Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
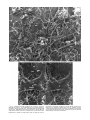
MICROSCOPY RESEARCH AND TECHNIQUE 44:415–429 (1999) Organisation of Xenopus Oocyte and Egg Cortices PATRICK CHANG, DANIEL PÉREZ-MONGIOVI, AND EVELYN HOULISTON Unité de Biologie Cellulaire Marine (UMR. 643 CNRS -Université Paris VI), Station Zoologique 06230 Villefranche-sur-mer, France KEY WORDS cytoskeleton; deep etch; microtubule; actin; intermediate filament; endoplasmic reticulum; determinant; germ plasm; mRNA localisation; cortical rotation; Surface Contraction Waves ABSTRACT The division of the Xenopus oocyte cortex into structurally and functionally distinct ‘‘animal’’ and ‘‘vegetal’’ regions during oogenesis provides the basis of the organisation of the early embryo. The vegetal region of the cortex accummulates specific maternal mRNAs that specify the development of the endoderm and mesoderm, as well as functionally-defined ‘‘determinants’’ of dorso-anterior development, and recognisable ‘‘germ plasm’’ determinants that segregate into primary germ cells. These localised elements on the vegetal cortex underlie both the primary animal-vegetal polarity of the egg and the organisation of the developing embryo. The animal cortex meanwhile becomes specialised for the events associated with fertilisation: sperm entry, calcium release into the cytoplasm, cortical granule exocytosis, and polarised cortical contraction. Cortical and subcortical reorganisations associated with meiotic maturation, fertilisation, cortical rotation, and the first mitotic cleavage divisions redistribute the vegetal cortical determinants, contributing to the specification of dorso-anterior axis and segregation of the germ line. In this article we consider what is known about the changing organisation of the oocyte and egg cortex in relation to the mechanisms of determinant localisation, anchorage, and redistribution, and show novel ultrastructural views of cortices isolated at different stages and processed by the rapid-freeze deep-etch method. Cortical organisation involves interactions between the different cytoskeletal filament systems and internal membranes. Associated proteins and cytoplasmic signals probably modulate these interactions in stage-specific ways, leaving much to be understood. Microsc. Res. Tech. 44:415–429, 1999. r 1999 Wiley-Liss, Inc. INTRODUCTION Developmental and cell biologists interested in the relationship between egg organisation and the establishment of the embryonic body plan have long appreciated the special role of the egg cortex (for instance see Elinson, 1980; Sardet and Chang, 1987; StewartSavage et al., 1991). In many types of embryo, developmental ‘‘determinants’’ identified by functional assays, and specific localised maternal mRNAs with known or suspected developmental functions tend to become associated with specific cortical regions during oogenesis (see Gavis, 1997; Micklem, 1995). Subsequent cortical and subcortical reorganisations associated with oocyte maturation, fertilisation, and/or early mitotic cleavage divisions result in the relocalisation of these localised elements and are responsible for establishment of the embryonic axes (Sardet et al., 1994). The nature of the cortical elements that participate in the localisation anchoring, release, and translocation of maternal determinants and mRNAs are not yet understood. Indeed the composition and organisation of the cortex are still poorly defined in most of the organisms. Xenopus offers a good opportunity to address this problem, for practical reasons (late stage oocytes and eggs are abundant, large, and exhibit clear external polarity) and because of the rapidly accumulating knowledge concerning early developmental events. In this article, we will summarise what is known about cortical organisation in Xenopus oocytes and eggs in relation to the mechanisms of determinant localisation and relocalisation, r 1999 WILEY-LISS, INC. and present some ultrastructural views of replicas prepared from isolated cortices of oocytes and eggs, which give a novel perspective of cortical organisation. A large number of ultrastructural and immunofluorescence studies using a variety of microscopy methods have been published concerning the amphibian oocyte and egg cortex. Filaments of all three cytoskeletal systems have been identified in the cortex at various times in various forms, essentially by immunofluorescence, while electron microscopy has revealed further details of the organisation of these and other components, notably cortical and subcortical ER (StewartSavage et al., 1991). Actin is clearly the predominant cortical protein, with cortical actin conferring contractility, in particular to the thickened animal cortex (Merriam and Sauterer, 1983; Merriam et al., 1983). However, despite the abundance of actin, definitive microfilaments have proven hard to detect in the cortex, except in the cleavage furrow (Franke et al., 1976; Larabell, 1995; Roeder and Gard, 1994). Microtubules come and go in the subcortical region according to spatial and temporal modulation of microtubule dynam- Contract grant sponsor: ARC; Contract grant numbers: 6985, 9144; Contract grant sponsor: CNRS; Contract grant sponsor: Ministère d’Education; Contract grant number: ACC4/Al. *Correspondence to: Evelyn Houliston, Unitée de Biologie Celulaire Marine (ERS.643 CNRS-Université Paris VI), Station Zoologique 06230 Villefrance-surmer, France. E-mail: [email protected] Received 3 September 1998; accepted in revised form 25 November 1998 416 P. CHANG ET AL. ics in the cell (see Houliston and Elinson, 1992) and intermediate filaments (IFs), essentially cytokeratincontaining, form a polarised cortical network that disassembles during meiosis (Klymkowsky, 1995). Disruption studies indicate that the actin cortex of the oocyte provides a polarised framework with respect to which microtubules and IFs are organised (Gard et al., 1997). The participation of microfilaments, intermediate filaments, and microtubules in specific cortical processes has also been investigated, and the accumulated data have been reviewed several times in recent years (Dent and Klymkowsky, 1989; Elinson and Houliston, 1990; Elinson, 1990; Gard et al., 1995, this issue; Klymkowsky and Karnovsky, 1994; Klymkowsky, 1995). For this reason we have chosen not to give extensive details of the literature again here, but rather to outline the possible participation of the cortex in the localisation and redistribution of developmental determinants. STEPS IN CORTICAL LOCALISATION The sequence of localisation events in Xenopus oocytes and eggs involving the cortex is shown in Figure 1. Specialisation of the cortex begins early during oogenesis when the oocyte is small and transparent (Stage I–II according to Dumont, 1972), with the arrival at the cell periphery of a curious dense structure known as the ‘‘Balbiani body’’ or ‘‘mitochondrial cloud’’ (Heasman et al., 1984). A number of specific mRNAs and proteins, as well as structurally defined fibrilo-granular material that gives rise to germinal granules and eventually to germ plasm, are associated together in the mitochondrial cloud (Al-Mukhtar and Webb, 1971; Heasman et al., 1984). The cortical site of mitochondrial cloud