Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
ß 2014. Published by The Company of Biologists Ltd | Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
RESEARCH ARTICLE
Regulation of axon growth by the JIP1–AKT axis
ABSTRACT
The polarisation of developing neurons to form axons and dendrites
is required for the establishment of neuronal connections leading to
proper brain function. The protein kinase AKT and the MAP kinase
scaffold protein JNK-interacting protein-1 (JIP1) are important
regulators of axon formation. Here we report that JIP1 and AKT
colocalise in axonal growth cones of cortical neurons and
collaborate to promote axon growth. The loss of AKT protein from
the growth cone results in the degradation of JIP1 by the
proteasome, and the loss of JIP1 promotes a similar fate for AKT.
Reduced protein levels of both JIP1 and AKT in the growth cone can
be induced by glutamate and this coincides with reduced axon
growth, which can be rescued by a stabilized mutant of JIP1 that
rescues AKT protein levels. Taken together, our data reveal a
collaborative relationship between JIP1 and AKT that is required for
axon growth and can be regulated by changes in neuronal activity.
KEY WORDS: Axon, JIP1, AKT, Proteasome
INTRODUCTION
As neurons develop in the brain, they polarise resulting in a single
axon and multiple dendrites. The axon conducts information
while the cell body and dendrites integrate this information by
making multiple synaptic connections. The polarisation of
neurons is therefore essential for the formation of neuronal
networks and the function of the brain (Barnes and Polleux,
2009). A number of factors determining axon formation and
growth have been uncovered, including the localisation of the
centrosome and Golgi, the regulation of gene expression, and
alterations in cytoskeletal dynamics mediated by many signalling
proteins. These include GTPases of the Ras and Rho families, Par
complexes, ubiquitin ligase complexes that target key proteins for
degradation, and the LKB1 and phosphoinositide 3-kinase (PI3K)
pathways (Barnes and Polleux, 2009; Cheng and Poo, 2012; de la
Torre-Ubieta and Bonni, 2011).
The protein kinase AKT is a component of the PI3K pathway.
It is recruited to the PI3K activity-dependent phosphatidylinositol
Faculty of Life Sciences, University of Manchester, Michael Smith Building,
Oxford Road, Manchester, M13 9PT, UK.
*Present address: School of Life Sciences, University of Nottingham, Queen’s
Medical Centre, Nottingham, NG7 2UH, UK. {Present address: Department of
Neurology and Neurosurgery, The Research Institute of the McGill University
Health Center, Montreal General Hospital, Montreal, H3G 1A4, Canada.
§
These authors contributed equally to this work
"
Authors for correspondence ([email protected]; [email protected])
Received 20 June 2013; Accepted 14 October 2013
230
(3,4,5)-trisphosphate [PtdIns(3,4,5)P3] in the membrane where it
is activated by 3-phosphoinositide-dependent protein kinase 1
(PDK1) phosphorylation at Thr308 (Bhaskar and Hay, 2007). A
second phosphorylation event at Ser473, which can be mediated
by the mTOR-containing complex TORC2, is required for full
AKT activity and may also regulate substrate selection (Bhaskar
and Hay, 2007). In developing neurons, AKT preferentially
localises to newly forming axons rather than dendrites (Shi et al.,
2003) and it has been proposed that this differential localisation
of AKT is maintained by its proteasomal degradation in the
dendrites (Yan et al., 2006). Indeed, inhibition of the proteasome
leads to a loss of polarisation and multiple AKT-positive neurites
(Yan et al., 2006). The presence of activated AKT is crucial
for axon formation through its targeting of effector proteins
that regulate the cytoskeleton and promote axon extension,
in particular through GSK3b–CRMP2 (Jiang et al., 2005;
Yoshimura et al., 2005).
It has previously been reported that AKT associates with the
JNK MAP kinase scaffold protein JIP1 (Kim et al., 2002; Song
and Lee, 2005a). JIP1 can also bind to multiple components of the
JNK pathway and facilitate JNK activation (Whitmarsh et al.,
1998). In response to excitotoxicity or oxygen–glucose deprivation,
JIP1 mediates JNK-dependent apoptosis of hippocampal neurons
(Whitmarsh et al., 2001). It has been shown that AKT dissociates
from JIP1 in response to excitotoxic or metabolic stress thereby
allowing JIP1 to recruit JNK and enhance its activation (Kim et al.,
2002; Song and Lee, 2005a; Song and Lee, 2005b). Work in our
laboratory has demonstrated that JIP1, in addition to its role in stress
responses, is required for the polarisation of cultured cortical
neurons and for axon growth (Dajas-Bailador et al., 2008). JIP1
specifically localises to the growth cone of the developing axon
through an interaction with the motor protein kinesin-1 (Verhey
et al., 2001; Dajas-Bailador et al., 2008) and, in association
with kinesin-1, it might regulate axonal vesicle transport through
interactions with transmembrane proteins, including APP (Muresan
and Muresan, 2005; Horiuchi et al., 2005; Whitmarsh, 2006; Fu and
Holzbaur, 2013).
In this study we explore the relationship between JIP1 and
AKT during axon outgrowth in cultured cortical neurons. We
demonstrate that JIP1 and AKT colocalise in the growth cone of
the developing axon and maintain each other’s stability. The loss
of either protein promotes the degradation of the other by the
proteasome and a cessation of axon growth. Furthermore, the
levels of JIP1 and AKT in the axon growth cone are regulated by
glutamate receptor stimulation.
RESULTS
JIP1 and AKT stabilize each other in axonal growth cones
JIP1 and AKT localise to axonal growth cones (Shi et al., 2003;
Dajas-Bailador et al., 2008) and have been reported to form
complexes (Kim et al., 2002; Song and Lee, 2005a); however, it
remains unclear whether these proteins can collaborate to regulate
axon formation and function. To investigate this we used cultured
Journal of Cell Science
Federico Dajas-Bailador*,§,", Ioannis Bantounas§, Emma V. Jones{ and Alan J. Whitmarsh"
primary cortical neurons from mouse embryos. In vitro, these
neurons undergo a series of developmental stages whereby they
extend a number of neurites (stages 1 and 2) prior to polarisation,
at which point one of the neurites forms an axon and the others
develop into dendrites (stage 3) (Dotti et al., 1988). At stage 2,
AKT is reported to be present at the tips of all the neurites (Yan
et al., 2006) whereas JIP1 has already localised to the future axon
(Dajas-Bailador et al., 2008). At stage 3, JIP1 and AKT colocalise
in the growth cones of newly formed axons along with the
axonal-specific microtubule binding protein Tau (Fig. 1A;
supplementary material Fig. S1). To determine whether JIP1
plays a scaffold role in maintaining AKT at the axon tip, shRNAmediated knockdown of JIP1 was performed. A substantial
reduction in JIP1 protein was observed by immunoblotting
(Fig. 1B). JIP1 knockdown had minimal effect on the total AKT
protein level in the neurons as determined by immunoblotting
(Fig. 1B); however, examination of immunostained axonal
growth cones revealed a significant loss of AKT (Fig. 1C,D).
