Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Adaptive immune system wikipedia , lookup
Molecular mimicry wikipedia , lookup
Sjögren syndrome wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
NK Cells in Central Nervous System
Disorders
This information is current as
of May 7, 2017.
Aurélie Poli, Justyna Kmiecik, Olivia Domingues, François
Hentges, Mathieu Bléry, Martha Chekenya, José Boucraut
and Jacques Zimmer
J Immunol 2013; 190:5355-5362; ;
doi: 10.4049/jimmunol.1203401
http://www.jimmunol.org/content/190/11/5355
Subscription
Permissions
Email Alerts
This article cites 96 articles, 28 of which you can access for free at:
http://www.jimmunol.org/content/190/11/5355.full#ref-list-1
Information about subscribing to The Journal of Immunology is online at:
http://jimmunol.org/subscription
Submit copyright permission requests at:
http://www.aai.org/About/Publications/JI/copyright.html
Receive free email-alerts when new articles cite this article. Sign up at:
http://jimmunol.org/alerts
The Journal of Immunology is published twice each month by
The American Association of Immunologists, Inc.,
1451 Rockville Pike, Suite 650, Rockville, MD 20852
Copyright © 2013 by The American Association of
Immunologists, Inc. All rights reserved.
Print ISSN: 0022-1767 Online ISSN: 1550-6606.
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
References
NK Cells in Central Nervous System Disorders
Aurélie Poli,*,† Justyna Kmiecik,† Olivia Domingues,* François Hentges,*
Mathieu Bléry,‡ Martha Chekenya,† José Boucraut,x,{ and Jacques Zimmer*
T
he CNS has long been considered an immune-privileged site because the intracerebral injection of an
Ag failed to generate a classical immune response.
The brain parenchyma, the interstitial fluid, and the cerebrospinal fluid (CSF) are isolated from the blood by two barriers:
the blood–brain barrier (BBB), which is localized at the level
of the endothelial cells, and the blood–CSF barrier (BCSFB) at
the epithelial layer of the choroid plexus. Both form complex
morphological features, such as tight junctions between endothelial cells to restrict the diffusion of soluble molecules,
pathogens, and immune cells. Despite these barriers, immune
cells are detected in the brain parenchyma and in the CSF,
where memory CD4+ T cells are predominant, supporting the
hypothesis of immune surveillance of the CNS (1). In pathological conditions, the integrity of the BBB and the BCSFB can
be disturbed, becoming permissive for the entry of inflammatory cells. Among them, NK cells were shown to be recruited to
the CNS following several pathological conditions (2–6).
*Laboratory of Immunogenetics and Allergology, Public Research Center for Health,
L-1526 Luxembourg, Luxembourg; †Translational Cancer Research Group, Department
of Biomedicine, University of Bergen, N-5009 Bergen, Norway; ‡Innate Pharma, 13276
Marseille, France; xAix Marseille Université, Centre de Recherche en NeurobiologieNeurophysiologie de Marseille, Centre National de la Recherche Scientifique Unité
Mixte de Recherche 7286, 13344 Marseille Cedex 15, France; and {Laboratoire
d’Immunologie, Assistance Publique–Hôpitaux de Marseille, 13009 Marseille, France
Received for publication December 11, 2012. Accepted for publication March 10, 2013.
This work was supported by the Ministry of Higher Education and Research of
Luxembourg, the Centre National de la Recherche Scientifique, the Bergen Medical
Research Foundation, the Meltzer Fund, and the Norwegian Cancer Society.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1203401
NK cells are the ‘‘founding’’ members of the innate lymphoid cell family. NK cells have been initially merely considered as nonspecific killers, whereas ‘‘primed’’ NK cells also
abundantly produce cytokines and chemokines and likewise
modulate the adaptive immune responses (7). Although NK
cells can assist in dendritic cell (DC) maturation and T cell
polarization, they can also prevent and limit immune responses via killing of APCs and lymphoid cells (7). These
pleiotropic functions of NK cells might be related to their
heterogeneity in phenotypes and functions. In mice, NK cells
were shown to differentiate from CD11blow to CD11bhigh
and could be further distinguished by their expression of
CD27. The immature CD11blowCD27+ subset has the
highest proliferative capacity and is mainly found in bone
marrow and lymph nodes (LNs), whereas CD11bhighCD27+
and CD11bhighCD272 NK cells present lower rates of proliferation and are found in spleen and peripheral blood (8).
In humans, NK cells can be subdivided into different populations based on the relative expression of the surface markers CD16 and CD56. The mature subtype CD56dimCD16+,
found mainly in peripheral blood, has reduced proliferative
capacity and produces negligible amounts of cytokines but is
highly cytotoxic (9). In contrast, the more immature subset,
CD56brightCD162/dim, mainly present in peripheral lymphoid organs, is able to proliferate and to secrete a large range
of cytokines, though it presents minimal cytotoxic capacity
(10).
Although NK cell biology remains understudied in neurologic diseases, there are arguments to think that specific
brain factors could modulate NK cell function and vice versa.
Indeed, NK cell physiology is modulated following the circadian rhythm through the influence of neurotransmitters,
such as norepinephrine, which inhibits cytotoxic function after
engagement of b-adrenergic receptors (11). Alternatively, NK
cells could impact CNS physiology by killing glial cells (12)
or by secreting IFN-g (13). Most knowledge about NK cells
in the CNS is coming from studies of experimental autoimmune encephalomyelitis (EAE) where conflicting results are
emerging, as NK cells have been shown to be either impliAddress correspondence and reprint requests to Dr. Jacques Zimmer, Centre de Recherche Public de la Santé, 84 Val Fleuri, L-1526 Luxembourg, Luxembourg. E-mail
address: [email protected]
Abbreviations used in this article: AD, Alzheimer disease; BBB, blood–brain barrier;
BCSFB, blood–cerebrospinal fluid barrier; CSF, cerebrospinal fluid; DC, dendritic cell;
EAE, experimental autoimmune encephalomyelitis; GBM, glioblastoma multiforme;
LN, lymph node; MS, multiple sclerosis; NKCA, NK cell cytotoxic activity; NPB,
nonpathological human brain; PD, Parkinson disease; TBI, traumatic brain injury.
Copyright Ó 2013 by The American Association of Immunologists, Inc. 0022-1767/13/$16.00
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
NK cells are important players in immunity against
pathogens and neoplasms. As a component of the innate
immune system, they are one of the first effectors on
sites of inflammation. Through their cytokine production capacities, NK cells participate in the development
of a potent adaptive immune response. Furthermore,
NK cells were found to have regulatory functions to
limit and prevent autoimmunity via killing of autologous immune cells. These paradoxical functions of NK
cells are reflected in CNS disorders. In this review, we
discuss the phenotypes and functional features of peripheral and brain NK cells in brain tumors and infections, neurodegenerative diseases, acute vascular and
traumatic damage, as well as mental disorders. We also
discuss the implication of NK cells in neurotoxicity and
neuroprotection following CNS pathology, as well as
the crosstalk between NK cells and brain-resident immune cells. The Journal of Immunology, 2013, 190:
5355–5362.
5356
NK cell modifications associated with CNS disorders
There is increasing knowledge about the bidirectional interconnection between the nervous system and the immune
system (19); nonetheless, data are patchy and complex. Furthermore, it is not clear whether the modifications of the
immune system (including NK cells) that are described in
several CNS pathologies are a cause or a consequence of the
disease.
