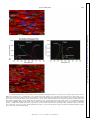
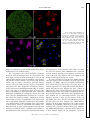
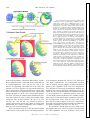
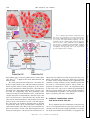
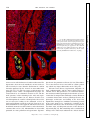
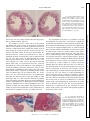
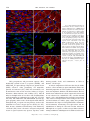
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cell growth wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cell culture wikipedia , lookup
List of types of proteins wikipedia , lookup
Cell encapsulation wikipedia , lookup
Tissue engineering wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cellular differentiation wikipedia , lookup
Physiol Rev 85: 1373–1416, 2005; doi:10.1152/physrev.00013.2005. Cardiac Stem Cells and Mechanisms of Myocardial Regeneration ANNAROSA LERI, JAN KAJSTURA, AND PIERO ANVERSA Cardiovascular Research Institute, Department of Medicine, New York Medical College, Valhalla, New York 1373 1375 1376 1378 1380 1382 1386 1386 1390 1391 1394 1395 1399 1402 Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 I. Introduction II. Exogenous Progenitor Cells and Cardiac Repair A. Human embryonic stem cells B. Hematopoietic stem cells C. Mesenchymal stem cells D. Controversy on bone marrow cell transdifferentiation and cardiac repair III. Cardiac Progenitor Cells A. Identification B. Origin of cardiac stem cells C. Cardiac stem cell niches IV. Cardiac Progenitor Cells and Myocardial Repair A. Myocardial regeneration and ischemic and nonischemic cardiomyopathy B. Cell fusion and myocardial regeneration V. Future Directions Leri, Annarosa, Jan Kajstura, and Piero Anversa. Cardiac Stem Cells and Mechanisms of Myocardial Regeneration. Physiol Rev 85: 1373-1416, 2005; doi:10.1152/physrev.00013.2005.—This review discusses current understanding of the role that endogenous and exogenous progenitor cells may have in the treatment of the diseased heart. In the last several years, a major effort has been made in an attempt to identify immature cells capable of differentiating into cell lineages different from the organ of origin to be employed for the regeneration of the damaged heart. Embryonic stem cells (ESCs) and bone marrow-derived cells (BMCs) have been extensively studied and characterized, and dramatic advances have been made in the clinical application of BMCs in heart failure of ischemic and nonischemic origin. However, a controversy exists concerning the ability of BMCs to acquire cardiac cell lineages and reconstitute the myocardium lost after infarction. The recognition that the adult heart possesses a stem cell compartment that can regenerate myocytes and coronary vessels has raised the unique possibility to rebuild dead myocardium after infarction, to repopulate the hypertrophic decompensated heart with new better functioning myocytes and vascular structures, and, perhaps, to reverse ventricular dilation and wall thinning. Cardiac stem cells may become the most important cell for cardiac repair. I. INTRODUCTION At the boundary of the second and third millennia, the discovery that hematopoietic stem cells (HSCs) can acquire cell lineages different from the organ of origin has started a new intriguing scientific revolution (23, 178, 206, 212, 253, 266, 384, 392, 401, 473). The unpredicted behavior of HSCs has surprised and distressed many of us; they disobey the dogma of embryonic specification and with plastic changes undergo unexpected metamorphoses (150, 384, 438, 487, 502). Because of their potential ability to transdifferentiate (Fig. 1), HSCs have been proposed as a novel form of cell therapy for damaged organs (27, 527). However, this possibility was perceived with excitement www.prv.org and mistrust, which equally divided the scientific and clinical community. The prevailing reaction on the part of the skeptics was disbelief for this alternative view of stem cell biology. Studies reporting negative findings were published, and questions were raised about the validity of the shift in paradigm advanced in the early work (508). This controversy is feeding a fire that may disrupt the stream of scientific consciousness and delay the clinical application of a prospectively revolutionary therapeutic tool. In addition to the plasticity of HSCs and their ability to regenerate damaged organs, the function of resident stem cells is to maintain tissue homeostasis in response to perturbations. The documentation of the existence of organ-specific adult stem cells has created great expecta- 0031-9333/05 $18.00 Copyright © 2005 the American Physiological Society 1373 1374 LERI, KAJSTURA, AND ANVERSA tions concerning their utilization as a strategy for the management of the human disease (36, 51, 59, 66, 139, 343, 374, 387, 457, 500). If the goal of medicine has been to treat symptoms of diseases and possibly remove their causes, scientists and physicians point now to a more ambitious target. The primary objective of regenerative medicine is the complete structural and functional recovery of the damaged organ (245, 315, 490). Regeneration coincides with tissue homeostasis and involves the replacement of cells lost by normal wear and tear and, ideally, following injury (143, 499). The regenerative capacity of organs is a property of particular significance in organisms with long lifespan; in fact, the preservation of the components of each tissue and their functional integration is essential for survival. Damage creates a barrier to restitutio ad integrum and promotes the initiation of a repair process that leads to the formation of a scar. Scar formation is crucial for rapid handling of the damage, to seclude the lesion from healthy tissue and to prevent a cascade of uncontrolled deleterious events (41, 325). However, the scar does not possess the biochemical, physical, and functional properties of the uninjured tissue and, therefore, negatively affects the overall performance of the organ (52, 263, 345, 377, 435, 534). In spite of the presence of adult stem cells in selfrenewing organs, tissue repair involves scar formation (280, 410, 512). Understanding the factors that dictate the evolution of a lesion into scarring instead of regeneration is at its beginning. Some insights come from embryonic Physiol Rev • VOL development. Tissue repair in embryos is rapid, efficient, and scar free (143). Skin wounds in the early mammalian embryo heal with restitutio ad integrum, whereas wounds in late gestational fetuses and adult mammals result in scarring. This difference may be related to the intrinsic characteristics of embryonic fibroblasts or to external stimuli. Fibroblasts in embryos are less prone to synthesize and release collagen, attenuating the amount of fibrosis following injury (287). The presence of hyaluronic acid and fibronectin in the amniotic fluid may also interfere with scar formation during wound healing (269). Moreover, the inflammatory response is attenuated; a low number of poorly differentiated inflammatory cells accumulate in the region of damage, and the growth factors present at the site of healing in embryos are different from those in the adult organism (143). The expression of transforming growth factor- (TGF-) isoforms, which have powerful profibrotic effects, and their receptors (TBRI, TBRII) vary in the skin of embryos at different gestational ages (92, 143). TGF-3 is more abundant in early embryo, while the amount of TGF-1 and TGF-2 increases progressively with development. Similarly, TBRI and TBRII increase with maturation. The different ratios of TGF- isoforms in the skin appear to condition the evolution of healing triggering profibrotic or antifibrotic signals in prenatal and postnatal life (5). Adult wounds in mice, rats, and pigs, exposed to the same profile of cytokines present in the embryo, experience a more effective repair process with reduction 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 1. Plasticity of adult stem cells. Stem cells acquire cell phenotypes different from their organ of origin. The confocal image illustrates the transdifferentiation of male c-kit positive bone marrow-derived cells (BMCs) into cardiomyocytes after infarction in a female mouse. Myocytes are labeled by cardiac myosin heavy chain (red) and nuclei by propidium iodide (PI; green). The Y chromosome in nuclei is shown by white dots. 1375 ADULT STEM CELLS II. EXOGENOUS PROGENITOR CELLS AND CARDIAC REPAIR In embryonic and early postnatal life, the increase in cardiac mass reflects the delicate balance between the addition of new myocytes and the death of unnecessary cells (102, 393). Shortly after birth, cardiac growth occurs by an increase in myocyte number and volume, which together contribute to the development of the adult heart phenotype (29, 31). In adulthood, the heart is characterized by a surprisingly high and rapid turnover of its parenchymal cells that is regulated by a stem cell compartment (51, 201, 293, 296, 308, 338). Under physiological conditions, the intrinsic growth reserve of the heart is sufficient to maintain cell homeostasis and adequate pump performance. However, when an increase in pressure and/or volume load is imposed on the heart, myocyte hypertrophy and proliferation in combination with myocyte apoptosis and necrosis constitute the elements of myocardial remodeling (18, 20, 25, 27, 327). Novel methodological approaches applied to the analysis of the myocardium have challenged the paradigm introduced more than 60 years ago that the heart is a postmitotic organ and myocytes are terminally differentiated cells that participate in cardiac function throughout life (7, 97, 288, 329, 339, 340, 354). A subset of parenchymal cells expresses the molecular components mediating the entry and progression of these cells through the cell cycle, and karyokinesis and cytokinesis. The recognition Physiol Rev • VOL that myocyte replication, hypertrophy, and death occur in the pathological heart has significantly enhanced our understanding of the dynamic nature of the myocardium and the critical role that these variables have in the preservation of cardiac performance or in the onset of ventricular dysfunction (22, 25, 27, 34, 326, 327). Contrary to the general belief, the restricted regenerative capacity of the myocardium does not represent the initial causal event of impaired cardiac function. The nonischemic failing heart, in its early phases of decompensation, has a number of myocytes that often exceeds the number of cells in a normal heart. Alternatively, the modest reduction in myocyte number chronically is not consistent with the severe deterioration in ventricular performance. A typical example is found in hypertensive cardiomyopathy or chronic aortic stenosis in animals and humans (29, 30, 497). The initiation and evolution of cardiac failure appears to depend mostly on two other crucial factors that are strictly interrelated in the determination of pump function: the accumulation of old, poorly contracting cells and the formation of multiple foci of myocardial scarring (52, 53, 67, 98, 484, 497). A severe reduction in cell number, however, occurs acutely after ischemic injury that inevitably results in scar formation not only in the heart but also in other organs, whether their parenchymal cells are highly proliferating, slowly cycling, or terminally differentiated (263, 280, 410, 512). The need to overcome this biological obstacle has favored the development of strategies aiming at the replacement of dead cells with viable cells. Several interventions have been used in an attempt to promote cardiac regeneration experimentally. These protocols have employed different cell types, including fetal tissue and fetal and adult cardiomyocytes (264, 395, 444, 544), skeletal myoblasts (306, 323, 466), embryonic-derived myocytes and endothelial cells (109, 142), bone marrow-derived immature myocytes (194), fibroblasts (165), smooth muscle cells (272), and bone marrow c-kit positive and negative progenitor cells (144, 217, 226, 228, 356, 357, 539, 545). Unexpectedly, these multiple approaches had a rather uniform outcome that consisted of variable degrees of improvement in cardiac contractile function. Most likely, the implanted cells formed a passive graft, which reduced negative remodeling by decreasing the stiffness of the scarred portion of the ventricular wall. The elastic properties of the implanted cells could be the major cause for enhanced heart function. An active graft, which dynamically contributed to myocardial performance, was observed only in a few cases (144, 228, 356, 357, 539). The possibility has also been advanced that the implanted cells exert a paracrine effect on resident cardiac stem cells (CSCs) activating myocardial regeneration (50, 539). Despite these efforts, however, the most appropriate form of cell therapy for actual restitutio ad integrum of the damaged myocardium has not yet been identified. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 in scar formation (143). Although scar-free regeneration of damaged organs is well known in amphibians and zebrafish (103, 385, 451), there is only one example in mammals in which repair occurs in the absence of fibrosis. Cartilage, skin, hair follicles, and cryoinjured myocardium regrow in the adult MRL mouse (197, 262). However, ischemia-reperfusion injury leads to comparable infarct size in wild-type and MRL mice (3), suggesting that the native capacity to reconstitute dead myocardium after infarction may be limited in this model. In summary, modulation of the inflammatory response may improve tissue repair in the adult organism. However, the removal of these extrinsic signals may not be sufficient for long-term regeneration of the injured organ. Resident stem cells in proximity to the lesion can change the properties of the microenvironment by secreting cytokines that contribute to cell homing, proliferation, and differentiation (50, 239, 240, 291, 482). Enhancement of the intrinsic growth reserve of the organ constitutes the foundation of regenerative medicine and regenerative cardiology. This review discusses current understanding of the role that endogenous and exogenous progenitor cells may have in the treatment of ischemic and nonischemic heart failure. 