatales Taxonomic order within the Euryarchaeota Contains 3 genera Thermoplasma Ferroplasma Picrophilus Thermophilic and/or extremely acidophilic Thermoplasma and Ferroplasma lack cell walls Thermoplasma Chemoorganotrophs Facultative aerobes via sulfur respiration Thermophilic Acidophilic Thermoplasma Species Thermoplasma acidophilum Figure 17.9a Thermoplasma Species Thermoplasma volcanium Isolated from Hot Springs Figure 17.9b A Self-Heating Coal Refuse Pile, Habitat of Thermoplasma Self-heats from microbial metabolism. Figure 17.10 Thermoplasma (cont’d) Evolved unique cytoplasmic membrane structure to maintain positive osmotic pressure and tolerate high temperatures and low pHs Membrane contains lipopolysaccharide-like material (lipoglycan) consisting of tetraether lipid monolayer membrane with mannose and glucose Membrane also contains glycoproteins but not sterols Structure of the Tetraether Lipoglycan of T. acidophilum Figure 17.11 Ferroplasma Chemolithotrophic Acidophilic Oxidizes Fe2+ to Fe3+, uses CO2 as carbon source Grows in mine tailings containing pyrite (FeS) - Generates acid (acid mine drainage) Picrophilus Extreme acidophiles Grow optimally at pH 0.7 Have an S-layer Model microbe for extreme acid tolerance 17.6 Thermococcales and Methanopyrus Three phylogenetically related genera of hyperthermophilic Euryarchaeota Thermococcus Pyrococcus Methanopyrus Comprise a branch near root of archaeal tree Detailed Phylogenetic Tree of the Archaea Figure 17.1 Thermococcales Distinct order that contains Thermococcus and Pyrococcus Thermococcus: organics + So, 55-95oC Pyrococcus: organics + So, opt. 100oC, 70-106oC ** In the absence of So, form H2 Indigenous to anoxic thermal waters Highly motile Spherical Hyperthermophilic Archaea Shadowed cells of Thermococcus celer Figure 17.12a Dividing cell of Pyrococcus furiosus Figure 17.12b Methanopyrus Methanogenic (CO2 + H2) Isolated from hot sediments near submarine hydrothermal vents and from walls of “black smoker” Opt. temp. 100oC, max. temp. 110oC (the most thermophilic of all known methangens) Contains unique membrane lipids: unsaturated form Contains 2,3-diphosphoglycerate in the cytoplasm (> 1 M) Functions as thermostabilizing agent Methanopyrus Electron Micrograph of a cell of Methanopyrus Kandleri Figure 17.13a Methanopyrus Structure of novel lipid of M. kandleri Figure 17.13b 17.7 Archaeoglobales Archaeoglobales Hyperthermophilic Couple oxidation of H2, lactate, pyruvate, glucose, or complex organic compounds to the reduction of SO42- to H2S Archaeoglobus Opt. temp. 83oC Produce methane, but lacks genes for methyl-CoM reductase Ferroglobus Opt. temp. 85oC Fe2+ + NO3- → Fe3+ + NO2- + NO Archaeoglobales TEM of Archaeoglobus fulgidus Figure 17.14a Freeze-etched Electron Micrograph of Ferroglobus placidus Figure 17.14b 17.8 Nanoarchaeum and Aciduliprofundum Nanorchaeum equitans One of the smallest cellular organisms (~0.4 µm) Obligate symbiont of the crenarchaeote Ignicoccus Contains one of the smallest genomes known (0.49 mbp) Lacks genes for all but core molecular processes Depends upon host for most of its cellular needs Nanoarchaeum Ignicoccus Nanoarchaeum Fluorescence micrograph of cells of Nanoarchaeum Figure 17.15a TEM of a thin section of a cell of Nanoarchaeum Figure 17.15b Aciduliprofundum Thermophilic: 55-75oC Acidophile: pH 3.3-5.8, lives in sulfide deposits in hydrothermal vents Oragnics + So or Fe3+ III. Crenarchaeota 17.9 Habitats and Energy Metabolism of Crenarchaeota 17.10 Hyperthermophiles from Terrestrial Volcanic Habitats 17.11 Hyperthermophiles from Submarine Volcanic Habitats 17.12 Nonthermophilic Crenarchaeota 17.9 Habitats and Energy Metabolism of Crenarchaeota Crenarchaeota Inhabit temperature extremes Most cultured representatives are hyperthermophiles Other representatives found in extreme cold environments Habitats of Crenarchaeota Habitats of Hyperthermophilic Archaea A typical Solfatara in Yellowstone National Park Figure 17.16a Sulfur-rich hot spring Figure 17.16b A typical boiling spring of neutral pH in Yellowstone Park; Imperial Geyser Figure 17.16c An acidic iron-rich geothermal spring Figure 17.16d Hyperthermophilic Crenarchaeota Most are obligate anaerobes