Primary brain tumors. Patients with glioma are known to
have impaired immune function, parallel to an increase of
anti-inflammatory molecules in plasma (20). Fadul et al.
(21) observed decreased absolute numbers of NK cells in
PBMCs isolated from glioblastoma multiforme (GBM)
patients who were treated with combined radiation therapy
and temozolomide. Furthermore, NK cell frequency and their
cytotoxic activity (NKCA) were shown to be affected in the
blood of brain tumor patients as well as in animal glioma
models (22, 23). Several factors could mediate this effect on
NK cell functions. The plasma levels of anti-inflammatory
molecules, such as TGF-b, were shown to be elevated in
the context of brain tumors and could inhibit NK cell functions,
as well as the level of expression of the activating receptor
NKG2D (24). Thus, decreased expression of NKG2D was
shown on NK cells that were isolated from GBM patients
(24). This downmodulation of NK cell functions in brain
tumors reflects a common mechanism of immune escape
found in other solid tumors (25).
Infections of the CNS. Viral infections. Shortly after they were
discovered, NK cells were shown to play an important role
in antiviral immune responses. The rare patients completely
devoid of NK cells were extremely susceptible to infections by
viruses of the herpes family (26, 27). Five consecutive children
with herpetic encephalitis were shown to have functional NK
cell deficiencies (27). By using a mouse model of HSV1 encephalitis, Adler et al. (28) directly demonstrated that NK
cells have an important role in the defense against this severe form of infection. Nonetheless, TAP-deficient patients
known to have hyporesponsive NK cells rarely present with
viral encephalitis (29). In contrast, TLR3–deficient patients
are susceptible to HSV encephalitis, and they present impaired NK cell IFN-g secretion following stimulation with
polyinosinic-polycytidylic acid, but normal production against
K562 (30), suggesting that TLR3 dysfunction could be at
the origin of this disease phenotype. Additionally, it has
been described that NK cells are important actors of the
antiviral immune response in mouse cerebral infections by
Theiler’s murine encephalomyelitis virus (31), in mouse hepatitis virus (32), and in Semliki Forest virus (4), with a rather
detrimental cytotoxic function in the latter case. Regarding
SIV infection in macaques (33), it has been shown that the
intensity of NK cell cytotoxicity was inversely related to the
severity of CNS lesions.
Bacterial infections of the CNS. Only a few studies addressed
the role of NK cells in bacterial infections of the CNS. Hayashi
et al. (34) developed a nonlethal mouse model of CNS listeriosis. Under these conditions, NK cells were identified as
the main source of bacterial growth-limiting IFN-g in the
early phase of infection. Mice are relatively protected from
Listeria neuroinvasion as long as they have NK cells and
can respond to IFN-g (35). In contrast, NK cell–derived
IFN-g was found to be responsible for the immune pathology
associated with Streptococcus pneumoniae meningitis, whereas
IFN-g KO mice are resistant to severe forms of this disease
(36).
Cerebral malaria. In 1986, Stach et al. (37) described that NK
cell cytotoxic function was normal in benign uncomplicated
malaria, whereas it was severely decreased in children with
cerebral malaria. In a mouse model of malaria induced by
Plasmodium berghei ANKA, it was shown that the NK cell
complex genetically influences pathogenesis, that NK cells are
necessary for both disease pathogenesis and control of parasitemia, and that NK cells infiltrate the brain of mice with
cerebral malaria (38). The same group demonstrated that a
reciprocal activation of NK cells and DCs is implicated in
the development of inflammatory pathways during malaria
(39). With IFN-g being known as a crucial factor for the
development of cerebral pathology in malaria, Villegas-Mendez
et al. (40) identified NK cells as the early source of this
cytokine, before T cells take center stage. Overall, in CNS
infections, NK cells can be beneficial or detrimental according
to the experimental model used. An emerging concept states
that neuroinflammation following infection may play an important role in the etiology of neurodegenerative diseases
(41).
Neurodegenerative diseases. Alzheimer disease and Parkinson
disease. Alzheimer (AD) and Parkinson (PD) diseases are
age-related neurodegenerative conditions characterized by
neuronal loss in hippocampus and substantia nigra, respectively. There are concepts supporting that AD and PD
pathogenesis could be originally related to systemic immune
system dysfunction (42). The frequency of NK cells seems
unmodified in the blood of AD patients (43, 44). Recently,
it was described that NK cells from these patients had increased
expression of the receptor 5-HT2C for the neurotransmitter
serotonin (45), which was shown to be implicated in either
increase or decrease of NKCA (46, 47). However, patients
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
cated in neuroprotection (14) or neurotoxicity (15). This is
highlighting the dual nature of NK cells in CNS homeostasis
as it was described for autoimmune diseases in general (16).
Thus, NK cells could either promote or limit autoimmune
disease by their intrinsic capacities to alter the issue of adaptive immune responses or preserve immune homeostasis (16).
This Jekyll and Hyde face of NK cells was observed in several
organs, including the uterus where they are essential for blood
supply to the fetus but where they could also be implicated in
pregnancy disorders (17). Although NK cell trafficking from
blood to specific organs is well characterized, the mechanisms
leading to the recruitment of NK cells to the CNS remain
unclear (18).
Monitoring NK cell subsets as surrogate markers for disease
activity as well as understanding the mechanism of recruitment and the function of brain-infiltrating NK cells are interesting goals for the development of new NK cell–based
therapies. Most studies are focusing on the modification of
systemic immunity following CNS disorders. In this review
we first discuss the impact of brain disease on circulating NK
cell physiology. We then focus on the recruitment of NK cells
within the healthy and pathological brain, which is so far
understudied.
BRIEF REVIEWS: NK CELLS IN CNS PATHOLOGY
The Journal of Immunology
with PD show an increased expression of the inhibitory receptor NKG2A (48, 49) and, depending on the study, no
modification (48) or an increased (49) expression of NKG2D
on NK cells. NKCA was shown to be either diminished (50)
or equal (51) in PD patients and controls. Furthermore, ex vivo
activation of NK cells with IL-2, IFN-a, or IFN-b (51)
resulted in increased NKCA in patients compared with control
donors. NKCA is positively correlated with PD severity (48).
Moreover, Solerte et al. (52) showed that purified NK
cells from blood of PD patients spontaneously secreted
more IFN-g and TNF-a than did healthy donors. All of
these modifications of NK cell physiology suggest that they
might play a role in the pathogenesis of PD. Nonetheless,
further investigations are required for delineating the possible
mechanisms.
Multiple sclerosis. Multiple sclerosis (MS) is the most common
tent with the reduction of their NKCA in the acute phase of
ischemic stroke. Interestingly, others reported that deficiency in
perforin expression was negatively correlated with Fas ligand
expression by NK cells in individual patients (67). The decrease
in IFN-g secretion was also observed in a mouse model of
middle cerebral artery occlusion leading to spontaneous septicemia and pneumonia (68). The adoptive transfer of NK cells
diminished bacterial load in the lung. These authors also
showed evidence that the sympathetic nervous system participates in the immunosuppression induced by middle cerebral
artery occlusion, as its blockage by a b2-adrenergic receptor
antagonist prevents the defective IFN-g response and the
occurrence of bacterial infections.
Mental disorders. A growing body of research has found that
mental disorders may influence somatic health and disease by
altering immunity. During acute stress, such as public speech,
there is a rapid mobilization of NK cells, with a CD56dim
CD16+CD62L2 phenotype, from various reservoirs into the
blood (69). Acute stress correlates with increased NKCA (70).