attachment, which marks the future vegetal pole of the oocyte, does not appear to be predetermined. The germ plasm and associated mRNAs spread out in distinctive islands across a patch of cortex in the stage II oocyte, marking the future vegetal pole. During mid-oogenesis (Stage III–IV) these are joined by a distinct set of mRNAs, which do not associate with the germ plasm but spread out evenly over a wider cortical region. The elaboration of the distinct animal and vegetal cortices continues throughout oogenesis as the oocyte accumulates yolk platelets, swelling enormously from less than 100 µm to over 1,200 µm in diameter. In the fully grown oocyte (Stage VI) the animal cortex is thickened and darkly pigmented, while the vegetal cortex is thin and transparent. On current evidence, mRNAs specific to the vegetal hemisphere tend to be associated with the cortex, whereas animal localised ones tend to be cytoplasmic (Hudson et al., 1996). Cortically-associated mRNAs and germ plasm are released to various extents from the vegetal cortex into the cytoplasm during meiotic maturation, and then further redistributed by post-fertilisation reorganisations. In particular, a process of ‘‘cortical rotation’’ is responsible for displacing dorso-anterior determinants from the vegetal pole to be activated at the equator of the egg. The entire cortex becomes displaced relative to the underlying cytoplasm by about 30°C about a horizontal axis (perpendicular to the plane of the page in Fig. 1). The direction of rotation and thus the equatorial site of dorsal determinant activation is not predetermined. At the end of the first cell cycle, successive ‘‘Surface Contraction Waves’’ (SCWs; Hara, 1971) that pass from the animal to vegetal poles of the egg at each mitosis, help to drive germ plasm islands towards the vegetal pole (Savage and Danilchik, 1993). Subsequently, subcortical ingression associated with cleavage furrow progression causes further redistribution of germ plasm and perhaps of other components deeper within the blastomeres of the early embryo (Danilchik and Denegre, 1991; Ressom and Dixon, 1988). The four cortical processes that modulate determinant localisation: localisation to the vegetal cortex during oogenesis, determinant release during maturation, cortical rotation, and the SCWs, will be discussed in more detail in the following sections. LOCALISATION OF DETERMINANTS ON THE OOCYTE VEGETAL CORTEX Germ Plasm Germ plasm is clearly implicated in germ line determination in Xenopus (Ikenishi, 1986), Drosophila (Gavis, 1997), and a number of other species from a wide range of phyla (Eddy, 1975; Wakahara, 1991; Ikenishi, 1998). The Xenopus mitochondrial cloud may be equivalent to similarly structured ‘‘sponge bodies’’ in Drosophila oocytes that also contain RNAs involved both in patterning the embryo and in germ line determination (WilschBrauninger et al., 1997). Molecular candidates for germ line determinant activity in Xenopus are emerging from a subset of identified localised mRNAs that associate with the mitochondrial cloud and subsequently with germ plasm: Xcat-2 (Mosquera et al., 1993) codes for a putative RNA-binding protein related to the Drosophila germ plasm component nanos, Xdaz1 is a functional homolog of Drosophila boule (Houston et al., 1998), and Xpat transcripts segregate with germ plasm and then with primordial germ cells during early development (Hudson and Woodland, 1998). Other mRNAs in the mitochondrial cloud, notably Xwnt11 (Ku and Melton, 1993), do not associate with the germ plasm and may play distinct patterning roles in the early embryo. It is possible that during metazoan evolution, germ line determinants were the first localised informational components in eggs (Denis, 1996, but see Dixon, 1994, for counter arguments). If germ plasm localisation does represent the original manifestation of oocyte polarity, molecules intervening in endoderm or mesendoderm development may subsequently have exploited the existing cortical localisation mechanisms to facilitate the organised development of associated tissues. The mechanism by which the mitochondrial cloud carries germinal granules and associated mRNAs to the cortex remains enigmatic, having proven refractile to cytoskeletal disruption (see Gard et al., 1995; Kloc et al., 1996). At around the time the mitochondrial cloud arrives, cortical microtubules are already present and the cytokeratin network is beginning to develop (Gard et al., 1997), with a loosely organised filament and ER network attached to the cortex (Figs. 2 and 3). Following the arrival of the mitochondrial cloud at the egg cortex, germinal granules disperse from it across the cortex and eventually assemble into distinct, mitochondria-rich, germ-plasm islands. During subsequent stages of oogenesis, the germ plasm material stays closely associated with the vegetal cortex. XENOPUS OOCYTE AND EGG CORTICES 417 Fig. 1. Summary of localisation events involving the cortex from oogenesis through to first cleavage in Xenopus. Dorso-Anterior Determinants There is clear experimental evidence that determinants necessary for the development of dorso-anterior embryonic structures are associated with the vegetal cortex of Xenopus oocytes and eggs. Removal of vegetal pole fragments from the egg prevents development of dorso-anterior sructures (Kikkawa et al., 1996; Sakai, 1996), while cytoplasm taken from close to the vegetal cortex (Fujisue et al., 1993; Holowacz and Elinson, 1993, 1995) is capable of directing the development of ectopic dorso-anterior structures if injected into other regions. In addition, vegetal cortices isolated from fertilised eggs and implanted into ectopic regions of the egg have been shown recently to harbour dorsalising activity (Kageura, 1998), substantiating the conclusions of earlier cortical grafting experiments (Curtis, 1960), which could not be interpreted clearly (Gerhart et al., 1981). The nature of the dorso-anterior determinants found on or close to the vegetal cortex is unresolved. They do not appear to act to induce mesoderm formation but rather to favour the development of dorsal structures in prospective mesoderm regions (Holowacz and Elinson, 1995), probably via the Wnt/ -catenin signal transduction pathway (Larabell et al., 1997; Marikawa et al., 1997; Rowning et al., 1997). Molecular components of dorso-anterior determinants are likely to be found amongst the ever-growing list of vegetally localised mRNAs, and/or the vegetal cortical proteins to which antibodies have been raised (Denegre et al., 1997). Certain of the known localised mRNAs appear to function in early patterning events although none has yet been clearly equated with dorso-anterior determinant activity: maternal VegT, a T-box containing transcription factor (Horb and Thomsen, 1997; Lustig et al., 1996; Stennard et al., 1996; Zhang and King, 1996) is involved in setting up the primary embryonic germ layers, being necessary for mesoderm and endoderm formation in the vegetal territories of the embryo (Horb and Thomsen, 1997; Zhang et al., 1998). Vg1, which codes for a TGF family growth factor able to induce mesoderm (Dale et al., 1993; Thomsen and Melton, 1993; Weeks and Melton, 1987), may be involved more specifically in formation and patterning