Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Caridoid escape reaction wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Synaptic gating wikipedia , lookup
Haemodynamic response wikipedia , lookup
Optogenetics wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
End-plate potential wikipedia , lookup
Proprioception wikipedia , lookup
Electromyography wikipedia , lookup
Central pattern generator wikipedia , lookup
Synaptogenesis wikipedia , lookup
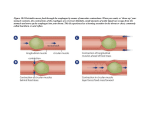
Diseases of the Esophagus (2012) 25, 292–298 DOI: 10.1111/j.1442-2050.2011.01180.x Original article dote_1180 292..298 Anatomy and physiology of the esophageal body E. Yazaki, D. Sifrim Barts and The London School of Medicine and Dentistry, Queen Mary University of London, London, UK SUMMARY. The primary role of the esophagus is to propel swallowed food or fluid into the stomach and to prevent or clear gastroesophageal reflux. This function is achieved by an organized pattern that involves a sensory pathway, neural reflexes, and a motor response that includes esophageal tone, peristalsis, and shortening. The motor function of the esophagus is controlled by highly complex voluntary and involuntary mechanisms. There are three different functional areas in the esophagus: the upper esophageal sphincter, the esophageal body, and the LES. This article focused on anatomy and physiology of the esophageal body. KEY WORDS: anatomy, esophagus, physiology. ANATOMY The esophagus is a tubular organ with approximately 18–26 cm of length in adults. The esophageal wall is composed of the mucosa, submucosa, and muscularis propria. In contrast to in other parts of the gastrointestinal tract, the esophagus does not have serosal covering, and the outer wall is bounded with a thin layer of connective tissue. The mucosa of the esophageal body is made of stratified squamous epithelium, composed of three sublayers: mucous membrane, lamina propria, and muscularis mucosae. Esophageal submucosa contains glands (esophageal submucosal glands), although numbers are fewer than that observed in the rest of the gut. The esophageal muscularis propria is composed of a circular muscle layer that is surrounded by a longitudinal muscle layer. The proximal esophagus (cervical esophagus) is composed of striated muscle, and the distal two third of the esophagus (thoracic esophagus) contains smooth muscles. The transition from striated to smooth muscle is gradual and there is a segment (4–6 cm long) that contains both striated and smooth muscles. Meyer et al.1 reported that this mixed zone of muscle fibers was approximately 35% of the esophageal length. This segment is usually present around the level of the aortic arch. EsoAddress correspondence to: Dr Etsuro Yazaki, MD, PhD, Wingate Institute of Neurogastroenterology, 26 Ashfield Street, London E1 2AJ, UK. Email: [email protected] 292 phageal manometry confirms this anatomical zone by demonstrating that the amplitude of the contraction wave reaches a minimum in this region. The esophagus runs posterior to the right of the aortic arch at the T4 vertebral level. From the level of T8 until the diaphragmatic hiatus, the esophagus lays anteriorly to the aorta. The esophagus then runs through an opening of the diaphragm that is located anterior to the opening for the descending aorta. Under normal anatomy, the esophagus is anchored to the diaphragm, but the esophagus does not tightly fill the hiatus because it needs to expand to accommodate luminal contents. With aging or other causes, the amount of elastic tissue in the phrenoesophageal membrane gradually declines, increasing its laxity and resulting in risk for developing hiatal hernia. The esophagus has parasympathetic and sympathetic innervation. The parasympathetic nerve supply comes from the nucleus ambiguus and the dorsal motor nucleus of the vagus (DMN) and provides motor innervation to the esophageal muscles and secretomotor innervation to the glands. The sympathetic nerve supply comes from the intermediolateral cell columns of the T1–T10 spine and regulates contractions of blood vessel (blood supply), esophageal sphincter tone, relaxation of the muscular wall, and increase in glandular function. The motor innervation of the esophagus is principally via the vagus nerves. The striated muscle part of the esophagus is controlled by lower motor neurons in the nucleus ambiguous in the brainstem, © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus Anatomy and physiology of the esophagus i.e. swallowing center. The nerve fibers are of the somatic type and terminate on motor endplates in the striated muscle fibers. In