1376 LERI, KAJSTURA, AND ANVERSA A. Human Embryonic Stem Cells Embryonic stem cells (ESCs), which are derived from the inner mass of the blastocyst (337, 369, 397, 477), possess unique properties. These cells can be grown in vitro and propagated indefinitely in their undifferentiated state while retaining a normal karyotype. Additionally, they maintain over time the property of multilineage commitment and, therefore, can differentiate in vitro into all cell types present in an organism (49, 88, 337, 525). Because of their enormous potential, human ESC lines have been obtained from the human blastocyst (369, 477) with the expectation of a future successful application of their broad therapeutic potential to patients (124, 233, 370). However, a controversy exists between the therapeutic relevance of ESCs and autologous adult progenitor cells (PCs). The debate concerning the efficacy of ESCs and adult PCs is unfortunate because the utilization of ESCs in clinical practice is at least a decade away while adult PCs present minimal risks and are the only applicable form of cell therapy for myocardial regeneration today. This does not exclude that ESCs may become in the future a powerful alternative to adult PCs. Human ESCs cultured in suspension form spontaneously cellular aggregates called embryoid bodies (EBs). EBs are composed of cells derived from the three germ layers (118). These teratoma-like structures consist of semiorganized tissues composed of cardiomyocytes, striated skeletal muscle cells, neuronal rosettes, and hemoglobin-containing blood islands. However, they lack critical features of embryonic patterning (216, 231). The morphology of solid EBs can be affected by the process of Physiol Rev • VOL cavitation that results in the formation of cystic EBs. During their formation, markers of the mesoderm (globin), ectoderm (neurofilament 68), and endoderm (␣fetoprotein) are expressed (187, 216). Synchronously pulsing EBs contain myogenic cells positive for ␣-cardiac actin, desmin, myogenin, and smooth muscle actin (44, 234). Consistent with these in vitro observations, in vivo delivery of human ESC lines to skeletal muscle or the testis gives rise to teratomas (397, 459, 477). Teratomas include endodermal, mesodermal, and ectodermal structures formed by mature cells (Fig. 2). Conversely, teratocarcinomas contain undifferentiated cells and possess a stem cell compartment (16, 88). Although the malignant tumorigenic potential of ESCs is not well defined yet, it is worrisome that ESCs and the embryonal carcinoma (EC) cells, i.e., the stem cell of teratocarcinomas, share several markers including Oct4 and Nanog (88, 89, 193, 342, 363). Moreover, the self-renewal property of ESCs and EC cells is modulated by similar pathways (88, 191, 458). Conversely, the ability to induce formation of human EBs containing cells of neuronal, hematopoietic, and cardiac origins is important for the recognition of early human embryonic development. In the last 15 years, this accessible in vitro system has been exploited to elucidate differentiation events in several tissues (118, 126, 397, 477). The study of ESCs may provide a unique understanding of the mechanisms of embryonic development that may lead to therapeutic interventions in utero and the correction of congenital malformations. This may become the most exciting outcome of ESC research. In view of the risks associated with the broad differentiation potential of ESCs in vivo, only sporadic reports employed these cells in their uncommitted state to repair the damaged heart (203, 243, 244, 312, 313, 509). In sharp contrast to the pluripotency of ESCs, cardiomyocytes were detected, but capillary structures and resistance arterioles were not found. The reconstituted tissue did not possess the characteristics of functionally competent myocardium. However, human ESCs differentiate in endothelial cells in vitro and form microvessels in vivo in the subcutaneous tissue of SCID mice (268). The differentiation of ESCs into cardiomyocytes has been attributed to the paracrine effects of host signals, including members of the TGF- superfamily of proteins (50, 244). Although some degree of functional improvement was observed, the therapeutic utilization of ESCs can be associated with dramatic side effects. Teratomas are benign tumors, but the cardiac localization confers on them a malignant biological behavior. Most importantly, ESCs injected intravenously can engraft and colonize all organs with the possible development of neoplastic lesions (509). In an attempt to avoid these complications, ESCs have been partially differentiated in vitro before their implantation in the injured heart (235, 284, 313, 535, 542). Some degree of cardiomyogenic commitment was 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 It is intuitively apparent that if the adult heart possesses a pool of primitive multipotent cells (51, 201, 293, 296, 338), these cells must be tested first before more complex and unknown cells are explored. The attraction of this approach is its simplicity. Cardiac regeneration would be accomplished by enhancing the normal turnover of myocardial cells. However, major difficulties exist in the acquisition of myocardial samples and the isolation and expansion of CSCs in quantities that can be employed therapeutically. Until protocols capable of resolving these limitations are implemented, CSC therapy remains a difficult task. Alternatively, CSCs may be activated locally within the myocardium, but the growth factor-receptor systems that regulate CSC proliferation and differentiation are largely unknown (277, 335). Although the importance of resident CSCs in organ homeostasis is obvious and, in the future, CSCs may become the most appropriate form of cell therapy for the diseased heart, currently, emphasis has been placed on the identification of exogenous sources of highly proliferating cells that can acquire the cardiac phenotypes. 1377 ADULT STEM CELLS claimed to enhance engraftment of the cells into the myocardium, attenuate the probability of ESCs to acquire undesired cell lineages, and thereby reduce the risk of teratoma formation (397, 477, 492). Although immature myocytes have been obtained from ESCs and their morphological, phenotypical, and functional properties characterized (196, 234, 235), the residual growth potential of the committed cells was not established. Additionally, the purity of the preparation remains a major problem since pluripotent ESCs may persist among the enriched population of developing myocytes and injection of this cell pool may engender tumor formation. There are no rigorous methodologies for ESC differentiation into cardiomyocytes in vitro (62, 196, 198, 234, 235, 519, 548). It is emblematic that eight distinct growth factors have forced ESCs to acquire only in part a specific cell phenotype while other undesired cells were consistently present in the preparations (417). None of the growth factors was capable of driving the commitment to a single cell type, making it impossible, at present, to obtain a uniform population of mature progeny from ESCs in culture. However, the plasticity of ESCs is greater than that of adult stem cells. The implantation of beating EBs reverses complete atrioventricular block in pigs (235). The morphology of the grafted cells resembled those of embryonic cardiomyocytes being of small size with disorganized myofibrils located at the periphery of the cells. The mechanical and electrical behavior of beating EBs indicates that human ESCs give rise to cells that display functional Physiol Rev • VOL properties typical of the embryonic human myocardium. When the electrophysiological analysis of the newly formed cells was associated with the analysis of their gene expression profile, the conclusion was reached that cardiomyocytes derived from ESCs maintain a phenotype reminiscent of the embryonic heart tube and hardly myocytes develop a chamberlike myocardial phenotype. Whether these immature myocytes can reach adult characteristics in vivo remains an unanswered question (147). The immunorejection of the allogeneic ESCs or their differentiated progeny constitutes an additional limitation inherent in the use of ESCs (128). Because of their early stage of development, ESCs were considered “immune-privileged” and therefore not recognizable by the immune defenses of the recipient (477). Conversely, recent evidence suggests that even in their undifferentiated state, human ESCs express discrete levels of HLA class I antigens that increase as the cells mature (127, 294). The immunosuppressive regimen is poorly tolerated, and this treatment severely impairs a patient’s quality of life. The immunodeficient state of mice that develop teratomas after ESC implantation (397, 477, 509) closely reflects the immunosuppressed condition that may have to be reached clinically to avoid rejection of ESCs. However, these limitations should not discourage the promoters of ESCs. Intense research in this area is expected to resolve these current biological problems and change the skepticism concerning the future use of ESCs for human disease. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 2. Embryonic stem cells. During development, embryonic stem cells (ESCs) give rise to the ectoderm, mesoderm, and endoderm. ESCs are obtained from the inner cell mass of the blastocyst and in vitro form embryoid bodies (EBs) that contain undifferentiated (yellow), ectodermal (green), mesodermal (blue), and endodermal (red) cells. Implantation of EBs in vivo can repair damaged organs and generate tumors. 1378 LERI, KAJSTURA, AND ANVERSA B. Hematopoietic Stem Cells Before discussing bone marrow cell (BMCs) transdifferentiation, a few comments concerning the developmental origin of the hematopoietic system and the heart may be relevant. Blood cells derive from the mesoderm (328, 516), and the hematopoietic progenitors accumulate first in blood islands in the early somites, migrate then to the fetal liver and, ultimately, reach the bone marrow, which represents their definitive localization in late embryogenesis and adulthood (328, 431, 516). Concurrently, the primitive heart is generated within the anterior lateral mesoderm (100, 133, 138). Eisenberg et al. (134) have shown that the early mesoderm has the capability of creating both blood and cardiac cells. The specification of organs that arise from adjacent regions in the developing embryo may differ by a single developmental decision, affecting the expression of a relatively small number of master switch genes (438, 485– 487). This modification in cell fate has been defined as direct transdifferentiation (56, 132, 426) to contrast the transdifferPhysiol Rev • VOL entiation that requires a dedifferentiation step (256, 257, 316) or the transdifferentiation that occurs through the involvement of a stem cell (10, 27, 178, 327, 392, 438, 487). If this were also the case for the bone marrow and the heart, differentiated BMCs could undergo a direct form of transdifferentiation and generate new myocardium. Although this remains a possibility, committed CD45 positive myeloid cells appear to be able to give rise to skeletal muscle cells or hepatocytes (79, 80). However, so far, only c-kit-positive immature myelomonocytic progenitors can contribute to muscle fibers while more mature CD11b positive myelomonocytic cells do not transdifferentiate into skeletal muscle (125). Embryonic hematopoietic progenitors acquire the cardiomyocyte phenotype in the presence of retinoic acid, dexamethasone, PGE2, interleukin (IL)-2, fibroblast growth factor (FGF)4, and bone morphometric protein (BMP)4 (136). Although the efficiency of transdifferentiation is low, the committed embryonic progenitors express transcription factors and contractile proteins of cardiac muscle cells. The creation of cardiomyocytes is markedly increased when lineage negative Sca-1-positive adult BMCs are cocultured with embryonic mesoderm. The new myocytes integrate functionally with the embryonic tissue (100). Together, these lines of evidence support the notion that BMCs can form cardiac and skeletal muscle cells in vitro and in vivo. This possibility has been strengthened by recent observations in which c-kit-positive fetal liver hematopoietic progenitor cells have been employed successfully to regenerate myocytes and coronary vessels in the infarcted heart (258). These hematopoietic progenitors were obtained from cloned embryos derived from somatic cell fusion between nuclei of LacZ-positive fibroblasts and enucleated oocytes of a different mouse strain (256, 257, 259). The newly formed myocardium replaced 38% of the infarct at 1 mo (258). The rebuilt tissue expressed LacZ and was composed of myocytes and vessels connected with the primary coronary circulation. Myocytes were functionally competent and expressed contractile proteins, desmin, connexin43, and N-cadherin. These structural characteristics indicated that the developing new myocytes were coupled electrically and mechanically (258). Similarly, the formed coronary arterioles and capillary structures contained blood cells and contributed, therefore, to tissue oxygenation. Cardiac replacement resulted in an improvement of ventricular hemodynamics, attenuation of cardiac remodeling, and reduction of diastolic wall stress. These beneficial effects were obtained by stem cell transdifferentiation and commitment to the cardiac cell lineages. In fact, myocardial growth was independent from fusion of the injected progenitor cells with preexisting partner cells. Although problems currently plague nuclear transplantation, including the potential for epigenetic and imprinting abnormalities, fetal hematopoietic progenitor cells derived from cloned embryos are sufficiently normal to repair dead myocardium in vivo. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 Finally, the potential application of human ESCs poses profound ethical concerns. A fundamental issue involves the notion of the moral status of human embryos. However, when the ethical principles are taken to the extreme, they threaten to paralyze public funding for ESCs (523), leaving experimentation to the private sector and, thereby, precluding the possibility to effectively monitor practices or the search for alternatives (149, 167, 285). The privatization of ESCs has reached a point that protocols of cryopreservation have been developed in spite of the fact that we are far from the actual documentation of the efficacy and safety of ESCs. Active research on human ESCs is highly desirable, but the duty to heal sick individuals cannot overcome the moral imperative to treat human beings as subjects and not as objects. An interesting viewpoint has recently been proposed (254). In parallel with the definition of organismic death of the adult, which occurs when the criteria for brain death are met, an embryo can be considered organismically dead when it has lost a fundamental function. This consists of continued, integrated cellular division, growth, and differentiation. This premise implies that irreversible arrest of cell division rather than the death of each and every cell corresponds to the organismic death of the embryo (189, 254). Nearly 60% of the in vitro fertilized embryos fail to cleave at 24 h (261). Often, this growth inhibition reflects severe genetic abnormalities but not necessarily all the cells of the embryo are abnormal. In these mosaic embryos, at least one normal diploid blastomere is found (254). Thus the ethical framework for organ donation can be applied to the harvesting of human ESCs from dead embryos. This would provide a common basis in which the need to protect human dignity and progress in biomedical research are no longer in conflict. 1379 ADULT STEM CELLS Physiol Rev • VOL Consistently, the injection of BMCs improves the performance of the pathological heart (144, 217, 228, 356, 539), but the mechanism by which the administration of BMCs results in enhanced cardiac function remains controversial (135, 137, 200, 247, 295). In spite of these uncertainties, clinical trials have been completed and some are ongoing (39, 69, 158, 163, 186, 273, 372, 373, 415, 421, 448, 449, 452, 528). The safety of this approach has been documented, and double-blind clinical studies are in progress. Limitations in the analysis of myocardial regeneration in humans due to the difficulty in obtaining cardiac biopsies, together with contrasting findings in animals, have prompted different interpretations of the positive outcome of BMC administration. Three possibilities have been advanced. They include the development of coronary vessels that rescue hibernating myocardium, de novo formation of myocytes and vascular structures, or the activation and growth of resident progenitor cells via a paracrine effect mediated by the implanted BMCs (17, 50, 135, 144, 217, 226, 228, 239, 356, 357, 460, 539, 545). These mechanisms of action of BMCs are not mutually exclusive and may be operative in the rescue of the injured heart. However, it is relevant to discuss available information to establish what has been shown so far and what has to be done to document unequivocally the actual role of BMCs in the management of cardiac diseases. This analysis can only be performed by comparing protocols and data accumulated in animal studies. The efficacy of adult BMCs for myocardial regeneration after infarction was documented four years ago (356). BMCs of male mice heterozygous for EGFP were collected, immunodepleted for lineage markers, and then sorted for the stem cell antigen c-kit. Enriched lineage negative c-kit-positive cells were injected in the border zone of infarcted mice where they colonized to the dead tissue and gave rise to contracting myocardium occupying 68% of the original infarct. The newly formed EGFP-Y-chromosome positive cells corresponded to functionally competent myocytes and vascular cells organized in coronary arterioles and capillary structures. Overall, the aspect of the newly formed myocardium was that of a rather immature tissue with myocytes small in size, ⬃500 m3, and vessels with a small lumen and a thick multilayered wall. Myocardial regeneration with amelioration of cardiac performance was obtained only in 40% of the treated mice. Coronary ligation in mice is a complex procedure with an inherent variability in infarct size and a 50% probability of correct injection (228, 356). The mouse heart beats ⬃600 times/min and has a left ventricular (LV) wall that is ⬍1 mm thick. These factors make the injection of cells within the LV wall highly problematic. When these technical difficulties were overcome by the mobilization of BMCs with the systemic administration of stem cell factor (SCF) and granulocyte-colony stimulating factor (G-CSF), myocardial regeneration was obtained in all infarcted treated animals. The new myocardium was com- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 The adult bone marrow contains several cell populations. In addition to differentiated cells, such as stroma cells, vascular cells, adipocytes, osteoblasts, and osteoclasts, primitive cells reside in the bone marrow. This class of undifferentiated