Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
12-Hydroxyeicosatetraenoic acid wikipedia , lookup
Monoclonal antibody wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Adaptive immune system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Innate immune system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
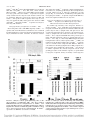
0041-1337/04/7712-1809/0 TRANSPLANTATION Copyright © 2004 by Lippincott Williams & Wilkins, Inc. Vol. 77, 1809–1814, No. 12, June 27, 2004 Printed in U.S.A. PROLONGATION OF ALLOGRAFT SURVIVAL IN CCR7-DEFICIENT MICE JAN H. BECKMANN,1 SHENG YAN,2 HEIKE LÜHRS,1 BETTINA HEID,1 SUSANNE SKUBICH,1 REINHOLD FÖRSTER,3 AND MATTHIAS W. HOFFMANN1,4 Background. Lymphocyte homing to secondary lymphoid organs is thought to be required for initiation of the alloreactive immune response. Because CCR7 is the essential chemokine receptor responsible for lymphocyte and dendritic cell homing to secondary lymphoid organs, allograft survival was analyzed in CCR7-deficient (CCR7ⴚ/ⴚ) mice. Methods. Heterotopic heart and skin allotransplantation was performed in CCR7ⴚ/ⴚ and wild-type (WT) recipients. Graft survival was monitored daily. Grafts and draining lymph nodes were analyzed by immunohistology and flow cytometry at different time points. Groups of mice were splenectomized at the day of allotransplantation. Results. A significant though modest prolongation of allograft survival in CCR7ⴚ/ⴚ recipients was observed for heart grafts (WT, 7.3ⴞ0.5 days; CCR7ⴚ/ⴚ, 10.7ⴞ2.8 days) and skin grafts (WT, 8.9ⴞ0.9 days; CCR7ⴚ/ⴚ, 12.3ⴞ0.9 days). This was accompanied by a delay in the cellular infiltration of allografts. T-cell accumulation and expansion in the draining lymph nodes in CCR7ⴚ/ⴚ recipients was severely impaired. Splenectomy had only a moderate prolongation effect on allograft survival in CCR7ⴚ/ⴚ mice. Conclusions. These results suggest that CCR7-dependent processes support allograft rejection yet are dispensable for the rejection response. Initiation of organ allograft rejection is thought to occur in secondary lymphoid organs (lymph nodes, spleen), where alloreactive T lymphocytes and antigen-presenting cells convene (1). Migration of T cells and antigen-presenting cells (APC) to secondary lymphoid organs is controlled by the homeostatic chemokine receptor CCR7 and its ligands, CCL19 (ELC) and CCL21 (SLC) (2). To prevent lymphocyte homing and activation during allograft rejection, CCR7 thus represents an ideal target. CCR7-deficient mice have been generated by one of us, and show a severe defect in T-cell and dendritic cell migration to secondary lymphoid organs, as well as diminished delayed-type hypersensitivity responses in vivo (3). They differ from mouse models, in which a different family of chemokine receptors, the inflammatory chemokine receptors, has been inactivated. Inflammatory chemokine receptors are involved in the effector phase of the immune response. CXCR3-, CCR1-, CCR5-, and CXC3CR1deficient mice show a limited prolongation of allograft survival that is significantly augmented by the addition of the immunosuppressive drug cyclosporine A (CsA) (4 –7). The present study was performed to investigate the role of T-cell and APC migration to secondary lymphoid organs during the initiation of the immune response. We demonstrate that CCR7 deficiency extends skin and heart allograft survival but cannot ultimately prevent allograft rejection. Supporting the proposed mechanism of a defect in T-cell and APC recirculation, activation of T cells in graft draining lymph nodes was severely impaired in CCR7-deficient recipients. In contrast to the effects observed in inflammatory chemokine receptor-deficient mice, the immunosuppressive drug CsA did not augment but rather abrogated the prolongation effect of CCR7 deficiency on allograft survival. MATERIALS AND METHODS Mice CCR7-deficient (CCR7⫺/⫺) and littermate control mice were originally on a mixed 129Sv(H-2b)/BALB/c (H-2d) background (3). 