The NK cell redistribution was proposed to be coordinated
via secretion of catecholamines from the sympathetic nervous
system into the blood and further NK cell mobilization regulated through b2-adrenergic receptors (71). In contrast, in the
context of chronic stress or depression, NKCA is markedly
reduced compared with controls (72). Diminished NKCA was
associated with a reduction of the CD16+ NK cell subset in
blood of depressed patients (73). Additionally, several studies
have found that antidepressants that block the reuptake of
serotonin are associated with upregulation of NK cell functions in depressed individuals (74). The mechanism by which
mental dysfunction acts on NK cells is still debated. Many
hypotheses are proposed, including the deregulation of the
hypothalamic/pituitary/adrenal axis leading to elevated levels
of blood glucocorticoids, which further inhibit NKCA through
epigenetic processes (75). Several other mental disorders are
correlated with a diminution of NK cell functions, such as
obsessive-compulsive disorder (76) and autism (77). In the
latter study, the authors proposed that NK cell impairment
in early life could impede neurogenesis, resulting in subsequent
mental dysfunction, especially because neuronal stem cells were
shown to express NKG2D ligands (78).
Phenotype and physiology of brain NK cells in health and disease
Healthy brain. NK cells have been found in most organs, where
they present various phenotypes and functions related to the
organ-specific environment (18). Although they could be
recruited to the brain in response to several CNS disorders,
there are sparse data about the phenotype of NK cells in
the nascent CNS. In the C57BL/6 mouse model, NK1.1+
abTCR2 NK cells represent 9.74% of total lymphoid cells
within the brain before infection with P. berghei (38). We
confirmed the presence of NK1.1+NKp46+ NK cells in the
brain of C57BL/6 mice where they represented 11.1 6 0.4%
(n = 3) of CD45high cells (Fig. 1D). These NK cells were
in the majority immature CD11blowCD27+ and CD11bhigh
CD27+ cells and in the minority mature CD11bhighCD272
cells, representing 58.9 6 4.3, 27.0 6 3.9, and 5.7 6
1.3%, respectively (Fig. 1E). This profile corresponds to
the one in LNs (Fig. 1F), which contrasts with a higher
percentage of mature NK cells in spleen or blood (Fig. 1G,
1H).
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
disabling neurologic disease in young adults. MS is a neuroinflammatory autoimmune disease affecting the CNS, characterized by widespread inflammation with infiltration of
immune cells and demyelination. The involvement of NK
cells in the pathogenesis of MS has been shown by several
approaches: MS relapses were correlated with a deficit of NK
cells, which were proposed to modulate the autoimmune response and to participate in the repair of neurologic damage
(53); the functional activity of NK cells decreases during active phases of the disease and is restored during remission
(54); and the treatment of MS patients with IFN-b (55) or
daclizumab (56) results in decreased disease flares accompanied by a growing population of regulatory CD56bright NK
cells. Moreover, during pregnancy, MS activity is reduced
and the regulatory NK cell population is increased in blood
(57). Interestingly, a reduction of blood NK cells expressing CX3CR1 correlated with disease severity (54). Because
CD56bright NK cells were shown to be CX3CR12 in normal
donors, it would be interesting to further look at the capacity
of daclizumab to modulate CX3CR1 expression on NK cell
subtypes (58). Manipulation of NK cells in animal models
of EAE leads to contradictory results. According to some
authors, NK cells play a deleterious role (15) during myelin
oligodendrocyte glycoprotein EAE in C57BL/6 mice. On
the contrary, numerous studies attribute, using the same EAE
model, a neuroprotective role to NK cells. Depletion of
NK cells, by injecting anti-NK1.1 or anti–asialo GM1 Abs,
induced worsening of the disease, associated with increased
proliferation of CD4+ Th17 cells and high production of
cytokines (14). Overall, this suggests that NK cells are a potential target for future MS therapy.
Acute stroke and trauma. Acute CNS diseases, such as stroke and
traumatic brain injury (TBI), have been associated with increased risk of infectious complications (59). It is thought
that CNS injuries break the delicate interplay between the
immune system and CNS, resulting in secondary immunosuppression (60). Many studies have shown a reduced NKCA
of blood NK cells of patients with spinal cord injury and
TBI (61–63). Although the CD56bright population stayed
unchanged, the CD56dim subtype was decreased, and this
correlated with the occurrence of infections after TBI (64).
Alternatively, in the context of stroke, the NK cell counts
showed a nonsignificant decrease (65–67). Nonetheless, there is
a decrease of NK cells expressing IFN-g and perforin, consis-
5357
5358
We demonstrated the presence of immature NK cells in the
brain of healthy mice. We excluded a contamination by blood
NK cells, as animals were extensively perfused with PBS and
because they display different phenotypes. To our knowledge,
there is no evidence for the presence of NK cells in nonpathological human brain (NPB) parenchyma because of
obvious ethical considerations. Nonetheless, we observed rare
NK cells in the CSF of patients with either peripheral neuropathy or MS, exhibiting mainly CD56bright phenotypes
(Fig. 2A). This repartition is typical of the phenotype of NK
cells found in LNs and is different from blood NK cells
(Ref. 10 and Fig. 2B). Recently, Hamann et al. (79) observed
an overall diminished frequency of NK cells in CSF compared
with blood with a CSF/blood ratio of 1:4.
The validity of comparing between human and mouse NK
cell subsets and their maturation is still debated, despite that
CD27high mouse NK cells show some functional and phenotypic similarity to the CD56bright human subset such as the
receptor expression pattern. However, the CD27high mouse
subset is highly cytotoxic as opposed to the CD56bright human
NK cells (8). Nonetheless, it has been proposed that human
NK cell maturation could be further delineated according to
the expression of CD11b and CD27 (80), such as in the
mouse. The same team demonstrated that CD56bright NK
cells are mainly CD27+, indicating that human brain NK cells
may display an immature phenotype, although a deeper analysis
is needed to confirm this.
NK cells are known to actively screen nonlymphoid and
lymphoid organs in search of infected targets and transformed
FIGURE 2. Lymphocyte subsets in paired (A, C) CSF and (B, D) blood
samples of representative patients with NPB (A, B) and with MS (C, D). NK
cells were gated as CD45highCD32CD56dim and CD56bright. Numbers represent the percentage of cells among the CD45high population.
cells, and the brain appears as a ‘‘new’’ compartment for future
investigation of their trafficking. In steady-state conditions,
CD4+ T cells were found to patrol from the blood through
the choroid plexus into the CSF to perform CNS immunosurveillance (1). Thus it is conceivable that NK cells follow
the same route to scan the CNS milieu for pathological components either directly through their surface receptors or by
interaction with brain-resident immune cells (Fig. 3). NK cell
subsets express a different array of chemokine receptors in
humans and mice. For example, human CD56bright NK cells
were shown to express high levels of CCR7 and CXCR3, so
they can migrate in response to CCL19 and CXCL10, which
are constitutively expressed in the CNS (81). Moreover,
TGF-b is constitutively present in the CSF and was found
to modulate macrophages (82). Because TGF-b was recently
shown to negatively regulate maturation of NK cells, it might
be relevant to investigate ex vivo whether particular CSF
constituents might modify NK cell phenotypes (83). The
reason for the presence of immature NK cells in the healthy
CNS needs further experiments to delineate whether the brain
influx of NK cells is dependent on their phenotype or whether
it is the CNS milieu that further influences such influx.