There were no obvious changes in the AKT levels in the cell
bodies and axon shafts (Fig. 1C), thus explaining why we do not
observe a substantial reduction in total AKT protein in the neurons
(Fig. 1B). To confirm that the reduced AKT protein level in the
growth cone was due to the loss of JIP1 and not an off-target effect
of the shRNA, we re-introduced recombinant JIP1 back into the
neurons and rescued the AKT staining (Fig. 1C,D). We next
determined whether the localisation of JIP1 to the axon tip was
required to maintain the AKT level there. JIP1 localises to the
growth cone through its interaction with the kinesin-1 motor
protein (Verhey et al., 2001). The kinesin-1-binding site on JIP1
resides at its C-terminus and mutation of Tyr705 to Ala within this
region prevents binding and results in the loss of JIP1 localisation
to the growth cone (Verhey et al., 2001; Dajas-Bailador et al.,
2008). Unlike wild-type JIP1, the expression of the JIP1 Y705A
mutant did not rescue the loss of AKT protein in the growth cone
following knockdown of endogenous JIP1 (Fig. 1C,D). These data
suggest that JIP1 is required to maintain AKT protein levels in the
axon growth cone. To further understand the relationship between
JIP1 and AKT, we investigated whether the loss of AKT affected
JIP1 localisation or stability. To do this we knocked down AKT
protein levels with shRNAs directed against the three AKT
isoforms (AKT1, AKT2 and AKT3). This led to a decrease in total
JIP1 protein (Fig. 1E) and in JIP1 immunostaining of the axon tip
(Fig. 1F,G). The loss of axonal JIP1 could be rescued by reintroducing recombinant human AKT1 into the neurons
(Fig. 1F,G). Taken together, these data suggest that the AKT and
JIP1 support each other’s presence in axonal growth cones.
AKT and JIP1 protect each other from degradation by the proteasome
One possibility was that AKT and JIP1 protect each other from
proteasome-mediated degradation in the axon tip. To address this,
neurons were incubated with the proteasome inhibitor MG132.
This rescued the loss of JIP1 in the axon tips of AKT knockdown
neurons (Fig. 2A) and also the reduction of AKT in the axon tips
of JIP1 knockdown neurons (Fig. 2B). This suggests that the
colocalisation of JIP1 and AKT to the axon growth cone prevents
their proteasome-dependent degradation. As the AKT pathway
mediates many cellular processes, including transcriptional
responses, we determined whether the loss of JIP1 protein that
occurs upon AKT knockdown might also reflect reduced
transcription of the Jip1 gene. However, no changes in transcript
levels were observed (Fig. 2C), thus providing additional evidence
that the major mechanism of regulation is through protein stability.
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
The role of proteasomal regulation of AKT and JIP1 in axon formation
It is known that the inhibition of the proteasome prevents
neuronal polarisation and results in multiple neurites with high
levels of AKT at the tips (Yan et al., 2006). We confirmed the
presence of AKT at many neurite tips following inhibition of the
proteasome (Fig. 3A,C) and also demonstrated that JIP1 was
present in many neurite tips (Fig. 3B,C). Indeed, JIP1 and AKT
were colocalised in the tips (Fig. 3C). These data demonstrate
that inhibition of the proteasome leads to loss of the asymmetric
distribution of AKT and JIP1 to a single axon. This correlated
with a significant reduction in neuronal polarisation (Fig. 3D)
with the majority of the neurites positive for the axonal marker
protein Tau (Fig. 3E,F). These data suggest a strong correlation
between the presence of JIP1 and AKT in a neurite and it
displaying axon-like properties, and support a model whereby the
specific localisation of both JIP1 and AKT proteins to the axon
is an important mechanism in the regulation of neuronal
polarisation and axon growth.
Neuronal activity regulates JIP1 and AKT stability
In addition to the role of JIP1 and AKT in axon formation, we
were interested in determining whether the JIP1–AKT axis is
responsive to neuronal activity. Glutamate is the major excitatory
neurotransmitter in the central nervous system and the principal
neurotransmitter of cortical efferent systems (Fonnum, 1984).
Previous studies have demonstrated that JIP1 can be regulated
following stimulation of the N-methyl-D-aspartate (NMDA) class
of glutamate receptors (Kim et al., 2002; Kennedy et al., 2007;
Centeno et al., 2007). We exposed developing cultured neurons to
glutamate and this led to a reduction in JIP1 protein levels that
could be rescued by the NMDA receptor antagonist MK-801, but
not by the AMPA/kainate antagonist CNQX (Fig. 4A). This
indicated that glutamate was acting through NMDA receptors to
decrease JIP1 protein levels. We next determined whether
glutamate was acting locally on NMDA receptors in the axon
or through receptors at the cell body or in dendrites, thereby
regulating JIP1 levels from a distance. To distinguish these
possibilities, we plated neurons in microfluidic chambers
that allow the compartmentalization and fluidic isolation of the
axons away from the cell bodies and dendrites (Fig. 4B).
Application of glutamate to the axonal compartment was
sufficient to reduce JIP1 immunostaining in the axon tip,
indicating that glutamate could act directly on the axon to
regulate JIP1 stability (Fig. 4C,D). To determine whether JIP1
stability was sensitive to the endogenous activation of glutamate
receptor neurotransmission, we exposed neurons to potassium
chloride (KCl). This causes cortical neurons to undergo
membrane depolarisation and subsequent activation of voltagesensitive channels leading to glutamate release. KCl caused a
reduction in overall JIP1 protein levels and this was rescued by
pre-incubation of neurons with an inhibitor of the L-type voltagedependent calcium channel (nifedipine) or the NMDA receptor
antagonist MK801 (Fig. 4E). Furthermore, KCl caused a
reduction of both JIP1 and AKT proteins in axon growth cones
(Fig. 4F,G). The activation of NMDA receptors by glutamate
increases calcium ion influx; therefore we investigated whether
this contributed to the loss of JIP1 stability. EGTA was used to
chelate extracellular calcium ions and prevent the NMDAdependent calcium ion influx. This rescued the loss of JIP1
protein that occurred following glutamate treatment (Fig. 4H) and
confirmed that intracellular calcium signalling is required to
destabilize JIP1.
231
Journal of Cell Science
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
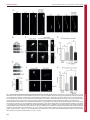
Fig. 1. JIP1 and AKT are required to maintain their respective protein levels in axonal growth cones. (A) Cortical neurons were fixed at 3 days in vitro (d.i.v.) and
stained with anti-JIP1, anti-AKT and anti-Tau antibodies. Left panels: JIP1, AKT and Tau localise to the tip of the emerging axon (indicated by a white arrow). Scale bar:
40 mm. Right panels: magnification of boxed growth cone demonstrating co-localisation of JIP1, AKT and Tau in the axon tip. Scale bar: 10 mm. (B) Immunoblot of JIP1
and AKT protein levels in lysates of neurons treated with lentivirus expressing JIP1 shRNA or control shRNA. Tubulin was used to control for equal protein loading.