129Sv/ BALB/c mice were intercrossed, and H-2b homozygous offspring were selected by flow cytometry. H-2b homozygous mice were used for most experiments and are termed CCR7⫺/⫺ or littermate mice, if not stated otherwise. For selected experiments, CCR7⫺/⫺ and littermate control mice were backcrossed to C57/Bl6 (B6) for three generations and are termed CCR7⫺/⫺ (B6) in the text. The latter mice had the same H-2 (H-2b) as the original CCR7⫺/⫺ mice, but differed at minor histocompatibility loci from the 129Sv/BALB/c mice. Therefore, these mice were used to study the role of minor histocompatibility differences in heart- and skin-grafted mice. C3H/HeN (H-2k) mice served as third-party controls. All mice were bred and maintained at the Central Animal Facility, Hannover Medical School, under specificpathogen–free conditions. All experiments were approved by the animal protection committee of the local authorities. This work was supported by a grant from the Roche Organ Transplantation Research Foundation to M.W.H. and R.F., project grant B10 of the SFB265 to M.W.H., and grant DFG Fo334/1-1 to R.F. Operative Procedures 1 Department of Visceral and Transplantation Surgery, Hannover Medical School, Hannover, Germany. Heterotopic heart grafting was performed according to Corry et al. 2 Department of Visceral and Transplantation Surgery, Hannover (8). Pulmonary artery and aorta were anastomosed to the recipient’s Medical School, Hannover, Germany. Currently, 1st Affiliated Hos- inferior vena cava and abdominal aorta, respectively. Graft function pital, Zhejiang University, China. was checked daily by palpation and was scored from 0 (no palpable 3 Institute of Immunology, Hannover Medical School, Hannover, heart beat) to 4 (strong, fast, rhythmic) according to Corry et al. (8). Germany. Cessation of heart function was confirmed by laparotomy. 4 Skin grafting was performed as described by Billingham and MeAddress correspondence to: Matthias W. Hoffmann, M.D., Ph.D., Visceral and Transplantation Surgery, Hannover Medical dawar (9). Recipients were anesthetized, shaved, and grafted with School, 30625 Hannover, Germany. Email: hoffmann.matthias@ tail skin on the back or flank. Bandages were removed after 8 days, mh-hannover.de. and grafts were checked daily for signs of rejection. Grafts were Received 21 December 2003. scored as rejected when they had shrunk to less than a third of their Revision requested 14 January 2004. Accepted 5 February 2004. original size. DOI: 10.1097/01.TP.0000131159.25845.EB 1809 1810 TRANSPLANTATION Vol. 77, No. 12 Splenectomy was performed transabdominally during the heart grafting procedure or through a left subcostal incision before skin grafting. The splenic vessels were ligated with 8-0 silk, and the abdominal wall and skin were closed in layers. contralateral axillary and brachial lymph nodes. Draining lymph nodes after syngeneic skin transplantation served as further controls. Immunosuppression Mean survival time and standard deviation were calculated for each group. Survival time between the two groups was compared by the independent-samples t test (SPSS 9.0; SPSS, Inc., Chicago, IL). P⬍0.05 was considered significant. Statistical Analysis