CNS disorders. Although the possible immune surveillance of
the CNS by NK cells is still enigmatic, their infiltration following brain pathology is more characterized. Nonetheless,
their mode of entry into CNS and the molecules leading
to this recruitment are poorly understood. This recruitment
could be coordinated by CNS resident cells, such as microglia,
astrocytes, and neurons, that secrete chemokines involved in
NK cell migration, such as CCL2, CXCL10, and CX3CL1
(3, 18, 38).
In intracranial tumors, NK cells account for a minor part
of infiltrating leukocytes as shown in GBM, pilocystic astrocytomas, and brain metastases of carcinoma (2, 84–86).
According to our observations, NK cells comprise 2.11 6
0.54% of infiltrating immune cells in GBM (J. Kmiecik,
A. Poli, N.H.C. Brons, A. Waha, G.E. Eide, P.O. Enger,
J. Zimmer, and M. Chekenya, unpublished observations).
These tumor-infiltrating NK cells were shown to be inhibited
by glioma-derived factors but also by other local immune cells
with anti-inflammatory properties (20). Nonetheless, immu-
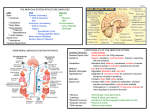
FIGURE 3. CNS NK cell physiology could be modulated by brain-resident
immune cells following CNS compartments. The passage of NK cells from
body vasculature through the CSF by the BCSFB at the choroid plexus may be
privileged at steady-states. Nonetheless, they may also cross the BBB when
circulating in brain vessels to enter the meninges, perivascular space, and brain
parenchyma.
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
FIGURE 1. Distribution of NK cell subsets in B6 mice. Tissues were disrupted using dissociation buffer containing benzonase (except whole blood);
brain cells were further enriched by Percoll density gradient. Cell suspensions
were stained for NK1.1, NKp46, CD45, CD27, CD11b, and live/dead. First,
(A) dead cells and (B) doublets were excluded, (C) immune cells (CD45+)
were gated, and then (D) NK cells were gated as NK1.1+NKp46+ events.
Representative dot plots show the profiles of CD27 and CD11b expression
of NK cells in (E) brain, (F) LNs, (G) spleen, and (H) whole blood.
BRIEF REVIEWS: NK CELLS IN CNS PATHOLOGY
The Journal of Immunology
5359
it is also dependent on interindividual genetic and environmental factors. The immune response following infection can
contribute to fatal disease, but is also necessary for recovery,
and the specific role of immune cells, including NK cells, is
not well described (89). Furthermore, many teams propose
that neurodegenerative diseases could be a consequence of
CNS infections (89). Thus, repeated infection by HSV was
proposed to be at the origin of AD and MS emergence, as
pathological features of this CNS infection share similarity
with neurodegenerative diseases, such as neuronal death (89).
Moreover, several models of CNS infection in mice were
associated with parenchymal demyelination (4). The recruitment of immune effectors, including NK cells, following
infection leads to the release of cytotoxic agents, such as
granzymes, perforin, and proinflammatory molecules, which
could generate direct or indirect cellular damages leading
to neurodegenerative foci (89) (Fig. 4). As a comparable
example, liver NK cells were implicated in the emergence
of fulminant hepatic failure following mouse hepatitis virus
infection, whereas the depletion of NK cells limited liver
damage and increased animal survival (90). Alternatively,
IFN-g was also associated with both neuronal protection and
repair by modulation of microglia properties (91), so we can
propose that NK cells secreting IFN-g following infection
may have a neuroprotective function (Fig. 4). Furthermore,
NK cells were shown to secrete IL-10 following viral infection
of the liver, and it could be interesting to investigate this in
the context of CNS infection and neuroprotection (92). All
of these facts indicate why NK cells could be implicated in
neurodegeneration and/or neuroprotection (Fig. 4).
During the last decade, the major increase in knowledge
about CNS NK cells arose from studies of MS patients and
animal models of EAE. The EAE model revealed that NK
cell homing to the CNS is essential to control neuroinflammation (Fig. 4). Indeed, recruited NK cells were shown to kill
microglia responsible for the priming of autoreactive T cells
Priming of
cytotoxic T cells
NEUROTOXICITY
IL-12
FIGURE 4. NK cells could initiate
neurotoxicity and neuroprotection following brain disorders. NK cells diminished animal survival in cerebral
malaria by activation of cytotoxic T cells
and DCs through secretion of IFN-g.
The exocytosis of cytotoxic granules
following CNS infection may also initiate inflammatory cascades leading to
brain injury. Alternatively, IFN-g could
generate neuroprotection through activation of microglia with neuroprotective
function. Finally, it was described that
in the context of EAE, NK cells reduce
neurotoxicity of autoreactive T cells by
killing proinflammatory microglia.
Perforin
Priming of
dendritic
cells
Recruitment of
cytotoxic T cells
IFN-γ
Th17 polarizing
cytokines
Perforin
Granzymes
Chemokines
Autoreactive
T cells
Killing of
proinflammatory
microglia
NK cells
GM-CSF
IFN-γ
IL-10
Priming of microglia
NEUROPROTECTION
Inhibition of
inflammatory cells
CNS recruitment of
inflammatory cells
Leukocytes
Microglia
Cytotoxic T cells
NK cells
Autoreactive T cells
Dendritic cells
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
notherapies are emerging to increase this NK cell recruitment and function. As an example, oncolytic HSV infection of GBM in mice resulted in a massive influx of NK
cells. Interestingly, although NK cells recruited to tumors in
control mice were mostly CD11bhighCD27low, oncolytic
HSV administration resulted in recruitment of CD11bhigh/low
CD27+ NK cells (87). This subpopulation was able to activate macrophages/microglia through secretion of IFN-g. In
a rat xenograph model of GBM, we also observed an increase
of proinflammatory macrophages/microglia following intracranial injection of activated NK cells leading to beneficial outcome (A. Poli, J. Wang, O. Domingues, C.B Rygh,
T. Yan, F. Thorsen, J. Planagumà, E. McCormack, F. Hentges,
P.H. Pedersen, J. Zimmer, P.O. Enger, and M. Chekenya,
submitted for publication). Thus, brain NK cells may influence the disease outcome by interaction with other CNS
immune cells.
Brain NK cells were observed in greater abundance following CNS infection (6, 38). However, NK cell recruitment
could be either beneficial or detrimental depending on the
infectious agent. Thus, although the recruitment of NK cells
was shown to be essential for controlling mouse hepatitis virus
and Semliki Forest virus replication within the CNS through
an IFN-g–dependent mechanism (3, 4), their cytolytic activity
(Fas and granule exocytosis dependent) was shown to potentiate the immunopathology, such as increased animal death
and brain demyelination (Fig. 4) (4). Alternatively, NK cells
were shown to be essential for controlling parasitemia;
however, their depletion protects C57BL/6 mice against
P. berghei–mediated cerebral malaria. It seems that NK cells
stimulated DCs to produce IL-12 required for T cell priming,
leading to subsequent cerebral disease (Fig. 4) (38, 39). In
neuroborreliosis, NK cell numbers in the CSF are significantly
higher than in tick-borne viral encephalitis, in correlation
with the level of IFN-g found in CSF (88). The outcome of
CNS infections varies according to the type of pathogens, and
5360
Conclusions
In this review we have discussed the systemic NK cell phenotypic and functional alterations involved in the most
common brain disorders. Note that the characterization of
NK cells was often not well performed, as many studies used
single staining of CD56+ or CD57+ cells (44, 45, 85, 86, 96).