(C) Immunostaining of AKT in axon growth cones and cell bodies of neurons electroporated with plasmids expressing JIP1 shRNA or control shRNA. AKT
immunostaining was also assessed in neurons transfected with pCDNA3-T7-JIP1 or pCDNA3-T7-JIP1(Y705A). The latter plasmid expresses a JIP1 mutant that does
not bind kinesin-1. Scale bars: 10 mm. (D) Quantification of AKT immunostaining in axon tips after electroporation of neurons with plasmids expressing control shRNA
or JIP1 shRNA. Neurons expressing JIP1 shRNA were also transfected with pCDNA3-T7-JIP1 or pCDNA3-T7-JIP1(Y705A). (E) Neurons were subjected to lentiviralmediated shRNA knockdown of AKT1/2/3 and the protein level of JIP1 was measured by immunoblotting. A representative immunoblot is shown as is the quantification
of blots from three experiments. (F) Immunostaining of JIP1 in axon growth cones after shRNA knockdown of AKT1/2/3. Neurons were also transfected with either
pCMV5 or pCMV5-HA-AKT1. Scale bar: 10 mm. (G) Quantification of JIP1 fluorescence in axon tips after lentiviral-mediated shRNA knockdown of AKT1/2/3 with or
without transfection of pCMV5 or pCMV5-HA-AKT1 (*P,0.05, **P,0.01).
232
Journal of Cell Science
RESEARCH ARTICLE
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
Glutamate stimulation causes the loss of JIP1 and AKT proteins in the
growth cone through the JNK pathway and the proteasome
As we had evidence that the proteasome regulates JIP1 and AKT
stability during axon development (Figs 2, 3), we determined
whether the loss of JIP1 and AKT in the growth cone in response
to glutamate stimulation was also being mediated by the
proteasome. The pre-incubation of glutamate-stimulated
neurons with the proteasome inhibitor MG132 rescued both
JIP1 and AKT protein levels in axon tips, indicating that
activation of glutamate neurotransmission can regulate the
stability of these proteins (Fig. 5A–C). It has previously been
reported that activation of kainate glutamate receptors in an
excitotoxic model can cause the dissociation of the JIP1–AKT
complex (Kim et al., 2002). We did not observe neuronal loss in
our experiments following stimulation of neurons with glutamate
(100 mM, 3 hours; supplementary material Fig. S2) suggesting
that the loss of JIP1 and AKT in axons is unlikely to be due to
excitotoxicity. However, it raised the possibility that glutamate
promotes the dissociation of the JIP1–AKT complex leading to
their instability. Co-immunoprecipitation experiments demonstrated
that glutamate stimulation did decrease JIP1 binding to AKT
(Fig. 5D). This occurred within 30 minutes of glutamate exposure,
prior to there being a significant decrease in JIP1 protein levels,
thus ruling out the possibility that the reduced binding was simply
due to a reduction in JIP1 protein in the neurons (Fig. 5D). Studies
in non-neuronal cells have shown that JNK phosphorylation of JIP1
controls its ability to bind to AKT (Song and Lee, 2005a; Song and
Lee, 2005b), whereas in cortical neurons treated with NMDA, it is
reported that inhibition of JNK activity can stabilize JIP1 (Centeno
et al., 2007). This suggested that glutamate stimulation induces
JNK phosphorylation of JIP1 thus causing the dissociation of the
JIP1–AKT complex and subsequent destabilization of the proteins
in the growth cone. If this is the case, inhibition of the JNK pathway
should rescue the glutamate-induced loss of JIP1 and AKT. Indeed,
when JNK activity was blocked by two different inhibitors, the
glutamate-induced degradation of JIP1 and AKT was prevented
(Fig. 5E–G; supplementary material Fig. S3). The major JNK
phosphorylation site on JIP1 that is proposed to regulate its binding
to AKT is Thr103 (Song and Lee, 2005a; Nihalani et al., 2003), so
we tested whether mutation of this site to a non-phosphorylatable
Ala residue prevented glutamate-induced JIP1 degradation.
Glutamate stimulation did not cause a reduction in JIP1 T103A
protein in the axon tips in contrast to the significant decrease in
wild-type JIP1 (Fig. 5H). Furthermore, the JIP1 T103A mutant
maintained AKT levels in the growth cone (Fig. 5I). Taken
together, these data support a model whereby glutamate regulates
the stability of JIP1 and AKT in growth cones by controlling their
association through the activation of the JNK pathway.
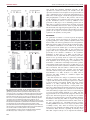
Fig. 2. AKT and JIP1 protect each other from degradation by the
proteasome. Cortical neurons at 4.5 d.i.v. were treated with the proteasome
inhibitor MG132 (0.2 mM, 16 hours) following lentiviral shRNA mediated
knockdown of AKT1/2/3 or JIP1. (A) Quantification of JIP1 fluorescence
intensity in axon tips after AKT1/2/3 knockdown and MG132 treatment.
(B) Quantification of AKT fluorescence intensity in axon tips after JIP1
knockdown and MG132 treatment. (C) qPCR of JIP1 and AKT1 mRNA in
neurons expressing shRNA against AKT1/2/3. Values are means 6 s.e.m. of
three experiments (*P,0.05).
The knockdown of either AKT (Fig. 6A) or JIP1 (Fig. 6B) (DajasBailador et al., 2008) reduces axonal growth in cultured neurons.
Similarly, glutamate stimulation can slow down axon growth
(Fig. 6B). However, there is no additive effect on axon growth of
glutamate treatment plus JIP1 knockdown (Fig. 6B), which would
be consistent with glutamate causing a slowing of axon growth by
reducing JIP1 stability. The glutamate-induced decrease in axon
growth could be rescued by inhibition of JNK activity (Fig. 6C),
which protects JIP1 from degradation (Fig. 5E,F). Furthermore,
when we expressed the stabilized JIP1 mutant (T103A) in neurons
we found that, unlike wild-type JIP1, it could rescue the decrease in
233
Journal of Cell Science
Stabilization of JIP1 reverses the glutamate-induced block in
axon growth
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
axon growth after glutamate stimulation (Fig. 6D). In this
experiment we transfected relatively low amounts of the JIP1
expression constructs because at high levels of expression JIP1 can
cause a substantial increase in axon length (Dajas-Bailador et al.,
2008). These results support a model whereby glutamate-induced
JNK phosphorylation of T103 in JIP1 promotes slowed axon
growth. It might be expected that the reduced axon growth caused
by JIP1 knockdown could be rescued by inhibition of the
proteasome if JIP1 plays a role in stabilizing proteins (such as
AKT) that are required for growth. Indeed, treatment of neuronal
cultures with MG132 did rescue the defect in growth following
JIP1 knockdown, thus supporting this hypothesis (Fig. 6E). Taken
together, these results indicate that the control of JIP1 levels in the
growth cone in response to neuronal activity is required for the
regulation of the dynamics of axon growth.