cells with stem cell properties is heterogeneous (383, 431, 457, 516), being composed of HSCs and mesenchymal stem cells (MSCs). These two subsets of cells have been employed in the repair of damaged organs giving rise to tissues distinct from the organ of origin (178, 206, 245, 253, 295, 315, 390, 392, 473, 490). However, a mixed population of bone marrow cells rather than a highly purified stem cell pool has commonly been used for the restoration of injured organs (64, 68, 131, 211, 275, 357, 360, 388, 475). In the context of this review, we have defined this heterogeneous cell population as BMCs, i.e., a cell preparation that contains an enriched pool of HSCs together with variable degrees of MSCs and endothelial progenitor cells. In 1961, Till and McCullock discovered clonogenic bone marrow cells that gave rise to multilineage hematopoietic colonies in the spleen (48, 436, 479). Till and McCullock (480) proposed that these cells were multipotent HSCs that possess the properties of self-renewal and multilineage differentiation. The definition of stem cells has not changed, and bone marrow HSCs are functionally defined by their capacity to self-renew, form clones, and differentiate into mature blood cell types (431, 516, 517). Although HSCs were discovered more than four decades ago, their isolation has become possible only more recently when surface markers were identified (445, 493, 517). Different epitopes characterize HSCs in different species. Mouse HSCs correspond to a subpopulation of BMCs that are c-kit⫹Sca-1⫹Thy-1-low (358, 493). Operationally, this population is multipotent and repopulates the bone marrow of irradiated mice (493). However, the c-kit⫹Sca-1⫹Thy-1-low cells are functionally heterogeneous. Based on the identification of additional surface markers and clonal analysis, the Mac-1-negative-CD4-negative cells within this category are enriched for long-term reconstituting cells (320). Conversely, the Mac-1-lowCD4-negative pool contains short-term repopulating cells. Finally, the Mac-1-low-CD4-low cells correspond to transient multipotent progenitors together with B lymphocyte progenitors (319). The CD34 antigen has been used to sort human HSCs (331) which, however, are also present in the CD34 negative fraction (123, 181). AC133 is another marker of human HSCs (57) but, so far, there is no specific epitope for the true stem cell. The discovery that adult HSCs or BMCs retain a remarkable degree of developmental plasticity and may have the potential to differentiate across boundaries of lineage and tissue (9, 10, 150, 438, 487, 502) has divided the scientific and clinical community (324, 507, 508, 518). Therefore, the therapeutic efficacy of BMCs for the damaged heart has been questioned (112, 137, 295, 309, 508). 1380 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL vascularization and cardiomyogenesis (539). Thus these findings support the notion that BMCs adopt the cardiac phenotype and potentiate the growth reserve of the adult heart. Collectively, these observations point to the therapeutic import of BMCs for cardiac diseases in humans (17, 39, 69, 158, 163, 239, 273, 372, 373, 415, 421, 448, 449, 452, 528). C. Mesenchymal Stem Cells There is no accepted definition of MSCs. Originally, MSCs were thought to correspond to the putative marrow cells that self-renew and give rise to one or more mesenchymal tissues (314). However, the marrow stromal cell population that contains the MSC pool can also differentiate into tissues other than those that originate in the embryonic mesoderm, raising questions about the appropriateness of the term mesenchymal for this type of stem cell (413, 505, 529). In the majority of studies, the adherent fibroblastic cells obtained from the unfractionated mononuclear cell class of the bone marrow are termed mesenchymal stem or progenitor cells (8). But this heterogeneous cell population is recognized to be too “crude” to represent the purified pool of MSCs (383). A common and widely used characterization of MSCs is that they are the nonhematopoietic multipotent stem cells of the bone marrow (314, 383). The notion of MSCs was introduced in 1961 by Friedenstein (154) who documented that marrow stroma cells contain osteogenic progenitors. Subsequently, the culture conditions necessary for the expansion and differentiation of this cell category were developed. Marrow stromal cells were found to be highly proliferative, clonogenic, and capable of forming colonies of different size and density, composed of several mesenchymal lineages (86, 87, 155, 453, 465). In the late 1980s and in the 1990s, MSCs acquired a significant biological role with the work performed by Caplan (84), who suggested that the actual MSC is an adherent fibroblastic cell isolated by Percoll density centrifugation. This postulated MSC expressed antigens reactive with the monoclonal antibodies SH2 and SH3 that recognize CD105 and CD73, respectively (195). The disadvantage of this traditional method of isolation is the inevitable contamination from HSCs and the cellular heterogeneity of the preparation. Over the past decade, Caplan’s definition has been questioned as being not restrictive enough for the ideal MSC. Although CD105 is a relevant epitope of MSCs and angiogenesis (314, 382), CD29, CD44, and CD90 have been proposed to be important determinants of the mesenchymal phenotype (382, 383) while STRO-1 may identify an immature state (175). The immunophenotype of MSCs has only been partially determined, although the SH2, SH3, CD29, CD44, CD71, CD90, CD106, CD120a, and CD124 epitopes have consis- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 posed of contracting myocytes and patent coronary vessels (357). These studies prompted other investigators to document the ability of BMCs to repair the damaged heart (144, 217, 226, 228, 334, 411, 539, 545). Initially, the hypothesis tested was whether spontaneous mobilization of BMCs occurs after infarction and whether these circulating cells home to the region of injury rescuing the dead tissue (217). Although the degree of engraftment and transdifferentiation in coronary vessels was low and markedly exceeded that of cardiomyocytes, this report confirmed the plasticity of BMCs. The difference in the degree of myocardial reconstitution between this and the previous studies can be attributed to several factors. They include the pathologic model, ischemia-reperfusion injury in irradiated mice (217) versus permanent coronary occlusion in nonablated mice (228, 356, 357), together with the modality of intervention, spontaneous mobilization of BMCs (217) versus intramyocardial injection of BMCs (228, 258, 356) or cytokine-mediated BMC mobilization (357), and the type of BMCs, c-kit enriched BMCs versus peripheral blood cells. The bone marrow of irradiated mice was repopulated with a subset of cells capable of excluding the Hoechst 33342 dye (217). These cells are positive for both c-kit and Sca-1 and are probably more enriched for true HSCs than the lineage negative c-kit positive cells (112) implemented in other studies (228, 356, 357). In spite of several unresolved issues, these observations are consistent with the notion that BMCs can adopt the cardiogenic fate by forming myocytes and coronary vessels. More recently, the possibility that cell therapy of the infarcted heart exerts its beneficial effects not only by reconstitution of dead myocardium but also by the activation of resident progenitor cells in the spared portion of the ventricular wall has been carefully examined (228, 291, 539). To properly evaluate the formation of cardiomyocytes and coronary vasculature in the region bordering the infarct and distant from the infarct, the accumulation of newly formed myocytes and vascular structures was measured utilizing markers of the cell cycle and morphometric methods. Although similar protocols were employed in the analysis of myocyte and vessel growth, positive and negative results were obtained concerning the paracrine effects of the administered BMCs (228, 539). However, an enriched population of mouse c-kit-positive BMCs was used in one case (228) and a novel human bone marrow stem cell in the other (539). Both cell populations differentiated into cardiac cell lineages and repaired the infarcted heart, but the latter also promoted a robust regenerative response in the surviving myocardium. Thus a specific human BMC has the ability to transdifferentiate in cardiac muscle cells, smooth muscle cells, and endothelial cells in vitro and in vivo (144, 539). Additionally, this unique cell population can induce endogenous neo- 1381 ADULT STEM CELLS Physiol Rev • VOL telomerase incompetent and have short telomeres. The senescent phenotype can be reversed by forced expression of human telomerase (2). During the phase of amplification, MSCs do not differentiate spontaneously but do so in the presence of growth factors and cytokines (72, 85, 456). They can acquire multiple phenotypes including osteoblasts, chondrocytes, adipocytes, endothelial cells, and neuronal-like cells (99, 221, 286, 382, 383, 405, 412, 422, 538). Bone marrow MSCs differentiate into cardiomyocytes in the presence of 5-aza-cytidine (159, 185, 290); the morphology of the cells changes from a spindle-shaped to a ball-like form and, subsequently, a rod-shaped configuration. The cells then fuse in a syncitium-like structure that resembles a myotube (160). Although these characteristics mimic the organization of skeletal muscle cells, the committed progeny expresses markers commonly found in cardiomyocytes at different stages of fetal development (185). Transcription factors of the cardiac and myocyte lineage, GATA4, Nkx2.5, and HAND1/2, have been detected (159). Interestingly, a switch in MEF2 isoform occurs. From early to late passages, MEF2C is replaced by MEF2A and MEF2D. Additionally, the -isoform of cardiac myosin heavy chain is more abundant than the ␣-isoform. Similarly, ␣-skeletal actin predominates, and ␣-cardiac actin is moderately present. Myosin light-chain 2v is also expressed. Functionally competent ␣- and -adrenergic and muscarinic receptors have been found (185). Cells beat spontaneously and synchronously, and the rate of contraction increases with stimulation by isoproterenol. Conversely, contractile activity is inhibited by 1-blocking agents (290). Thus the differentiation process of these cardiomyogenic cells recapitulates in part the developmental program of gene expression of prenatal life. An elusive stem cell that may or may not belong to the MSC category is the primitive cell identified by Verfaillie and co-workers (225). This human or mouse multipotent adult progenitor cell (MAPC) can be obtained by growing CD45 and glycoprotein A-depleted BMCs in selective culture conditions (Fig. 3). When this cell population is plated at low density in laminin-coated dishes with a low serum medium containing EGF and PDGF, MAPCs appear several months later after ⬃20 population doublings. In this in vitro system, MAPCs differentiate into cells of the three germ layers including hepatocytes, endothelial cells, and neurons (398, 419). This multilineage differentiation occurs also in vivo when MAPCs are injected in the early blastocysts of mice. However, in adult sublethally irradiated immunodeficient mice, the distribution of systemically delivered MAPCs is much more restricted. MAPCs engraft only in hematopoietic tissues, lung, liver, and intestine but not in the heart (Fig. 3). Whether MAPCs injected locally in the myocardium acquire the myocyte lineage has not been tested. In sum- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 tently been found in MSCs (85, 383, 456). Moreover, MSCs are negative for markers of the hematopoietic lineage, including CD34 and the common leukocyte antigen CD45 (43, 383). However, Prockop and others (105, 423, 439) support the notion that MSCs cannot be distinguished solely by antigen expression but necessitate functional assays demonstrating their multipotent growth and differentiation behavior. Thus a definitive consensus on the properties of MSCs has not been reached yet. Putative MSCs have been identified in embryonic, fetal, and postnatal organs (84, 389). During prenatal life, mesenchymal progenitors accumulate in sites harboring HSCs (307). MSCs appear in the aorta-gonad-mesonephros region and colocalize with HSCs (117). Additionally, MSCs are found in the embryonic circulation including the cord blood and amniotic fluid (213, 214, 400, 489). In adulthood, the bone marrow and the systemic circulation constitute the main sources of MSCs (389, 402), although they have also been detected in tissues distant from the bone marrow. The oval cells of the liver, prostatic stem cells, metanephric mesenchymal cells, precursors of the Leydig cells in the testis, primitive osteoprogenitors, preadipocytes, and satellite cells of the skeletal muscle have been classified as MSCs (13, 114, 399, 488, 543, 552). Because not all studies agree that MSCs reside in these mesenchymal tissues, the possibility has been raised that a long-distance traffic of MSCs occurs between the bone marrow and distant sites through the bloodstream. Alternatively, embryonic primordia of MSCs may be stored in organs of mesodermal origin and, in response to injury, may participate in tissue repair (383). Characteristically, MSCs adhere quickly to the culture dish and grow, forming colonies that become visible 1 wk after plating (314, 383, 389). Only 0.001– 0.01% of the initial unfractionated bone marrow cell population consists of MSCs (383, 389). Some preparations of MSCs can be expanded over 15 cell doublings, while others cease replicating after ⬃4 cell doublings (389). This difference may depend on several determinants, including the procedure used to harvest the marrow (314), the low frequency of MSCs in marrow harvests (84), and the age or condition of the donor from which MSCs were prepared (450). This limited growth potential is associated with preservation of the karyotype, telomerase activity, and telomere length (381, 382). The constant level of telomerase activity and the absence of telomeric shortening have been interpreted as unique properties of MSCs, which were considered capable of escaping replicative senescence. However, MSCs from young donors experience ⬃20 population doublings while MSCs from old donors are compromised in their proliferative capacity (381, 450). Extensive subcultivation impairs cell function, and evident signs of senescence (119) and/or apoptosis (314) become apparent. Both young and old MSCs undergo senescence-associated growth arrest (314); they become 1382 LERI, KAJSTURA, AND ANVERSA mary, MAPCs could represent a rare population of MSCs or a by-product of long-term culture. Although MSCs are capable of self-renewal and multilineage differentiation, whether they are involved in the physiological turnover of cells in organs in which they reside remains unclear. However, MSCs are considered a powerful cell for reconstructive and gene-targeting therapy (245). They are easily transfected with engineered DNA (15, 291) and possess an extended survival after transplantation (461) together with the inherent ability to differentiate into several cell classes. The clinical application of MSCs for the regeneration of cartilage and bone is a good example of MSC safety and efficacy (40, 46). Clinical trials based on the therapeutic potential of MSCs have been conducted or are in progress throughout the world; MSCs have been injected intravenously in children with a genetic disorder of the bone, osteogenesis imperfecta (207–209). This was the first study demonstrating that bone marrow MSCs engraft outside of the organ of origin and produce a beneficial effect (208). Recently, autologous MSCs have been given to patients affected by cancer in an effort to enhance hematopoietic recovery during chemotherapy (242). Reports in animals have shown that bone marrow MSCs home to the heart and promote regeneration of the infarcted myocardium in different species (434, 483), possibly involving a paracrine effect (169, 291). Whether MSCs will be implemented in the management of ischemic heart failure in humans is difficult to predict, but it is a likely possibility. Autologous MSC-derived myogenic cells (321) and autologous undifferentiated MSCs (424) have been emPhysiol Rev • VOL ployed for cardiac repair after infarction or ischemiareperfusion injury in pigs. Cells were injected directly in proximity or within the injured myocardium, acutely or chronically after the ischemic event, and the effects of these interventions were determined several weeks later. In all cases, myocardial regeneration was documented and the reconstitution of dead myocardium correlated with the improvement in ventricular function and reappearance of wall motion activity (321, 424). Moreover, cotransplantation of human MSCs and human fetal cardiomyocytes resulted in an amelioration of cardiac performance greater than with MSCs alone (311). Formation of myocytes and coronary vessels has also been observed in rats after cryoinjury and MSC implantation (355). However, the intracoronary delivery of MSCs after infarction led to the differentiation of MSCs into fibroblasts in the scarred region and to the regeneration of myocytes in the surviving, unaffected portion of the ventricular wall (506). Importantly, in the absence of injury, MSCs engraft into the myocardium but remain in a viable quiescent state and do not participate in the physiological turnover of myocytes as well as vascular smooth muscle and endothelial cells. Thus the microenvironment significantly conditions the developmental pathway of MSCs in vivo. D. Controversy on Bone Marrow Cell Transdifferentiation and Cardiac Repair Stem cell transdifferentiation in the adult organism is a highly questioned mechanism of growth (9, 10, 150, 178, 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 3. Multipotent adult progenitor cells (MAPCs). MAPCs are isolated from the bone marrow and have the potential to differentiate in many different cell types in vitro and in vivo. However, MAPCS give rise to a more limited number of somatic cells when injected in vivo in the adult mouse than in the early blastocyst. 