the smooth muscle segment, the nerve fibers originate in the DMN. There are two distinct types of DMN preganglionic neurons. One type (short latency fibers), in the caudal part of the DMN, synapses on postganglionic nitrergic inhibitory neurons in the myenteric (Auerbach’s) plexus, and the other type (long latency fibers), in the rostral part of the DMN, synapses on the postganglionic cholinergic excitatory neurons in the myenteric plexus.2,3 The ganglia of the submucosal (Meissner’s) plexus regulate secretion and the contraction of the muscularis musosae. The significance of a scattered myenteric plexus in the proximal esophageal striated muscle remains unclear.4,5 Arterial blood supply to the esophagus is provided by three sources: branches of the inferior thyroid artery provide arterial blood supply to the upper esophageal sphincter (UES) and the cervical esophagus; paired aortic esophageal arteries or terminal branches of bronchial arteries provide blood supply to the thoracic esophagus; and the left gastric artery and a branch of the left phrenic artery provide blood supply to the LES and the most distal segment of the esophagus. A continuous intensive network arteries located in the submucosa connects all the extramural vessels, which results in the copious blood supply to the esophagus. Blood from the proximal and distal esophagus is drained via the azygous system, which in turn drains directly into the superior vena cava. The left gastric vein, a branch of the portal vein, receives venous blood from the mid-esophagus. The submucosal connections of veins between the portal and systemic venous systems in the distal esophagus can form esophageal varices with portal hypertension. PHYSIOLOGY Esophageal secretion The esophageal submucosa contains glands, although there are fewer numbers of glands than in the rest of the gastrointestinal tracts. The esophageal submucosal glands secrete the following substances: water, bicarbonate, mucins, epidermal growth factor, and prostaglandins. Secretions of the esophageal submucosal glands are involved in mucosal clearance along with peristalsis and salivary secretion. The most important secreted substance is bicarbonate, which plays a protective role during gastroesophageal reflux.6 This is more significant when non-salivary source of bicarbonate is required to neutralize luminal acid, e.g. during sleeping.7 Sensory physiology Vagal afferents merging from the esophageal smooth muscle layer and serosa are sensitive to muscle 293 stretch, whereas vagal afferents in the mucosa are sensitive to various stimuli including chemical (acid), thermal (cold or hot), and mechanical intraluminal stimuli.8 In general, vagal afferents do not play a direct role in visceral pain transmission to the brainstem, but spinal afferents, which have their cell bodies in the dorsal root ganglia, are predominantly acting as nociceptors.8 Spinal afferents terminate in the dorsal column nuclei (gracilia and cuneatus nuclei) and project stimuli to the brain through the spinoreticular, spinomesencephalic, spinohypothalamic, and spinothalamic tracts.9,10 Esophageal pain is known to resemble one from cardiac origin, i.e. noncardiac chest pain. This resemblance is due to convergence of sensory afferent fibers from the heart and esophagus in the same spinal dorsal horn neuron in the cervical and thoracic spinal cord.11–13 The concept of visceral hypersensitivity is now well-established in esophageal symptoms.14 Changes in esophageal sensitivity can be caused by peripheral or central sensitization (neurons in the spinal cord and brain). Peripheral sensitization has been reported to be due to upregulation of acidsensing receptors. Acid can activate two proton-gated ion channels: transient receptor potential vanilloid-1 and acid-sensing ion channels, and activation of these channels can cause neurogenic inflammation resulting in sensitization of peripheral neurons.15,16 More recently, dilated intercellular spaces in the basal layer of the esophageal epithelium have been suggested to play an important role in peripheral sensitization in nonerosive reflux disease by favoring increased acid penetration through the esophageal mucosa.17,18 For the central sensitization, acid could induce the release of proinflammatory substances that can sensitize sensory neurons and would result in the central sensitization.19,20 Another concept, psychoneuroimmune interactions, has recently been described as one of the mechanisms for hypersensitivity in nonerosive gastroesophageal reflux disease.14 The mechanisms of psychoneuroimmune interactions can be explained by alterations in