Fig. 3. Proteasomal regulation of JIP1 and AKT proteins in axon
formation. Neurons were immunostained for either AKT (A) or JIP1
(B) following treatment with the proteasome inhibitor MG132 (0.2 mM,
16 hours) at 2 d.i.v. The number of neurite tips positive for these proteins
was quantified. Accumulated data from three independent experiments,
each with more than 50 neurons, is shown. (C) Co-staining for JIP1 and
AKT in 2 d.i.v. neurons following MG132 treatment. Neurite tips positive
for AKT and JIP1 are indicated by white arrows. Scale bar: 20 mm.
(D) Cortical neurons (2 d.i.v.) were treated with MG132 (0.2 mM, 16 hours)
and assessed for polarisation. A polarised axon was defined as a neurite that
was longer than 80 mm and at least three times the length of other
processes. Data are from three independent experiments, each with more
than 50 neurons. (E) The MG132-treated neurons were immunostained for
Tau and the number of neurites positive for Tau quantified.
(F) Immunostaining of neurons for Tau and tubulin following treatment
with MG132 (0.2 mM, 16 hours) at 2 d.i.v. Scale bar: 20 mm.
234
The polarisation of neurons is a crucial step in the development
of the nervous system and involves the integration of many
signalling pathways to control cytoskeletal dynamics (Barnes and
Polleux, 2009). One of the key pathways involves the lipid kinase
PI3K that promotes AKT activation (Read and Gorman, 2009).
The asymmetric distribution of active AKT in the developing
axon tip, but not in the dendrite tips, is essential for polarisation
to occur (Read and Gorman, 2009). It has previously been
proposed that selective degradation of AKT by the proteasome
specifically in the dendrites contributes to the asymmetric
distribution of AKT (Yan et al., 2006). In this study we
demonstrate that the JNK MAP kinase scaffold protein JIP1,
which we have previously shown is required for polarisation
and axon growth (Dajas-Bailador et al., 2008), can protect AKT
from degradation in the axon tip (Figs 1, 2). Furthermore, we
demonstrate a collaborative relationship between these proteins,
as the stability of JIP1 requires the presence of AKT (Figs 1, 2). It
is currently unclear how the asymmetric distribution of AKT and
JIP1 is maintained. Our data would be consistent with a model
whereby JIP1 is localised to the emerging axon tip through
association with the motor protein kinesin-1 (Verhey et al., 2001;
Dajas-Bailador et al., 2008; Jacobson et al., 2006), where it
associates with AKT, resulting in a stabilized complex that
promotes axon growth.
How JIP1 facilitates the changes in cytoskeletal architecture
required to promote axon growth is also unclear. JIP1 can
cooperate with fasciculation and elongation protein zeta-1 (FEZ1)
to activate kinesin-1 microtubule binding and motor activity
(Blasius et al., 2007), indicating that it might enhance the
transport of other proteins specifically to axonal growth cones
where they could maintain polarisation and promote axon growth.
This is supported by studies in Drosophila where loss of function
of the JIP1 orthologue Aplip1 leads to defects in axonal transport
of vesicles and mitochondria (Horiuchi et al., 2005). Our data
suggest that JIP1 is not essential for the transport of AKT to axon
tips because AKT is still present in the growth cones of neurons
that have been incubated with a proteasome inhibitor following
knockdown of JIP1 (Fig. 2B). AKT can regulate many proteins
that influence axon growth (Read and Gorman, 2009). Some of
the best characterized are microtubule-associated proteins,
including CRMP2 and Tau that can be targeted by AKT control
of GSK3b activity. Also, AKT can directly target proteins
involved in actin organization including girdin and ezrin (Read
and Gorman, 2009). Therefore, regulating the stability of the
JIP1–AKT complex is likely to be an important mechanism in
Journal of Cell Science
DISCUSSION
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
rapid modulation of the cytoskeleton and axon growth dynamics
in response to developmental cues or neuronal activity.
Furthermore, the manipulation of the JIP1–AKT axis could
represent a potential route for promoting nerve regeneration.
There is already evidence that JIP1 promotes regeneration of
adult mouse dorsal root ganglion neurons (Barnat et al., 2010)
and a number of studies have proposed a role for AKT-mediated
signalling in axonal regeneration (Namikawa et al., 2000; Kim
et al., 2011; Song et al., 2012).
It is becoming increasingly apparent that the regulation of
protein levels constitutes an important mechanism in the
modulation of neuronal function (Gallo and Letourneau, 2002;
DiAntonio and Hicke, 2004). The demonstrated role of glutamate
stimulation through NMDA receptors in promoting instability of
AKT and JIP1 in axons may be particularly relevant since the
establishment of neuronal connections in the developing brain is
not only guided by specific signalling molecules and growthpromoting factors, but also by specific patterns of neuronal
activity. It has been proposed that NMDA receptors can have a
role in experience-dependent circuit refinement through
regulating axonal structural plasticity (Gogolla et al., 2007). For
example, conditional cortical knockout of the NMDA receptor
component NR1 in mice leads to thalamocortical axons
developing more extensive arborizations (Lee et al., 2005), and
in the tadpole tectum, selective branch elimination is prevented
by NMDA receptor blockade (Ruthazer et al., 2003). It is possible
that the proteasomal regulation of JIP1 and AKT may contribute
to the plasticity of presynaptic boutons and axon branches in the
developing and adult brain.
The mechanism by which NMDA receptor signalling mediates
JIP1 and AKT degradation involves the influx of calcium ions
(Fig. 4H) and activation of the JNK pathway (Fig. 5E–G). It will
clearly be of interest in the future to also identify the ubiquitin
ligase complexes that target JIP1 and AKT for degradation. The
involvement of calcium and JNK in regulating JIP1 stability is
supported by other studies that demonstrate: (1) calcium added to
cell lysates in vitro can promote JIP1 ubiquitylation (AllamanPillet et al., 2003), and (2) NMDA treatment of neurons causes a
235
Journal of Cell Science
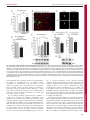
Fig. 4. Neuronal activity regulates JIP1 and AKT stability. (A) Neurons at 10 d.i.v. were treated with glutamate (100 mM, 3 hours) together with either CNQX
(20 mM) or MK801 (5 mM), and JIP1 protein level assessed by immunoblotting. JIP1 protein levels from three experiments were quantified by densitometry.
(B) Neurons developed in microfluidic chambers. Cell bodies and nuclei were stained with anti-Sox1 antibody and axons were stained with acetylated tubulin
antibody at 5 d.i.v. (C) Glutamate (100 mM, 3 hours) was added to the axonal compartment. Images of glutamate-induced loss of JIP1 staining are shown for two
axons. (D) The intensity of JIP1 immunostaining in axon tips was quantified. (E) Neurons at 10 d.i.v. were treated with KCl (20 mM) and either nifedipine (Nif;
5 mM), MK-801 (5 mM) or CNQX (20 mM) and lysates immunoblotted for JIP1 and actin protein levels. JIP1 protein levels from three experiments were quantified
by densitometry. (F,G) JIP1 and AKT immunostaining in axon tips were quantified in the absence and presence of KCl (20 mM, 3 hours). (H) Neurons were
treated with glutamate (100 mM, 3 hours) in the presence or absence of the calcium ion chelator EGTA (5 mM). JIP1 and actin protein levels were monitored by
immunoblotting (*P,0.05).
