Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Feature detection (nervous system) wikipedia , lookup
Signal transduction wikipedia , lookup
Multielectrode array wikipedia , lookup
Haemodynamic response wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Optogenetics wikipedia , lookup
Synaptogenesis wikipedia , lookup
Neuroanatomy wikipedia , lookup
Subventricular zone wikipedia , lookup
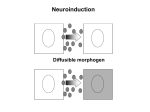
Neural Induction Chapter One Development of the Nervous System Fertilization Cleavage (Blastula, Gastrula) Neuronal InductionNeuroblast Formation Mesodermal Induction Lateral Inhibition Axis Formation Cell Migration Axon Growth/Target innervation Differentiation Functional Nervous System Cell Lineage Cell-cell Interactions Spermann Organizer Transcription Factors Homeobox genes Chemical Gradients Cell-Cell Interactions Chemical Gradients Cell-Cell Interactions Synaptic Formation Electrical Differentiation Cell-Cell Interactions Trophic Factors Mechanisms of nervous system development Gastrula (Mesodermal induction via BMP signal inhibition) Induction (Lateral Inhibition via regulation of transcription factors) Neuroblast (Cell migration, Differentiation) Functional Nervous System Questioning Developmental Biologist started looking: • Determine the time line of differentiation into different cell types • What was determine by cell’s position? • What part did cell to cell interactions play? • What was the inducing factor? • What was required to form neuronal tissue? Factors regulating development of the nervous system Before After • Interactions with neighboring tissue is required for ectodermal differentiation into nervous tissue • Neuronal lineage arises during gastrulation • Isolation of animal cap before gastrulation gives origin to epidermal cells. • If the animal cap is removed after gastrulation, it will give origin to neurons. • These experiments suggest that tissue arrangements during gastrulation is necessary for induction of nerve tissue • In Amphibian, the involuting mesoderm (dorsal lip of the blastopore) induces neuronal differentiation of the ectoderm • Transplant of a dorsal blastopore gives rise to a new body axis. Therefore the dorsal lip of the blastopore “organize” the formation of a new body axis. This structure has received the named of Spemann organizer …even at the gastrula stage the head organizer is not an equipotential entity, but it is subdivided into specialized inductors although distinct boundaries between then do not seem to exist….Holtfreter, 1939 • Tissue in the dorsal lip of the blastopore contain multiple inducers: subdivision of the dorsal lip and transplant to another embryo give origin of either tails or head-like structures. Medial portion of DLB Lateral portion of DLB Generate head Generate tail structures What chemical factors are involved in neuronal induction by the mesoderm? • The use of a expression cloning system identified noggin as the first neuronal inducer – Noggin • Noggin is a 26 kD protein expressed in the dorsal lip of the blastopore during gastrulation (but not in the blastula stage) • Noggin cDNA injection into UV treated embryos result in normal development of the nervous system • Follistatin signaling has been identifies as another important pathway of neuronal induction • During development follistatin binds to and inhibits activin (a TGF-β-like molecule) • Follistatin is expressed in the organizer region of the embryo during gastrulation • Overexpression of a truncated form of the activin receptor result in differentiation of animal cap cells into neurons without any neuronal inducing molecule • Misexpression of follistatin causes the formation of a secondary axis • These experiments lead to the idea that neuronal induction by the mesoderm may involve inhibition by activin receptors • Neuronal differentiation of ectoderm in actively inhibited by cell-cell interactions • Dissociation of animal caps cells causes most cells to become neurons, whereas cells maintained as aggregates develop as epidermal cells • So it appears that the ectoderm is actively inhibited from becoming neuronal tissue Adding BMP4 Blocks dissociated cells from becoming neural tissue What chemical factors are involved in neuronal induction? • The idea that the ectoderm is actively inhibited from differentiating into neuronal tissue was further proved with the discovery of chordin • Chordin overexpression cause generation of a secondary axis similar to transplanting a Spemann organizer into another embryo How does chordin induce neuronal differentiation? Fly Sog Dpp = = Frog Chordin BMP-4 • Studies in Drosophila indicate that the chordin-like protein sog bindings to the TGF-β-like receptor dpp related to the vertebrate genes bone-morphogenic proteins, BMP-2 and BMP-4 • Sog and Dpp antagonize each other in Drosophila • Overexpression of sog (ie, chordin) stimulate neuronal differentiation. Injection of BMP-4 inhibits neuronal differentiation. • The antagonistic action of sog and dpp lead to the idea that neuronal inducers (noggin, follistatin, chordin) stimulate neuronal differentiation by inhibiting BMP-4 signaling • Overexpression of BMP-4 inhibits neuronal differentiation of animal caps treated with noggin, chordin, or follistatin • Antisense BMP-4 RNA causes neuronal differentiation without any any neuronal inducing molecule Multiple Inducers Selectively Promote Different Regions of the