of dorsal mesoderm and endoderm (Joseph and Melton, 1998). Vegetal Localised mRNAs The localised vegetal cortical RNAs identified so far can be divided into two sets on the basis of their time and pathway of localisation (Forristall et al., 1995; Kloc 418 P. CHANG ET AL. Fig. 2. General view of a replica from a deep-etched cortex prepared from a Stage I– II oocyte. Cortices were prepared directly on polylysine-coated glass chips from oocytes scalpeled in half in PHEM buffer (Schliwa et al., 1981) by a modification of the method of Elinson et al. (1993). After 15 minutes fixation in 1% glutaraldehyde in PHEM, the samples were fast frozen and deep-etched as described (Chang et al., 1996). A complex system of subcortical filaments (f) is present, interacting with frequent sheets of rough ER (rER). Large (0.7–0.8 µm) vesicular structures can be tentatively identified as cortical granules (cg), distributed unevenly through the subcortical cytoplasm, and the residual envelopes of yolk platelets (Y), membrane bound packets of pinocytosed vitellogenin. The asterisk marks the position of the stereo pairs shown in Figure 3. Magnification ⫻14,200 Fig. 3. A,B: High magnification stereo views of the Stage I-II oocyte cortex shown in Figure 2 (from region marked with asterisk). To appreciate fully the 3D view, inexpensive stereo viewers obtainable from Electron Microscopy supply companies should be used. Based on filament diameters, we can tentatively identify microtubules (mt), intermediate filaments (if), and maybe also microfilaments (mf). Coated pits (cp) are found at the plasma membrane (pm). Magnification ⫻71,000. 420 P. CHANG ET AL. and Etkin, 1995). A first set arrives in association with the mitochondrial cloud. This includes all the germ plasm associated mRNAs mentioned above along with Xwnt11 and Xlsirt, a non-coding, repetitive sequence RNA (Kloc and Etkin, 1994). Different transcripts take up different positions within the cloud, and appear to attach to the cortex in a temporal sequence so that they take on distinct distributions within the flattened ‘‘galette’’ that forms on the vegetal cortex (Kloc and Etkin, 1995; Kloc et al., 1996). The resultant layering of the messages (Xcat-2, then Xlsirt, then Xwnt11) is later lost as the mRNAs disperse more widely across the cortex (Kloc and Etkin, 1995). The second set of localised mRNAs includes Vg1 and VegT. These ‘‘late’’ localised mRNAs are synthesised in the very early oocyte but only concentrate to the vegetal cortex once the germ plasm localisation has been established, active localisation occurring in oocytes between stages II and IV. The transport mechanism used by the late mRNAs does not appear to be highly specific, since it can also function to localise inappropriate messages including the early mRNAs Xcat-2 (Zhou and King, 1996) and Xpat (Hudson and Woodland, 1998), and tau mRNA, which is localised in axons (Litman et al., 1996). These mRNAs all have ‘‘E2’’ motifs in the 3’UTR, which recognise the ER-associated RNA binding protein Vera (Deshler et al., 1997, 1998). Additional elements in the Vg1 3’UTR appear to have overlapping roles in localisation, binding Vera and other cytoplasmic proteins. This suggests that complexes containing multiple proteins associate with the RNAs prior to localisation, distinct proteins mediating specificity or localisation pathways (Gautreau et al., 1997; Mowry, 1996). The first step in localisation of late mRNAs is their accumulation in a wedge-shaped cytoplasmic region between the GV and the newly established germ plasm patch on the cortex. This region is rich in Vera, apparently associating with a subpopulation of the oocyte’s ER (Deshler et al., 1997; Kloc and Etkin, 1998). The RNAs subsequently accumulate on the vegetal cortex, and then spread laterally to cover a larger area than that of the early RNAs. Large flat ER sheets and tubules are predominant features of the cortex at this time (Fig. 4A), appearing to fuse directly to the plasma membrane as well as being tethered by cytokeratin-containing filaments (Fig. 4B). Localisation of Vg1 to the vegetal pole, and perhaps the initial steps in its cortical anchoring, depend on microtubules (Kloc and Etkin, 1995; Yisraeli et al., 1989). Transport may be mediated by direct binding of RNP particles to microtubules via the Vg1 binding protein Vg1RBP (Elisha et al., 1995) either in combination with the ER and Vera or in a parallel pathway. Vg1RBP is highly homologous to a protein implicated in localisation of actin mRNA in fibroblasts, a process mediated by microfilaments rather than microtubules (Havin et al., 1998), suggestive that both cytoskeletal systems can participate in mRNA localisation. The polarity of the cytoplasmic microtubules between the oocyte nucleus and the cortex is not known so it is hard to predict which kinds of motor molecules might be involved in message transport. Microtubule regrowth experiments suggest that minus ends are likely to lie at the base of the nucleus (Gard, 1991), while the specific detection of ␥-tubulin at the vegetal cortex implicates microtubule nucleation in the opposite direction (Gard, 1994; see also Gard, this issue). The anchorage of vegetally localised mRNAs to the cortex is distinguishable from transport since it is not disrupted by microtubule depolymerizing drugs (Kloc and Etkin, 1995; Yisraeli et al., 1989, 1990). Unsurprisingly, given the major role of actin in maintaining cortical integrity, cytochalasin treatment, was found to release Vg1 message from the cortex into the vegetal cytoplasm (Yisraeli et al., 1989). In another study (Kloc and Etkin, 1995), Xlsirt, Xcat-2, and Xwnt11 as well as Vg1 RNAs were detached from the cortex of stage III and IV oocytes by cytochalasin treatment, but appeared to remain associated in a disc that delaminated from the cortex. More drastic disruption of cortical anchoring of Vg1 mRNA has been achieved with oligonucleotides targeted to destroy Xlsirt RNAs (Kloc and Etkin, 1994). These repetitive sequence RNAs, which arrive with the mitochondrial cloud (Kloc et al., 1993), are proposed to play a structural role in the cortical anchoring of Vg1, but not Xcat-2 mRNAs (Kloc and Etkin, 1994). Although Vg1 mRNA associates with an IF-rich cellular residue upon detergent extraction (Pondel and King, 1988), manipulation of meiotic maturation suggests that Vg1 mRNA is not directly linked to cortical cytokeratin filaments (Klymkowsky and Maynell, 1989; Klymkowsky et al., 1991; see Klymkowsky, 1995). Studies of whole stage VI oocytes (Canman and Bement, 1997; Gard et al., 1997) and cortices isolated from them, which retain the localised mRNAs (Elinson et al., 1993), indicate that all three filament systems, as well as mitochondria and extensive ER interact in the oocyte cortex (Figs. 5 and 6) (Elinson et al., 1993). Microtubules and microfilaments appear to direct cytokeratin filament organisation (Gard et al., 1997), with the IF bundles more tightly organised in the vegetal hemisphere (Klymkowsky et al., 1987). MODIFICATION OF DETERMINANT AND mRNA ATTACHMENT DURING MEIOTIC MATURATION UV irradiation of the vegetal cortex of the immature Stage VI oocyte, but not of the unfertilised egg, significantly reduces dorso-anterior determinant activity, suggesting