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
JNK-dependent decrease in JIP1 (Centeno et al., 2007). These
observations, combined with our own findings, suggest a model
whereby glutamate promotes JNK-mediated phosphorylation of
JIP1 leading to its dissociation from AKT and subsequent
degradation of both proteins in the axon tip. This loss of JIP1
and AKT mediates glutamate-induced cessation of axon growth.
Crucially, AKT protein levels in the axon tip and axon growth can
both be restored by the expression of a mutant JIP1 with a key JNK
phosphorylation site mutated (Fig. 5I; Fig. 6D), thus providing
236
strong evidence that the proposed mechanism is important for
regulating axonal growth. It has previously been observed that
inhibition of the JNK pathway prior to polarisation can suppress
axonogenesis (Oliva et al., 2006), whereas our results suggest that
JNK may not be essential for axon growth following polarisation
(Fig. 6C), but can play a role in modulating the response of the
growth cone to glutamate stimulation.
Taken together, our study demonstrates an important
collaboration between JIP1 and AKT to promote axon formation
Journal of Cell Science
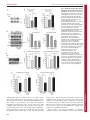
Fig. 5. Glutamate regulates JIP1 and AKT
stability through the JNK pathway and the
proteasome. Neurons at 8 d.i.v. were treated
with glutamate (Glut; 100 mM, 3 hours) with or
without MG132 (10 mM). (A–C) JIP1 protein
levels were monitored by immunoblotting
(A) and the fluorescence intensity of JIP1 and
AKT immunostaining in the axon tips was
quantified (B,C). (D) AKT was
immunoprecipitated from lysates of neurons
treated with glutamate (100 mM) for the
indicated times. The AKT immunoprecipitates
(IP) were analysed for the presence of JIP1.
The protein level of JIP1 in the AKT
immunoprecipitations and in the lysates was
quantified by densitometry from four
experiments. (E) Neurons were treated with
glutamate (100 mM, 2 hours) after pretreatment with the JNK inhibitor XVI (JNK-i;
2 mM) and JIP1 levels analysed by
immunoblotting. (F,G) The fluorescence
intensities of JIP1 and AKT immunostaining in
the axon tips of neurons treated with
glutamate (100 mM, 3 hours) after pretreatment with JNK inhibitor XVI (JNK-i; 2 mM)
were quantified. (H) Neurons were transfected
with constructs expressing either T7-tagged
JIP1 or JIP1 T103A and treated with
glutamate (100 mM, 3 hours). The
fluorescence intensities of JIP1
immunostaining in the axon tips were
quantified. For each construct the JIP1
fluorescence in untreated neurons (Con) was
set to 100%. (I) The fluorescence intensity of
AKT immunostaining in the axon tips was
quantified for neurons transfected with
constructs expressing either T7-tagged JIP1
or JIP1 T103A and treated with glutamate
(*P,0.05, **P,0.01, N.S, not significant).
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
Fig. 6. Stabilized JIP1 reverses the glutamate-induced block in axon growth. (A) Neurons at 8 d.i.v. were subjected to siRNA-mediated knockdown of AKT
isoforms, and axon length measured. (B) Neurons subjected to siRNA-mediated knockdown of JIP1 were treated with glutamate (100 mM, 6 hours) or left
untreated, and axon length measured. (C) Neurons incubated with or without JNK inhibitor XVI (JNK-i; 2 mM) were treated with glutamate (100 mM, 6 hours)
or left untreated, and axon length measured. (D) Neurons were transfected with pEGFPC2 and either a plasmid expressing T7-tagged JIP1 or JIP1 (T103A),
and treated with glutamate or left untreated. The length of axons in GFP-positive neurons was measured. (E) Neurons were subjected to lentiviral-mediated
knockdown of JIP1 (shJIP1) and were treated with MG132 (0.2 mM, 16 hours) or left untreated, and axon length was measured (*P,0.05, **P,0.01, ***P,0.001).
MATERIALS AND METHODS
Neuronal cell culture
Primary embryonic cortical neurons were obtained and cultured as described
before (Dajas-Bailador et al., 2008). Briefly, cortices of E17 embryos of C57
mice were dissected in HBSS (Life Technologies Ltd, Paisley, UK) and
trypsinised. Following trituration, dissociated neurons were plated in
Neurobasal medium supplemented with 2% B27 and 16 GlutaMAX (all
from Life Technologies Ltd), on poly-D-lysine (Sigma, Dorset, UK)-coated
plates. Cell density was 56105 neurons per well in six-well plates for western
blotting and immunoprecipitation experiments and 26104 or 46104 neurons
per well in 24-well plates, on glass coverslips, for immunocytochemistry
experiments. JNK inhibitor II (SP600125) (Bennett et al., 2001) and JNK
inhibitor XVI (JNK-IN-8) (Zhang et al., 2012) (both from Merck Millipore,
Watford, UK) were used at a final concentration of 10 mM and 2 mM,
respectively. MG132 (Merck Millipore) was added at a concentration of
0.2 mM, overnight, except in glutamate stimulation experiments, where it
was used at 10 mM for 30 minutes before the addition of glutamate.
Glutamate was used at a final concentration of 100 mM.
Compartmentalized culture in microfluidic chambers
Primary cortical neurons (E17) were cultured for 5 days in microfluidic
devices with 150 mm long microgrooves (Xona Microfluidics LLC,
Temecula, CA, USA), as described previously (Dajas-Bailador et al.,
2012). The chambers allow for fluidic isolation of the axonal from the
cell body (somal) compartment. Glutamate was applied only to the
axonal side of the microfluidic device.