Chemoorganotrophs or chemolithotrophs with diverse electron donors and acceptors Energy-Yielding Reactions of Hyperthermophilic Archaea Properties of Some Hyperthermophilic Crenarchaeota 17.10 Hyperthermophiles from Terrestrial Volcanos Sulfolobales An order containing the genera Sulfolobus and Acidianus Sulfolobus Grows in sulfur-rich acidic hot springs Aerobic chemolithotrophs that oxidize reduced sulfur or iron Acidianus Also lives in acidic sulfur hot springs Able to grow using elemental sulfur both aerobically and anaerobically (as an electron donor and electron acceptor, respectively) Acidophilic Hyperthermophilic Archaea, the Sulfolobales Sulfolobus acidocaldarius Figure 17.17a Acidianus infernus Figure 17.17b Thermoproteales An order containing the key genera Thermoproteus, Thermofilum, and Pyrobaculum Inhabit neutral or slightly acidic hot springs or hydrothermal vents Rod-Shaped Hyperthermophiles, the Thermoproteales Thermoproteus neutrophilus Figure 17.18a Thermofilum librum Figure 17.18b Pyrobaculum aerophilum Figure 17.18c 17.11 Hyperthermophiles from Submarine Volcanos Shallow-water thermal springs and deep-sea hydrothermal vents harbor the most thermophilic of all known Archaea Pyrodictium and Pyrolobus Desulfurococcus and Ignicoccus Staphylothermus Desulfurococcales with Temperature Optima > 100°C Pyrodictium occultum Thin-section electron micrograph of P. occultum Figure 17.19a Thin section of a cell of Pyrolobus fumarii Figure 17.19c Negative stain of a cell of strain 121, the most heat-loving of all known Figure 17.19d Desulfurococcales with Temperature Optima Below Boiling Thin section of a cell of Desulfurococcus saccharovorans Figure 17.20a Extremely large periplasm Thin section of a cell of Ignicoccus islandicus Figure 17.20b The Hyperthermophile Staphylothermus marinus Figure 17.21 17.12 Nonthermophilic Crenarchaeota Nonthermophilic Crenarchaeota have been identified in cool or cold marine waters and terrestrial environments by culture-independent studies Abundant in deep ocean waters Appear to be capable of nitrification Cold-Dwelling Crenarchaeota DAPI (diamidino-2-phenylindole) stained Photo of the Antarctic peninsula Flouresence photomicrograph of seawater treated with FISH probe Figure 17.22 IV. Evolution and Life at High Temperatures 17.13 An Upper Temperature Limit for Microbial Life 17.14 Adaptations to Life at High Temperature 17.15 Hyperthermophilic Archaea, H2, and Microbial Evolution 17.13 An Upper Temperature Limit for Microbial Life What are the upper temperature limits for life? Laboratory experiments with biomolecules suggest 140–150°C Thermophilic and Hyperthermophilic Prokaryotes Figure 17.23 17.14 Adaptations to Life at High Temperature Stability of Monomers Protective effect of high concentration of cytoplasmic solutes Use of more heat-stable molecules e.g., use of nonheme iron proteins instead of proteins that use NAD and NADH Protein Folding and Thermostability Amino acid composition similar to that of nonthermostable proteins Structural features improve thermostability Highly hydrophobic cores Increased ionic interactions on protein surfaces Chaperonins Class of proteins that refold partially denatured proteins Thermosome A major chaperonin protein complex in Pyrodictium DNA Stability High intracellular solute levels stabilize DNA Reverse DNA gyrase Introduces positive supercoils into DNA, which stabilizes DNA Found only in hyperthermophiles High intracellular levels of polyamines (e.g., putrescine, spermidine) stabilize DNA and RNA DNA-binding proteins (archaeal histones) compact DNA into nucleosome-like structures Archael Histones and Nucleosomes Figure 17.25 SSU rRNA Stability Higher GC content Lipid Stability Possess dibiphytanyl tetraether type lipids; form a lipid monolayer membrane structure 17.15 Hyperthermophilic Archaea, H2, and Evolution Hyperthermophiles may be the closest descendants of ancient microbes Hyperthermophilic Archaea and Bacteria are found on the deepest, shortest branches of the phylogenetic tree The oxidation of H2 is common to many hyperthermophiles and may have been the first energyyielding metabolism Upper Temperature Limits for Energy Metabolism Figure 17.26