Recipients were treated with subtherapeutic doses of cyclosporine A (Sandimmune; Novartis Pharma, Basel, Switzerland). Four hours after heart transplantation, mice were injected intraperitoneally with 10 mg/kg of CsA in normal saline daily according to published protocols (4, 5). Skin-grafted mice received daily doses of 20 mg/kg of CsA administered intraperitoneally according to Larsen et al. (10). RESULTS Prolonged Survival of CCR7⫺/⫺ Skin and Cardiac Allografts Immunopathology Skin and heart grafts were harvested at different time points, embedded in Tissue Tek freezing medium (Reichert-Jung, Nuûloch, Germany), snap-frozen, and stored in liquid nitrogen. Then, 5-m cryostat sections (Frigocut 2800 E; Reichert-Jung) were air-dried overnight and fixed in acetone at room temperature before hematoxylin-eosin staining (Merck, Germany; Sigma Chemical Co., St. Louis, MO) or incubation with monoclonal antibodies. Primary and secondary antibodies were RM4-5/L3T4 (anti-CD4; PharMingen, San Jose, CA), 53-6.7 (anti-CD8), RB6.8C5 (Gr-1; anti-Ly-6G, neutrophils), and M1/70.15 (Mac1; anti-CD11b, macrophages). Antibodies were detected with horseradish peroxidase-conjugated goat anti-rat antibodies (Dianova, Hamburg, Germany), and were visualized with aminoethylcarbazole (Sigma, Deisenhofen, Germany). High-power fields of 200⫻ magnification were digitally scanned for counting immunostained cells of 20 fields (Olympus B202 microscope; Olympus Optical Co., Tokyo, Japan; AxioCam MRC; Carl Zeiss). Results are expressed as immunostained cells per field. To determine the role of CCR7 in the rejection of primarily vascularized heart and nonvascularized skin allografts, fully major histocompatibility complex (MHC)-mismatched C3H (H-2k) hearts and skin were grafted to CCR7⫺/⫺ mice and to wild-type (WT) littermates (H-2b). Whereas WT mice rejected C3H hearts within 7.3 days, allograft survival in CCR7⫺/⫺ mice was prolonged to 10.7 days (P⬍0.01) (Table 1). Similarly, rejection of C3H skin grafts was significantly faster in WT (8.9 days) compared with CCR7⫺/⫺ recipients (12.3 days, P⬍0.01) (Table 2). Similar results were observed in mice that were matched at the MHC but mismatched at non-MHC loci. To this end, CCR7⫺/⫺ and WT skin (H-2b, minor histocompatibility antigens derived from 129Sv and BALB/c) was grafted to CCR7⫺/⫺ (B6) and WT (B6) mice (H-2b, minor histocompatibility antigens derived from C57Bl/6). Whereas WT recipients rejected minor histocompatibility disparate skin grafts within 11.3 days, graft survival in CCR7⫺/⫺ mice was significantly prolonged to 17.5 days (P⬍0.01) (Table 2). To examine the role of graft-derived cells in allograft rejection (e.g., dendritic cells), CCR7⫺/⫺ hearts and skin (H-2b) were grafted to MHC and minor disparate C3H recipients. Neither heart-grafted (7.6 days in WT vs. 8.2 days in CCR7⫺/⫺) nor skin-grafted mice (12.0 days in WT vs. 11.0 days in CCR7⫺/⫺) showed any signs of prolonged graft survival (Tables 1 and 2). Finally, deficiency of CCR7 in both donor and recipient was analyzed in CCR7⫺/⫺ recipients by transplanting allografts from CCR7⫺/⫺ mice. To this end Flow Cytometry At different time points after skin transplantation to the left flank, recipients were killed. Left axillary and brachial (draining) lymph nodes, right axillary and brachial (nondraining) lymph nodes, inguinal lymph nodes, and spleen were harvested, and isolated cells were stained with the antibodies 53-6.7 (CD8FITC), GK1.5/L3T4 (CD4PE) (Becton Dickinson, Franklin Lakes, NJ), KT3 (CD3FITC), and 1D3 (CD19bio). Streptavidin APC (Caltag Laboratories, Burlingame, CA) was