Both of these molecules can also be expressed by CD3+ T cells
and should by consequence be excluded. Furthermore, there
are many unanswered questions, and the future challenge is to
delineate the precise influence of NK cells on CNS pathology,
as well as the characterization of brain-resident NK cells,
which have so far been understudied. Thus, it is not well
understood whether brain NK cells are the origin or the
consequence of CNS pathogenesis. Some evidence suggests
that NK cells can control CNS autoimmunity and inflammation by their capacity to kill autoreactive or proinflammatory microglia (5, 12, 14). Alternatively, NK cell response
in CNS following infection may be deleterious (39). Future
studies providing novel insights into the role of NK cells in
brain pathogenesis might result in the development of NK
cell–based approaches to cure patients with inflammatory and
autoimmune disorders of the CNS.
Disclosures
M.B. is an employee of Innate Pharma. The other authors have no financial
conflicts of interest.
References
1. Ransohoff, R. M., and B. Engelhardt. 2012. The anatomical and cellular basis of
immune surveillance in the central nervous system. Nat. Rev. Immunol. 12: 623–
635.
2. Domingues, P. H., C. Teodósio, J. Ortiz, P. Sousa, A. Otero, A. Maillo, P. Bárcena,
M. C. Garcı́a-Macias, M. C. Lopes, C. de Oliveira, et al. 2012. Immunophenotypic
identification and characterization of tumor cells and infiltrating cell populations in
meningiomas. Am. J. Pathol. 181: 1749–1761.
3. Trifilo, M. J., C. Montalto-Morrison, L. N. Stiles, K. R. Hurst, J. L. Hardison,
J. E. Manning, P. S. Masters, and T. E. Lane. 2004. CXC chemokine ligand 10
controls viral infection in the central nervous system: evidence for a role in innate
immune response through recruitment and activation of natural killer cells. J. Virol.
78: 585–594.
4. Alsharifi, M., M. Lobigs, M. M. Simon, A. Kersten, K. Müller, A. Koskinen, E. Lee,
and A. Müllbacher. 2006. NK cell-mediated immunopathology during an acute
viral infection of the CNS. Eur. J. Immunol. 36: 887–896.
5. Hao, J., D. Campagnolo, R. Liu, W. Piao, S. Shi, B. Hu, R. Xiang, Q. Zhou,
T. Vollmer, L. Van Kaer, et al. 2011. Interleukin-2/interleukin-2 antibody therapy
induces target organ natural killer cells that inhibit central nervous system inflammation. Ann. Neurol. 69: 721–734.
6. Cédile, O., N. Popa, F. Pollet-Villard, N. Garmy, E. C. Ibrahim, and J. Boucraut.
2010. The NKG2D ligands RAE-1d and RAE-1ε differ with respect to their receptor affinity, expression profiles and transcriptional regulation. PLoS ONE 5:
e13466.
7. Lünemann, A., J. D. Lünemann, and C. Münz. 2009. Regulatory NK-cell functions
in inflammation and autoimmunity. Mol. Med. 15: 352–358.
8. Hayakawa, Y., and M. J. Smyth. 2006. CD27 dissects mature NK cells into two
subsets with distinct responsiveness and migratory capacity. J. Immunol. 176: 1517–
1524.
9. Caligiuri, M. A. 2008. Human natural killer cells. Blood 112: 461–469.
10. Poli, A., T. Michel, M. Thérésine, E. Andrès, F. Hentges, and J. Zimmer. 2009.
CD56bright natural killer (NK) cells: an important NK cell subset. Immunology 126:
458–465.
11. Logan, R. W., A. Arjona, and D. K. Sarkar. 2011. Role of sympathetic nervous
system in the entrainment of circadian natural-killer cell function. Brain Behav.
Immun. 25: 101–109.
12. Lünemann, A., J. D. Lünemann, S. Roberts, B. Messmer, R. Barreira da Silva,
C. S. Raine, and C. Münz. 2008. Human NK cells kill resting but not activated
microglia via NKG2D- and NKp46-mediated recognition. J. Immunol. 181: 6170–
6177.
13. Walter, J., S. D. Honsek, S. Illes, J. M. Wellen, H. P. Hartung, C. R. Rose, and
M. Dihné. 2011. A new role for interferon gamma in neural stem/precursor cell
dysregulation. Mol. Neurodegener. 6: 18.
14. Hao, J., R. Liu, W. Piao, Q. Zhou, T. L. Vollmer, D. I. Campagnolo, R. Xiang,
A. La Cava, L. Van Kaer, and F. D. Shi. 2010. Central nervous system (CNS)resident natural killer cells suppress Th17 responses and CNS autoimmune pathology. J. Exp. Med. 207: 1907–1921.
15. Winkler-Pickett, R., H. A. Young, J. M. Cherry, J. Diehl, J. Wine, T. Back,
W. E. Bere, A. T. Mason, and J. R. Ortaldo. 2008. In vivo regulation of experimental autoimmune encephalomyelitis by NK cells: alteration of primary adaptive
responses. J. Immunol. 180: 4495–4506.
16. Flodström-Tullberg, M., Y. T. Bryceson, F. D. Shi, P. Höglund, and H. G. Ljunggren.
2009. Natural killer cells in human autoimmunity. Curr. Opin. Immunol. 21: 634–
640.
17. Hiby, S. E., J. J. Walker, K. M. O’shaughnessy, C. W. Redman, M. Carrington,
J. Trowsdale, and A. Moffett. 2004. Combinations of maternal KIR and fetal
HLA-C genes influence the risk of preeclampsia and reproductive success. J. Exp.
Med. 200: 957–965.
18. Shi, F. D., H. G. Ljunggren, A. La Cava, and L. Van Kaer. 2011. Organ-specific
features of natural killer cells. Nat. Rev. Immunol. 11: 658–671.
19. Wrona, D. 2006. Neural-immune interactions: an integrative view of the bidirectional relationship between the brain and immune systems. J. Neuroimmunol. 172:
38–58.
20. Parney, I. F. 2012. Basic concepts in glioma immunology. Adv. Exp. Med. Biol. 746:
42–52.
21. Fadul, C. E., J. L. Fisher, J. Gui, T. H. Hampton, A. L. Côté, and M. S. Ernstoff.
2011. Immune modulation effects of concomitant temozolomide and radiation
therapy on peripheral blood mononuclear cells in patients with glioblastoma multiforme. Neuro-oncol. 13: 393–400.
22. Servade, F., R. Parente, M. Bucci, E. Beltrandi, F. Tognetti, and G. Gaist. 1983.
Particular features of cell-mediated immunity in patients with anaplastic gliomas:
a comparison with kidney and bladder cancer patients. J. Neurooncol. 1: 327–332.
23. Alizadeh, D., L. Zhang, C. E. Brown, O. Farrukh, M. C. Jensen, and B. Badie.
2010. Induction of anti-glioma natural killer cell response following multiple lowdose intracerebral CpG therapy. Clin. Cancer Res. 16: 3399–3408.
24. Crane, C. A., S. J. Han, J. J. Barry, B. J. Ahn, L. L. Lanier, and A. T. Parsa. 2010.
TGF-b downregulates the activating receptor NKG2D on NK cells and CD8+
T cells in glioma patients. Neuro-oncol. 12: 7–13.
25. Groth, A., S. Klöss, E. P. von Strandmann, U. Koehl, and J. Koch. 2011. Mechanisms of tumor and viral immune escape from natural killer cell-mediated surveillance. J. Innate Immun. 3: 344–354.