Plasmid construction
For shRNA vector construction, inserts were designed as complementary
oligonucleotides coding for the appropriate shRNA, producing blunt ends
and XhoI ends upon annealing and were ligated into HpaI–XhoI-digested
pLL3.7 plasmid (Rubinson et al., 2003). Oligonucleotide sequences
were: shJIP1: 59-TGAGCAAACCCATCGGGCTATTCAAGAGATAGCCCGATGGGTTTGCTCTTTTTTC-39, 59-CGAGAAAAAAGAGCAAACCCATCGGGCTATCTCTTGAATAGCCCGATGGGTTTGCTCA-39;
shAkt1/2: 59-TGTGCAGTGGACCACAGTCATTTCAAGAGAATGACTGTGGTCCACTGCATTTTTTC-39, 59-TCGAGAAAAAATGCAGTGGACCACAGTCATTCTCTTGAAATGACTGTGGTCCACTGCCA-39;
shAkt3: 59-TGGCAAGATGTATATGACAATTCAAGAGATTGTCATATACATCTTGCCTTTTTTC-39, 59-TCGAGAAAAAAGGCAAGATGTATATGACAATCTCTTGAATTGTCATATACATCTTGCCA-39;
shControl: 59-TGGCACATTAGAAGCCATACATTTCAAGAGAATGTATGGCTTCTAATGTGCCTTTTTTC-39, 59-TCGAGAAAAAAGGCACATTAGAAGCCATACATTCTCTTGAAATGTATGGCTTCTAATGTGCCA-39. The pRRL.sin.cppt.CMV-IRES-EGFP-WPRE (empty) vector
was created by digesting pIRES2-EGFP (Clontech) with NotI–Blunt–NheI
and ligating the IRES-EGFP fragment into SalI–Blunt–XbaI-digested
pRRL.sin.cppt.CMV-EGFP-WPRE (provided by J. Uney, University of
Bristol). The expression vectors pcDNA3-T7-JIP1 and pcDNA3-T7JIP1(Y705A) were described previously (Whitmarsh et al., 1998; DajasBailador et al., 2008). To obtain pcDNA3-T7-JIP1(T103A), site-directed
mutagenesis was performed using the QuikChange kit (Agilent
Technologies, Lakeside, UK) and the primers: 59-GCGGCAGGTGACGCTCCGGGCGCCGAG-39 and 59-CTCGGCGCCCGGAGCGTCACCTGCCGC-39.
Lentiviral vector production and concentration
HEK293T cells were co-transfected with the appropriate shuttle vector
and the helper plasmids pMDLg/pRRE, pMD2.G and pRSV-Rev by
calcium phosphate precipitation. Cell medium from transfected cells was
collected over 2 days and centrifuged at 6000 g overnight, followed by
re-suspension of the pellet in phosphate-buffered saline (PBS) and
ultracentrifugation at 50,000 g in a SW40-Ti rotor (Beckman Coulter
Ltd, High Wycombe, UK) for 1.5 hours. The final viral pellets were
re-suspended in PBS at 1:2000 of the original media volumes. Viral
titres were calculated by fluorescence-activated cell sorting (detecting
EGFP-expressing cells), after HEK293T cell transduction.
237
Journal of Cell Science
and provides evidence that the JIP1–AKT axis responds to
neuronal activity in order to modulate axonal growth dynamics.
RESEARCH ARTICLE
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
Plasmid transfection and lentiviral transduction of cells
Immunocytochemistry
Plasmids and JIP1 siRNA (sc-35723, Santa Cruz, Heidelberg, Germany)
or AKT siRNA (sc-43610, Santa Cruz) were either electroporated into
neurons using the Mouse Neuron Nucleofector Kit (Lonza Biosciences,
Wokingham, UK) or transfected using Lipofectamine 2000 (Life
Technologies Ltd) according to the manufacturers’ instructions.
Typically, 1–1.56106 neurons were electroporated with 5–10 mg of
plasmid, before plating. Neurons were transduced with lentivirus on the
day of plating at a multiplicity of infection (MOI) of 10 for each vector. In
AKT knockdown experiments, the LV-shCTRL vector was used at MOI of
20 to control for the total viral load resulting from using both LV-shAkt1/2
and LV-shAkt3 at MOI of 10 each in the knockdown condition.
Neurons were fixed for 15 minutes in 4% paraformaldehyde and
permeabilised in 0.2% Triton X-100 in PBS for 5 minutes, followed by
blocking in 3% BSA-PBS for 1 hour. Primary antibodies were used at
1:100 (AKT) or 1:200 (JIP1) dilution (in 3% BSA/PBS), overnight at
4 ˚C, followed by three washes with 0.1% Triton-X100 in PBS.
Appropriate secondary Alexa Fluor antibodies were used at 1:250–
1:500 dilution in 3%BSA-PBS for 1 hour at room temperature, followed
by three washes with 0.1% Triton-X100/PBS before the cells were
mounted on glass slides using Prolong Gold Antifade Reagent with
DAPI (Life Technologies Ltd). In some experiments, to visualise the
neurons, coverslips were incubated with FITC-conjugated anti-b-tubulin
antibody (1:500 dilution) for 1 hour at room temperature, after the
secondary antibody and before mounting. Images were collected on an
Olympus BX51 upright microscope using a 206/0.50 NA UPlanFLN or
a 606/0.65–1.25 NA UPlanFLN objective and captured using a
Coolsnap HQ camera (Photometrics, Tucson, AZ, USA) through
MetaVue Software (Molecular Devices; Sunnyvale, CA, USA).
Quantification of mean fluorescence intensity of JIP1 and AKT at
axon tips was performed using ImageJ (http://rsb.info.nih.gov/ij) on
images obtained using the 1006 objective. Images of fields of neurons
were taken with the 606 objective. More than 50 axon tips were imaged
per condition in each of at least three independent experiments. For the
measurement of axons, an axon was defined as a neurite that was longer
than 80 mm and at least three times the length of other processes. Axon
length was measured using ImageJ and data expressed as the means 6
s.e.m. The probability distribution of the data set was analysed before
further statistical analysis (,100 axons measured for each condition
from n53 or 4). Statistical analysis of test data compared with the
respective control was performed using Prism (V 5.0c) (*P,0.05,
**P,0.01, ***P,0.01). Multiple group comparisons were carried out
using ANOVA (post-hoc Tukey’s test) or Kruskal–Wallis one-way
analysis of variance (post-hoc Dunn’s test) for non-parametric data.
Pairwise comparisons were carried out using Student’s t-test or Mann–
Whitney test for non-parametric analysis.
Total RNA was extracted using the RNeasy Kit (Qiagen, Manchester,
UK) according to the manufacturer’s instructions and RNA samples were
treated with DNase (DNAfree Kit; Life Technologies Ltd). Quantitative
PCR was performed using Multiscribe Reverse Transcriptase and
SYBR-Green PCR Reaction Mix (Life Technologies Ltd) in a singlestep reaction according to the manufacturer’s instructions, and run
on a BioRad C1000 Thermal Cycler. The primers used were: Jip1,
59-GCTATATTCAGGTTTGTGCCTC-39 and 59-GTCGCCTCCCAATTTCAG-39; Akt1, 59-GTATGAGAAGAAGCTGAGCCC-39 and 59GATCATCTGAGCTGTGAACTCC-39. Typically, 20 ng of total RNA
was used per reaction.
Antibodies
Primary antibodies used in this study were: mouse anti-JIP1 (sc-25267;
Santa Cruz), rabbit anti-AKT and anti-AKT(p-S473) (9272 and 4060;
Cell Signaling) that recognize all AKT isoforms, anti-AKT1 (sc-1618,
Santa Cruz), mouse anti-Tau (MAB3420; Merck Millipore), Alexa-Fluor488-conjugated anti-Tau (MAB3420A4; Merck Millipore), rabbit anti-btubulin and anti-b-actin (ab6046 and ab8227; Abcam, Cambridge, UK),
mouse anti-T7 (69522; Merck Millipore), SOX1 (Abcam), mouse antiHA, mouse FITC-conjugated anti-b-tubulin and acetylated tubulin (all
from Sigma). Western blot secondary antibodies were: Odyssey goat
anti-mouse IRDye 680LT and goat anti-rabbit IRDye 680LT (Li-Cor
Biosciences, Cambridge, UK). Immunocytochemistry secondary
antibodies were: Alexa Fluor 594 anti-mouse, Alexa Fluor 647 antimouse, Alexa Fluor 594 anti-rabbit, Alexa Fluor 555 anti-rabbit, and
Alexa Fluor 488 anti-rabbit (all from Life Technologies Ltd).