used as the secondary antibody to detect biotinylated primary antibodies. Multiple color analysis was performed on a FACSCalibur (Becton Dickinson). Absolute and relative numbers of immunostained cells per lymph node were compared between ipsilateral and TABLE 1. CCR7–/– mice reject MHC-mismatched heart allografts in a prolonged fashiona Donor C3H (H-2k) C3H (H-2k) WT (H-2b) CCR7–/– (H-2b) Recipient WT (H-2b) CCR7–/– (H-2b) C3H (H-2k) C3H (H-2k) Survival (days) 7, 7, 7, 5, 7, 8, 7, 7, 7, 9, 8, 8, 7, 9, 8, 8, 8, 8 10, 10, 12, 12, 14, 16 8 13 Mean⫾SD p value 7.3⫾0.5 10.7⫾2.8 7.6⫾0.5 8.2⫾2.9 ⬍0.01 NS a WT and CCR7–/– mice received C3H vascularized heart allografts. NS, Not significant. TABLE 2. Rejection of skin allografts in fully mismatched (donor, H-2k; recipient, H-2b) combinations and in non–MHCdisparate but MHC-matched combinations (donor, H-2b; minor H, 129Sv/BALB/c; recipient, H-2b; minor H, C57Bl/6) Donor C3H (H-2k) C3H (H-2k) WT (H-2b) CCR7–/– (H-2b) WT (H-2b) WT (H-2b) CCR7–/– (H-2b) CCR7–/– (H-2b) Recipient WT (H-2b) CCR7–/– (H-2b) C3H (H-2k) C3H (H-2k) WT B6 (H-2b) CCR7–/– B6 (H-2b) WT B6 (H-2b) CCR7–/– B6 (H-2b) Survival (days) 8, 8, 8, 8, 8, 9, 9, 9, 10, 10, 10, 10 11, 12, 12, 12, 12, 12, 13, 13, 14 9, 11, 11, 13, 14, 14 10, 10, 10, 11, 12, 13 10, 10, 11, 11, 13, 13 14, 14, 14, 19, 21, 23 11, 12, 12, 12, 17, 17 19, 19, 21, 22, 23, 23 Mean⫾SD p value 8.9⫾0.9 12.3⫾0.9 12.0⫾2.0 11.0⫾1.3 11.3⫾1.4 17.5⫾4.0 13.5⫾2.7 21.2⫾1.8 ⬍0.01 NS ⬍0.01 ⬍0.01 June 27, 2004 ⫺/⫺ 1811 BECKMANN ET AL. b CCR7 skin (H-2 , minor H: 129Sv/BALB/c) was grafted to WT (B6) and CCR7⫺/⫺ mice (B6; H-2b, minor H: B6). Whereas CCR7⫺/⫺ skin was rejected by WT recipients within 13.5 days, graft survival in CCR7⫺/⫺ recipients was significantly extended to 21.2 days (P⬍0.01) (Table 2). There was no significant difference between the survival of WT and CCR7⫺/⫺ skin grafts in WT recipients (11.3 days vs. 13.5 days, not significant [NS]) and between the survival of WT and CCR7⫺/⫺ allografts in CCR7⫺/⫺ recipients (17.5 days vs. 21.2 days, NS) (Table 2). Intragraft Leukocyte Infiltration in CCR7⫺/⫺ Mice Heart allografts with a full MHC disparity (CCR7⫺/⫺ recipients and control littermates, C3H donors) were examined by immunohistology at days 4 and 6 after transplantation. At day 4, infiltration of CD4⫹ and CD8⫹ T cells was consider- FIGURE 1. Analysis of heart graft infiltrating cells. (a) Reduced numbers of infiltrating CD8ⴙ cells after complete MHC-mismatched heart transplantation to CCR7ⴚ/ⴚ compared with WT recipients (anti-CD8; magnification ⴛ200) (b) Significant reduction of infiltrating CD4ⴙ and CD8ⴙ T cells at day 4 after heterotopic heart transplantation in CCR7ⴚ/ⴚ (open bars) vs. WT mice (closed bars). *P<0.05. ⫺/⫺ ably reduced in CCR7 recipients compared with WT controls. However, analyzing grafts 6 days after transplantation revealed similar levels of leukocyte infiltration in both groups (Fig. 1). Cell numbers of infiltrating neutrophils and macrophages in CCR7⫺/⫺ recipients compared with WT mice showed no significant differences at any time. Lack of T-Cell Expansion in Draining Lymph Nodes of CCR7⫺/⫺ Compared with WT Recipients after Fully Mismatched Unilateral Skin Transplantation To examine the intensity of the allogeneic immune response in CCR7⫺/⫺ mice, C3H skin was transplanted to the recipient’s left flank. Recipients were killed at different time points (days 4 and 7). Lymph nodes and spleen were harvested and analyzed by flow cytometry. As described previously (3), lymph node cell counts were reduced to a tenth in CCR7⫺/⫺ mice compared with WT