that the dorso-anterior determinants are released from the cortex and/or transformed during meiotic maturation (Elinson and Pasceri, 1989; Holowacz and Elinson, 1993). In contrast, UV irradiation of oocyte and egg vegetal surfaces affects germ cell formation to similar degrees (Holwill et al., 1987). Dorso-anterior determinants may move deeper into the cytoplasm during maturation, but are not lost from the cortical/subcortical region since they relocate with the vegetal cortex upon experimental 180° rotation of the egg (Marikawa et al., 1997). Vg1 mRNA is released into the vegetal cytoplasm from its tight cortical association early during meiotic maturation, whereas Xcat-2 remains associated with the cortical germ plasm throughout meiosis (Mosquera et al., 1993; Forristall et al., 1995; Weeks and Melton, 1987). The small islands of mitochondria and germinal granules present beneath the vegetal oocyte surface (Heasman et al., 1984; Savage and Danilchik, 1993) coalesce during oocyte maturation to form more clearly distinct germ plasm Fig. 4. Deep-etched cortex prepared from a Stage III oocyte as described in Figure 1. Immunolabeling of IFs with anticytokeratin monoclonal 1h5 (Klymkowsky et al., 1987) was performed as described (Chang et al., 1996). A: General view, magnification ⫻28,500. B: High magnification stereo view from a different area of the same cortex. Magnification ⫻99,500. A large tubular patch of rough ER (rER) is connected to the plasma membrane (pm) both directly (large arrowhead) and via filaments lableled with the 1h5 antibody (arrows indicate gold-liganded secondary antibody). Annotations as in Figures 2 and 3. Note the rER lumen (lumen), and the wall (w) of rER tethered by filaments (f) to the plasma membrane. Fig. 5. General view of a deep-etched cortex prepared from the animal hemisphere of a Stage VI oocyte. Abundant intermediate-sized filaments run in wide loose bundles (large arrows) across the cytoplasmic side of a complex thick layer containing cortical granules (cg). Magnification ⫻14,200. Annotations as in Figures 2 and 3. Fig. 6. General view of a deep-etched cortex prepared from the vegetal hemisphere of the same St VI oocyte as shown in Figure 5. The IF bundles (large arrows) are more compact and appear to run between the cortical granules (cg), closer to the plasma membrane (pm) than in the animal half. Magnification ⫻14,200. Annotations as in Figures 2 and 3. 424 P. CHANG ET AL. islands (Czolowska, 1969) but remain close to the vegetal surface. Determinant/localised mRNA dispersal during oocyte maturation accompanies extensive remodelling of the whole egg, including the cortex. Some structural changes reflect the change in cell cycle state from interphasic (prophase of meiosis I) to mitotic (metaphase of meiosis II), while others are related to the change from the stable organisation of the stage VI oocyte, which may remained unchanged for months in the ovary, to one that is unstable and poised for activation by the fertilising sperm. The most striking change to the cortex during maturation is the setting up of a specialised cortical endoplasmic reticulum (cER), most highly developed in the animal hemisphere and implicated in the fertilisation response (Campanella et al., 1984; Charbonneau and Grey, 1984; Dersch et al., 1991; Stewart-Savage et al., 1991). This extensive cER network is established very close to, and connecting with, the plasma membrane (Gardiner and Grey, 1983), now enveloping cortical granules that reorganise from a deep to a regular array very close to the plasma membrane (Campanella et al., 1984). Remodelling of the actin cortex occurs in parallel with the setting up of the cortical granule-associated cER, manifest as the development of contractability, inducible by fertilisation or calcium ionophore. The robust contraction of the animal cortex of the unfertilised egg cannot be prevented with cytochalasins (Merriam and Sauterer, 1983), suggesting that an extensive actomyosin filament system has been set up below the animal surface (Merriam et al., 1983; see Larabell, 1995). Vinculin is also recruited into the cortex during the period of oocyte maturation and oviposition (Evans et al., 1990). Cortical cytokeratin filaments throughout the egg are also extensively remodelled during meiotic maturation (Klymkowsky, 1995). In the animal hemisphere, there is a brief assembly of IF cable network, less developed in this region of the Stage VI oocyte, then a disassembly to phosphorylated oligomers induced indirectly by activation of the universal mitotic kinase MPF (Klymkowsky and Maynell, 1989; Klymkowsky et al., 1991). In the vegetal hemisphere, the preformed cytokeratin cable network disintegrates completely. The release of Vg1 mRNA from the cortex precedes this IF disassembly and can be distinguished from it experimentally (Klymkowsky et al., 1991). Thus, Vg1 mRNA anchoring to the vegetal cortex appears not to be simply dependent on any one of the cytoskeletal systems, but probably on a combination of these, and/or other as yet unanalysed structural components. CORTICAL ROTATION Cortical rotation is necessary for the development of dorso-anterior structures in the frog embryo (Gerhart et al., 1989). During the cortical rotation, dorsoanterior determinants relocate from around the vegetal pole to a broad area spreading toward the equator in the direction of cortical movement (Fujisue et al., 1993; Kikkawa et al., 1996; Sakai, 1996). This relocation is necessary to allow the determinants to activate certain downstream targets in prospective mesoderm regions that stimulate ‘‘organiser’’ activity (Darras et al., 1997). Whether determinant substances are carried with the cortex or translocated independently along aligned subcortical microtubules on vesicles or particles (Rowning et al., 1997) is not known. The mechanism of cortical rotation depends on interaction between subcortical microtubules and elements anchored to the cortex proper. Prior to the cortical rotation, microtubules tend to grow outwards from the center of the egg (Houliston and Elinson, 1991b; Elinson and Palacek, 1993). The interaction of these microtubules with the vegetal cortex, the region where the rotation movement is likely to be generated (see Houliston and Elinson, 1992), is different from that at the animal cortex (Houliston and Elinson, 1991b; Schroeder and Gard, 1992). In the animal hemisphere, outwardgrowing microtubules abut the thickened, pigmented cortex abruptly with minimal bending. This behaviour, and/or the relative fluidity of the animal cytoplasm, may be important to allow sperm aster expansion to drive pronuclear migration earlier in the cell cycle (see Reinsch and Gönczy, 1998). In contrast, microtubules that cross the dense vegetal yolk mass turn in the yolk-poor subcortical vegetal cytoplasm and continue to polymerise parallel to the vegetal cortex. Differences in the interactions of microtubules with the animal and vegetal cortices have also been demonstrated by observing ectopic meiotic spindle formation and rotation (Gard, 1993). Outward-growing vegetal microtubules, together with ones nucleated locally in the vegetal cortical region, form a striking array of aligned microtubules covering the whole vegetal surface