26. Biron, C. A., K. S. Byron, and J. L. Sullivan. 1989. Severe herpesvirus infections in
an adolescent without natural killer cells. N. Engl. J. Med. 320: 1731–1735.
27. Almerigogna, F., F. Fassio, M. G. Giudizi, R. Biagiotti, C. Manuelli, E. Chiappini,
L. Galli, S. Romagnani, and M. De Martino. 2011. Natural killer cell deficiencies in
a consecutive series of children with herpetic encephalitis. Int. J. Immunopathol.
Pharmacol. 24: 231–238.
28. Adler, H., J. L. Beland, N. C. Del-Pan, L. Kobzik, R. A. Sobel, and I. J. Rimm.
1999. In the absence of T cells, natural killer cells protect from mortality due to
HSV-1 encephalitis. J. Neuroimmunol. 93: 208–213.
29. Zimmer, J., E. Andrès, L. Donato, D. Hanau, F. Hentges, and H. de la Salle. 2005.
Clinical and immunological aspects of HLA class I deficiency. QJM 98: 719–727.
30. Zhang, S. Y., E. Jouanguy, S. Ugolini, A. Smahi, G. Elain, P. Romero, D. Segal,
V. Sancho-Shimizu, L. Lorenzo, A. Puel, et al. 2007. TLR3 deficiency in patients
with herpes simplex encephalitis. Science 317: 1522–1527.
31. Paya, C. V., A. K. Patick, P. J. Leibson, and M. Rodriguez. 1989. Role of natural
killer cells as immune effectors in encephalitis and demyelination induced by
Theiler’s virus. J. Immunol. 143: 95–102.
32. Walsh, K. B., R. A. Edwards, K. M. Romero, M. V. Kotlajich, S. A. Stohlman, and
T. E. Lane. 2007. Expression of CXC chemokine ligand 10 from the mouse hep-
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
(5, 14). Furthermore, there is an exacerbation of brain lesions
and a concomitant reduction of brain NK cells during EAE
in CX3CR1-deficient mice. This emphasizes that CX3CR1+
NK cells are essential for brain homing to further modulate
EAE pathogenesis (93). In contrast, in a postmortem study of
MS patients, Durrenberger et al. (94) detected rare NKp46+
NK cells in demyelinated areas. Additionally, in the CSF,
Hamann et al. (79) described a reduction of NK cell frequency in the MS patients compared with NPB samples.
However, a preliminary investigation of four patients with
suspected MS described an increased absolute NK cell number in their CSF compared with patients with other neurologic defects (unpublished data). These NK cells display an
immature phenotype with high CD56 (Fig. 2C) and low
CX3CR1 (79) expression. Overall, the mechanism leading
to the recruitment is not clear; nonetheless, it seems that the
genetic background might influence this phenomenon. Thus,
Nimer et al. (95) found that the influx of CD32CD161bright
NK cells in experimental traumatic brain contusion in DA
rats was more marked than in PVG rats. This interindividual
genetic heterogeneity might influence the outcome and treatment response following CNS pathology and partially explain
the differences observed between studies.
BRIEF REVIEWS: NK CELLS IN CNS PATHOLOGY
The Journal of Immunology
33.
34.
35.
36.
37.
38.
39.
40.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58. Hamann, I., N. Unterwalder, A. E. Cardona, C. Meisel, F. Zipp, R. M. Ransohoff,
and C. Infante-Duarte. 2011. Analyses of phenotypic and functional characteristics
of CX3CR1-expressing natural killer cells. Immunology 133: 62–73.
59. Lee, K., and F. Rincon. 2012. Pulmonary complications in patients with severe
brain injury. Crit. Care Res. Pract. 2012: 207247.
60. Meisel, C., J. M. Schwab, K. Prass, A. Meisel, and U. Dirnagl. 2005. Central
nervous system injury-induced immune deficiency syndrome. Nat. Rev. Neurosci. 6:
775–786.
61. Quattrocchi, K. B., E. H. Frank, C. H. Miller, A. Amin, B. W. Issel, and
F. C. Wagner, Jr. 1991. Impairment of helper T-cell function and lymphokineactivated killer cytotoxicity following severe head injury. J. Neurosurg. 75: 766–773.
62. Cruse, J. M., R. E. Lewis, Jr., G. R. Bishop, W. F. Kliesch, E. Gaitan, and R. Britt.
1993. Decreased immune reactivity and neuroendocrine alterations related to
chronic stress in spinal cord injury and stroke patients. Pathobiology 61: 183–192.
63. Campagnolo, D. I., D. Dixon, J. Schwartz, J. A. Bartlett, and S. E. Keller. 2008.
Altered innate immunity following spinal cord injury. Spinal Cord 46: 477–481.
64. Mrakovcic-Sutic, I., V. S. Tokmadzic, G. Laskarin, H. Mahmutefendic, P. Lucin,
Z. Zupan, and A. Sustic. 2010. Early changes in frequency of peripheral
blood lymphocyte subpopulations in severe traumatic brain-injured patients. Scand.
J. Immunol. 72: 57–65.
65. Vogelgesang, A., U. Grunwald, S. Langner, R. Jack, B. M. Bröker, C. Kessler, and
A. Dressel. 2008. Analysis of lymphocyte subsets in patients with stroke and their
influence on infection after stroke. Stroke 39: 237–241.
66. Urra, X., A. Cervera, N. Villamor, A. M. Planas, and A. Chamorro. 2009. Harms
and benefits of lymphocyte subpopulations in patients with acute stroke. Neuroscience 158: 1174–1183.
67. Peterfalvi, A., T. Molnar, M. Banati, G. Pusch, E. Miko, L. Bogar, J. Pal,
L. Szereday, and Z. Illes. 2009. Impaired function of innate T lymphocytes and NK
cells in the acute phase of ischemic stroke. Cerebrovasc. Dis. 28: 490–498.
68. Prass, K., C. Meisel, C. Höflich, J. Braun, E. Halle, T. Wolf, K. Ruscher,
I. V. Victorov, J. Priller, U. Dirnagl, et al. 2003. Stroke-induced immunodeficiency
promotes spontaneous bacterial infections and is mediated by sympathetic activation
reversal by poststroke T helper cell type 1-like immunostimulation. J. Exp. Med.
198: 725–736.
69. Bosch, J. A., G. G. Berntson, J. T. Cacioppo, and P. T. Marucha. 2005. Differential
mobilization of functionally distinct natural killer subsets during acute psychologic
stress. Psychosom. Med. 67: 366–375.
70. Segerstrom, S. C., and G. E. Miller. 2004. Psychological stress and the human
immune system: a meta-analytic study of 30 years of inquiry. Psychol. Bull. 130:
601–630.
71. Benschop, R. J., M. Schedlowski, H. Wienecke, R. Jacobs, and R. E. Schmidt.
1997. Adrenergic control of natural killer cell circulation and adhesion. Brain Behav.
Immun. 11: 321–332.
72. Irwin, M., U. Lacher, and C. Caldwell. 1992. Depression and reduced natural killer
cytotoxicity: a longitudinal study of depressed patients and control subjects. Psychol.
Med. 22: 1045–1050.
73. Evans, D. L., J. D. Folds, J. M. Petitto, R. N. Golden, C. A. Pedersen, M. Corrigan,
J. H. Gilmore, S. G. Silva, D. Quade, and H. Ozer. 1992. Circulating natural killer
cell phenotypes in men and women with major depression: relation to cytotoxic
activity and severity of depression. Arch. Gen. Psychiatry 49: 388–395.