mice. The percentages of CD4- and CD8-expressing cells were significantly lower in CCR7⫺/⫺ mice (Fig. 2). After fully MHC-mismatched skin transplantation (donor, C3H; recipient, CCR7⫺/⫺ or WT), cell numbers in the draining lymph nodes of CCR7⫺/⫺ recipients were 5 to 10 times lower compared with WT mice. Whereas cell numbers in draining lymph nodes in WT recipient mice increased at day 7, lymph node cells in CCR7⫺/⫺ mice showed no further expansion. To exclude nonspecific effects of the grafting procedure on the immune response, we grafted skin to syngeneic recipients. On day 4 and day 7 after syngeneic FIGURE 2. Leukocyte subsets in draining and nondraining lymph nodes at different time points after unilateral MHCmismatched skin transplantation. Nondraining and draining lymph nodes were analyzed 4 and 7 days after transplantation. Draining lymph nodes in CCR7ⴚ/ⴚ mice failed to expand (right) compared with syngeneic (left) and allogeneic (center) transplanted WT recipients. 1812 TRANSPLANTATION Vol. 77, No. 12 grafting, increased cell numbers were observed in the draining lymph nodes. These were considerably lower than in allografted mice but significantly higher compared with CCR7⫺/⫺ mice (Fig. 2). In contrast, the number of lymphocytes in the spleen of recipients of syngeneic grafts showed no significant differences in CCR7⫺/⫺ and WT 129Sv mice. As shown previously (3), the number of CD4⫹ and CD8⫹ cells in the spleen of CCR7⫺/⫺ mice was increased two- to threefold: naive CCR7⫺/⫺ mice contained 23.2⫾11.3⫻106 CD4 T cells, whereas syngeneically grafted mice contained 20.7⫾3.8⫻106 and allogeneically grafted mice 24.1⫾7.9⫻106 CD4 T cells. In contrast, WT mice contained consistently fewer CD4 T cells (naive, 10.0⫾2.5⫻106; syngeneic graft, 9.6⫾1.5⫻106; allogeneic graft, 6.8⫾1.8⫻106 CD4 T cells). A similar result was observed for CD8⫹ cells in the spleen of CCR7⫺/⫺ recipients (naive, 9.1⫾3.4⫻106; syngeneic graft, 8.9⫾0.1⫻106; allogeneic graft, 9.4⫾1.6⫻106 CD4 T cells) and WT recipients (naive, 5.3⫾0.3⫻106; syngeneic graft, 4.7⫾0.7⫻106; allogeneic graft, 3.1⫾0.9⫻106 CD4 T cells). Additional Prolongation of Allograft Survival in Splenectomized CCR7⫺/⫺ Mice The above results demonstrate that CD4 and CD8 T-cell numbers are increased in the spleens of CCR7⫺/⫺ mice. In accordance with previously reported results in alymphoid aly/aly mice (11), which after splenectomy were unable to reject allogeneic heart grafts, we attempted to extend allograft survival in CCR7⫺/⫺ mice by splenectomy. To this end, WT and CCR7⫺/⫺ mice were splenectomized and grafted with C3H skin. Compared with nonsplenectomized recipients, splenectomy significantly prolonged the survival of skin grafts in WT recipients by more than 4 days (8.9⫾0.9 days vs. 13.6⫾1.3 days, P⬍0.01) (Fig. 3). In CCR7⫺/⫺ mice, splenectomy extended skin graft survival by 5 days (12.3⫾0.9 days vs. 17.3⫾2.2 days, P⬍0.01) (Fig. 3). Heart graft survival could also slightly be enhanced by splenectomy in CCR7⫺/⫺ recipients (10.7⫾2.8 days vs. 13.3⫾1.0 days, P⬍0.05), whereas WT recipients showed no significant increase in graft survival (7.3⫾0.5 days vs. 8.3⫾1.2 days, NS) (Fig. 3). Subtherapeutic Doses of CsA Abrogated the CCR7⫺/⫺ Effect Synergistic effects between cyclosporine and the deficiency of different chemokine receptors (e.g., CXCR3, CCR5, CX3CR1, and CCR1) have previously been reported (4 –7). To elucidate whether CsA might augment allograft survival in CCR7⫺/⫺ mice, WT and CCR7⫺/⫺ mice, grafted with C3H (H-2k) heart or skin, were treated with subtherapeutic daily doses of CsA. In WT mice, CsA did not significantly