of the egg (Elinson and Rowning, 1988). This vegetal subcortical microtubule array is strongly implicated in the process of cortical rotation, and the cortical rotation movement and the alignment of the microtubules mutually reinforce one another (see Houliston and Elinson, 1992; Gerhart et al., 1989). The aligned vegetal subcortical microtubules stay attached to experimentally isolated cortices and can thus be visualised in deep etch replicas, where they are seen to be sandwiched in layers between extensive sheets of rough ER (Fig. 7; Chang et al., 1996). This observation reinforces the hypothesis that ER could provide the cortical substrate for microtubule motor attachment to create the cortical rotation movement. Interactions generating the rotation movement could also occur directly between subcortical microtubules and cortical actin, or between microtubules and associated cytokeratin filaments, although the latter possibility seems less likely since cytokeratin filaments are sparse at this time and are mainly located towards the cytoplasmic side of the subcortical microtubule layer (Houliston and Elinson, 1991a). Various observations suggest that microtubule motor(s) cause the cortical rotation by moving cortically attached elements towards the plus-ends of microtubules: the uniform polarity of the microtubules in the direction of cortical movement (Houliston and Elinson, 1991a), the observed displacement of the majority of vegetal subcortical microtubules relative to the cortex (Houliston, 1994; Larabell et al., 1996), and the independence of cortical rotation from continuing microtubule polymerisation (Houliston, 1994). Immunofluorescence studies on isolated cortices indicate that the architypical plus-end directed motor kinesin is associated with the aligned subcortical vegetal microtubules and ER, and so could XENOPUS OOCYTE AND EGG CORTICES contribute to the cortical movement by generating movement between these structures (Houliston and Elinson, 1991a). In contrast, immunogold localisation of the abundant plus-end directed motor protein Eg5 on replicas of cortices isolated during the cortical rotation revealed an exclusive association with microtubules and microtubule bundles (Chang et al., 1996), despite indications from whole mount immunofluorescence that Eg5 might also be associated with the ER (Houliston et al., 1994). Thus, if Eg5 contributes to the rotation movement, it must do so by some sort of microtubulemicrotubule sliding mechanism (Larabell et al., 1996). High-resolution localisation of kinesin and the many other kinesin-like proteins present in Xenopus eggs has yet to be accomplished. To understand the mechanism of cortical rotation it will be necessary to determine which microtubule subpopulations are involved in force generation, as well as to identify the motor molecules(s) utilized. SURFACE CONTRACTION WAVES AND CORTICAL CONTRACTIONS Germ plasm loses its tight cortical association following fertilisation but remains in a subcortical band 25–30 µm thick (Ressom and Dixon, 1988). This moves with the vegetal cortex if it is displaced animally by 180° rotation, disrupting subsequent formation and migration of primary germ cells (Cleine and Dixon, 1985). Another indication that the germ plasm remains under the influence of the cortex, or at least of organisational changes in the subcortical cytoplasm, is that the SCWs of successive mitotic cycles participate in moving the germ plasm islands toward the vegetal pole (Savage and Danilchik, 1993). The SCWs traverse the egg from animal to vegetal poles at the time of each mitosis (Hara, 1971; Yoneda et al., 1982; Sawai, 1982). There are two distinct SCWs in each cell cycle. The first appears to be a wave of general relaxation of the egg rather than a contraction. It coincides with the activation of MPF at the onset of mitosis. MPF activation is initiated in the animal cytoplasm and propagates by a post-translational autocatalytic activation mechanism to the vegetal pole (Pérez-Mongiovi et al., 1998; Rankin and Kirschner, 1997). The first SCW appears, at least in part, to reflect depolymerisation of subcortical microtubules as a consequence of the wave of MPF activation (Pérez-Mongiovi et al., 1998; Schroeder and Gard, 1992). MPF activation in the vegetal hemisphere coincides with the end of the cortical rotation, which stops precisely as the first SCW passes. Microtubule depolymerisation provoked by the MPF activation wave would explain the influence of the first SCW on the germ plasm, since many aspects of germ plasm redistribution following fertilisation are sensitive to microtubule depolymerisation (Ressom and Dixon, 1988; Savage and Danilchik, 1993). Germ plasm aggregates are known to associate with subcortical microtubules via the kinesin-like motor protein Xklp1 (Robb et al., 1996). It has been proposed that the first SCW may, like the second SCW (see below), depend on a propagated wave of calcium release (Jaffe, 1999). Although calcium transients are implicated in both activation and inactivation of MPF in Xenopus egg cytoplasm (Lindsay et al., 1995), no calcium waves have yet been visualised in the cortex concurrent with the first SCW. 425 The second SCW, which immediately precedes the advancing cleavage furrow, is a true cortical contraction wave. Its progress coincides with the passage of MPF inactivation across the egg (Pérez-Mongiovi et al., 1998; Rankin and Kirschner, 1997). The mechanism of the second SCW is clearly likely to be closely tied in with the actin-based contraction that drives the cleavage furrow. Increased contractility of actin in the cortex at the end of mitosis is thought to involve actomyosin interactions favoured by phosphorylation of myosin light chain kinase upon MPF inactivation (Satterwhite et al., 1992). Intracellular calcium release appears likely to play a role in cleavage furrow progression and the second SCW, since localised calcium release has been detected at the time of the second SCW, associated with the furrow (Keating et al., 1994; Muto et al., 1996). A more experimentally tractable actin-based cortical contraction induced by cytoplasmic calcium release occurs following fertilisation in amphibian eggs (Merriam and Sauterer, 1983). This violent isotropic contraction of the actin-thickened animal cortex, which drags the entering sperm nucleus toward the animal pole, has been shown to represent a cortical response to the massive IP3-mediated wave of cytoplasmic calcium release triggered by the fertilising sperm (Kubota et al., 1987; Larabell and Nuccitelli, 1992; see Elinson, 1980; Larabell, 1995). The wave of calcium release from the cER following fertilisation also promotes exocytosis of cortical granules (Andreuccetti et al., 1984). Like the second SCW, the fertlisation-triggered cortical contraction accompanies MPF inactivation, in this case the release from meiotic metaphase arrest. Waves of peristaltic cortical contraction accompanied by calcium transients can also be triggered artificially during interphase by microinjection of IP3 (Muto and Mikoshiba, 1998). Germ plasm movement accompanying the second SCW is less distinct than at the first SCW and difficult to discriminate from cleavage-associated movements. The participation of the cortex