1383 ADULT STEM CELLS Physiol Rev • VOL cardium to a modest recovery of ventricular performance in the absence of tissue regeneration. The lack of beneficial consequences on the heart has also been reported. However, predominant observations have been positive supporting the feasibility of this form of cell therapy for the failing heart. Although these experimental results and the therapeutic efficacy of BMCs in patients with ischemic and nonischemic heart failure were indicative of BMC transdifferentiation (Fig. 4), the plasticity of BMCs has been questioned. Recent publications in mice have emphasized negative results, criticizing the documentation that BMCs can convert into cardiac cell lineages. The ability of BMCs to regenerate dead myocardium in the mouse heart was questioned, and the claim was made that the fate of the injected cells was to acquire only the hematopoietic lineages (45, 324). An additional report suggested that the engraftment of BMCs in the damaged heart is transient and hematopoietic in nature. BMC-derived cardiomyocytes were observed at low frequency and only outside the infarcted myocardium (336). Six possibilities can account for these contrasting results: 1) differences in the experimental protocol, 2) differences in the methodology of detection of newly formed structures, 3) differences in the viability of the injected cells, 4) artifacts due to autofluorescence and erroneous interpretation of immunolabeled structures, 5) relevance of animal models of parabiosis and repopulation of ablated bone marrow with an ideal HSC, and 6) a combination of these factors. We will discuss each of these variables. The intramyocardial delivery of BMCs in the mouse heart is complex and faced with difficulties dictated by the extremely high heart rate and very thin LV wall. Laboratories with years of experience in inducing coronary artery occlusion, coronary artery narrowing, or ischemia-reperfusion injury in mice are well aware that the successful imposition of myocardial infarction is ⬃80%, with ⬃20% unsuccessful surgery (115, 228, 356, 357, 539). Additionally, in positive cases, infarct size can vary from ⬃20 to ⬃90% of the entire LV inclusive of the interventricular septum. Infarcts involving ⬃50 – 60% of the LV carry a nearly 50% mortality within 24 – 48 h after surgery (33). Even more problematic is the recognition of a successful administration of BMCs within the LV myocardium, ⬃40 –50%, and the identification of the site of injection in proximity of the infarcted tissue (228, 356). Because of these unpredictable factors, methodologies have been developed in which rhodamine-labeled polystyrene spheres are mixed with BMCs to have a reliable reference point for the recognition of the actual positioning of BMCs in the LV wall and with respect to the infarct. When this simple protocol was applied, myocardial regeneration was demonstrated in 100% of mice, which had rhodamine particles and BMCs implanted in 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 206, 253, 279, 384, 392, 438, 473, 487, 502, 507, 508, 518). The most versatile cell is the bone marrow progenitor cell. The documentation that adult BMCs are capable of generating mature cells beyond their own tissue boundaries has provided an unexpected and powerful new form of cell therapy. The wave of enthusiasm created by this discovery and the lack of effective new drugs for the treatment of heart failure has prompted cardiologists to the rapid implementation of BMCs in the management of the infarcted human heart. Currently, bone marrow cells or circulating bone marrow cells, including endothelial progenitor cells (EPCs), are utilized in patients, and several initial clinical trials have been performed (39, 69, 158, 163, 273, 372, 373, 415, 421, 448, 449, 452, 528). Because of the compelling need to seek new treatments for severely ill patients, clinicians are leading the field of cell therapy and myocardial regeneration. Conversely, statements of caution have been made concerning the necessity to acquire a better understanding of the mechanisms of recovery of cardiac function before these protocols are introduced in the treatment of the diseased human heart. The fundamental question to be addressed is whether this more conservative position is justified because strong criticisms have been made not only against the notion of stem cell plasticity (45, 95, 279, 324, 474, 507, 508, 518) but also against observations that have challenged the perennial view of the heart and the brain as postmitotic organs incapable of regenerating after birth (508). In fact, the revolutionary work on the brain and the heart that has led to the identification of neural stem cells (139, 500) and cardiac stem cells (51, 201, 293, 296, 338) has been attacked as inconclusive, methodologically incorrect (508), and, more recently, a collection of artifacts (45, 95, 324, 495). The early studies on BMCs and cardiac repair were followed by numerous reports in which different types of BMCs and modalities of interventions were employed for the treatment of the damaged heart (24, 26, 34). These cells of bone marrow origin included several subsets making it difficult to dissect the impact that each cell type had on the function and structure of the diseased heart. For example, lineage-negative c-kit-positive cells (228, 258, 356), lineage-negative c-kit-positive Sca-1-positive cells, c-kit-positive-Thy1.1-low-lineage-negative Sca-1-positive, long-term reconstituting HSCs (45), CD34-positive cells (545), mesenchymal-like progenitor cells (424, 434), EPCs (281, 282), mononuclear BMCs, and clonally expanded human multipotent stem cells (539) have been tested. Moreover, BMCs with the potential of restoring the dead myocardium have been mobilized from the bone marrow into the systemic circulation utilizing several cytokines; SCF, G-CSF, and SDF-1␣ have been administered alone or in combination in various animal species (1, 38, 162, 188, 334, 357, 462). The effects of these various modalities of therapy ranged from improvement in regional cardiac function together with the restoration of the dead myo- 1384 LERI, KAJSTURA, AND ANVERSA the border zone of the infarcted myocardium (228). Therefore, differences in the protocols employed in the administration of BMCs could account for the differences observed experimentally between positive studies of myocardial regeneration (228, 356, 539) and reports challenging the ability of BMCs to form myocytes and coronary vessels (45, 324). Moreover, the negative studies claim that cell transplantation was 100% successful, that a mortality of 8% occurred with infarcts of 60%, and that a Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 4. Myocardial regeneration by BMCs. A: newly formed myocytes (troponin I; red) created by the injection of mouse male EGFP tagged c-kit positive BMCs in a female infarcted mouse heart. Nuclei are stained by PI (blue). Laminin defines the boundaries of the cells (white lines). The male phenotype of the regenerated myocytes is demonstrated by the presence of the Y chromosome in nuclei (yellow dots). B: a significant fraction of new myocytes expresses EGFP in the cytoplasm (green). C: the merge of A and B shows the colocalization of troponin I and EGFP (yellow-green) in the male myocytes. normal LV end-diastolic pressure was found with infarcts commonly incompatible with life in this model (45). A critical issue in studies of myocardial regeneration mediated by transplantation of BMCs involves the morphological analysis of the heart. This is an important point because one of the reports questioning the ability of BMCs to form myocytes and coronary vessels utilized conventional light microscopy and immunoperoxidase staining to demonstrate the presence or absence of newly formed structures within the infarcted myocardium (324). This approach reflects the contention that conventional light microscopy is superior to fluorescence labeling and confocal microscopy (250, 467). The resolution in images obtained by light microscopy is markedly inferior to that provided by confocal microscopy (28). Additionally, only the very superficial layer of the section can be examined by light microscopy (Fig. 5), whereas the entire thick histological section can be viewed by confocal microscopy (391). At high-magnification light microscopy, the observer is faced with a blurred image with loss of detail and undefined cell boundaries. This inherent problem with light microscopy was recognized immediately when confocal technology became available (521). Clear examples were published documenting the impossibility of identifying microtubules, mitotic spindle, cytoplasmic proteins, and chromosomal structures by light microscopy of cultured cells. Conversely, these morphological details were apparent when the same cells were evaluated by confocal microscopy. The difference between epifluorescence microscopy or bright-field microscopy and confocal microscopy is magnified in tissue sections. In fact, focal depth in a 5- to 6-m tissue section examined by epifluorescence microscopy or bright-field microscopy is only 8%, while the focal depth in a 0.5-m optical section analyzed by confocal microscopy is 12.5-fold higher, i.e., 100% (29, 283; Fig. 5). Problematic is the decision whether frozen tissue samples or paraffin-embedded specimens have to be employed for the immunohistochemical analysis of the myocardium by confocal microscopy. Both preparations have their advantages and their limitations. Perfusion fixation and paraffin processing of the heart facilitates morphological studies but inevitably leads to the extraction of some proteins and RNAs complicating the interpretation of negative results. Conversely, frozen section immunolabeling preserves better antigenicity but compromises structural detail. For the injured heart in particular, frozen samples introduce artifacts affecting the quality of the sections and microscopic resolution. The infarct is rarely preserved in frozen sections, and this problem interferes with the recognition of the region of the ventricle being examined. Paraffin-embedded tissue sections result, at most, in an underestimation of specific immunostaining not in an overestimation and, therefore, differences in the technical 1385 ADULT STEM CELLS approach could explain why positive and negative results concerning BMC transdifferentiation have been obtained. Hearts studied immunocytochemically should be the same examined functionally and with routine histology. This was not the case in the negative studies (45, 324). Whether coronary ligation was unsuccessful or a small or large infarct was generated was not determined. This is critical, because of the complexity of the model and the difficulty of producing an infarct and a correct injection of BMCs. Moreover, the utilization of frozen sections may interfere with the recognition of small cells of the size of newly formed myocytes (115, 228, 258, 356). Double labeling for the identification of the transgene present in the administered BMCs and the detection of myocyte cytoplasmic proteins was never performed on the same section (324), and negative claims were based on only two animals (45) that were supposedly properly infarcted and Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 5. BMC transdifferentiation in the infarcted heart: light microscopy histochemistry and confocal microscopy immunofluorescence. A comparison is made between a section of 5– 6 m in thickness examined by light microscopy (A) obtained from Reference 324 and optical sections of 0.5 m analyzed by confocal microscopy (B). The image in A was used to demonstrate the lack of transdifferentiation of BMCs into myocytes. Conversely, the image in B documents the ability of BMCs to adopt the cardiomyogenic fate. The brown cells in A should correspond to EGFP-positive cells that were not stained by any myocyte marker and were considered adequate for the unequivocal demonstration of the absence of BMC transdifferentiation. There is no double staining in this preparation for EGFP and myocyte cytoplasmic proteins. In contrast, B illustrates newly formed myocytes, ⬃400 m3 in volume, that are positive for EGFP (yellow-green). These cells express GATA-4 in their nuclei (yellow), and connexin43 is detected at the cell boundary (white, arrows). Focal depth in A is 8% and in B is 12.5-fold higher, 100%. injected with cells comparable to those used in the positive studies (356). This limited cohort calls the statistical validity of this type of experimentation into question, an issue that has recently been recognized in a large number of published reports in Nature and Nature Medicine (496). When these problems are dealt with, myocardial regeneration mediated by BMC transdifferentiation can also be documented by immunolabeling of frozen sections of infarcted myocardium (539). The animal models of parabiosis with complete blood chimerism or with reconstituted bone marrow through the injection of a single EGFP positive HSC (45, 507, 508) have been introduced to question the ability of BMCs to acquire a cardiomyocyte lineage. The utilization of the irradiation protocol in studies aiming at the documentation of the transdifferentiation potential of adult stem cells has been criticized (4, 297). Irradiation represents a form of injury that complicates the interpretation of the results. Most importantly, it cannot be used as documentation of physiological trafficking of cells from the bone marrow to distant organs. In fact, bone marrow transplantation in mice affected by amyotropic lateral sclerosis typically shows cell engraftment in the central nervous system (CNS) shortly after the intravenous injection of BMCs. But this process decreases with time (110). Because the damage is not repaired and the CNS continues to send the same signals for eventual cell recruitment, the progressive decline in BMCs colonized to the CNS strongly suggests that homing of these cells occurred only during the phase of high concentration of repopulating cells in the bloodstream. The administered BMCs targeted the depleted bone marrow, and their circulating pool markedly decreased with time. After the initial engraftment, there was no longer translocation of cells from the bone marrow to the CNS (110). Thus positive and negative results obtained with this protocol are difficult to interpret. The limitation of the therapeutic potential of circulating BMCs is not new. If circulating BMCs had the ability to spontaneously repair damaged organs, infarcts of the heart, brain, skin, kidney, and intestine would be easily reconstituted and the majority of current human diseases would not exist. These models are of little value to resolve the controversy at hand and can only add confusion to the confusion. The issue in need of resolution is whether BMCs injected directly in the infarct, in the border zone or systemically, differentiate into cardiac cell lineages and contribute to myocardial regeneration. The paradigms offered by the models of parabiosis and repopulated bone marrow with single cell injection