Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Quantitative trait locus wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Essential gene wikipedia , lookup
X-inactivation wikipedia , lookup
Long non-coding RNA wikipedia , lookup
Microevolution wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Genome evolution wikipedia , lookup
Designer baby wikipedia , lookup
Genome (book) wikipedia , lookup
Minimal genome wikipedia , lookup
Ridge (biology) wikipedia , lookup
Nutriepigenomics wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Genomic imprinting wikipedia , lookup
Gene expression programming wikipedia , lookup
Biology and consumer behaviour wikipedia , lookup
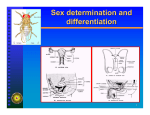
Developmental homology and dissociation Homologous genes need not function in the development of homologous structures (HOX genes, Notch signaling) Expression of a homologous gene does not imply that developmental pathways are also homologous (engrailed and metamerism) Homologous developmental pathways may control the development of non-homologous structures (Dll in appendages, Pax6 in the eyes) Homologous structures need not be specified by homologous genes (insect segmentation) Segmentation of the Drosophila embryo Genetic control of segmentation in Drosophila Segment polarity genes make up the bottom level of the regulatory hierarchy Segment polarity genes establish boundaries between segments and control patterning within each segment Expression of segment polarity genes is conserved in beetles… en wg Tribolium Grasshoppers en Schistocerca Myriapods en/wg Lithobius Crustaceans Artemia wg Spiders en Cupiennius salei wg The functions of en and wg in subdividing the embryo into segments appear to be conserved krusty, a gap mutants in Tribolium en Pair-rule mutants in Tribolium eve pair-rule function is conserved in beetles Chromophore-assisted laser inactivation ftz deletion does not affect segmentation in Tribolium Antennae Pair-rule gene expression in Schistocerca Drosophila Tribolium eve ftz pby even-skipped expression in Lithobius even-skipped expression in Lithobius eve/ en eve expression does not show two-segment periodicity Expression of pair-rule genes in Chelicerates primary pair-rule genes in Cupiennius paired - secondary pair-rule in Tetranychus Evolution of Arthropod segmentation - Some parts of the segmentation pathway are conserved. - There is some turnover of genes within the overall pathway - Pair-rule patterning may be a higher insect innovation Segmentation in long germ band insects Q ui ck Ti m e ™ an d a P l an ar RG B de co m p re ss or ar e n ee de d t o s ee t h is pi ct u re . - Simultaneous generation of segments - Segmentation independent of growth - Occurs in syncytial environment Cellularization before blastoderm formation in grasshoppers Rhodamine dextran injection Segmentation in short germ band insects (Tribolium) -Sequential generation of segments -Segmentation coupled to growth - Occurs in a cellular environment The global patterning mechanisms cannot operate in the same way as in Drosophila Maternal gradient of bicoid establishes AnteriorPosterior axis in the Drosophila embryo The roles of maternal gradients in Drosophila Gradients form from maternally deposited transcripts by diffusion or transport in a cell-free environment bicoid function is conserved in Cyclorhapha Maternal gradients in Drosophila and Megaselia RNA interference Inhibition of Bcd protein synthesis in Megaselia results in posterior duplication (an embryo with two butts…) bicoid does not exist outside higher Dipterans hunchback is a conserved component of the Anterior determination system Megaselia RNA interference Schistocerca Tribolium hunchback is correctly regulated in Drosophila Tribolium hb Tribolium hb transgene in Drosophila Some maternal system must therefore exist in beetles But how does it work without bicoid? hunchback can substitute for bicoid Making anterior hunchback stripe in the absence of bicoid In Tribolium, anterior patterning is controlled by orthodenticle and hunchback otd hb otd; hb otd is deposited maternally Removal of otd and hb eliminates anterior structures A new mechanism for a new mode of development Drosophila Maternal Zygotic bcd otd hb hb Anterior structures Tribolium otd hb Anterior structures A maternal protein gradient can only work in a syncytium bcd has taken over the ancestral functions of otd and hb ? A parasitic wasp, Copidosoma floridanum Polyembryonic development Polyembryonic development Primary morula Polymorula Secondary morulae Polyembryonic development in Copidosoma floridanum Polyembryonic development evolved independently as an adaptation to parasitism Segmentation without maternal gradients engrailed expression in Aphidius ervi Segmentation without pair-rule genes? eve eve Bracon en Aphidius How do developmental pathways diverge from a common ancestral state? - By recruitment and loss of component genes - By re-deployment of old genes in new patterns - By changing regulatory interactions between genes Somatic sex determination pathway in Drosophila Sex determination is cell-autonomous (X:A ratio or dsx expression) Somatic sex in Drosophila is controlled by a splicing cascade Establishment (X:A ratio) Maintenance (autoregulation) Regulation of downstream target genes by doublesex Genotype/sex Yolk protein expression Sex determination