in determinant localisation can be considered to be essentially complete by the time of first cleavage, as germ plasm and probably the dorso-anterior determinants move into subcortical and deeper cytoplasm, eventually reaching nuclei of different embryonic blastomeres to have their developmental effects. PERSPECTIVES Despite the wealth of descriptions and functional studies concerning the organisation of different components of the Xenopus egg cortex at different times, there remain large gaps in our knowledge. It is difficult to integrate the different pieces of published information and to form a full or clear picture of the structure of this dynamic region at the times when determinants are translocated, anchored or released. With the growing realisation that different cytoskeletal systems do not function independently but interact intimately (Canman and Bement, 1997; Gard et al., 1997), we must consider that the distribution of motor molecules, membrane systems, and determinants is likely to depend on all different types of filaments as well as the regulation of their interaction by changing cytoplasmic conditions, for instance during cell cycle transitions. Further structural studies of the cortex, as well as, of course, the molecular characterisation of the dorso-anterior and Fig. 7. View of a deep-etched cortex isolated manually from a fertilised egg during cortical rotation and immunolabelled with antitubulin antibody YL1/2 (Kilmartin et al., 1982) and subsequent goat anti-rat liganded with 5-nm gold particles prior to freezing and replica preparation (methods described in Chang et al., 1996). Bundled microtubules (arrowheads) and microtubules (small arrows) can be seen. Magnification ⫻28,500. Annotations as in Figures 2 and 3. XENOPUS OOCYTE AND EGG CORTICES germ line determinants and the analysis of their localisation at the ultrastructural level, will be required to unravel this problem. ACKNOWLEDGMENTS Anti-cytokeratin monoclonal 1h5 developed by M. Klymkowsky was obtained from the Developmental Studies Hybridoma Bank maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA 52242 under contract N01-HD-7–3263 from the NICHD. We thank Clare Hudson (Marseille) for useful hints about mRNA localisation sequences and Christian Sardet for support. The original research described was funded by ARC grants 6985 and 9144 to E.H, and by financement from the CNRS and Ministère d’Education (ACC4/ AI ‘‘Biocell’’). REFERENCES Al-Mukhtar KAK, Webb AC. 1971. An ultrastructural study of primordial germ cells, oogonia and early oocytes in Xenopus laevis. J Embryol Exp Morphol 26:195–217. Andreuccetti P, Denis-Donini S, Burrini A, Campanella. 1984. Calcium ultrastructural localization in Xenopus laevis eggs following activation by pricking or by calcium ionophore A 23187. J Exp Zool 229:295–308. Campanella C, Reuccetti P, Taddei C, Talevi R. 1984. The modifications of cortical endoplasmic reticulum during in vitro maturation of Xenopus laevis oocytes and its involvement in cortical granule exocytosis. J Exp Zool 229:283–93. Canman JC, Bement W M. 1997. Microtubules suppress actomyosinbased cortical flow in Xenopusoocytes. J Cell Sci 110:1907–1917. Chang P, LeGuellec K, Houliston E. 1996. Immunodetection of cytoskeletal structures and the Eg5 motor protein on deep-etch replicas of Xenopus egg cortices isolated during the cortical rotation. Biol Cell 88:89–98. Charbonneau M, Grey R. 1984. The onset of activation responsiveness during maturation coincides with the formation of the cortical endoplasmic reticulum in oocytes of Xenopus laevis. Dev Biol 102:90–97. Cleine JH, Dixon K E. 1985. The effect of egg rotation on the differentiation of primordial germ cells in Xenopus laevis. J Embryol Exp Morphol 90:79–99. Curtis ASG. 1960. Cortical grafting in Xenopus laevis. J Embryol Exp Morphol 8:163–173. Czolowska R 1969. Observations on the origin of the ‘‘germinal cytoplasm’’ inXenopus laevis. J Embryol Exp Morphol 22:229–51. Dale L, Matthews G, Colman A. 1993. Secretion and mesoderminducing activity of the TGF--related domain of Xenopus Vg1. EMBO J 12:4471–4480. Danilchik M, Denegre J. 1991. Deep cytoplasmic rearrangements during early development in Xenopus laevis. Development 111:845– 856. Darras S, Marikawa Y, Elinson R P, Lemaire P. 1997. Animal and vegetal pole cells of early Xenopus embryos respond differently to maternal dorsal determinants: implications for the patterning of the organiser. Development 124:4275–4286. Denegre JM, Ludwig ER, Mowry KL. 1997. Localized maternal proteins in Xenopus revealed by subtractive immunisation. Dev Biol 192:446–454. Denis H. 1996. Cytosquelette et polarité ovulaire. Méd/Sci 12:1145– 1158. Dent JA, Klymkowsky MW. 1989. Whole mount analyses of cytoskeletal reorganisation and function during oogenesis and embryogenesis in Xenopus. In: Schatten H, Schatten G, editors. The cell biology of fertilization. New York: Academic Press, p 63–103. Dersch MA, Bement WM, Larabell CA, Mecca MD, Capco DG. 1991. Cortical membrane-trafficking during the meiotic resumption of Xenopus laevis oocytes. Cell Tissue Res 263:375–383. Deshler J, Highett M, Schnapp B. 1997. Localization of Xenopus Vg1 mRNA by Vera protein and the endoplasmic reticulum. Science 276:1128–31. Deshler JO, Highett MI, Abramson T, Schnapp BJ. 1998. A highly conserved RNA-binding protein for cytoplasmic mRNA localisation in vertebrates. Curr Biol 8:489–496. Dixon KD. 1994. Evolutionary aspects of primordial germ cell forma- 427 tion. In: Germline development (Ciba Foundation symposium). Chichester: Wiley, p 92–120. Eddy EM. 1975. Germ plasm and the differentiation of the germ cell line. Int Rev Cytol 43:229–281. Elinson RP. 1980. The amphibian egg cortex in fertilization and early development. Symp Soc Dev Biol 38:217–234. Elinson R P. 1990. Cytoskeleton and embryo polarity. Curr. Opin. Cell Biol, 2:75–79. Elinson RP, Houliston E. 1990. Cytoskeleton in Xenopus oocytes and eggs. Semin Cell Biol 1:349–357. Elinson RP, Palacek J. 1993. Independence of microtubule systems in fertilized frog eggs: the sperm aster and the vegetal parallel array. Roux’s Arch Dev Biol 202:224–232. Elinson RP, Pasceri P. 1989. Two UV-sensitive targets in dorsoanterior specification of frog embryos. Development 106:511–518. Elinson RP, Rowning B. 1988. A transient array of parallel microtubules in frog eggs: potential tracks for a cytoplasmic rotation that specifies the dorso-ventral axis. Dev Biol 128:185–197. Elinson RP, King ML, Forristall C. 1993. Isolated vegetal cortex from Xenopus oocytes selectively retains localized mRNAs. Dev Biol 160:554–562. Elisha Z, Havin L, Ringel I, Yisraeli JK. 1995. Vg1 RNA binding protein mediates the association of Vg1 RNA with microtubules in Xenopus oocytes. EMBO J 14:5109–5114. Evans JP, Page BD, Kay BK. 1990. Talin and vinculin in the oocytes, eggs and early embryos of Xenopus laevis: a developmentally regulated change in distribution. Dev Biol 137:403–413. Forristall C, Pondel M, King ML. 1995. Patterns of localization and cytoskeletal association of two vegetally localized RNAs Vg1 and Xcat–2. Development 121:201–208. Franke WW, Rathke PC, Seib E, Trendelenburg MF, Osborne M, Weber K. 1976. Distribution and mode of arrangement of microfilamentous structures and actin