are nonphysiological. More than 90% of the animals die, and the question is how relevant is an experimental procedure that represents the exception rather than the rule. Intrinsic genetic markers have been proposed as today’s gold standard for these types of studies (45, 95, 218, 1386 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL to false collection of data and erroneous interpretations and conclusions. Collectively, the data available so far demonstrate that myocardial regeneration occurs and reestablish the plasticity of BMCs and their ability to acquire a phenotype different from the organ of origin. Whether the process of cardiac repair occurs independently from the formation of synkaryons and heterokaryons or requires fusion of the progenitor cell with a preexisting partner cell is discussed in section IV.B of this review. III. CARDIAC PROGENITOR CELLS A. Identification Traditionally the heart has been viewed as a static organ incapable of repairing any form of damage. According to this paradigm, the number of myocytes is established at birth, and this population of terminally differentiated myocytes is irreplaceable throughout the life of the organ and the organism (7, 97, 288, 329, 338, 339, 354). Myocyte aging is dictated by the age of the organism, and myocytes can only increase in size or die. In agreement with this notion of the heart, myocytes are permanently lost, and the heart has no reserve mechanism that compensates for cell death and the wear and tear resulting from the physiological demands of daily life. There are men and women 100 years old and older, and according to this paradigm, all of their myocytes would have lived 100 years or more. In other words, the age of the individuals and the age of their myocytes should coincide, and myocytes would survive by continuously replacing intracellular organelles, leaving intact the viability of the cells. The demonstration that small amplifying myocytes can divide after infarction (54, 229) or pressure overload (30, 497) in humans has been vigorously challenged, and criticisms varied from technical errors in the identification of replicating cells (443) to the lack of relevance of the findings (329). In spite of the evidence of myocyte regeneration (27), the concept of the heart as a postmitotic organ is still widely accepted. The static view of the myocardium implies that myocyte death and regeneration have little role in cardiac homeostasis. Although stem cells have been identified in several organs including the blood, skin, central nervous system, liver, gastrointestinal tract, and skeletal muscle (59), the search for a CSC was perceived as a futile effort given the accepted lack of regenerative potential of the myocardium. However, data challenging this belief began to accumulate (63, 322, 391). In the past several years, the existence of cycling ventricular myocytes in the normal and pathological heart has been published (21, 29, 54, 229, 391, 497). Although these data provided an alternative understanding of cardiac homeostasis, they also raised 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 324, 495). Although the detection of the Y chromosome and EGFP protein in the regenerated myocytes (228, 356) falls within this category, recent experiments were performed with a genetic marker consisting of a cardiomyocyte-restricted transgene. Donor c-kit-positive BMCs were obtained from male transgenic mice carrying a cmyc tagged nuclear Akt under the control of the cardiac specific ␣-myosin heavy chain promoter (430). BMCs from these animals were injected acutely after infarction in wild-type female mice, and myocardial regeneration within the infarct was identified a few days later. The band of formed myocardium contained small myocytes that expressed ␣-sarcomeric actin, troponin I, ␣-actinin, connexin43, and N-cadherin. In all cases, the nuclei of these developing myocytes were positive for the c-myc tag (Fig. 6A). Vascular structures and CD45 labeled cells in the reconstituted tissue were negative for the c-myc tag. The Y chromosome was detected in endothelial and smooth muscle cell nuclei but not in CD45 positive cells. Therefore, BMCs injected in the dead myocardium do not adopt the hematopoietic fate (45) but assume the cardiac phenotype and repair the infarcted heart. Recently, a new technology was implemented. By this novel approach, antibodies are labeled by small semiconductor particles termed quantum dots. These particles are 10 nm in diameter and have a unique property, which is dictated by quantum mechanics of electrons confined to a small space (11, 71). The excitation wavelength of quantum dots is widely separated from the emission wavelength. The common site of excitation by confocal microscopy using blue diode laser is 405 nm, while the emission wavelength of quantum dots varies and the most frequently used have emissions between from 525 to 705 nm. The spectrum of autofluorescence under this setting totally escapes the emission wavelength; autofluorescence ranges between 440 and 520 nm. The advantage of this approach is remarkable; it is impossible to detect autofluorescence of tissue sections using this combination of excitation and emission wavelengths (Fig. 6B). Conversely, a minimal level of autofluorescence cannot be avoided when standard fluorochromes such as rhodamine or fluorescein are used. However, the degree of autofluorescence is always much lower than the emission peak. By employing primary and secondary antibodies conjugated directly with distinct quantum dots, the ability of BMCs to regenerate infarcted myocardium was confirmed in the absence of autofluorescence. Importantly, this protocol was combined with a gene reporter assay (Fig. 6C). The emphasis placed on genetic markers for the “objective” recognition of newly formed structures is only in part correct. Any genetic marker requires its subsequent identification by histochemical (218, 324) or immunocytochemical (45, 218) procedures. If limitations exist in these protocols, then the powerful genetic markers lead 1387 ADULT STEM CELLS Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 6. BMC transdifferentiation in the infarcted heart by quantum dot (Qdot) analysis. A: confocal microscopy with organic fluorescent dyes: BMCs from transgenic mice overexpressing c-myc tagged nuclear Akt, driven by the ␣-myosin heavy chain promoter, formed myocytes (␣sarcomeric actin; red) in the infarcted mouse heart. Nuclei are stained by DAPI (blue). Laminin defines the boundaries of the cells (green lines). The donor origin of the regenerated myocytes is demonstrated by c-myc in nuclei (yellow speckles). B: the excitation and emission wavelengths of the semiconductor particles Qdot 655 are distinct from the autofluorescence wavelength of tissue sections. This cannot be accomplished by organic fluorochromes (TRITC). The dotted lines indicate the excitation wavelength for blue diode laser and Qdots, 405 nm, or for the argon laser and TRITC, 568 nm. C: a field of myocardial regeneration comparable to that shown in A is illustrated in C by Qdot labeling of antibodies. The new myocytes are recognized by ␣-sarcomeric actin conjugated with Qdot 655 (red), c-myc in nuclei by Qdot 605 (yellow speckles), and laminin by Qdot 525 (green lines). Nuclei are stained by DAPI (blue). Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org 1388 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL organ, the replenishment of its parenchymal and nonparenchymal cells is regulated by a stem cell compartment and by the ability of these primitive cells to selfrenew and differentiate. Regeneration conforms to a hierarchical archetype: slowly dividing stem cells give rise to highly proliferating, lineage-restricted progenitor cells, which then become committed precursors that, eventually, reach growth arrest and terminal differentiation. Although stem cells have been known for quite some time, there is little understanding of the significance of stem cell surface antigens in the growth and differentiation potential of these cells (76, 113, 246). These limitations are not restricted to the heart and the CSC but apply to stem cells in all self-renewing organs including the bone marrow. Moreover, the surface antigens c-kit, MDR1, or Sca-1 are present in cells undergoing lineage differentiation, complicating recognition of the actual primitive cells in the population. FACS analysis is extremely helpful in separating stem cells in various categories according to the expression of one or more surface epitopes (101, 146, 481), but the distinction between undifferentiated and early committed cells necessitates caution. Sophisticated analyses of nuclear and cytoplasmic proteins can be performed by FACS, but they require fixation of the cells to make them permeable and amenable to the detection of intracellular components (174, 192, 255). However, the viability of the cells is lost precluding any subsequent in vivo or in vitro study. Similar problems are encountered by immunocytochemistry that also demands cell fixation before the antibody or a cocktail of antibodies is utilized to determine the stemness or commitment of the cells (305, 318, 447). The first attempt to describe an endogenous myocardial stem cell was made by Rudnicki and co-workers (201), who isolated a Hoechst effluxing cell population from the heart of 2-mo-old mice. Similar cells, which express MDR1 or a comparable ABC transporter, have been identified in other organs, and they correspond to the so-called side population (37, 73, 172, 299, 328, 546). These putative CSCs are negative for markers of hematopoietic cells including CD34, c-kit, Sca-1, Flk-2, and Thy1.1. Additionally, they give rise to colonies in methylcellulose medium and, in coculture with mature myocytes, differentiate into cardiac muscle cells expressing connexin43. Their pool size is markedly reduced in hearts of mice overexpressing a dominant negative form of MEF2C, suggesting that under this stressful condition there is a massive recruitment and commitment of these primitive cells to the myocyte lineage. These Hoechst effluxing cells also display a robust capacity to form hematopoietic colonies and to fuse with skeletal myoblasts in the absence of cell transdifferentiation (201). Although the characterization of these primitive cells as true resident CSCs remained incomplete, this work introduced the concept of a myocardial stem cell pool that 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 questions concerning two apparent contradictory lines of evidence: the well-documented irreversible withdrawal of cardiac myocytes from the cell cycle soon after their formation and the presence of cycling myocytes undergoing mitosis and cytokinesis. These results raised the question of the origin of dividing myocytes and their increase in response to an acute or chronic overload, pointing to the presence of a CSC. The pool size of adult stem cells is most likely set during prenatal life or early after birth, and stem cell migration, proliferation, maturation, and death regulate the homeostasis of the adult heart. Stem cells divide rarely while committed transient amplifying cells are the actual group of replicating cells. The less primitive amplifying cells possess a unique property; they undergo rounds of doublings and simultaneously differentiate (431, 516). The most relevant observation that dramatically challenged the old paradigm of the heart as a postmitotic organ was the identification of male cells in female hearts transplanted in male recipients (391). In these cases of sex-mismatched cardiac transplants in humans, the female heart in a male host had a significant number of Y-chromosome positive myocytes and coronary vessels. Although discrepancies existed among groups in terms of the degree of cardiac chimerism (116, 168, 322, 391, 471, 472), these results raised the possibility that these male cells colonized the female heart and subsequently differentiated into myocytes and functional vascular structures. The presence of male cells in the female heart was consistent with the contention that stemlike cells can migrate to the cardiac allograft and give rise to the three main cardiac cell progenies: myocytes, smooth muscle cells, and endothelial cells. Importantly, primitive cells of donor and recipient origin that expressed the stem cell surface antigens c-kit, Sca-1-like, and MDR1 were identified. Of relevance, identical cells were found in human control hearts (391). Since in fetal life, c-kit-positive cells colonize the yolk sack, liver, and probably other organs, and the colonized organs express SCF, the ligand of the c-kit receptor (249, 470), it was reasonable to assume that stemlike cells are present in the heart from fetal life. Additionally, the rapid induction of SCF during myocardial ischemia (152) strengthened the notion that SCF was involved in the activation of resident primitive c-kit-positive cells and, thereby, in the increased formation of myocytes in the acutely infarcted heart (54). The presence of these undifferentiated cells together with early committed progenies was suggestive of a true CSC as the critical modulator of the homeostasis of the normal and stressed myocardium. These observations were the foundation for the work that ultimately led to the identification and characterization of a resident CSC pool in the adult heart (51). A new, more biologically interesting model of cardiac growth, aging, and death was emerging. If the heart is a self-renewing 1389 ADULT STEM CELLS adapts in response to growth modifications. These observations have been confirmed (293). The c-kit-positive CSC was the first stem cell identified in the rat heart and, up to date, the c-kit-positive CSC is the most extensively characterized. These CSCs do not express transcription factors or membrane and cytoplasmic proteins of bone marrow cells, neural cells, skeletal muscle cells, or cardiac cells and are, therefore, lineage negative cells (51). Cardiac lineage negative c-kit-positive cells possess the three properties of stemness: clonogenicity, self-renewal, and multipotentiality (Fig. 7). In vitro, these cells grow as a monolayer when seeded in substrate-coated dishes or form spheroids when cultured in suspension, mimicking the biological behavior of neural stem cells (82). Transplantation of clonogenic c-kit-positive cells in the infarcted ventricle confirmed the in vitro characteristics of this cell population. This intervention led to engraftment, migration, proliferation, and multilineage differentiation of CSCs resulting in the replacement of dead tissue with new functional myocardium (51). When this class of CSCs is delivered intracoronarily after ischemia reperfusion injury, it promotes myocardial repair, limits infarct size, attenuates ventricular remodeling, Physiol Rev • VOL and ameliorates cardiac function (115). Thus c-kit-positive CSCs are effective when administered in a clinically relevant manner, pointing to an approach in which the heart’s own stem cells could be collected, expanded, and stored for subsequent therapeutic repair. Recently, CSCs have been defined as undifferentiated cells that express on the membrane the stem cell-related antigens, c-kit, MDR1, and Sca-1, in variable combinations (277). CSCs sorted separately for each of the stem cell antigens appear to have comparable growth and differentiation behavior in vitro. Whether the three classes of CSCs have the same potential in vivo is currently unclear. MDR1 belongs to the class of ABC transporters that mediate the Hoechst dye efflux in the side population of the bone marrow (73, 74) and skeletal muscle (172, 289). ABCG2 represents a more specific marker of this cell population, since mice defective for ABCG2 do not possess SP cells in the bone marrow (547). MDR1-ABCG2positive cells in the bone marrow give rise to the myeloid, lymphoid, and erythroid cell lineages (73) while in skeletal muscle they regenerate muscle fibers (184). Additionally, MDR1-ABCG2-positive cells from skeletal muscle are of bone marrow origin and can repopulate a depleted 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 7. Cardiac stem cells (CSCs). A: large clone of c-kit positive cells (green) generated by a single rat c-kit positive CSC. B–D: clonogenic cells in differentiating medium acquire the myocyte (B; ␣-sarcomeric actin, red), smooth muscle cell (C; ␣-smooth muscle actin, magenta), and endothelial cell (D; von Willebrand factor, yellow) phenotype. 