mucosal permeability, immune activation and pain perception.13,14 Motor physiology Primary and secondary peristalsis. The coordinated motor pattern of the esophagus initiated by the act of swallowing is called primary peristalsis. A rapidly progressing pharyngeal contraction wave transfers the bolus through the relaxed UES into the esophageal body, and a progressive circular contraction begins in the upper esophagus and proceeds distally along the esophageal body to propel the bolus through the relaxed LOS. The mean peak pressures amplitudes are 53 ⫾ 9 mmHg in the proximal esophagus, 35 ⫾ 6 mmHg in the middle, and 70 ⫾ 12 mmHg © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus 294 Diseases of the Esophagus in the distal esophagus.21 The duration of the wave is normally 2–7 seconds, and the propagation velocity is 2 to 4 cm/second.21 Primary peristalsis usually clears most contents of the esophagus into the stomach, but with ineffective peristalsis, there may be food residue. Secondary peristalsis is provoked by residual food or reflux events, and it is not accompanied by pharyngeal contraction or UES relaxation. The amplitude and velocity of secondary peristaltic waves are similar to those of primary peristalsis. Secondary peristalsis in the striated muscle esophagus is regulated by similar central mechanism as primary peristalsis.22 The control of secondary peristalsis in the smooth muscle part of the esophagus is a local reflex without demonstrated deglutitive inhibition.23 In secondary peristalsis, mechanoreceptors in the esophageal wall are excited by distension to initiate a reflex that may traverse both the extrinsic and intrinsic nerve pathways. The peripheral reflex excites an inhibitory innervation, which releases nitric oxide (NO) to produce inhibition distal to the level of distension.24 Deglutitive inhibition. Studies using animal models demonstrated that esophageal smooth muscle was hyperpolarized before the occurrence of a peristaltic contraction.25–28 The muscle hyperpolarization lasts progressively longer in the distal esophageal segments and is followed by depolarization. Because muscle contraction is inhibited during hyperpolarization, the timing of esophageal peristaltic contractions is influenced by the strength and duration of this inhibition. Because of methodological limitations, electrophysiological measurements of this hyperpolarization in the esophageal smooth muscle was never demonstrated in humans, although this was suggested by the phenomenon called ‘deglutitive inhibition’ where swallowing inhibits ongoing muscular activity in the esophagus.3,29,30 During multiple rapid swallowing (MRS), the esophageal muscle activity is inhibited, and only the last swallow is associated with a powerful rebound, peristaltic sequence known as the after MRS contraction. Sifrim et al.31 reported the first direct evidence that deglutition produces in the human esophagus a wave of inhibition that precedes the primary peristaltic contraction. In this study, a balloon placed in the esophagus was inflated at various positions, and esophageal motor responses to swallows were studied. They reported that the duration of swallow-induced fall in pressure (inhibition) at 13 cm from the LES is shorter than that at 8 cm (Fig. 1). This study showed that the latency period before the onset of the peristalsis was a period of inhibition and that there was a gradient of increasing duration of deglutitive inhibition distally along the human esophagus. Control of esophageal peristalsis. Peristalsis in the striated muscle part of the esophagus is entirely Fig. 1 Sifrim et al. demonstrated the first direct evidence of a deglutitive inhibition in the human esophagus. A balloon placed in the esophagus was inflated at various positions, and esophageal motor responses to swallows were studied. The duration of swallow-induced fall in pressure (inhibition) at 13 cm from the LES is shorter than that at 8 cm. dependent on central vagal pathways. It is mediated by sequential excitation of lower motor neurons originated in the nucleus ambiguus through the vagus nerve (Fig. 2).32,33 Control of peristalsis in the smooth muscle part of the esophagus is more complicated. Peristalsis in the thoracic esophagus is mediated by both central and peripheral mechanisms. In contrast to the striated muscle portion, sequential excitation of vagal efferent fibers is not essential to exhibit peristaltic waves.3,34 The timing of peristalsis in the smooth muscle segment is based on the duration of the deglutitive inhibition that increases distally along the esophagus 1 Nucleus Ambiggus 2 3 UES 1 2 3 Vagus