in the cortex of the amphibian oocyte. Cytobiologie 14:111–130. Fujisue M, Kobayakawa Y, Yamana K. 1993. Occurrence of dorsal axis-inducing activity around the vegetal pole of an uncleaved Xenopus egg and displacement to the equatorial region by cortical rotation. Development 118:163–170. Gard DL. 1991. Organization, nucleation, and acetylation of microtubules in Xenopus laevis oocytes: a study by confocal immunofluorescence microscopy. Dev Biol 143:346–362. Gard DL. 1993. Ectopic spindle assembly during maturation of Xenopus oocytes: evidence for functional polarization of the oocyte cortex. Dev Biol 159:298–310. Gard DL. 1994. Gamma-tubulin is asymmetrically distributed in the cortex of Xenopus oocytes. Dev Biol 161:131–140. Gard DL, Cha BJ, Schroeder MM. 1995. Confocal immunofluorescence microscopy of microtubules, microtubule-associated proteins, and microtubule-organizing centers during amphibian oogenesis and early development. Curr Top Dev Biol 31:383–431. Gard DL, Cha BJ, King E. 1997. Organisation and animal-vegetal asymmetry of cytokeratin filaments in Stage VI Xenopus oocytes is dependent upon F-actin and microtubules. Dev Biol 184:94–114. Gardiner DM, Grey RD. 1983. Membrane junctions in Xenopus eggs: their distribution suggests a role in calcium regulation. J Cell Biol 96:1159–1163. Gautreau D, Cote C, Mowry K. 1997. Two copies of a subelement from the Vg1 RNA localization sequence are sufficient to direct vegetal localization in Xenopus oocytes. Development 124:5013–5020. Gavis ER. 1997. Expeditions to the pole: RNA localisation in Xenopus and Drosophila.Trends Cell Biol 7:485–492. Gerhart JC, Ubbels G, Black S, Hara K, Kirschner M. 1981. A reinvestigation of the role of the grey crescent in axis formation in Xenopus laevis. Nature (Lond) 292:511–516. Gerhart J, Danilchik M, Doniach T, Roberts S, Rowning B, Stewart R. 1989. Cortical rotation of the Xenopus egg: consequences for the anteroposterior pattern of embryonic dorsal development. Development 107:37–51. Hara K. 1971. Cinematographic observation of ‘‘surface contraction waves’’ (SCW) during the early cleavage of axolotl eggs. Wilhelm Roux Arch Entw Mech Org 167:183–186. Havin L, Git A, Elisha Z, Oberman F, Yaniv K, Pressmen Schwartz S, Standart N, Yisraeli JK. 1998. RNA-binding protein conserved in both microtubule- and microfilament-based RNA localization. Genes Dev 12:1593–1598. Heasman J, Quarmby J, Wylie CC. 1984. The mitochondrial cloud of Xenopus oocytes: the source of germinal granule material. Dev Biol 105:458–469. Holowacz T, Elinson R. 1993. Cortical cytoplasm, which induces dorsal axis formation in Xenopus, is inactivated by UV irradiation of the oocyte. Development (Suppl) 119:277–285. 428 P. CHANG ET AL. Holowacz T, Elinson RP. 1995. Properties of the dorsal activity found in the vegetal cortical cytoplasm of Xenopus eggs. Development 121:2789–2798. Holwill S, Heasman J, Crawley CR, Wylie CC. 1987. Axis and germ line deficiencies caused by u.v. irradiation of Xenopus oocytes cultured in vitro. Development 100:735–743. Horb M E, Thomsen GH. 1997. A vegetally localized T-box transcription factor in Xenopuseggs specifies mesoderm and endoderm and is essential for embryonic mesoderm formation. Development 124: 1689–98. Houliston E. 1994. Microtubule translocation and polymerisation during cortical rotation in Xenopus eggs. Development 120:1213– 1220. Houliston E, Elinson RP. 1991a. Evidence for the involvement of microtubules ER, and kinesin in the cortical rotation of fertilized frog eggs. J Cell Biol 114:1017–1028. Houliston E, Elinson RP. 1991b. Patterns of microtubule polymerization relating to cortical rotation in Xenopus laevis eggs. Development 112:107–117. Houliston E, Elinson RP. 1992. Microtubules and cytoplasmic reorganization in the frog egg. Curr Top Dev Biol 26:53–70. Houliston E, Le Guellec R, Kress M, Philippe M, Le Guellec K. 1994. The kinesin-related protein Eg5 associates with both interphase and spindle microtubules during Xenopus early development. Dev Biol 164:147–159. Houston D W, Zhang J, Maines J Z, Wasserman S A, King M L. 1998. A XenopusDAZ-like gene encodes an RNA component of germ plasm and is a functional homologue of Drosophila boule. Development, 125:171–180. Hudson C, Woodland H. 1998. Xpat, a gene expressed specifically in germ plasm and primordial germ cells of Xenopus laevis. Mech Dev 73:159–68. Hudson JW, Alarcon VB, Elinson RP. 1996. Identification of new localized RNAs in the Xenopus oocyte by differential display PCR. Dev Genet 19:190–198. Ikenishi K. 1986. Direct evidence for the presence of germ cell determinant in vegetal pole cytoplasm of Xenopus laevis and in a subcellular fraction of it. Dev Growth Differ 28:563–568. Ikenishi K. 1998. Germ plasm in Caenorhabditis elegans, Drosophila and Xenopus. Dev Growth Differ 40:1–10. Jaffe LF. 1998. Calcium patterns and development. BioEssays. Joseph EM, Melton DA. 1998. Mutant Vg1 ligands disrupt endoderm and mesoderm formation in Xenopus embryos. Development 125: 2677–2685. Kageura H. 1998. Activation of dorsal development by contact between the cortical dorsal determinant and the equatorial core cytoplasm in eggs of Xenopus laevis. Development, 124:1543–1551. Keating TJ, Cork RJ, Robinson KR. 1994. Intracellular free calcium oscillations in normal and cleavage-blocked embryos and artificially activated eggs of Xenopus laevis. J Cell Sci 107:2229–2237. Kikkawa M, Takano K, Shinagawa A. 1996. Location and behavior of dorsal determinants during first cell cycle in Xenopus eggs. Development 122:3687–3696. Kilmartin JV, Wright B, Milstein C. 1982. Rat monoclonal anti-tubulin antibodies derived by using a new nonsecreting rat cell line. J Cell Biol 93:576–582. Kloc M, Etkin LD. 1994. Delocalization of Vg1 mRNA from the vegetal cortex in Xenopus oocytes after destruction of Xlsirt RNA. Science 265:1101–1103. Kloc M, Etkin LD. 1995. Two distinct pathways for the localization of RNAs at the vegetal cortex in Xenopus oocytes. Development 121:287–297. Kloc M, Etkin LD. 1998. Apparent continuity between the messenger transport organiser and late RNA localisation pathways during oogenesis in Xenopus. Mech Dev 73:95–106. Kloc M, Spohr G, Etkin LD. 1993. Translocation of repetitive RNA sequences with the germ plasm in Xenopus oocytes. Science 262: 1712–1714. Kloc M, Larabell C, Etkin LD. 1996. Elaboration of the messenger transport organizer pathway for localization of RNA to the vegetal cortex of Xenopus oocytes. Dev Biol 180:119–130. Klymkowsky MW. 1995. Intermediate filament organisation, reorganisation and function in the clawed frog Xenopus. Curr Top Dev Biol 31:455–485. Klymkowsky MW, Karnovsky A. 1994. Morphogenesis and the cytoskeleton: Studies of the Xenopus embryo. Dev Biol 165:372–384. Klymkowsky MW, Maynell LA. 1989. MPF-induced breakdown of cytokeratin filament organization in the maturing Xenopus oocyte depends on the translation of maternal RNAs. Dev Biol 134:479– 485. Klymkowsky MW, Maynell LA, Polson AG. 1987. Polar asymmetry in the organization of the cortical cytokeratin system of Xenopus laevis oocytes and embryos. Development 100:543–557. Klymkowsky MW, Maynell LA, Nislow C. 1991. Cytokeratin phosphorylation, cytokeratin filament severing and the solubilization of the maternal mRNA