1390 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL atria, and outflow tract have been linked to the homozygous deletion of Isl1 (77). The expression of Isl1 corresponds to the onset of myocyte commitment since Isl1, together with GATA-4, is a transcriptional activator of the myocyte transcription factor MEF2C (121). Recently, the claim has been made that Isl1 positive cells are the “true” CSCs and are highly important for myocardial regeneration in the diseased adult human heart (260). Conversely, the expression of Isl1 in progenitor cells clustered in the niches or scattered throughout the atrial and ventricular myocardium of the adult mouse heart is, at best, extremely rare. In our hands, not a single convincing example has been found. Similarly, no Isl1 cells have been detected in the failing human heart. Although these initial results do not exclude the possibility that some Isl1 cells may be present in the adult myocardium, the functional impact of these cells on cardiac pathology, if it exists, is debatable. Even during development, Isl1 cells are not implicated in the formation of the left ventricle. Myocardial repair requires the formation of myocytes and coronary vessels, and it cannot be accomplished by a cell already committed to the myocyte lineage. In the presence of an infarct, the generation of myocytes alone cannot restore contractile performance in the akinetic region; myocytes would not grow or survive in the absence of vessel formation. Resistance arterioles are critical for blood supply, and oxygen delivery is controlled by the capillary network. Similarly, the creation of vessels alone would not restore the dead myocardium or reinstitute contractile activity in the infarcted portion of the ventricular wall; vessels do not produce force. Myocardial regeneration necessitates the utilization of a more primitive multipotent cell such as the c-kit-positive CSC (51, 277). In vitro data suggest that the growth potential of c-kit-positive CSCs is greater than that of Sca-1-like or MDR1-positive cells, although these cell categories give rise to all cardiac cell lineages (277). B. Origin of Cardiac Stem Cells Following the recognition of a tissue-specific stem cell in the adult heart, the question concerned whether this primitive cell population originates, lives, and dies within the myocardium or whether other organs continuously replenish the heart with an undifferentiated stem cell pool that subsequently acquires cardiac characteristics. The bone marrow constitutes the main reservoir of primitive cells in the organism, and these cells can egress from the marrow niches, enter the systemic circulation, and chronically repopulate the heart and other organs (178, 253, 392, 473). However, this possibility has never been definitely proven. Under the condition of parabiosis and in the absence of myocardial injury, circulating HSCs do not home to the heart (45, 507, 508) or engraft efficiently in the bone marrow (300), 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 bone marrow (289). In humans (391, 497, 498) and animal models (277), MDR1-positive cells differentiate in progenitors and precursors of myocytes, endothelial and vascular smooth muscle cells, and fibroblasts. Sca-1 has been considered a marker of stem cells restricted to the mouse bone marrow (328, 431, 516). Contrary to expectation, Sca-1-like protein has been found by confocal microscopy and Western blot in primitive cells of the mouse, rat, dog, and human heart (277, 497, 498). Most likely, this epitope belongs to the large family of Ly6 proteins and corresponds to a known or unidentified member of these antigens. The recognition of this antigen by a Sca-1 antibody is not surprising given the high degree of homology among Ly6 proteins. Recent studies have indicated that Sca-1-positive progenitor cells are the predominant stem cell population in the mouse heart (296, 338). According to these observations, Sca-1-positive cells are 100- to 700-fold more frequent than c-kit-positive cells (51). Similarly, MDR1-positive cells have been found to represent a small fraction of all CSCs in the mouse heart (293). Comparable results about HSCs have been reported in the mouse bone marrow (219). Quantitative data in the mouse, rat, dog, and human heart from our laboratory have demonstrated that there is one CSC per ⬃30,000 – 40,000 myocardial cells: ⬃65% of all CSCs possess the three stem cell antigens (c-kit, MDR1, Sca-1-like); ⬃20% two stem cell antigens and ⬃15% only one. Approximately 5% each of all CSCs express exclusively c-kit, MDR1, or Sca-1 (277, 498). However, this work did not resolve the issue of whether the variability in the distribution of these surface markers actually reflects cells with distinct functional import. Some information can be obtained from the available transgenic mice: the Sca-1⫺/⫺ mouse and the Mdr1⫺/⫺ mouse (215, 416) have essentially a normal phenotype with modest defects of the hematopoietic system and other organs. In contrast to the deletion of Sca-1 or MDR1, mutation of the c-kit receptor in the W/WV mouse has profound effects on the phenotype of the animal. The W/WV mouse has a spontaneous point mutation in one allele coupled with an amino acid deletion in the other allele of the c-kit receptor (166, 333, 396). The homozygous W/W mouse dies shortly after birth, while the heterozygous W/WV has defects of the hematopoietic system with the development of anemia, melanocytes with loss of skin pigmentation, and mastocytes with altered immunoresponse (75, 476). The W/WV mouse reaches adulthood, and the heart shows alterations in cardiac anatomy and function at 8 –9 mo that precede the overt manifestations of anemia. Older W/WV mice develop anemia with painful skin lesions and behavioral abnormalities (94, 476). Nearly 5 years ago, the Isl1 transcription factor was shown to be associated with cardiac progenitor cells that condition heart morphogenesis in the mouse embryo (541). Defects in the development of the right ventricle, 1391 ADULT STEM CELLS Physiol Rev • VOL mented (302, 304). According to this theory, the primitive left ventricle and the outflow tract are derived from a single lineage, while the other regions including the primitive atria and the right ventricle are colonized by both lineages. The differential expression of transcription factors, Nkx2.5, Tbx5, Isl1, eHAND, and dHAND in distinct regions of the heart supports this possibility (364, 440). The common origin of cardiomyocytes was recognized by retrospective clonal analysis of the cells composing the mouse embryonic heart. Although clones of the founder cells appear in a temporal and spatial sequence, i.e., in different days of development and in distinct regions, the uniform genetic background of developing myocytes documented their derivation from a shared pool of founder cells. These precursors, which are ⬃140 at E8.5, participate in the formation of the entire length of the heart tube (302–304). Therefore, the initiation of separate transcriptional programs in the cells of the cardiac chambers (440) is an independent process and not the consequence of a diverse clonal origin from unipotent progenitors. To confirm this possibility, the EGFP gene was introduced at the stage of the morula-blastocyst transition (Fig. 8). EGFP was placed under the control of the desmin or ␣-cardiac actin promoter leading to the accumulation of EGFP in cells that express these myocyte-specific proteins (129). By this sophisticated technology, the patterns of myocardial histogenesis have been defined and the existence of an individual common progenitor of cardiomyocytes in prenatal and postnatal life has been demonstrated (129). Most importantly, this study has led to the unequivocal documentation of the intracardiac origin of adult myocytes and has excluded the possibility that a secondary immigration of stem cells from distant organs occurs in adulthood under physiological conditions (Fig. 8). Homing and engraftment of stem cells through the systemic circulation and the blood vessels would lead to a scattered localization of extra-cardiac-derived myocytes with small accumulation in the proximity of capillaries and arterioles (129, 391). However, single cardiomyocytes or miniclones distributed independently from the original embryonic EGFP-positive patches were never observed. Thus “late comers” do not contribute to the postnatal growth of the heart and, most likely, progenitor cells from other organs do not have a direct spontaneous role in myocardial homeostasis. C. Cardiac Stem Cell Niches Although stem cell antigens have been unequivocally detected in CSCs, there is no single marker capable of providing an absolute identification of stem cells in vivo. Stem cells are stored in niches that are located deep in the tissue for protection from damaging stimuli (157, 361, 362, 446, 491, 513). The niche constitutes the microenviron- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 supporting the notion that bone marrow homeostasis is primarily maintained by division of HSCs in situ (431, 516). In this regard, the model of heterochronic parabiosis (298, 464) has confirmed that rejuvenation of aged muscle mass depends on circulating molecules contained in the peripheral blood of young animals and not on young HSCs homed to old skeletal muscle (106 –108, 524). This argues against the bone marrow as the source of primitive cells in the heart. The contribution of bone marrow cells to cardiac chimerism has been proposed (247). Interestingly, a comparison has been made between the degree of chimerism in cardiac allografts and in hearts of patients who received allogeneic bone marrow transplantation (472). In the latter case, only 2–5% chimeric myocytes were detected, while 14 –16% of chimeric myocytes and endothelial cells were found in transplanted hearts. These observations suggest the intracardiac origin of the recipient cells in the donor heart and the extracardiac origin of chimeric cells in the resident heart following bone marrow transplantation. In the first case, host cells may have migrated from the residual atrial stumps to the donor heart (391) and, in the second, donor cells may have reached the myocardium because of the high level of blood chimerism (472). Thus blood-borne cardiac cells may be detected exclusively when the peripheral blood contains a large number of HSCs. Experimental results support this contention (217, 357). Whether HSCs represents the precursors of CSCs during the regenerative response of the damaged heart remains an unresolved issue. The striking discrepancy between the incidence of heart failure and bone marrow failure and the lack of comorbidity of these disease states in the same patient indicates that HSCs do not typically migrate from the bone marrow and repopulate the decompensated heart. If the bone marrow continuously replenishes the heart with new functionally competent HSCs, the decline in myocyte number with cardiac diseases would not occur, and the poorly contracting myocytes would be constantly replaced by a bone marrow-derived progeny. Current knowledge supports the notion that primitive cells are present in the heart during embryonic life and regulate heart morphogenesis and postnatal development. Organogenesis begins with the specification of a progenitor cell population (364). The pool size of embryonic cardiac progenitors conditions the final dimension of the organ. For example, retinoic acid reduces the number of prenatal cardiac progenitors leading to a hypoplastic heart (232). Heart morphogenesis is a complex process that results from the assembly of subsets of myocardial cells that exert contractile function but express different genes (432, 440, 520). Initially, the hypothesis was advanced that distinct unipotent progenitors give rise to the myocytes that populate the different parts of the heart (310). More recently, the early segregation of two lineages of myocytes from a common precursor has been docu- 1392 LERI, KAJSTURA, AND ANVERSA ment in which primitive cells divide, differentiate, and die. The recognition of stem cells within their natural milieu is of crucial importance; stem cells do not exist in the absence of supporting cells within the niche (446). The structure of a niche appears to be specifically tailored to suit the particular needs of its resident stem cells. In turn, stem cells may play an important role in the organization and specification of a niche (157). Niches share similarities in the activation of common signal transduction pathways that regulate the slow-cycling, self-renewing, undifferentiated state of their resident cells (12, 55, 83, 140, 202, 220, 223, 361, 446, 491, 513). However, niches in various self-renewing organs may differ in architectural organization and cellular composition that establish a tissue-specific microenvironment. Stem cells divide rarely, and cell replication is mostly confined to the niches (446). Stem cells undergo symmetPhysiol Rev • VOL ric or asymmetric division (141, 220, 223, 513). When stem cells divide symmetrically, two self-renewing daughter cells are formed, and the purpose of this mechanism of growth is the expansion of the stem cell compartment (513). When stem cells engage in asymmetric division, one daughter stem and one daughter amplifying cell are obtained. The objective of this division is cell differentiation, i.e., the production of a committed progeny. Stem cells can also divide symmetrically into two committed amplifying cells, decreasing the number of primitive cells (513). The developmental choice made by stem cells at any given time has a direct impact on the stem cell pool size, the number of progenitors and precursors and, ultimately, the number of mature cells (157, 251, 446, 513). The systematic identification of niches within an organ requires that stem cells be recognized together with the cells responsible for their anchorage to the niche 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 8. Heart histogenesis and mouse chimeras. The combination of two 8-cell morulas of different genetic origin (blue ⫽ LacZ; green ⫽ EGFP) gives rise to a chimeric morula that evolves into a chimeric blastocyst. The implantation of the early chimeric blastocyst in pseudopregnant mice results in balanced (equal parental contribution) or unbalanced (unequal parental contribution) mouse chimeras. By this approach, that is termed aggregation method, the number of progenitor cells that contribute to the development of the heart or other organs can be determined. The heart of these chimeric mice in which the two morulas carry either EGFP or LacZ originates from a single progenitor cell through a mechanism of coherent expansion of clones. In this case, patchy areas of the heart will be -gal positive (blue) and others EGFP positive (green). However, intermingling of clonogenic cells occurs and foci of -gal-positive cells (blue) are found in EGFP clonal patches (green) or vice versa. Limited intermingling of clonogenic cells and their progeny interferes partly with the coherent expansion of clonal patches. This pattern of embryonic heart growth is shown within the red box. The possibility that this model of cardiac growth is mediated by a large number of progenitors would require a massive loss of clones early in development, a phenomenon that cannot be excluded. If thousands of progenitors were implicated in embryonic cardiac growth, a mechanism of coherent expansion together with extensive intermingling of clonogenic cells would lead to a dispersed pattern of cardiomyocyte formation illustrated in the scheme. Importantly, the persistence of the embryonic clonal patches with small, intermingled foci in the adult myocardium excludes that new clonal regions (red foci) contribute to the growth of the heart postnatally as graphically represented in the red box. 1393 ADULT STEM CELLS Physiol Rev • VOL advantage with respect to the remaining endogenous CSCs. Stem cells in solid organs are dispersed in the tissue and, after their identification, it is critical to analyze the microenvironment where they reside and whether, upon activation, they can repair the damaged organ. Recently, CSC clusters have been found in the adult heart (22, 51, 98, 326, 498). Although CSC clusters are scattered throughout the myocardium, their distribution appears to be conditioned by the distinct levels of wall stress. In fact, the frequency of CSC clusters is inversely related to the hemodynamic load sustained by the anatomical regions of the heart: they accumulate in the atria and apex and are less numerous at the base and midportion of the left ventricle (Fig. 9). Physical forces, mechanical deformation, and high wall stress can be transduced in intracellular responses that regulate cell behavior and fate. In a fashion similar to chemical