Immunoprecipitation
For endogenous JIP1 and AKT pulldown experiments, 36106 neurons
were lysed in 300 ml 1% NP40 lysis buffer (Kim et al., 2002) containing
10 mg/ml aprotinin, 10 mg/ml leupeptin, 1 mM phenylmethanesulfonyl
fluoride and 1 mM NaVO3, and protein complexes were incubated
overnight with 3 mg of anti-AKT1 antibody. Following antibody
incubation, complexes were precipitated by incubation for 1 hour, at
4 ˚C, with 15 ml Dynabeads Protein-G (Life Technologies Ltd).
Immunoblotting
Cells were lysed in TLB buffer (1% Triton X-100, 10% glycerol, 20 mM
Tris-HCl pH 7.4, 137 mM NaCl, 25 mM sodium b-glycerophosphate,
2 mM sodium pyrophosphate, 2 mM EDTA, plus protease inhibitors).
Samples (lysates or immunoprecipitates) were mixed with an appropriate
volume of a 66SDS loading buffer (200 mM Tris-HCl pH 6.8, 7.5% SDS,
0.5 M dithiothreitol, 30% glycerol, 0.1% Bromophenol Blue) and loaded
onto a 10% SDS-polyacrylamide gel, followed by transfer onto a PVDF
membrane. Membranes were blocked with 5% milk in TBS (15 mM TrisHCl pH 7.4, 150 mM NaCl) followed by primary antibody incubation at
4 ˚C overnight: anti-JIP1 at 1:1000, anti-AKT at 1:500, anti-b-tubulin and
anti-b-actin at 1:5000. All dilutions were in 5% milk with TBS–Tween-20
(0.1%). Blots were then washed four times over 30 minutes with TBS–
Tween (0.1%) and incubated for 1 hour, at room temperature with the
corresponding secondary antibody at a dilution of 1:40,000 in TBS–Tween
(0.1%) with 0.01% SDS, followed by three 10-minutes washes with TBS–
Tween (0.1%). Proteins were detected using a Li-Cor OdysseyH System
scanner according to the manufacturer’s instructions.
238
Acknowledgements
We thank James Uney (University of Bristol) for providing the pRRL.sin.cppt.CMVEGFP-WPRE plasmid. We thank Karel Dorey and Richard Monaghan for helpful
comments on the manuscript and Peter March for his advice on microscopy. The
Faculty of Life Sciences Bioimaging Facility microscopes used in this study were
purchased with grants from BBSRC, Wellcome Trust and the University of
Manchester Strategic Fund.
Competing interests
The authors declare no competing interests.
Author contributions
A.J.W., F.D.B. and I.B. designed research; F.D.B., I.B. and E.V.J. conducted
experiments; A.J.W. and F.D.B. wrote the paper.
Funding
This work was supported by the Wellcome Trust [grant number 088232/Z/09/Z to
A.J.W]; and Medical Research Council [grant number G0400620 to A.J.W.]
Biotechnology and Biological Sciences Research Council [studentship number
02/B1/C/08253 to E.V.J. and A.J.W.]. Deposited in PMC for release after 6
months.
Supplementary material
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.137208/-/D
References
Allaman-Pillet, N., Størling, J., Oberson, A., Roduit, R., Negri, S., Sauser, C.,
Nicod, P., Beckmann, J. S., Schorderet, D. F., Mandrup-Poulsen, T. et al.
(2003). Calcium- and proteasome-dependent degradation of the JNK scaffold
protein islet-brain 1. J. Biol. Chem. 278, 48720-48726.
Barnat, M., Enslen, H., Propst, F., Davis, R. J., Soares, S. and Nothias,
F. (2010). Distinct roles of c-Jun N-terminal kinase isoforms in neurite initiation
and elongation during axonal regeneration. J. Neurosci. 30, 7804-7816.
Barnes, A. P. and Polleux, F. (2009). Establishment of axon-dendrite polarity in
developing neurons. Annu. Rev. Neurosci. 32, 347-381.
Journal of Cell Science
Quantitative RT-PCR
Bennett, B. L., Sasaki, D. T., Murray, B. W., O’Leary, E. C., Sakata, S. T., Xu, W.,
Leisten, J. C., Motiwala, A., Pierce, S., Satoh, Y. et al. (2001). SP600125, an
anthrapyrazolone inhibitor of Jun N-terminal kinase. Proc. Natl. Acad. Sci. USA
98, 13681-13686.
Bhaskar, P. T. and Hay, N. (2007). The two TORCs and Akt. Dev. Cell 12, 487-502.
Blasius, T. L., Cai, D., Jih, G. T., Toret, C. P. and Verhey, K. J. (2007). Two
binding partners cooperate to activate the molecular motor Kinesin-1. J. Cell
Biol. 176, 11-17.
Centeno, C., Repici, M., Chatton, J. Y., Riederer, B. M., Bonny, C., Nicod, P.,
Price, M., Clarke, P. G., Papa, S., Franzoso, G. et al. (2007). Role of the JNK
pathway in NMDA-mediated excitotoxicity of cortical neurons. Cell Death Differ.
14, 240-253.
Cheng, P. L. and Poo, M. M. (2012). Early events in axon/dendrite polarization.
Annu. Rev. Neurosci. 35, 181-201.
Dajas-Bailador, F., Jones, E. V. and Whitmarsh, A. J. (2008). The JIP1 scaffold
protein regulates axonal development in cortical neurons. Curr. Biol. 18, 221-226.
Dajas-Bailador, F., Bonev, B., Garcez, P., Stanley, P., Guillemot, F. and
Papalopulu, N. (2012). microRNA-9 regulates axon extension and branching by
targeting Map1b in mouse cortical neurons. Nat. Neurosci. 15, 697-699.
de la Torre-Ubieta, L. and Bonni, A. (2011). Transcriptional regulation of neuronal
polarity and morphogenesis in the mammalian brain. Neuron 72, 22-40.
DiAntonio, A. and Hicke, L. (2004). Ubiquitin-dependent regulation of the
synapse. Annu. Rev. Neurosci. 27, 223-246.
Dotti, C. G., Sullivan, C. A. and Banker, G. A. (1988). The establishment of
polarity by hippocampal neurons in culture. J. Neurosci. 8, 1454-1468.
Fonnum, F. (1984). Glutamate: a neurotransmitter in mammalian brain.
J. Neurochem. 42, 1-11.
Fu, M. M. and Holzbaur, E. L. (2013). JIP1 regulates the directionality of APP axonal
transport by coordinating kinesin and dynein motors. J. Cell Biol. 202, 495-508.