mechanisms in insects Y-chromosomal genes (Tipulidae, Tephritidae) Autosomal genes (Culex, Anopheles) Mobile genes (Megaselia, Musca) X:Autosome ratio (Drosophila) Genotype of the mother (Chrysomia, Sciara) Haploid/ diploid (Hymenopterans) Environmental factors (Pseudacteon) Sex determination in the medfly Ceratitis capitata (Tephritidae) Sxl Sex is controlled by a maledetermining factor on the Y tra Differential splicing of transformer in Ceratitis transformer controls sexual differentiation in Ceratitis Female Male Intersexes produced by tra RNAi Sxl and dsx in Megaselia scalaris (Phoridae) Sxl dsx Sex determination in Megaselia Megaselia lacks differentiated sex chromosomes The Maleness factor is mobile and can be located on different chromosomes This can create new Y chromosomes from former autosomes Sex determination systems in Musca domestica In male-heterogametic strains, sex is determined by a single masculinizing factor (M), which can be located either on a Y chromosome, or on 4 different autosomes Some female-heterogametic strains are homozygous for M, and sex is determined by a dominant feminizing factor F Other female-heterogametic strains lack M, and sex is determined by a recessive masculinizing factor Fman In arrhenogenic strains, sex of the offspring depends on the genotype of the mother Sexually dimorphic splicing of dsx is conserved in Musca Musca doublesex expression is sexually dimorphic Somatic sex correlates with sexually dimorphic doublesex splicing, irrespective of the upstream sex determination mechanism Musca dsxF induces vitellogenin synthesis in males Musca dsxM promotes male-specific morphology in Drosophila Musca dsxM has the same phenotypic effect as Drosophila dsxM The mechanism of sex-specific differentiation appears to be conserved, even if the upstream sex determination signal is not dsx is required for the development of reproductive organs in Musca A model for Musca sex determination? M Suppose that: F Then: dsx dsxM dsx produces a male-specific product by default F is required for female-specific splicing of dsx M represses F dsxF F-D could be an M-insensitive allele of F F-man could be a non-functional allele of F Ag and tra could be leaky alleles of F or M expressed in the germiline?? Could F be a homologue of the Drosophila transformer gene? doublesex controls sexual differentiation in Lepidoptera Bombyx mori Sex determination in Lepidoptera In Lepidopterans, females are the heterogametic sex (males ZZ, females ZW) W? or Z:A? Splicing regulator (tra?) Differential splicing of dsx dsxM Male development dsxF Female development Evolution of the sex determination pathway The function of doublesex in sexual differentiation and the sexually dimorphic splicing of dsx are highly conserved The immediate upstream regulator of dsx splicing (tra) may also be conserved? The primary sex determination signals evolve rapidly and vary among closely related groups Presumably, the downstream targets of dsx are also different in different species dsx How does the fly count to 2? Sxl “measures” the X:A ratio reflected in the balance between “numerator” and “denominator” gene products Numerator genes are located on the X chromosome (sisA, sisB, sisC, and runt); a single denominator element (deadpan) is located on an autosome Sxl was recently recruited in the sex determination cascade Sxl protein is highly conserved in all Dipterans: Musca (83%), Chrysomia, Ceratitis, Megaselia, etc. Sxl is spliced in a sexually dimorphic fashion in other Drosophila species (D. virilis, D. subobscura), but not in other Dipterans (Musca, Megaselia, Ceratitis). Dipterans outside Drosophila also lack the male-specific, translation-terminating exon Although the RNA binding domain of Sxl is highly conserved, Sxl proteins from Musca or Ceratitis cannot regulate the splicing of tra in transgenic Drosophila Sex determination mechanism based on Sxl is a Drosophila innovation So where did the numerator elements come from? The origin of numerator genes The role of Sxl in sex determination is very recent, so the numerator genes must also have acquired their functions in sex determination recently All numerator genes have other functions in development that clearly predate their roles in sex determination: segmentation (runt), neurogenesis (sc, da, dpn), signaling (upd) What does it take to be a numerator gene? Genes must be located on the X chromosome Must be able to regulate gene expression Must be expressed very early in development One of the numerator genes is scute bHLH transcription factor with a very ancient function as a proneural gene Part of a gene cluster evolved by tandem duplications (scute, achaete, l(sc)) Shares regulatory elements and overlapping expression with achaete Changes in scute regulation were responsible for its recruitment in the X:A sex determination signal Recruitment of scute for the numerator function The duplication of achaete and scute predates the recruitment of scute as a numerator element In contrast to neurogenesis, achaete cannot substitute for scute in its sex determination role Changes in scute coding sequence were not required for the acquisition of its new function The numerator function of scute depends on specific cis-regulatory elements Models of pathway evolution Retrograde growth from a simple ancestral state Component replacement Emancipation and regrowth