cues, i.e., cytokines and hormones, local forces modulate cell migration, proliferation, differentiation, and death (93, 140, 265, 407). CSCs nested in the atrial microenvironment may be implicated in the preservation of the CSC pool through symmetric division and the formation of self-renewing daughter cells. Conversely, the very high degree of stress at the base and midregion of the LV wall may condition the turnover of mature progeny and CSC commitment. Ventricular niches could be mostly involved in the formation of transient amplifying cells while apical niches may exert a dual function. Lineage negative cells expressing stem cell antigens, separately or in combination, have been detected, clustered together with cardiac progenitors and precursors. In close proximity to lineage negative cells, myocytes or other cells in an early stage of differentiation are commonly visible. The structural organization of these sites of accumulation of poorly differentiated cells differs from that described in the bulge region of the skin (65, 491, 513), seminiferous tubes of the testis (429), and gut crypts (386). In contrast, similarities in the spatial arrangement of candidate stem cells and committed cells can be found in the mouse bone marrow (540) and rat brain (122, 361). However, nurse cells in these organs have been characterized while they remain to be identified in the heart. Stem cells and supporting cells in the niches interact structurally and functionally through specialized gap and adherent junctional proteins (157). Connexins are gap junction channel proteins that mediate passage of small molecules and signals involved in cell-to-cell communication (170). Additionally, these gap junctions interfere with the activation, commitment, and migration out of the niches of stem cells (157, 317, 365). Survival factors and mitogens traverse gap junctions to oppose cell death and favor cell growth (Fig. 9). The identity of these molecules is largely unknown. However, Ca2⫹, ATP, adenosine, and cyclic nucleotides can translocate from one cell to another via gap junctions (170). Ca2⫹ influx or release from 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 structure, i.e., the supporting cells. Additionally, the existence of a progenitor-product relationship has to be established (157, 446). The direct physical interactions between stem cells and their non-stem cell neighbors in the niche are critical in keeping stem cells in this specialized compartment and in maintaining stem cell characteristics (35, 78, 276). The identification of a stem cell niche is a difficult challenge in mammals. This search is more readily accomplished in simpler organisms such as the Drosophila (276, 341, 446, 531). The precise architectural organization of germ cells in the Drosophila gonads simplifies the recognition of the niches (171, 441, 442, 532, 533, 549). In the fruit fly, there is a consistent reference point for the identification of the supporting cells. Mammalian stem cell niches have been documented in some organs. The better defined are those in organs that possess an epithelial lining, such as the skin niches in the bulge of the hair follicles and the intestinal niches in the crypts. In these cases, the supporting cells or nurse cells are the dermal papilla cells and the mesenchymal cells underlying the crypt (238, 491). The niche characteristics are applicable to all organs whether bone marrow, brain, or heart. The niches define the growth potential of an organ, and the recognition of their microenvironment is more important than the requirements postulated to be fundamental for the definition of stem cells. This is relevant because the criteria employed in the study of HSCs cannot be transferred to neural stem cells or CSCs without caveats. For example, the radiation protocol commonly used for lethal irradiation and bone marrow reconstitution would not be effective in the heart. In fact, the radiation dose required to reach and kill CSCs is 30 Gy. The need for 30 Gy is dictated by the structural and physical properties of the cardiac muscle (511). This very high dose, even though restricted to the heart, results in profound alterations of the entire organ, diffuse apoptosis, and death of the animals in congestive heart failure in 3–5 days. Also, these data demonstrate that the circulating pool of hematopoietic progenitor cells cannot replenish a depleted tissue such as the myocardium. The limitations inherent in the therapeutic potential of the circulating blood should not come as a surprise. If this were not the case, spontaneous regeneration would occur throughout the organism, and ischemic foci in all organs would be rapidly repaired. Models of parabiosis and bone marrow transplantation with a single ideal HSC (45, 324, 507) have little to contribute to our understanding of the human disease and the future impact of regenerative medicine (228). A similar comment can be made concerning the viewpoint that the “true” CSC has to be identified and a single CSC has to be shown to possess the ability to repopulate the depleted heart (95, 508). The heart cannot be ablated of its CSC population, and the injected single cell would have no competitive growth 1394 LERI, KAJSTURA, AND ANVERSA intracellular stores activates proliferation of neural stem cells. Also, Ca2⫹ is implicated in their differentiation and survival (406). The molecular glue that holds stem cells within the niches is represented by the cadherin family of proteins, which, together with -catenin, participate in the formation of specialized intercellular junctions, called adherens junctions (536). When differentiating stem cells lose contact with the neighboring cells, they proliferate more easily and change location within the niche. Cadherins are calcium-dependent transmembrane adhesion molecules (371, 536), which have a dual function; they anchor stem cells to their microenvironment and promote interaction between stem cells and between stem cells and supporting cells. Stem cell anchorage depends also on integrin receptors that bind cells to a basal lamina composed of extracellular matrix (222). Elevated levels of integrins are often characteristic of stem cells. Both integrins and adherent junctions play a critical role in the maintenance of Physiol Rev • VOL adhesiveness and quiescent status of stem and early committed cells within the organ (371). Thus the recognition of cardiac niches and the identity of the supporting cells constitute a major challenge for the definitive proof that the heart is a self-renewing organ in which cardiac homeostasis is regulated by a stem cell compartment (Fig. 9). The niches are expected to control the physiological turnover of myocardial cells and the growth, migration, and commitment of primitive cells leaving the niches to replace old dying cells in the myocardium. IV. CARDIAC PROGENITOR CELLS AND MYOCARDIAL REPAIR The recognition that the mammalian heart possesses a stem cell compartment that regenerates myocytes and coronary vessels raises the unique possibility to reconstitute dead myocardium after infarction, to repopulate the 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 9. Schematic representation of CSC niches. Cardiac niches are prevalently located in the atria and apex and consist of differentiated myocytes that surround clusters of CSCs and highly dividing amplifying cells. The amplifying cells are committed cells and express transcription factors of the cardiac (GATA-4), myocyte (MEF2C), smooth muscle cell (GATA-6), and endothelial cell (Ets1) lineages. The interaction among CSCs, early committed cells (ECCs), and supporting cells occurs via junctional proteins (cadherins and connexins). 1395 ADULT STEM CELLS Physiol Rev • VOL A. Myocardial Regeneration and Ischemic and Nonischemic Cardiomyopathy Myocardial regeneration occurs in humans following ischemic (52, 54, 229, 346, 498) and nonischemic injury (53, 229, 346, 348, 349, 497). However, myocyte proliferation appears to be restricted to the viable myocardium adjacent to and remote from the infarct or to areas of intact myocardium in other pathological states (497). The identification of CSCs is consistent with the notion that replicating myocytes constitute a subpopulation of rapidly growing amplifying cells originated from progenitors or more primitive cells. CSCs are distributed in all regions of the heart, suggesting that the reconstitution of parenchymal cells and coronary vessels can occur throughout the myocardium offering great potential for cardiac repair. CSCs located within the infarct or in its proximity could divide and differentiate reconstituting dead myocardium (Fig. 10). If this were the case, strategies can be implemented to enhance myocyte and vascular growthpromoting partial restoration of the infarct. This response would increase the number of myocytes and vessels, reduce infarct size, improve function, and decrease mortality. An identical argument can be made for other cardiac diseases associated with increases in pressure and/or volume loads on the heart (346, 348, 349, 497). Before discussing the role that CSCs may have in the treatment of the pathological heart, some comments have to be made concerning the complexity of the problem and the heterogeneity of cardiac pathology in the patient population that inevitably conditions the feasibility and efficacy of CSC therapy. Currently, it is difficult to anticipate how the etiology of heart failure and the unpredictable path of the disease influence the CSC compartment and, thereby, cardiac reserve. Ischemic heart disease and hypertension are the major causes of congestive heart failure (224, 274, 514, 551). However, myocardial infarction and high blood pressure lead to heart failure by mechanisms that are initially different, remain distinct during the progression of the pathological processes, and may become comparable only in the late evolution to terminal failure experimentally (345, 352, 353, 377) and in humans (54, 156, 179, 229, 344, 346, 348, 351, 497). Myocardial infarction results in acute ventricular decompensation (351, 359, 378) if the loss of mass and, thereby, of myocytes, reaches a critical value (Fig. 11). If cardiac dysfunction does not occur, the segmental loss of myocardium conditions ventricular remodeling and the clinical course of the disease (104). The postinfarcted heart typically shows cavitary dilation, wall thinning, decreased systolic ventricular pressure, and an increase in diastolic and systolic wall stress (29). Conversely, pressure overload hypertrophy induced by systemic hypertension is characterized by the preservation of chamber volume with thickening of the wall and 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 hypertrophic decompensated heart with new, better functioning myocytes and vascular structures, and, perhaps, to reverse ventricular dilation and wall thinning, restoring the physiological and anatomical characteristics of the normal heart. This hypothesis is supported by the identification of CSCs in the human heart (47, 497, 498). Additionally, CSCs injected locally in the infarcted myocardium of immunodeficient animals repair the necrotic tissue and improve ventricular function (47, 308). These observations have formed the basis of a new paradigm in which multipotent CSCs are implicated in the normal turnover of myocytes, endothelial cells, smooth muscle cells, and fibroblasts. Understanding the mechanisms of cardiac homeostasis would offer the extraordinary opportunity to potentiate this naturally occurring process and promote myocardial regeneration following tissue injury. Hypothetically, stimulation of CSCs is unlikely to induce malignant neoplasms since they are an extremely rare form of cardiac pathology (414). CSCs should be more effective in making new myocardium than progenitor cells from other organs. CSCs are programmed to create heart muscle and, upon activation, can rapidly engender parenchymal cells and coronary vessels possibly rescuing the failing heart (115, 391, 497, 498). However, the field of regenerative cardiology is in its infancy, and great caution has to be exercised in the implementation of this form of cellular therapy in humans before we have obtained the basic information concerning the ability of CSCs to migrate, divide, and differentiate. Similarly, ischemic and nonischemic pathological states may have profound and distinct implications for the function of the CSC pool (199, 498) including growth and lineage commitment. Unfortunately, there is no good animal model that can be employed to collect the information that is of tremendous importance in the application of CSC treatment to the patient population. In an attempt to develop strategies relevant to the future treatment of patients, new hypotheses have to be raised and tested to move the field in a direction that defines CSC therapy on an individual basis. This would maximize the efficacy of CSC administration in heart failure. Novel approaches have to be developed to impact on the future directions of regenerative cardiology. Negative experimental findings resulting from differences in protocols, methodological shortcomings, and suboptimal techniques for the analysis of myocardial repair have muddied the arena of regenerative cardiology. Therefore, the promoters of the field of stem cell therapy have to acknowledge these recent condemnations, repeat some of the work, and provide unequivocal answers to the criticisms. But mostly they have to continue to search for new and more innovative strategies for the reconstitution of the severely damaged pathological heart. 1396 LERI, KAJSTURA, AND ANVERSA an increase in wall thickness-to-chamber radius ratio. The hypertensive heart is faced initially by an increased systolic load only, because diastolic dysfunction is a minor alteration promoted by the increase in myocardial stiffness (176, 177, 551). The late stages of hypertensive hypertrophy when cardiac decompensation develops are characterized by an abnormal elevation in LV end-diastolic pressure, relative thinning of the wall, and expansion in chamber volume (29, 30). Myocyte death is scattered throughout the ventricle and frequently affects clusters of myocytes leading to the formation of foci of replacement fibrosis across the wall (20). Defects in coronary blood flow become apparent and involve increases in minimal coronary vascular resistance and reductions in coronary flow reserve (42). Abnormalities in myocardial perfusion are more prominent in hypertensive hypertroPhysiol Rev • VOL phy than in the postinfarcted heart (42, 230). Thus differences exist between these two forms of cardiac pathology, which may impact differently on the CSC pool. Ischemic heart disease, hypertension, idiopathic dilated cardiomyopathy, and the unsuccessful repair of a valvular defect with persistence of ventricular dysfunction and myocardial hypertrophy lead with time to severe ventricular decompensation. Although cavitary dilation is a common characteristic of these pathological states, the phenotypic architecture and loading of the heart vary significantly among these conditions and among patients in the same group. If we consider the evolution of the postinfarcted heart, the size of the infarct is a predictor of the short-, mid-, and long-term outcome of the disease in all patients (Fig. 11). But remodeling and accumulation of damage in the viable myocardium is the critical determi- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 10. Myocardial regeneration in the human heart. A: an area of spontaneous formation of myocytes (␣-sarcomeric actin, red) is present in proximity of dead, infarcted myocardium and a region of inflammatory reaction. B and C: metaphase chromosomes (PI, blue) are apparent in two small dividing myocytes (cardiac myosin heavy chain, red) in the heart of patients affected by chronic aortic stenosis. [Modified from Urbanek et al. (497).] 