prolong heart allograft survival (7.8⫾1.8 days vs. 7.3⫾0.5 days without CsA, NS). Interestingly, CsA abrogated the previously observed prolongation of heart graft survival in CCR7⫺/⫺ mice (8.0⫾2.0 days with CsA vs. 10.7⫾2.8 days without CsA, NS). In contrast, in the presence of CsA, skin graft survival was extended in both WT (15.5⫾2.4 days vs. 8.9⫾0.9 days without CsA, P⬍0.01) and CCR7⫺/⫺ mice (15.7⫾1.5 days vs. 12.3⫾0.9 days without CsA, P⬍0.05). Similar to the heartgrafted mice, there was no significant difference in allograft survival between CsA-treated WT and CCR7⫺/⫺ mice (15.5⫾2.4 days in WT vs. 15.7⫾1.5 days in CCR7⫺/⫺ mice, NS) (Fig. 4). FIGURE 3. MHC-mismatched (H-2k) allograft survival in WT and CCR7ⴚ/ⴚ recipients (H-2b) with or without splenectomy. (open diamonds) WT recipients; (filled diamonds) WT splenectomized recipients; (open squares) CCR7ⴚ/ⴚ recipients; (filled squares) CCR7ⴚ/ⴚ splenectomized recipients. (a) Heart graft survival time. (b) Skin graft survival time. DISCUSSION The above results show a significant though modest prolongation of allograft survival in mice lacking the chemokine receptor CCR7. Because CCR7 is essential for T-lymphocyte and APC migration to secondary lymphoid organs (3), this limited effect indicates that homing to secondary lymphoid organs accomplishes a nonessential role in allograft rejection. In this respect, the results resemble those obtained in lymphotoxin-␣– deficient and lymphotoxin--receptor– deficient mice that are devoid of lymph nodes and Peyer’s patches. Splenectomized lymphotoxin-␣– deficient and lymphotoxin--receptor– deficient mice reject fully allogeneic heart allografts in a delayed fashion, but are not tolerant (12, 13). These results contrast to experiments by Lakkis et al. (11), who used the alymphoid aly/aly mouse strain for similar experiments. Splenectomized aly/aly mice were proposed to June 27, 2004 BECKMANN ET AL. FIGURE 4. MHC-mismatched (H-2k) allograft survival in WT and CCR7ⴚ/ⴚ recipients (H-2b) with or without CsA treatment. (open diamonds) WT recipients; (filled diamonds) WT recipients treated with CsA; (open squares) CCR7ⴚ/ⴚ recipients; (filled squares) CCR7ⴚ/ⴚ recipients treated with CsA. (a) Mice that underwent heart transplantation received 10 mg/kg of CsA administered intraperitoneally per day. (b) Skin-grafted recipients received 20 mg/kg of CsA per day. represent a mouse model lacking secondary lymphoid organs, and were tolerant to allogeneic skin and heart allografts. Because both of these models, however, have severe defects in other lymphoid compartments, it remained uncertain whether the observed effects were mediated solely by the absence of secondary lymphoid organs or might result from support of hidden factors. The presented results obtained in CCR7⫺/⫺ mice address the question of lymphocyte homing in a model where lymph nodes and spleen are morphologically intact but where deficiency of a single receptor, CCR7, impairs T-lymphocyte and APC migration into the secondary lymphoid organs. Our results suggest that homing of these cells by means of CCR7 is not an absolute requirement for allograft rejection. Alternatively, it might be argued that CCR7⫺/⫺ mice have developed CCR7-independent pathways 1813 of T-cell activation to fight environmental pathogens. This view is supported by the finding that in lymph nodes and spleen of CCR7⫺/⫺ mice a large proportion of T cells express the activation and memory marker CD44 at high density (3). Therefore, it appears possible that cross-reactive memory T cells might be involved in the rejection of allografts in