Vg1. J Cell Biol 114:787–797. Ku M, Melton DA. 1993. Xwnt–11: a maternally expressed Xenopus wnt gene. Development 119:1161–1173. Kubota HY, Yoshimoto Y, Yoneda M, Hiramoto Y. 1987. Free calcium wave upon activation in Xenopus eggs. Dev Biol 119:129–136. Larabell CA. 1995. Cortical cytoskeleton of the Xenopus oocyte, egg, and early embryo. Curr Top Dev Biol 31:433–453. Larabell C, Nuccitelli R. 1992. Inositol lipid hydrolysis contributes to the Ca2⫹ wave in the activating egg of Xenopus laevis. Dev Biol 153:347–55. Larabell CA, Rowning BA, Wells J, Wu M, Gerhart JC. 1996. Confocal microscopy analysis of living Xenopus eggs and the mechanism of cortical rotation. Development 122:1281–1289. Larabell CA, Torres M, Rowning BA, Yost C, Miller JR, Wu M, Kimelman D, Moon RT. 1997. Establishment of the dorso-ventral axis in Xenopus embryos is presaged by early asymmetries in beta-catenin that are modulated by the Wnt signaling pathway. J Cell Biol 136:1123–1136. Lindsay HD, Whitaker MJ, Ford CC. 1995. Calcium requirements during mitotic cdc2 kinase activation and cyclin degradation in Xenopus egg extracts. J Cell Sci 108:3557–3568. Litman P, Behar L, Elisha Z, Yisraeli JK, Ginzburg I. 1996. Exogenous tau RNA is localized in oocytes: possible evidence for evolutionary conservation of localization mechanisms. Dev Biol 176:86–94. Lustig KD, Kroll KL, Sun EE, Kirschner MW. 1996. Expression cloning of a Xenopus T-related gene (Xombi) involved in mesodermal patterning and blastopore lip formation. Development 122:4001– 4012. Marikawa Y, Li Y, Elinson RP. 1997. Dorsal determinants in the Xenopus egg are firmly associated with the vegetal cortex and behave like activators of the Wnt pathway. Dev Biol 191:69–79. Merriam RW, Sauterer R. 1983. Localization of a pigment containing structure near the surface of Xenopus eggs which contracts in response to calcium. J Embryol Exp Morphol 76:51–65. Merriam RW, Sauterer R, Christensen K. 1983. A subcortical, pigmentcontaining structure in Xenopus eggs with contractile properties. Dev Biol 95:437–446. Micklem DR. 1995. mRNA localisation during development. Dev Biol 172:377–395. Mosquera L, Forristall C, Zhou Y, King ML. 1993. A mRNA localised to the vegetal cortex of Xenopus oocytes encodes a protein with a nanos-like zinc finger domain. Development 117:377–386. Mowry K. 1996. Complex formation between stage-specific oocyte factors and a Xenopus mRNA localization element. Proc Natl Acad Sci USA 93:14608–13. Muto A, Mikoshiba K. 1998. Activation of Inositol 1,4,5-triphosphate receptors induces transient changes in cell shape of fertilised Xenopus eggs. Cell Motil Cytoskeleton 39:201–208. Muto A, Kume S, Inoue T, Okano H, Mikoshiba K. 1996. Calcium waves along the cleavage furrows in cleavage-stage Xenopus embryos and its inhibition by heparin. J Cell Biol 135:181–190. Pérez-Mongiovi D, Chang P, Houliston E. 1998. A propagated wave of MPF activation accompanies surface contraction waves at first mitosis in Xenopus. J Cell Sci 111:385–393. Pondel MD, King M L. 1988. Localized maternal mRNA related to transforming growth factor mRNA is concentrated in a cytokeratinenriched fraction from Xenopus oocytes. Proc Natl Acad Sci USA 85:7612–7616. Rankin S, Kirschner MW. 1997. The surface contraction waves of Xenopus eggs reflect the metachronous cell-cycle state of the cytoplasm. Curr Biol 7:451–454. Reinsch S, Gönczy P. 1998. Mechanisms of nuclear positioning. J Cell Sci 111:2283–2295. Ressom RE, Dixon K E. 1988. Relocation and reorganization of germ plasm in Xenopus embryos after fertilization. Development 103:507– 518. Robb DL, Heasman J, Raats J, Wylie C. 1996. A kinesin-like protein is required for germ plasm aggregation in Xenopus. Cell 87:823–831. Roeder AD. Roeder AD, Gard DL. 1994. Confocal microscopy of F-actin distribution in Xenopus oocytes. Zygote 2:111–124. Rowning BA, Wells J, Wu M, Gerhart JC, Moon RT, Larabell CA. 1997. Microtubule-mediated transport of organelles and localization of XENOPUS OOCYTE AND EGG CORTICES beta-catenin to the future dorsal side of Xenopus eggs. Proc Natl Acad Sci USA 94:1224–1229. Sakai M. 1996. The vegetal determinants required for the Spemann organiser move equatorially during the first cell cycle. Development 122:2207–2214. Sardet C, Chang P. 1987. The egg cortex: from maturation through fertilization. Cell Differ 21:1–19. Sardet C, McDougall A, Houliston E. 1994. Cytoplasmic domains in eggs. Trends Cell Biol 4:166–172. Satterwhite LL, Lohka MJ, Wilson KL, Scherson TL, Cisek LJ, Cordu JL, Pollard TD. 1992. Phosphorylation of myosin II regulatory light chain by cyclin-p34cdc2. A mechanism for the timing of cytokinesis. J Cell Biol 118:595–605. Savage R, Danilchik M. 1993. Dynamics of germ plasm localization and its inhibition by ultraviolet irradiation in early cleavage Xenopus embryos. Dev Biol 157:371–82. Sawai T. 1982. Wavelike propagation of stretching and shrinkage in the surface of the newt’s egg before the first cleavage. J Exp Zool 222:59–68. Schliwa M, Euteneuer U, Bunlinsky JC, Izant JG. 1981. Calcium lability of cytoplasmic microtubules and its modulation by microtubule associated proteins. Proc Natl Acad Sci USA 78:1037–1041. Schroeder MM, Gard DL. 1992. Organization and regulation of cortical microtubules during the first cell cycle of Xenopus eggs. Development 114:699–709. Stennard F, Carnac G, Gurdon JB. 1996. A Xenopus T-box gene, antipodean, encodes a vegetally localised maternal mRNA that can trigger mesoderm formation. Development 122:2359–2366. Stewart-Savage J, Grey RD, Elinson RP. 1991. Polarity of the surface and cortex of the amphibian egg from fertilization to first cleavage. J Electron Microsc Tech 17:369–383. 429 Thomsen GH, Melton DA. 1993. Processed Vg1 protein is an axial mesoderm inducer in Xenopus. Cell 74:433–441. Wakahara M. 1991. Cytoplasmic localization and organization of germ-cell determinants. In: Malacinski G, ed. Cytoplasmic organization systems. New York: McGraw Hill, p 219–242. Weeks DL, Melton DA. 1987. A maternal mRNA localised to the vegetal hemisphere in Xenopus eggs codes for a growth factor related to TGF-. Cell 51:861–867. Wilsch-Brauninger H, Schwarz H, Nusslein-Volhard C. 1997. A spongelike structure involved in the association and transport of maternal products during Drosophila oogenesis. J Cell Biol 139:817–829. Yisraeli JK, Sokol S, Melton DA. 1989. The process of localizing a maternal messenger RNA in Xenopus oocytes. Development 107: 31–36. Yisraeli JK, Sokol S, Melton DA. 1990. A two-step model for the localization of maternal mRNA in Xenopus oocytes: involvement of microtubules and microfilaments in the translocation and anchoring of Vg1 mRNA. Development 108:289–298. Yoneda M, Kobayakawa Y, Kubota H, Sakai M. 1982. Surface contraction waves in amphibian eggs. J Cell Sci 54:35–46. Zhang J, King ML. 1996. Xenopus VegT RNA is localised to the vegetal cortex during oogenesis and encodes a novel T-box transcription factor involved in mesodermal patterning. Development 122:4119– 4129. Zhang J, Houston DW, King ML, Payne C, Wylie C, Heasman J. 1998. The role of maternal VegT in establishing the primary germ layers in Xenopus embryos. Cell 94:515–524. Zhou Y, King ML. 1996. RNA transport to the vegetal cortex of Xenopus oocytes. Dev Biol 179:173–183.