Striated muscle Smooth muscle part Fig. 2 Peristalsis in the striated muscle part of the esophagus is entirely dependent on central vagal pathways. It is mediated by sequential excitation of lower motor neurons originated in the nucleus ambiguus through the vagus nerve. © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus Anatomy and physiology of the esophagus (the latency gradient) followed by deglutitive rebound excitation.35,36 Weisbrodt and Christensen37 first reported that a latency gradient along the esophagus was due to noncholinergic inhibitory nerves in the esophageal wall. Crist et al.2 reported that the influence of noncholinergic nerve is minimal at the proximal esophagus and increasing distally and the cholinergic influence is maximal in the proximal esophagus and decreasing distally. These studies suggested that that preprogrammed intramural mechanisms, composed of inhibitory (noncholinergic) and excitatory (cholinergic) neurons, were responsible for peristalsis in the esophageal smooth muscle together. A myogenic control system (MCS) is also reported to regulate esophageal smooth muscle contractions.38–40 MCS is a fundamental property of contraction of gastrointestinal smooth muscle.40 There are two important characteristics in MCS: oscillation of the electrical control activity of the so-called slow waves and coupling of the smooth muscle cells in order to work as a functional unit. A central mechanism also plays an important role in control of peristalsis in the esophageal smooth muscle. There are two types of vagal motor fibers that originate in the DMN: short and long-latency fibers.3 On swallowing, there is near-simultaneous activation of the short-latency inhibitory vagal fibers that inhibits the entire length of the esophagus.3 This inhibition is due to a release of NO released from the myenteric inhibitory neurons.41 There is a delayed and sequential activation of the cholinergic pathway in the longlatency vagal fibers. These cholinergic excitations only occur after the sequential termination of deglutitive inhibition, and the balance of timing in inhibition and excitation is the fundamental mechanism that regulates esophageal peristalsis (Fig. 3). New insights into esophageal motor physiology based on observations during high resolution esophageal manometry. Kahrilas et al.42 described using simultaneous manometry and videofluoroscopy abnormal bolus clearance associated with hypotensive peristalsis in the aortic arch region of the esophagus. Based on this study, Li et al.43 developed a mathematical model of the esophageal bolus transport. They hypothesized that bolus retention in the aortic arch region, i.e. transition zone (TZ), could be due to ‘mismatch’ of the upper (striated muscle part) and lower (smooth muscle part) peristaltic waves. This hypothesis was further studied by Ghosh et al.,44 who analyzed data obtained from concurrent 21 lumen manometry and videofluoroscopy. They reported that two separate waves – above and below the TZ – were a part of normal esophageal motor physiology. The same group, using concurrent 36ch high-resolution manometry (HRM) and digital fluoroscopy, confirmed that impaired coordination between these two waves was associated with bolus retention in the TZ.45 295 rDMN cDMN Excitation Inhibition Inhibition Inhibition Smooth muscle Inhibition & contractions LES Fig. 3 On swallowing, there is near-simultaneous activation of the short-latency inhibitory vagal fibers in the caudal dorsal motor nucleus of the vagus (cDMN) that inhibits the entire length of the esophagus by releasing nitric oxide from the myenteric inhibitory neurons. There is a delayed activation of the cholinergic pathway in the long-latency vagal fibers in the rostral DMN (rDMN). These cholinergic excitations only occurs after the sequential termination of deglutitive inhibition. In 1991, Clouse and Staiano46 first described esophageal pressure topography using conventional manometry and noted different segmental contractions (S1–S3). Details of these segmental contractions have been described further with HRM.47,48 The S1 segment originates from the UES and involves the striated muscle esophagus. S2 and S3 segments involve smooth muscle esophagus and are predominantly controlled by excitatory (cholinergic) and inhibitory (NO) neurons, respectively.2,35 Pandolfino et al.48 recently reported the concept of the contractile deceleration point (CDP) as an important physiological landmark on esophageal pressure topography. Using combined HRM and videofluoroscopy, they found that the CDP of esophageal contractile front velocity did not occur at the