74. Frank, M. G., S. E. Hendricks, D. R. Johnson, J. L. Wieseler, and W. J. Burke.
1999. Antidepressants augment natural killer cell activity: in vivo and in vitro.
Neuropsychobiology 39: 18–24.
75. Bush, K. A., K. Krukowski, J. L. Eddy, L. W. Janusek, and H. L. Mathews. 2012.
Glucocorticoid receptor mediated suppression of natural killer cell activity: identification of associated deacetylase and corepressor molecules. Cell. Immunol. 275:
80–89.
76. Denys, D., S. Fluitman, A. Kavelaars, C. Heijnen, and H. Westenberg. 2004.
Decreased TNF-a and NK activity in obsessive-compulsive disorder. Psychoneuroendocrinology 29: 945–952.
77. Enstrom, A. M., L. Lit, C. E. Onore, J. P. Gregg, R. L. Hansen, I. N. Pessah,
I. Hertz-Picciotto, J. A. Van de Water, F. R. Sharp, and P. Ashwood. 2009. Altered
gene expression and function of peripheral blood natural killer cells in children with
autism. Brain Behav. Immun. 23: 124–133.
78. Popa, N., O. Cedile, X. Pollet-Villard, C. Bagnis, P. Durbec, and J. Boucraut.
2011. RAE-1 is expressed in the adult subventricular zone and controls cell proliferation of neurospheres. Glia 59: 35–44.
79. Hamann, I., J. Dörr, R. Glumm, C. Chanvillard, A. Janssen, J. M. Millward,
F. Paul, R. M. Ransohoff, and C. Infante-Duarte. 2013. Characterization of natural
killer cells in paired CSF and blood samples during neuroinflammation. J. Neuroimmunol. 254: 165–169
80. Fu, B., F. Wang, R. Sun, B. Ling, Z. Tian, and H. Wei. 2011. CD11b and CD27
reflect distinct population and functional specialization in human natural killer cells.
Immunology 133: 350–359.
81. Krumbholz, M., D. Theil, F. Steinmeyer, S. Cepok, B. Hemmer, M. Hofbauer,
C. Farina, T. Derfuss, A. Junker, T. Arzberger, et al. 2007. CCL19 is constitutively
expressed in the CNS, up-regulated in neuroinflammation, active and also inactive
multiple sclerosis lesions. J. Neuroimmunol. 190: 72–79.
82. Wilbanks, G. A., and J. W. Streilein. 1992. Fluids from immune privileged sites
endow macrophages with the capacity to induce antigen-specific immune deviation
via a mechanism involving transforming growth factor-b. Eur. J. Immunol. 22:
1031–1036.
83. Marcoe, J. P., J. R. Lim, K. L. Schaubert, N. Fodil-Cornu, M. Matka,
A. L. McCubbrey, A. R. Farr, S. M. Vidal, and Y. Laouar. 2012. TGF-b is responsible for NK cell immaturity during ontogeny and increased susceptibility to
infection during mouse infancy. Nat. Immunol. 13: 843–850.
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017
41.
atitis virus genome results in protection from viral-induced neurological and liver
disease. J. Immunol. 179: 1155–1165.
Shieh, T. M., D. L. Carter, R. L. Blosser, J. L. Mankowski, M. C. Zink, and
J. E. Clements. 2001. Functional analyses of natural killer cells in macaques infected
with neurovirulent simian immunodeficiency virus. J. Neurovirol. 7: 11–24.
Hayashi, T., S. Nagai, H. Fujii, Y. Baba, E. Ikeda, T. Kawase, and S. Koyasu. 2009.
Critical roles of NK and CD8+ T cells in central nervous system listeriosis. J.
Immunol. 182: 6360–6368.
Jin, Y., L. Dons, K. Kristensson, and M. E. Rottenberg. 2001. Neural route of
cerebral Listeria monocytogenes murine infection: role of immune response mechanisms in controlling bacterial neuroinvasion. Infect. Immun. 69: 1093–1100.
Mitchell, A. J., B. Yau, J. A. McQuillan, H. J. Ball, L. K. Too, A. Abtin, P. Hertzog,
S. L. Leib, C. A. Jones, S. K. Gerega, et al. 2012. Inflammasome-dependent IFN-g
drives pathogenesis in Streptococcus pneumoniae meningitis. J. Immunol. 189: 4970–
4980.
Stach, J. L., E. Dufrenoy, J. Roffi, and M. A. Bach. 1986. T-cell subsets and natural
killer activity in Plasmodium falciparum-infected children. Clin. Immunol. Immunopathol. 38: 129–134.
Hansen, D. S., N. J. Bernard, C. Q. Nie, and L. Schofield. 2007. NK cells stimulate
recruitment of CXCR3+ T cells to the brain during Plasmodium berghei-mediated
cerebral malaria. J. Immunol. 178: 5779–5788.
Ryg-Cornejo, V., C. Q. Nie, N. J. Bernard, R. J. Lundie, K. J. Evans, B. S. Crabb,
L. Schofield, and D. S. Hansen. 2013. NK cells and conventional dendritic cells
engage in reciprocal activation for the induction of inflammatory responses during
Plasmodium berghei ANKA infection. Immunobiology 218: 263–271.
Villegas-Mendez, A., R. Greig, T. N. Shaw, J. B. de Souza, E. Gwyer Findlay,
J. S. Stumhofer, J. C. Hafalla, D. G. Blount, C. A. Hunter, E. M. Riley, and
K. N. Couper. 2012. IFN-g-producing CD4+ T cells promote experimental cerebral
malaria by modulating CD8+ T cell accumulation within the brain. J. Immunol.
189: 968–979.
De Chiara, G., M. E. Marcocci, R. Sgarbanti, L. Civitelli, C. Ripoli, R. Piacentini,
E. Garaci, C. Grassi, and A. T. Palamara. 2012. Infectious agents and neurodegeneration. Mol. Neurobiol. 46: 614–638.
Pellicanò, M., A. Larbi, D. Goldeck, G. Colonna-Romano, S. Buffa, M. Bulati,
G. Rubino, F. Iemolo, G. Candore, C. Caruso, et al. 2012. Immune profiling of
Alzheimer patients. J. Neuroimmunol. 242: 52–59.
Speciale, L., E. Calabrese, M. Saresella, C. Tinelli, C. Mariani, L. Sanvito,
R. Longhi, and P. Ferrante. 2007. Lymphocyte subset patterns and cytokine production in Alzheimer’s disease patients. Neurobiol. Aging 28: 1163–1169.
Richartz-Salzburger, E., A. Batra, E. Stransky, C. Laske, N. Köhler, M. Bartels,
G. Buchkremer, and K. Schott. 2007. Altered lymphocyte distribution in Alzheimer’s disease. J. Psychiatr. Res. 41: 174–178.
Martins, L. C., N. P. Rocha, K. C. Torres, R. R. Dos Santos, G. S. França, E. N. de
Moraes, M. A. Mukhamedyarov, A. L. Zefirov, A. A. Rizvanov, A. P. Kiyasov, et al.
2012. Disease-specific expression of the serotonin-receptor 5-HT2C in natural killer
cells in Alzheimer’s dementia. J. Neuroimmunol. 251: 73–79.
Hellstrand, K., and S. Hermodsson. 1987. Role of serotonin in the regulation of
human natural killer cell cytotoxicity. J. Immunol. 139: 869–875.