CCR7⫺/⫺ mice. It has been debated whether priming of alloreactive immune responses occurs in secondary lymphoid organs or in the allograft itself. Results from this study document a limited effect of CCR7 deficiency on allograft survival, indicating that homing to secondary lymphoid organs is not absolutely required for rejection. However, it cannot be formally ruled out that additional chemotactic factors or adhesion molecules might compensate for the deficiency of CCR7. For instance, L-selectin has been shown to be important for the adhesion of leukocytes to high endothelial venules in secondary lymphoid organs. Similar to our results in CCR7⫺/⫺ mice, L-selectin– deficient mice rejected allogeneic skin grafts in a delayed fashion (14). Furthermore, Tang et al. (14) reported a delay in the cellular infiltration of skin allografts in L-selectin– deficient mice. Recent experiments by Kreisel et al. (15) support the concept that nonhematopoietic cells within the allograft can directly activate CD8⫹ T cells. Further on, they showed that endothelial cells have the capacity to activate CD8⫹ T cells in a B7-dependent fashion (16). These findings suggest that nonprofessional APC, such as endothelial cells in case of vascularized heart grafts, are sufficient to initiate the alloreactive immune response, independent of secondary lymphoid organs. Dendritic cells (DC) have been shown to migrate from the allograft to secondary lymphoid organs (17). The authors speculated that DC migration from the allograft to SLO might initiate the alloimmune response. Because CCR7 is a key regulator of DC immigration into secondary lymphoid organs, one aspect of the current study was to determine whether CCR7-mediated migration of DC from the allograft to SLO might affect the rejection response. Our results show that CCR7⫺/⫺ skin and heart allografts were rejected with similar kinetics as WT allografts. This indicates that CCR7mediated lymphocyte trafficking to SLO has no superior role in allograft rejection. Further experiments aimed to determine whether CCR7 deficiency of both donor and recipient might improve allograft survival. The results demonstrate that in allografts differing in multiple minor antigens, allograft survival was not significantly prolonged in CCR7⫺/⫺ recipients of CCR7⫺/⫺ grafts compared with allograft survival of WT grafts in CCR7⫺/⫺ recipients. Therefore, CCR7mediated migration of passenger DC to secondary lymphoid organs appears not to be essential to trigger an alloresponse. Several chemokines and their receptors have been shown to influence allograft survival. Distinct from the homeostatic chemokine receptor CCR7, all of these are inflammatory chemokines. The most pronounced effect was observed when CXCR3 was inactivated in CXCR3-deficient mice or by antiCXCR3 monoclonal antibody therapy. MHC-disparate heart allograft survival was prolonged to 58 days in CXCR3-deficient mice compared with 9 days in WT controls (4). Interestingly, permanent graft survival was observed when subtherapeutic doses of CsA were administered during the first 14 days after grafting. These results contrast with the findings in the present study. In CCR7⫺/⫺ recipients, the prolon- 1814 TRANSPLANTATION gative effect of CCR7 deficiency in heart-grafted mice was completely abrogated by the application of CsA. Similarly, in skin-grafted mice, subtherapeutic CsA caused a significant prolongation of skin graft survival in WT mice. In contrast, addition of CsA in CCR7⫺/⫺ mice caused no prolongation compared to untreated mice. These results suggest that CsA had a negative rather