border between S2 and S3 as previously thought,49 but CDP occurred within S3 and was indicative of the transition from peristaltic bolus transport to onset of emptying of the phrenic ampulla. The concept of CDP reflects the progression of ampullary emptying and can help to better understand bolus transport across the esophagogastric junction. Apart from the aforementioned physiological observations, based on the HRM, a new classification of esophageal motility disorders has been described (Chicago Classification),47,50 providing a new approach to esophageal motor physiology and pathophysiology. Using the HRM, normal esophageal peristalsis can be defined as >30 mmHg isobaric contour with <3 cm defect, contraction front © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus 296 Diseases of the Esophagus velocity <8 cm, intrabolus pressure <15 mmHg, and distal contractile integral (DCI) <5000 mmHg/ second/cm.47,50 Longitudinal muscle contraction – esophageal shortening. Contraction of the esophageal longitudinal smooth muscle is responsible for shortening of the esophageal wall during peristalsis.51 Although half of the esophageal mucularis propria is composed of longitudinal muscle, its role in human esophageal physiology and pathophysiology is not completely understood. This is partly due to technical difficulty in measuring longitudinal muscle contractions in human and studies were mainly performed on animal models.52,53 A number of techniques have been reported to assess longitudinal muscle contractions in vivo. Radio-opaque markers endoscopically clipped along the long axis of the esophagus have been used.54,55 This technique involves X-ray fluoroscopy and is not suitable for prolonged measurements. Recently, highfrequency intraluminal ultrasound (HFIU) has been introduced as a valid tool to measure longitudinal muscle contractions in human.56–58 Longitudinal muscles can play an important role in motor physiology of the esophagus. Dodds et al.59 suggested that both the longitudinal and circular esophageal muscular contractions were vital to achieve efficient peristaltic transport of a bolus through the esophagus. Pouderoux et al.54 reported that both circular and longitudinal muscle contractions occurred during peristalsis, and the longitudinal muscle contractions led circular muscle contractions. Mittal et al.57 suggested that synchrony between circular and longitudinal muscle contractions was important as the synchrony created maximal increase in esophageal muscle thickness, and therefore, circular muscles could efficiently exhibit peak pressure contractility. Esophageal shortening is also very important to produce lower esophageal sphincter axial movement and opening. This is true during swallowing and also during gastroesophageal reflux. A localized distal esophageal shortening has been reported during transient lower esophageal sphincter relaxation (TLESR),60,61 and it seems to be critical for reflux to occur during a TLESR. Abnormal longitudinal muscle contraction and shortening may be associated to pathology and symptoms: esophageal acidification is known to induce contraction of the longitudinal muscle.62,63 Paterson et al.63 have demonstrated in an opossum’s esophagus that intraluminal acid activates a reflex pathway involving mast cell degranulation, activation of capsaicin-sensitive afferent neurons, and the release of substance P or a related neurokinin, which evokes sustained contraction of the esophageal longitudinal smooth muscle. This reflex could be initiated by changes in mucosal permeability (dilated intercellular spaces) induced by mucosal acidification.64 These results suggest that increased esophageal acid exposure can be a possible cause of exaggerated chronic longitudinal muscle contraction, leading to formation of hiatus hernia.65 Several studies have demonstrated that the esophageal hypermotility (peristalsis) is poorly correlated to noncardiac chest pain.66–69 Studies using HFIU described long-lasting thickening of the esophageal wall (sustained esophageal contraction) associated with chest pain or heartburn.56,70,71 Longitudinal muscle spasm or shortening can therefore be an important mechanism for esophageal symptoms. Esophageal body tone. Based on in vitro studies and manometric studies in vivo, it has been traditionally believed that the esophageal body has no resting tone. In 1993, Mayrand and Diamant72 first demonstrated using a modified barostat that active tone was indeed present in the smooth muscle esophagus, and compliance and resting tone differed between the smooth and striated muscle segments of the esophagus. This finding