Oláh, T., I. Ocsovszki, Y. Mándi, R. Pusztai, M. Bakay, and E. Balint. 2005.
Opposite effects of serotonin and interferon-alpha on the membrane potential and
function of human natural killer cells. In Vitro Cell. Dev. Biol. Anim. 41: 165–170.
Mihara, T., M. Nakashima, A. Kuroiwa, Y. Akitake, K. Ono, M. Hosokawa,
T. Yamada, and M. Takahashi. 2008. Natural killer cells of Parkinson’s disease
patients are set up for activation: a possible role for innate immunity in the pathogenesis of this disease. Parkinsonism Relat. Disord. 14: 46–51.
Niwa, F., N. Kuriyama, M. Nakagawa, and J. Imanishi. 2012. Effects of
peripheral lymphocyte subpopulations and the clinical correlation with Parkinson’s
disease. Geriatr. Gerontol. Int. 12: 102–107.
Araga, S., H. Kagimoto, K. Funamoto, and K. Takahashi. 1991. Reduced natural
killer cell activity in patients with dementia of the Alzheimer type. Acta Neurol.
Scand. 84: 259–263.
Solerte, S. B., M. Fioravanti, A. Pascale, E. Ferrari, S. Govoni, and F. Battaini.
1998. Increased natural killer cell cytotoxicity in Alzheimer’s disease may involve
protein kinase C dysregulation. Neurobiol. Aging 19: 191–199.
Solerte, S. B., L. Cravello, E. Ferrari, and M. Fioravanti. 2000. Overproduction of
IFN-g and TNF-a from natural killer (NK) cells is associated with abnormal NK
reactivity and cognitive derangement in Alzheimer’s disease. Ann. N. Y. Acad. Sci.
917: 331–340.
Morandi, B., P. Bramanti, I. Bonaccorsi, E. Montalto, D. Oliveri, G. Pezzino,
M. Navarra, and G. Ferlazzo. 2008. Role of natural killer cells in the pathogenesis
and progression of multiple sclerosis. Pharmacol. Res. 57: 1–5.
Infante-Duarte, C., A. Weber, J. Krätzschmar, T. Prozorovski, S. Pikol, I. Hamann,
J. Bellmann-Strobl, O. Aktas, J. Dörr, J. Wuerfel, et al. 2005. Frequency of blood
CX3CR1-positive natural killer cells correlates with disease activity in multiple
sclerosis patients. FASEB J. 19: 1902–1904.
Saraste, M., H. Irjala, and L. Airas. 2007. Expansion of CD56bright natural killer
cells in the peripheral blood of multiple sclerosis patients treated with interferon-b.
Neurol. Sci. 28: 121–126.
Bielekova, B., M. Catalfamo, S. Reichert-Scrivner, A. Packer, M. Cerna,
T. A. Waldmann, H. McFarland, P. A. Henkart, and R. Martin. 2006. Regulatory
CD56bright natural killer cells mediate immunomodulatory effects of IL-2Ra-targeted
therapy (daclizumab) in multiple sclerosis. Proc. Natl. Acad. Sci. USA 103: 5941–5946.
Airas, L., M. Saraste, S. Rinta, I. Elovaara, Y. H. Huang, and H. Wiendl; Finnish
Multiple Sclerosis and Pregnancy Study Group. 2008. Immunoregulatory factors in
multiple sclerosis patients during and after pregnancy: relevance of natural killer
cells. Clin. Exp. Immunol. 151: 235–243.
5361
5362
84. Vaquero, J., S. Coca, J. Escandón, R. Magallón, and R. Martı́nez. 1990. Immunohistochemical study of IOT-10 natural killer cells in brain metastases. Acta
Neurochir. (Wien) 104: 17–20.
85. Stevens, A., I. Klöter, and W. Roggendorf. 1988. Inflammatory infiltrates and
natural killer cell presence in human brain tumors. Cancer 61: 738–743.
86. Yang, I., S. J. Han, M. E. Sughrue, T. Tihan, and A. T. Parsa. 2011. Immune cell
infiltrate differences in pilocytic astrocytoma and glioblastoma: evidence of distinct
immunological microenvironments that reflect tumor biology. J. Neurosurg. 115:
505–511.
87. Alvarez-Breckenridge, C. A., J. Yu, R. Price, J. Wojton, J. Pradarelli, H. Mao,
M. Wei, Y. Wang, S. He, J. Hardcastle, et al. 2012. NK cells impede glioblastoma
virotherapy through NKp30 and NKp46 natural cytotoxicity receptors. Nat. Med.
18: 1827–1834.
88. Holub, M., Z. Klucková, O. Beran, V. Aster, and A. Lobovská. 2002. Lymphocyte
subset numbers in cerebrospinal fluid: comparison of tick-borne encephalitis and
neuroborreliosis. Acta Neurol. Scand. 106: 302–308.
89. Amor, S., F. Puentes, D. Baker, and P. van der Valk. 2010. Inflammation in
neurodegenerative diseases. Immunology 129: 154–169.
90. Zou, Y., T. Chen, M. Han, H. Wang, W. Yan, G. Song, Z. Wu, X. Wang, C. Zhu,
X. Luo, and Q. Ning. 2010. Increased killing of liver NK cells by Fas/Fas ligand and
NKG2D/NKG2D ligand contributes to hepatocyte necrosis in virus-induced liver
failure. J. Immunol. 184: 466–475.
BRIEF REVIEWS: NK CELLS IN CNS PATHOLOGY
91. Baron, R., A. Nemirovsky, I. Harpaz, H. Cohen, T. Owens, and A. Monsonego.
2008. IFN-g enhances neurogenesis in wild-type mice and in a mouse model of
Alzheimer’s disease. FASEB J. 22: 2843–2852.
92. Gaddi, P. J., M. J. Crane, M. Kamanaka, R. A. Flavell, G. S. Yap, and T. P. SalazarMather. 2012. IL-10 mediated regulation of liver inflammation during acute murine
cytomegalovirus infection. PLoS ONE 7: e42850.
93. Huang, D., F. D. Shi, S. Jung, G. C. Pien, J. Wang, T. P. Salazar-Mather, T. T. He,
J. T. Weaver, H. G. Ljunggren, C. A. Biron, et al. 2006. The neuronal chemokine
CX3CL1/fractalkine selectively recruits NK cells that modify experimental autoimmune encephalomyelitis within the central nervous system. FASEB J. 20: 896–
905.
94. Durrenberger, P. F., A. Ettorre, F. Kamel, L. V. Webb, M. Sim, R. S. Nicholas,
O. Malik, R. Reynolds, R. J. Boyton, and D. M. Altmann. 2012. Innate immunity
in multiple sclerosis white matter lesions: expression of natural cytotoxicity triggering receptor 1 (NCR1). J. Neuroinflammation 9: 1.
95. Al Nimer, F., R. Lindblom, M. Ström, A. O. Guerreiro-Cacais, R. Parsa,
S. Aeinehband, T. Mathiesen, O. Lidman, and F. Piehl. 2013. Strain influences on
inflammatory pathway activation, cell infiltration and complement cascade after
traumatic brain injury in the rat. Brain Behav. Immun. 27: 109–122.
96. Vaquero, J., M. Zurita, and S. Coca. 2012. Clinical behavior in metastatic brain
disease is not influenced by the immunological defense mediated by CD57 NKcells. Int. J. Surg. Oncol. 2012: 834852.
Downloaded from http://www.jimmunol.org/ by guest on May 7, 2017