than positive effect on allograft survival in the absence of CCR7. The reasons for this effect are currently not clear. It may be speculated that CCR7-mediated allograft prolongation might involve an activity of responsive T cells, possibly with protective capacities, that might be inhibited in the presence of CsA. CONCLUSION We have shown that CCR7 has a significant but overall limited role in the prolongation of heart and skin allograft survival in the mouse. This effect could be further enhanced in splenectomized mice. Conventional immunosuppression prevented rather than augmented the prolongation effect in CCR7⫺/⫺ mice. The results suggest that recirculation of leukocytes to secondary lymphoid organs has a significant but nonessential effect on the alloreactive immune response. Acknowledgments. The authors thank M. Lipp for providing CCR7⫺/⫺ mice, G. Bernhardt for critically reading the article, and C. Ziegowski for excellent animal husbandry. REFERENCES 1. Ascher NL, Hoffman RA, Hanto DW, et al. Cellular basis of allograft rejection. Immunol Rev 1984; 77: 217–232. 2. Cyster JG. Chemokines and cell migration in secondary lymphoid organs. Science 1999; 286: 2098 –2102. 3. Forster R, Schubel A, Breitfeld D, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in sec- Vol. 77, No. 12 ondary lymphoid organs. Cell 1999; 99: 23–33. 4. Hancock WW, Lu B, Gao W, et al. Requirement of the chemokine receptor CXCR3 for acute allograft rejection. J Exp Med 2000; 192: 1515–1520. 5. Gao W, Topham PS, King JA, et al. Targeting of the chemokine receptor CCR1 suppresses development of acute and chronic cardiac allograft rejection. J Clin Invest 2000; 105: 35– 44. 6. Gao W, Faia KL, Csizmadia V, et al. Beneficial effects of targeting CCR5 in allograft recipients. Transplantation 2001; 72: 1199 –1205. 7. Robinson LA, Nataraj C, Thomas DW, et al. A role for fractalkine and its receptor (CX3CR1) in cardiac allograft rejection. J Immunol 2000; 165: 6067– 6072. 8. Corry RJ, Winn HJ, Russell PS. Heart transplantation in congenic strains of mice. Transplant Proc 1973; 5: 733–735. 9. Billingham RE, Medawar PB. The technique of free skin grafting in mammals. J Exp Biol 1951; 28: 385– 401. 10. Larsen CP, Elwood ET, Alexander DZ, et al. Long-term acceptance of skin and cardiac allografts after blocking CD40 and CD28 pathways. Nature 1996; 381(6581): 434 – 438. 11. Lakkis FG, Arakelov A, Konieczny BT, et al. Immunologic ‘ignorance’ of vascularized organ transplants in the absence of secondary lymphoid tissue. Nat Med 2000; 6(6): 686 – 688. 12. Chin R, Zhou P, Alegre ML, et al. Confounding factors complicate conclusions in aly model. Nat Med 2001; 7(11): 1165–1166. 13. Zhou P, Hwang KW, Palucki D, et al. Secondary lymphoid organs are important but not absolutely required for allograft responses. Am J Transplant 2003; 3: 259 –266. 14. Tang ML, Hale LP, Steeber DA, et al. L-selectin is involved in lymphocyte migration to sites of inflammation in the skin: Delayed rejection of allografts in L-selectin-deficient mice. J Immunol 1997; 158: 5191– 5199. 15. Kreisel D, Krupnick AS, Gelman AE, et al. Non-hematopoietic allograft cells directly activate CD8⫹ T cells and trigger acute rejection: An alternative mechanism of allorecognition. Nat Med 2002; 8: 233–239. 16. Kreisel D, Krupnick AS, Balsara KR, et al. Mouse vascular endothelium activates CD8⫹ T lymphocytes in a B7-dependent fashion. J Immunol 2002; 169: 6154 – 6161. 17. Larsen CP, Steinman RM, Witmer-Pack M, et al. Migration and maturation of Langerhans cells in skin transplants and explants. J Exp Med 1990; 172: 1483–1493.