was confirmed by other investigators reporting the presence of the tonic contractions in the esophageal body.53,73,74 Little is known about the relevance and neural modulation of esophageal tone. Zhang et al.75 demonstrated that tonic contractile activity was strongly modulated by a cholinergic neural excitatory input. Bilateral vagotomy and atropine provoked more than 50% decrease in esophageal tone. They also suggested that NO may have a complementary inhibitory role. Esophageal tone was decreased by sildenafil and increased by L-NNA. Esophageal body tone might compensate the negative intrathoracic pressure induced by the respiration. A tonic esophageal wall might be relevant as a background for the occurrence and organization of efficient phasic peristaltic contractions. It has been reported that changes in esophageal tone might modulate the proximal extent of gastroesophageal reflux.76 It is thought that changes in esophageal tone could be one of the important components in normal swallowing along with circular muscle contraction (peristalsis) and contraction of longitudinal muscle (shortening), although further studies are required to understand the significance of this low resting tone in motor physiology of esophagus. References 1 Meyer G, Austin R, Brady C R, Castell D. Muscle anatomy of the human esophagus. J Clin Gastroenterol 1986; 8: 131–4. 2 Crist J, Gidda J, Goyal R. Intramural mechanism of esophageal peristalsis: roles of cholinergic and noncholinergic nerves. Proc Natl Acad Sci USA 1984; 81: 3595–9. 3 Gidda J, Goyal R. Regional gradient of initial inhibition and refractoriness in esophageal smooth muscle. Gastroenterology 1985; 89: 843–51. 4 Christensen J, Stiles M, Rick G, Sutherland J. Comparative anatomy of the myenteric plexus of the distal colon in eight mammals. Gastroenterology 1984; 86: 706–13. © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus Anatomy and physiology of the esophagus 5 Aziz Q, Rothwell J, Barlow J et al. Esophageal myoelectric responses to magnetic stimulation of the human cortex and the extracranial vagus nerve. Am J Physiol 1994; 267 (Pt 1): G827–35. 6 Long J, Orlando R. Esophageal submucosal glands: structure and function. Am J Gastroenterol 1999; 94: 2818–24. 7 Dent J, Dodds W, Friedman R et al. Mechanism of gastroesophageal reflux in recumbent asymptomatic human subjects. J Clin Invest 1980; 65: 256–67. 8 Fass R. Sensory testing of the esophagus. J Clin Gastroenterol 2004; 38: 628–41. 9 Mayer E, Gebhart G. Basic and clinical aspects of visceral hyperalgesia. Gastroenterology 1994; 107: 271–93. 10 Almeida T, Roizenblatt S, Tufik S. Afferent pain pathways: a neuroanatomical review. Brain Res 2004; 1000: 40–56. 11 Qin C, Chandler M, Foreman R. Esophagocardiac convergence onto thoracic spinal neurons: comparison of cervical and thoracic esophagus. Brain Res 2004; 1008: 193–7. 12 Qin C, Farber J, Foreman R. Gastrocardiac afferent convergence in upper thoracic spinal neurons: a central mechanism of postprandial angina pectoris. J Pain 2007; 8: 522–9. 13 Garrison D, Chandler M, Foreman R. Viscerosomatic convergence onto feline spinal neurons from esophagus, heart and somatic fields: effects of inflammation. Pain 1992; 49: 373–82. 14 Knowles C, Aziz Q. Visceral hypersensitivity in non-erosive reflux disease. Gut 2008; 57: 674–83. 15 Patterson L, Zheng H, Ward S, Berthoud H. Vanilloid receptor (VR1) expression in vagal afferent neurons innervating the gastrointestinal tract. Cell Tissue Res 2003; 311: 277–87. 16 Miranda A, Nordstrom E, Mannem A, Smith C, Banerjee B, Sengupta J. The role of transient receptor potential vanilloid 1 in mechanical and chemical visceral hyperalgesia following experimental colitis. Neuroscience 2007; 148: 1021–32. 17 van Malenstein H, Farré R, Sifrim D. Esophageal dilated intercellular spaces (DIS) and nonerosive reflux disease. Am J Gastroenterol 2008; 103: 1021–8. 18 Farré R, Fornari F, Blondeau K et al. Acid and weakly acidic solutions impair mucosal integrity of distal exposed and proximal non-exposed human oesophagus. Gut 2010; 59: 164–9. 19 Woolf C. Evidence for a central component of post-injury pain hypersensitivity. Nature 1983; 306: 686–8. 20 Woolf C, Shortland P, Coggeshall R. Peripheral nerve injury triggers central sprouting of myelinated afferents. Nature 1992; 355: 75–8. 21 Richter J, Wu W, Johns D et al. Esophageal manometry in 95 healthy adult volunteers. Variability of pressures with age and frequency of ‘abnormal’ contractions. Dig Dis Sci 1987; 32: 583–92. 22 Bardan E, Xie P, Aslam M, Kern M, Shaker R. Disruption of primary and secondary esophageal peristalsis by afferent stimulation. Am J Physiol Gastrointest Liver Physiol 2000; 279: G255–61. 