Nervous System The Spemann's organizer secretes many proteins that bind to different growth factors in the extracellular space. Each one blocks the signaling through their cognate receptors Antagonist Ligands Mesodermal Induction in Vertebrates • Experiment in zebra fish have confirmed that in vertebrates formation of the neural tube requires inhibition of BMP signal Gene Interactions • Ectoderm cells start out fairly similar • Bone Morphogenetic Proteins (BMP) cause them to form skin and bone • BMP inhibitors cause them to form neurons instead – Examples Chordin and Noggin Cell-cell Interactions Control Neuroblast Segregation • The noggin, chordin, follistatin molecules only determine the generation of a neurogenic region in the embryo • The process of neurobalst formation (ie, neuronal precursors) has been studied in Drosophila where delamination results in the formation of precursor neuronal tissue Delamination and Formation of Ganglion Mother Cells in Drosophila • First division of neuroblast cells generate a ganglion mother cell (GMC) • Each GMC generates one pair of neurons (or glial cells) Does neuroblast formation require interaction among neighboring cells? • Laser ablation of a delaminating cell causes another cells to delaminate • This lead to the idea that lateral inhibition maybe responsible for the neuroblast formation in Drosophila • It was found that a family of transcription factors are required for this process to occur Lateral Inhibition • Ectodermal cells are actively inhibited from becoming neuronal cells • If they are physically separated • Or if they do not receive inhibition signal then they will become neuronal cells • In Drosophila, segregation of neuroblast from epidermal cells requires the expression of bHLH transcription factors (basic-helixloop-helix transcription factors) • bHLH transcription factors activate transcription by binding to specific DNA sites called Eboxes (CANNTG) • In Drosophila, the achaete scute family of transcription factors (ACS) is required for proper development of neuroblasts. Because their role in neuroblasts formation these genes are called proneural genes • This family of genes were identified for their effect on the development of the fly’s bristles How does the process of lateral inhibition works? Some ectodermal cells express ACS proneural genes ACS expression increases in one of the cells in the center of the cluster Lateral inhibition blocks proneural genes in neighboring cells Proneural Genes • Achaete scute (ACS) – Transcription factors • Induce neuronal development • Once one cell begins expressing ACS, surrounding cells are laterally inhibited Lateral Inhibition Requires the Delta-Notch System • Delta is the endogenous ligand for the Notch receptor (a large transmembrane protein) • Low Notch activity induces neuroblast formation. Thus, in Notch mutants most of the cells in the neurogenic region become neuroblasts Delta Y Notch • Delta mutants undergo change in same direction The Delta-Notch Signaling • Cleavage of the Notch receptor after binding to Delta allows for the diffusion of the Notch tail to the nucleus and regulation of transcription • During development interactions between Delta and Notch induce molecular differences between neighboring cells Lateral Inhibition Requires the Delta-Notch System • Cells within the proneural cluster express high levels of ASC as well as Delta • If by chance one cell express higher levels of ACS, this cell will express higher levels of Delta Ectodermal cell Notch Activation Results in Cells Adopting an Epidermal Fate • Upregulation of Delta expression in the central cell will activate Notch in surrounding cells • This will lead to release of the SuH (supressor hairless) from Notch • SuH translocate to the nucleus where it suppresses neuronal gene expression (ie, ASC) Ectodermal cell Notch Activation Results in Cells Adopting an Epidermal Fate • SuH inhibits neuronal gene expression by regulating the expression of another set of bHLH proteins called enhancer of split complex (Espl) • E-spl act as repressors of transcription (notice that ASC genes are transcriptional activators) Ectodermal cell • E-spl proteins act as repressors of transcription by binding to the E-boxes in the ASC promoter Tight Control • Delta/Notch and SuH block neuronal differentiation in surrounding cells • This is how lateral inhibition is controlled • ACS is proneural transcription factor • SuH is inhibiting transcription factor • Vertebrate homologs of Drosophila ASC have been identified (Mash1, Cash1, Xash1, NeuroD) • These genes act as transcriptional activators similar to ACS • Overexpression of the bHLH gene NeuroD in frog embryos result in significant neuronal formation throughout the epidermis Current Model: Default is epidermis Fly Delta Notch SuH ACS Vertebrates Wnt Frizzled Msx Sox Necessary vs. Sufficient • Necessary = must be present • Sufficient = all that is needed • BMP antagonism is necessary and sufficient to induce a cell to become a neuron • Notch/Delta pathway is necessary and sufficient for inhibition of surrounding cells so that they do NOT become neurons Necessary vs. Sufficient • If you had no BMP antagonism what would the ectoderm look like? • If you had only BMP antagonism what would the ectoderm look like? • If you had only the Notch/Delta pathway? • What if you have both BMP antagonism and the Notch/Delta pathway? Any Questions? Read Chapter Two