Gallo, G. and Letourneau, P. (2002). Axon guidance: proteins turnover in turning
growth cones. Curr. Biol. 12, R560-R562.
Gogolla, N., Galimberti, I. and Caroni, P. (2007). Structural plasticity of axon
terminals in the adult. Curr. Opin. Neurobiol. 17, 516-524.
Horiuchi, D., Barkus, R. V., Pilling, A. D., Gassman, A. and Saxton, W. M.
(2005). APLIP1, a kinesin binding JIP-1/JNK scaffold protein, influences the
axonal transport of both vesicles and mitochondria in Drosophila. Curr. Biol. 15,
2137-2141.
Jacobson, C., Schnapp, B. and Banker, G. A. (2006). A change in the selective
translocation of the Kinesin-1 motor domain marks the initial specification of the
axon. Neuron 49, 797-804.
Jiang, H., Guo, W., Liang, X. and Rao, Y. (2005). Both the establishment and the
maintenance of neuronal polarity require active mechanisms: critical roles of
GSK-3beta and its upstream regulators. Cell 120, 123-135.
Kennedy, N. J., Martin, G., Ehrhardt, A. G., Cavanagh-Kyros, J., Kuan, C. Y.,
Rakic, P., Flavell, R. A., Treistman, S. N. and Davis, R. J. (2007). Requirement
of JIP scaffold proteins for NMDA-mediated signal transduction. Genes Dev. 21,
2336-2346.
Kim, A. H., Yano, H., Cho, H., Meyer, D., Monks, B., Margolis, B., Birnbaum, M.
J. and Chao, M. V. (2002). Akt1 regulates a JNK scaffold during excitotoxic
apoptosis. Neuron 35, 697-709.
Kim, S. R., Chen, X., Oo, T. F., Kareva, T., Yarygina, O., Wang, C., During, M.,
Kholodilov, N. and Burke, R. E. (2011). Dopaminergic pathway reconstruction
by Akt/Rheb-induced axon regeneration. Ann. Neurol. 70, 110-120.
Lee, L. J., Lo, F. S. and Erzurumlu, R. S. (2005). NMDA receptor-dependent
regulation of axonal and dendritic branching. J. Neurosci. 25, 2304-2311.
Journal of Cell Science (2014) 127, 230–239 doi:10.1242/jcs.137208
Muresan, Z. and Muresan, V. (2005). Coordinated transport of phosphorylated
amyloid-beta precursor protein and c-Jun NH2-terminal kinase-interacting
protein-1. J. Cell Biol. 171, 615-625.
Namikawa, K., Honma, M., Abe, K., Takeda, M., Mansur, K., Obata, T., Miwa,
A., Okado, H. and Kiyama, H. (2000). Akt/protein kinase B prevents injuryinduced motoneuron death and accelerates axonal regeneration. J. Neurosci.
20, 2875-2886.
Nihalani, D., Wong, H. N. and Holzman, L. B. (2003). Recruitment of JNK to JIP1
and JNK-dependent JIP1 phosphorylation regulates JNK module dynamics and
activation. J. Biol. Chem. 278, 28694-28702.
Oliva, A. A., Jr, Atkins, C. M., Copenagle, L. and Banker, G. A. (2006). Activated
c-Jun N-terminal kinase is required for axon formation. J. Neurosci. 26, 9462-9470.
Read, D. E. and Gorman, A. M. (2009). Involvement of Akt in neurite outgrowth.
Cell. Mol. Life Sci. 66, 2975-2984.
Rubinson, D. A., Dillon, C. P., Kwiatkowski, A. V., Sievers, C., Yang, L.,
Kopinja, J., Rooney, D. L., Zhang, M., Ihrig, M. M., McManus, M. T. et al.
(2003). A lentivirus-based system to functionally silence genes in primary
mammalian cells, stem cells and transgenic mice by RNA interference. Nat.
Genet. 33, 401-406.
Ruthazer, E. S., Akerman, C. J. and Cline, H. T. (2003). Control of axon branch
dynamics by correlated activity in vivo. Science 301, 66-70.
Shi, S. H., Jan, L. Y. and Jan, Y. N. (2003). Hippocampal neuronal polarity specified
by spatially localized mPar3/mPar6 and PI 3-kinase activity. Cell 112, 63-75.
Song, J. J. and Lee, Y. J. (2005a). Dissociation of Akt1 from its negative regulator
JIP1 is mediated through the ASK1-MEK-JNK signal transduction pathway
during metabolic oxidative stress: a negative feedback loop. J. Cell Biol. 170,
61-72.
Song, J. J. and Lee, Y. J. (2005b). Crosstalk between JIP3 and JIP1 during
glucose deprivation: SEK1-JNK2 and Akt1 act as mediators. J. Biol. Chem. 280,
26845-26855.
Song, Y., Ori-McKenney, K. M., Zheng, Y., Han, C., Jan, L. Y. and Jan, Y. N.
(2012). Regeneration of Drosophila sensory neuron axons and dendrites is
regulated by the Akt pathway involving Pten and microRNA bantam. Genes
Dev. 26, 1612-1625.
Verhey, K. J., Meyer, D., Deehan, R., Blenis, J., Schnapp, B. J., Rapoport, T. A.
and Margolis, B. (2001). Cargo of kinesin identified as JIP scaffolding proteins
and associated signaling molecules. J. Cell Biol. 152, 959-970.
Whitmarsh, A. J. (2006). The JIP family of MAPK scaffold proteins. Biochem.
Soc. Trans. 34, 828-832.
Whitmarsh, A. J., Cavanagh, J., Tournier, C., Yasuda, J. and Davis, R. J.
(1998). A mammalian scaffold complex that selectively mediates MAP kinase
activation. Science 281, 1671-1674.
Whitmarsh, A. J., Kuan, C. Y., Kennedy, N. J., Kelkar, N., Haydar, T. F., Mordes,
J. P., Appel, M., Rossini, A. A., Jones, S. N., Flavell, R. A. et al. (2001).
Requirement of the JIP1 scaffold protein for stress-induced JNK activation.
Genes Dev. 15, 2421-2432.
Yan, D., Guo, L. and Wang, Y. (2006). Requirement of dendritic Akt degradation
by the ubiquitin-proteasome system for neuronal polarity. J. Cell Biol. 174,
415-424.
Yoshimura, T., Kawano, Y., Arimura, N., Kawabata, S., Kikuchi, A. and
Kaibuchi, K. (2005). GSK-3beta regulates phosphorylation of CRMP-2 and
neuronal polarity. Cell 120, 137-149.
Zhang, T., Inesta-Vaquera, F., Niepel, M., Zhang, J., Ficarro, S. B., Machleidt,
T., Xie, T., Marto, J. A., Kim, N., Sim, T. et al. (2012). Discovery of potent and
selective covalent inhibitors of JNK. Chem. Biol. 19, 140-154.
Journal of Cell Science
RESEARCH ARTICLE
239