23 Pandolfino J, Shi G, Zhang Q, Kahrilas P. Absence of a deglutitive inhibition equivalent with secondary peristalsis. Am J Physiol Gastrointest Liver Physiol 2005; 288: G671–6. 24 Christensen J. Mechanisms of secondary esophageal peristalsis. Am J Med 1997; 103: 44S–6S. 25 Goyal R, Gidda J. Relation between electrical and mechanical activity in esophageal smooth muscle. Am J Physiol 1981; 240: G305–11. 26 Rattan S, Gidda J, Goyal R. Membrane potential and mechanical responses of the opossum esophagus to vagal stimulation and swallowing. Gastroenterology 1983; 85: 922–8. 27 Sugarbaker D, Rattan S, Goyal R. Mechanical and electrical activity of esophageal smooth muscle during peristalsis. Am J Physiol (Pt 1) 1984; 246: G145–50. 28 Paterson W. Electrical correlates of peristaltic and nonperistaltic contractions in the opossum smooth muscle esophagus. Gastroenterology 1989; 97: 665–75. 29 Meyer G, Gerhardt D, Castell D. Human esophageal response to rapid swallowing: muscle refractory period or neural inhibition? Am J Physiol 1981; 241: G129–36. 30 Vanek A, Diamant N. Responses of the human esophagus to paired swallows. Gastroenterology 1987; 92: 643–50. 31 Sifrim D, Janssens J, Vantrappen G. A wave of inhibition precedes primary peristaltic contractions in the human esophagus. Gastroenterology 1992; 103: 876–82. 297 32 Bieger D. Neuropharmacologic correlates of deglutition: lessons from fictive swallowing. Dysphagia 1991; 6: 147– 64. 33 Janssens J, De Wever I, Vantrappen G, Hellemans J. Peristalsis in smooth muscle esophagus after transection and bolus deviation. Gastroenterology 1976; 71: 1004–9. 34 Gidda J, Goyal R. Influence of successive vagal stimulations on contractions in esophageal smooth muscle of opossum. J Clin Invest 1983; 71: 1095–103. 35 Diamant N. Physiology of esophageal motor function. Gastroenterol Clin North Am 1989; 18: 179–94. 36 Eslami M, Richards W, Sugarbaker D. Esophageal physiology. Chest Surg Clin N Am 1994; 4: 635–52. 37 Weisbrodt N, Christensen J. Gradients of contractions in the opossum esophagus. Gastroenterology 1972; 62: 1159–66. 38 Sarna S, Daniel E, Waterfall W. Myogenic and neural control systems for esophageal motility. Gastroenterology 1977; 73: 1345–52. 39 Preiksaitis H, Diamant N. Myogenic mechanism for peristalsis in the cat esophagus. Am J Physiol 1999; 277 (Pt 1): G306– 13. 40 Daniel E, Bardakjian B, Huizinga J, Diamant N. Relaxation oscillator and core conductor models are needed for understanding of GI electrical activities. Am J Physiol 1994; 266 (Pt 1): G339–49. 41 Yamato S, Spechler S, Goyal R. Role of nitric oxide in esophageal peristalsis in the opossum. Gastroenterology 1992; 103: 197–204. 42 Kahrilas P, Dodds W, Hogan W. Effect of peristaltic dysfunction on esophageal volume clearance. Gastroenterology 1988; 94: 73–80. 43 Li M, Brasseur J, Dodds W. Analyses of normal and abnormal esophageal transport using computer simulations. Am J Physiol 1994; 266 (Pt 1): G525–43. 44 Ghosh S, Janiak P, Schwizer W, Hebbard G, Brasseur J. Physiology of the esophageal pressure transition zone: separate contraction waves above and below. Am J Physiol Gastrointest Liver Physiol 2006; 290: G568–76. 45 Ghosh S, Janiak P, Fox M, Schwizer W, Hebbard G, Brasseur J. Physiology of the oesophageal transition zone in the presence of chronic bolus retention: studies using concurrent high resolution manometry and digital fluoroscopy. Neurogastroenterol Motil 2008; 20: 750–9. 46 Clouse R, Staiano A. Topography of the esophageal peristaltic pressure wave. Am J Physiol 1991; 261 (Pt 1): G677–84. 47 Pandolfino J, Fox M, Bredenoord A, Kahrilas P. Highresolution manometry in clinical practice: utilizing pressure topography to classify oesophageal motility abnormalities. Neurogastroenterol Motil 2009; 21: 796–806. 48 Pandolfino J, Leslie E, Luger D, Mitchell B, Kwiatek M, Kahrilas P. The contractile deceleration point: an important physiologic landmark on oesophageal pressure topography. Neurogastroenterol Motil 2010; 22: 395–400, e90. 49 Clouse R, Staiano A, Bickston S, Cohn S. Characteristics of the propagating pressure wave in the esophagus. Dig Dis Sci 1996; 41: 2369–76. 50 Kahrilas P, Ghosh S, Pandolfino J. Esophageal motility disorders in terms of pressure topography: the Chicago Classification. J Clin Gastroenterol 2008; 42: 627–35. 51 Nicosia M, Brasseur J, Liu J, Miller L. Local longitudinal muscle shortening of the human esophagus from highfrequency ultrasonography. Am J Physiol Gastrointest Liver Physiol 2001; 281: G1022–33. 