Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Gene regulatory network wikipedia , lookup
Photosynthesis wikipedia , lookup
NADH:ubiquinone oxidoreductase (H+-translocating) wikipedia , lookup
Basal metabolic rate wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Adenosine triphosphate wikipedia , lookup
Microbial metabolism wikipedia , lookup
Oxidative phosphorylation wikipedia , lookup
Multi-state modeling of biomolecules wikipedia , lookup
Biochemistry wikipedia , lookup
Citric acid cycle wikipedia , lookup
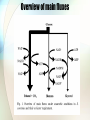
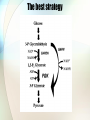
In silico aided metaoblic engineering of Saccharomyces cerevisiae for improved bioethanol production Christoffer Bro et al. 2005 The problem • Under anaerobic conditions, S. cerevisiae produces only four major products from glucose: • CO2, ethanol, biomass and glycerol • To increase the ethanol yield, the flow of carbon going to biomass or glycerol should be redirected towards ethanol. Overview of main fluxes Previous work • Some of the carbon flowing to biomass can be redirected towards ethanol by increasing the consumption of ATP for biomass production or reducing the amount of ATP formed in association with ethanol production. (Nissen et al. 2000) • Deletion of the structural genes in glycerol biosynthesis is not a successful strategy. • The maximum specific growth rate is severely lowered in such strains • Formation of glycerol is necessary for maintaining the redox balance by oxidizing NADH Strategy #1 • Substitution of NADPH-oxidizing reactions in biomass formation with NADH-oxidizing reactions Strategy #2 • Substitution of NAD+-reducing reactions in biomass formation by NADP+-reducing reactions. Strategy #3 • Introduction of a reaction which either directly or via a cycle converts NADH into NADPH. Strategy #4 • Substitution of the glycerol production with production of ethanol, which has a net oxidation of NADH. In silico model • iFF708 (Forster et al., 2003) • 708 genes • 584 metabolites • 1175 reactions Method • A database of 3800 biochemical reactions is derived from the LIGAND database of KEGG. • Each gene (corresponding to a specific biochemical reaction) was inserted one at a time into the genome-scale metabolic model, and the performance of the engineered strain was evaluated. • Two other engineered strains: • Heterologous expression of a non-phosphorylating, NADP+-dependent D-GAPN • Deletion of GDH1 combined with simultaneous overexpression of GDH2 or GLN1 and GLT1. • • • • GDH1: AKG + NH3 + NADPH -> GLU + NADP GDH2: GLU + NAD -> AKG + NH3 + NADH GLN1: GLU + NH3 + ATP -> GLN + ADP + PI GLT1: AKG + GLN + NADH -> NAD + 2 GLU Eight best strains predicted The best strategy In vivo testing of the best strategy • Ethanol production increased by 3% • Reasons for disagreement between experiment and model: • Limited GAPN activity in vivo • Low intracellular NADP+ concentrations compared with NADPH Discussion • “The success of the strategies is due to the tight linking of the different parts of the metabolic network through the common usage of cofactors like NADH, NADPH and ATP, and the genome-scale metabolic model represents a valuable tool for studying how these co-factors link the different parts of the metabolism in a quantitative fashion.” Efficiency of amino acid production in Escherichia coli Anthony Burgard & Costas Maranas, 2001 iJR904 20 8.2609 15.68 18.627 9.4501 11.322 11.81 26.122 7.8728 7.4219 7.5 7.8512 5.7951 5.4913 10.074 20 12.601 4.4907 5.6886 10 Universal model • The universal model is constructed by incorporating 3400 cellular reactions from the KEGG into the modified Keasling stoichiometric model. Arginine production