52 Sifrim D, Lefebvre R. Role of nitric oxide during swallowinduced esophageal shortening in cats. Dig Dis Sci 2001; 46: 822–30. 53 Zhang X, Geboes K, Depoortere I, Tack J, Janssens J, Sifrim D. Effect of repeated cycles of acute esophagitis and healing on esophageal peristalsis, tone, and length. Am J Physiol Gastrointest Liver Physiol 2005; 288: G1339–46. 54 Pouderoux P, Lin S, Kahrilas P. Timing, propagation, coordination, and effect of esophageal shortening during peristalsis. Gastroenterology 1997; 112: 1147–54. 55 Shi G, Pandolfino J, Joehl R, Brasseur J, Kahrilas P. Distinct patterns of oesophageal shortening during primary peristalsis, secondary peristalsis and transient lower oesophageal sphincter relaxation. Neurogastroenterol Motil 2002; 14: 505–12. © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus 298 Diseases of the Esophagus 56 Mittal R, Liu J, Puckett J et al. Sensory and motor function of the esophagus: lessons from ultrasound imaging. Gastroenterology 2005; 128: 487–97. 57 Mittal R, Padda B, Bhalla V, Bhargava V, Liu J. Synchrony between circular and longitudinal muscle contractions during peristalsis in normal subjects. Am J Physiol Gastrointest Liver Physiol 2006; 290: G431–8. 58 Korsapati H, Bhargava V, Mittal R. Reversal of asynchrony between circular and longitudinal muscle contraction in nutcracker esophagus by atropine. Gastroenterology 2008; 135: 796–802. 59 Dodds W, Stewart E, Hodges D, Zboralske F. Movement of the feline esophagus associated with respiration and peristalsis. An evaluation using tantalum markers. J Clin Invest 1973; 52: 1–13. 60 White R, Zhang Y, Morris G, Paterson W. Esophagitis-related esophageal shortening in opossum is associated with longitudinal muscle hyperresponsiveness. Am J Physiol Gastrointest Liver Physiol 2001; 280: G463–9. 61 Shi G, Pandolfino J, Zhang Q, Hirano I, Joehl R, Kahrilas P. Deglutitive inhibition affects both esophageal peristaltic amplitude and shortening. Am J Physiol Gastrointest Liver Physiol 2003; 284: G575–82. 62 Paterson W. Role of mast cell-derived mediators in acidinduced shortening of the esophagus. Am J Physiol 1998; 274 (Pt 1): G385–8. 63 Paterson W, Miller D, Dilworth N, Assini J, Lourenssen S, Blennerhassett M. Intraluminal acid induces oesophageal shortening via capsaicin-sensitive neurokinin neurons. Gut 2007; 56: 1347–52. 64 Farré R, van Malenstein H, De Vos R et al. Short exposure of oesophageal mucosa to bile acids, both in acidic and weakly acidic conditions, can impair mucosal integrity and provoke dilated intercellular spaces. Gut 2008; 57: 1366–74. 65 Dunne D, Paterson W. Acid-induced esophageal shortening in humans: a cause of hiatus hernia? Can J Gastroenterol 2000; 14: 847–50. 66 Peters L, Maas L, Petty D et al. Spontaneous noncardiac chest pain. Evaluation by 24-hour ambulatory esophageal motility and pH monitoring. Gastroenterology 1988; 94: 878–86. 67 Breumelhof R, Nadorp J, Akkermans L, Smout A. Analysis of 24-hour esophageal pressure and pH data in unselected patients with noncardiac chest pain. Gastroenterology 1990; 99: 1257– 64. 68 Janssens J, Vantrappen G, Ghillebert G. 24-hour recording of esophageal pressure and pH in patients with noncardiac chest pain. Gastroenterology 1986; 90: 1978–84. 69 Lam H, Dekker W, Kan G, Breedijk M, Smout A. Acute noncardiac chest pain in a coronary care unit. Evaluation by 24-hour pressure and pH recording of the esophagus. Gastroenterology 1992; 102: 453–60. 70 Balaban D, Yamamoto Y, Liu J et al. Sustained esophageal contraction: a marker of esophageal chest pain identified by intraluminal ultrasonography. Gastroenterology 1999; 116: 29–37. 71 Mittal R. Motor and sensory function of the esophagus: revelations through ultrasound imaging. J Clin Gastroenterol 2005; 4 (Suppl 2): S42–8. 72 Mayrand S, Diamant N. Measurement of human esophageal tone in vivo. Gastroenterology 1993; 105: 1411–20. 73 Mayrand S, Tremblay L, Diamant N. In vivo measurement of feline esophageal tone. Am J Physiol 1994; 267 (Pt 1): G914– 21. 74 González M, Mearin F, Vasconez C, Armengol J, Malagelada J. Oesophageal tone in patients with achalasia. Gut 1997; 41: 291–6. 75 Zhang X, Tack J, Janssens J, Sifrim D. Neural regulation of tone in the oesophageal body: in vivo barostat assessment of volume–pressure relationships in the feline oesophagus. Neurogastroenterol Motil 2004; 16: 13–21. 76 Sifrim D, Tack J, Lerut T, Janssens J. Transient lower esophageal sphincter relaxations and esophageal body muscular contractile response in reflux esophagitis. Dig Dis Sci 2000; 45: 1293–300. © 2011 Copyright the Authors Journal compilation © 2011, Wiley Periodicals, Inc. and the International Society for Diseases of the Esophagus