1397 ADULT STEM CELLS FIG. 11. Myocardial infarcts incompatible with survival. A and B: large transverse myocardial sections of paraffin-embedded tissue illustrating the left ventricle (LV), interventricular septum (IS), and right ventricle (RV). Tissue necrosis is present in a large portion of the IS and in some areas of the anterior aspect of LV (arrowheads). Foci of myocardial scarring are also detected (*). [Modified from Olivetti et al. (351).] It is remarkable that there is a persistent view that the formation of vessels alone can result in a significant restoration of performance of the failing heart (120). Minimal amounts of hibernating or stunned myocardium may be rescued by regional improvement of coronary blood flow, but this modest reinstitution of contractile activity in viable tissue cannot be confused with actual cardiac repair or myocardial regeneration (497, 498). The objective of cellular therapy is the creation of cardiomyocytes and vascular structures, which together replace dead or scarred myocardium, mending the broken heart. Whether healed infarcts or foci of replacement fibrosis contain a few vessels or a large number of vessels, ventricular function does not change. The increases in ejection fraction demonstrated in several clinical trials may very well reflect regeneration of myocytes and coronary vasculature, mimicking the results obtained experimentally (27, 51, 228, 258, 277, 539). There is a tight relationship between myocytes and the coronary vasculature, particularly the capillary network, with myocardial growth (31, 102, 393). This occurs during fetal and postnatal maturation of the heart as well as in conditions of accelerated growth induced by physiological overloads in the adult, such as dynamic exercise (32). The increased work demand may be counteracted by the activation of CSCs, which could generate new tissue with optimal quantities of myocytes and vessels. FIG. 12. Ventricular remodeling after myocardial infarction. Large transverse myocardial sections of paraffin-embedded tissue illustrating a healed myocardial infarct (HMI) with thinning of the wall (A) and multiple sites of replacement fibrosis (RF) in the noninfarcted viable tissue (B). [Modified from Beltrami et al. (52).] Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 nant of the onset of cardiac dysfunction and its progression to terminal failure (Fig. 12). The number of acute events varies in the patient population and, by the nature of the damage, segmental losses of myocardium necessitate approaches that are by far more complex than those required by minute areas of tissue injury or scattered myocyte death across the ventricular wall. This notion is based on observations obtained in humans, acutely after infarction (54, 498), or sustained pressure overload (497), and in animal models (51, 115, 227). The degree of differentiation that newly formed parenchymal cells acquire when they are clustered together in a large area of lost tissue markedly differs from that of regenerated myocytes scattered throughout the viable myocardium in close proximity to mature preexisting cells. In the former case, myocytes are small and the myofibrils are rare and occupy only a minimal portion of the cytoplasm resembling fetal-neonatal cells in humans (Fig. 10) and animals (Fig. 13). In the latter case, the regenerated myocytes are indistinguishable from the adjacent muscle cells and exhibit an adult phenotype (Fig. 13). This attractive biological problem raises interesting questions about the microenvironment and the cross-talk between differentiated and developing myocytes. Similarly, from a clinical standpoint, the reconstitution of infarcted myocardium with properties mimicking the structural organization of the normal heart is critical for the complete recovery of cardiac function. 1398 LERI, KAJSTURA, AND ANVERSA CSCs, progenitors, and precursors express c-Met, insulin-like growth factor I receptors (IGF-IR), and ANG II AT1 receptor subtype. Hepatocyte growth factor (HGF) activates c-Met promoting cell migration, growth, and survival (277). IGF-I has no influence on locomotion in vitro and in vivo but enhances cell proliferation, differentiation, and viability (277). ANG II induces multiplication of vascular cells and fibroblasts and hypertrophy of cardiomyocytes (148, 267, 278, 408, 409, 418). The HGF-cMet and IGF-I/IGF-IR systems oppose apoptotic and necrotic death signals while ANG II, through the AT1 receptor effector pathway, leads to the formation of reactive oxygen species (ROS) (93, 265). ROS, in turn, stimulates DNA damage and, ultimately, cell death (148). Stem cell surface antigens and growth factor receptor systems may operate together, condiPhysiol Rev • VOL tioning growth, death, and commitment of CSCs to distinct cell lineages. A critical component of CSCs is related to the ability of these cells to translocate upon stimulation. Three fundamental properties of CSCs regulate the cell turnover of the heart: migration, division, and differentiation. The fact that the myocardium is not a static organ and cell renewal is not restricted to endothelial cells, vascular smooth muscle cells, and fibroblasts but includes parenchymal cells, offers a reinterpretation of the biology of the heart and mechanisms of cardiac aging. Aging may promote alterations in the degree of CSC proliferation, maturation, and locomotion, attenuating cell replacement and the substitution of damaged cells and vascular structures. Although the acceptance of this concept finds resistance among traditional cardiologists and cardiovascular scien- 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 13. Myocardial regeneration after infarction in rats. A–F: the local intramyocardial injection of EGFP-tagged CSCs in syngeneic rats acutely after permanent coronary artery occlusion results in the formation of new myocardium. A–C: in the infarcted region, the regenerated myocytes are small (A; ␣-sarcomeric actin, red) and consistently show EGFP in the cytoplasm (B; green). C: the merge of A and B. The regenerated myocytes express both ␣-sarcomeric actin and EGFP (yellow-green). These myocytes are ⬃700 m3 in volume and, because of their size, cannot be the product of cell fusion. Nuclei are labeled by PI (blue). The localization of laminin at the periphery of the developing myocytes is illustrated by white lines. D–F: two newly formed myocytes (arrows) are detected within the surviving myocardium of the border zone. They are positive for ␣-sarcomeric actin (D; red) and EGFP (E; green). F: the merge of D and E. The new myocytes are indistinguishable from the adjacent myocytes, ⬃20,000 m3 in volume. 1399 ADULT STEM CELLS tists, recent information strongly supports this new view of the heart (484). B. Cell Fusion and Myocardial Regeneration Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 The notion of cell fusion, which was extremely popular in the 1980s, has recently been reintroduced successfully in the field of regenerative medicine (58, 60, 61, 368). Originally, cell fusion referred to the joining of two fully differentiated cells (60, 61). This phenomenon was frequently observed in the skeletal muscle and liver that are typical fusogenic organs (180, 182, 190, 454). The concept of cell fusion has evolved and, currently, indicates the coalescence of a stem cell and a differentiated cell (90, 111, 205, 501, 507, 508, 510). The merge of an adult stem cell and a committed cell results in the formation of a binucleated heterokaryon or a mononucleated hyperploid synkarion (58, 60, 61, 236, 368). The growth of the binucleated heterokaryon depends on the nucleus of the primitive cell that dominates the nucleus of the somatic cell by transferring its replication properties. However, the destiny of the heterokaryon is regulated by the nucleus of the differentiated cell (58, 60, 61, 153, 368, 384). When cell fusion is accompanied by nuclear fusion, a mononucleated hybrid cell with a hyperploid DNA content is formed. In this case, the bulky burden of the high nuclear DNA content leads to genetic instability and reduced or abrogated replicative potential (70, 130, 182, 190, 292, 384, 454). Cell fusion in mononucleated cells with 2C DNA content has been proposed and justified on the basis of reductive mitosis (510). Theoretically, diploid cells can result from hybrid cells that underwent reductive cell division converting the hyperploid cell into a diploid karyotype, which concealed fusion history (91, 376, 437, 526). Reductive cell division of hybrid cells in vivo has only been suggested in hepatocytes generated under stringent selection pressure that conferred survival and growth advantage on them (510). These “pseudo-diploid” cells accumulate slowly and might be present exclusively when a large number of fused cells are generated. The notion of myocardial regeneration by spontaneous or induced activation of CSCs as well as by transplantation or mobilization of progenitor cells from the bone marrow has been questioned, and the possibility that cardiac repair is not a primary event but a secondary process has been alleged (151). Fusion of a resident primitive cell or BMC with a preexisting myocyte that subsequently reenters the cell cycle and leads to the formation of a myocyte progeny has been proposed as an alternative mechanism of growth in the adult heart (14, 45, 151, 324, 336, 338) or other organs (14, 79, 80, 248, 301, 332, 501, 507, 508, 510, 522, 530, 545). If this were the case, spontaneous myocardial regeneration by activation of CSCs would be limited in scope and BMCs would not be able to differentiate across lineage boundaries and reconstitute damaged myocardium. Moreover, the turnover of cardiac cells would not be modulated by a stem cell compartment but by a complex biological process, that does not replace old dying myocytes or vascular smooth muscle cells and endothelial cells. Old cells would persist and somehow would constitute a necessary requirement for cell division to occur. Essentially, the engrained paradigm of the heart as a postmitotic organ comes back in a different manner, postulating once more that the life of myocytes corresponds to the life of the organ and the organism. The intrinsic capacity of the heart to heal itself would be negligible and of no biological and clinical importance. The wave of enthusiasm for cell fusion was prompted by two reports in 2002 that demonstrated the “extremely” rare occurrence of cell fusion under unusual in vitro settings that have no in vivo counterpart (469, 537). Bone marrow progenitor cells (469) or neural progenitors (537) were plated with ESCs. These coculture systems involved not only two types of undifferentiated cells but also highly stringent conditions favoring the occurrence of fusion events (135). In spite of this peculiar environment, cell fusion was occasional at best: 2–11 ⫻ 106 cells (469). The experimental design largely precluded the survival of ESCs that may have fused with the adult progenitor cells to acquire the antibiotic resistance gene necessary to remain viable under the selective in vitro protocol. Whether the formation of hybrid cells was dictated by the properties of the tissue-derived adult progenitor cells or ESCs was not determined (135). Surprisingly, this little evidence of cell fusion was used by skeptics of stem cell plasticity who questioned the “unexpected” regeneration of adult tissues including the brain, the liver, and the heart and proposed cell fusion as the only form of repair in adult organs (14, 79, 80, 248, 301, 332, 336, 338, 501, 507, 508, 510, 522). One-year later, mutant mice with an enzymatic defect in liver parenchymal cells were rescued by the injection of BMCs (501, 510), and the beneficial effect was totally attributed to fusion of BMCs with hepatocytes lacking the fumarylacetoacetate hydrolase (FAH). Four issues have been overlooked. First, the normal adult liver is a highly fusogenic organ (130, 182, 183, 454), such as the skeletal muscle (90, 130, 190, 205), and pathological states enhance the generation of polyploidy in both tissues (70, 173, 180, 182, 241, 433, 463). Second, the FAH mutant mouse has an unusually large number of hepatocytes with an abnormal karyotype, which facilitates cell fusion (135, 510). Third, 6 –15% of hepatocytes with normal DNA content were of donor origin, showing a female phenotype in male recipients (510). This finding is consistent with BMC transdifferentiation more than the complex, unproven suggestion of reductive mitosis. Fourth, the engraftment of BMCs and the incidence of cell fusion is very low in the liver of FAH mice, 1/150,000 cells (501, 510). However, 1400 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL role of progenitor cells. It is also unlikely that resident progenitor cells follow a growth pathway distinct from the intrinsic hierarchical process of commitment and differentiation. Additionally, the discovery that nanotubular highways are formed between cells could explain not only the translocation of enzymes between adjacent cells but also the migration of entire organelles from one cell to the neighboring cell (404). These nanotubules consist of Factin-positive structures that protrude from one cell to establish connection with another cell (164, 550). However, this phenomenon has been demonstrated in vitro, and its functional significance in vivo is unclear. Whether the twist in fate of a stem cell occurs by transdifferentiation or fusion, reprogramming of chromatin configuration is required (153, 237, 270, 271). The reorganization of chromatin is mediated by an alternate turning on and off of transcription factors to drive the adult stem cell towards the creation of a specific progeny (237). This process is slow and limited in efficiency (145, 150, 487). If cell fusion is involved, the replicative potential of the fused cell is at best transient (173, 180, 241). In organs such as the brain and the heart, cell fusion may lead only to a temporary rescue of an old preexisting terminally differentiated parenchymal cell. Heterokaryon formation in the Purkinje cells of the brain results in a short-lived rejuvenation of old or damaged cells, which is not associated with the acquisition of replicative potential (515). Nuclear reprogramming of transdifferentiated cells or synkaryons involves epigenetic modifications of chromatin (210, 394). Extrinsic signals present in the new environment drive chromatin remodeling. The principal mechanisms by which the expression pattern of tissuespecific genes or global gene silencing is established or maintained involve chromatin modification. This includes DNA methylation as well as histone acetylation, phosphorylation, methylation, and ubiquitinylation (161, 270). Dynamic changes in these epigenetic modifications underlie chromatin remodeling that is responsible for cell transdifferentiation or nuclear fusion (161, 210). To assess whether cell fusion plays a role in cardiac repair, myocardial regeneration mediated by the delivery of BMCs in mice (228, 356) and following CSC implantation in rats was analyzed (51). Since female rat clonogenic CSCs were injected in female infarcted rats and the probe for the rat X-chromosome was not available, we measured the number of chromosome 12 in the newly formed myocytes. In all cases, at most two chromosomes 12 were found excluding cell fusion (Fig. 14). Moreover, the DNA content was measured in regenerating myocytes and surviving myocytes. In all cases, a 2C DNA content was identified, and higher levels of DNA content reflected cycling myocytes. The absence of cell fusion was consistent with the number and size of newly formed myocytes within the band of regenerating myocardium (51). Importantly, the injection of male mouse BMCs in infarcted 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 this small number of fused cells grows rapidly under the selective pressure of the enzymatic deficiency, making this a particularly favorable situation for uncommon hepatocyte replacement (135, 510). The cre-lox genetic system is frequently used to detect cell fusion. Cre is a recombinase enzyme that cuts DNA segments flanked by binding sequences termed LoxP sites (6, 96, 403, 487). The cre-lox system can be generated by cross-breeding two transgenic mice or by implanting cells from a Cre mouse into a LacZ or EGFP mouse (6, 96, 260, 338, 403, 487). Importantly, a tissuespecific promoter drives the Cre recombinase. The Cre mouse is bred with a mouse carrying an inactive LacZ or EGFP gene placed under the control of a ubiquitous promoter in which a stop codon flanked by LoxP sites is inserted between the promoter and the LacZ or EGFP gene. The fusion of cells carrying the Cre recombinase with cells carrying the LacZ or EGFP gene leads to the binding of Cre to the LoxP sites. In the event of cell fusion, the Cre recombinase excises the stop codon or inhibitory segment resulting in the activation of LacZ or EGFP gene. Therefore, fused cells are labeled by -Gal or EGFP (487). More recently, a conditional Cre mouse has been developed in which Cre is regulated by the presence of the mouse estrogen receptor (mer-cre-mer). In this mouse, the activity of Cre is dependent on the administration of tamoxifen (260, 375), since mer is not activated by the endogenous 17-estradiol. A few studies have employed this approach to determine whether the injection of cardiac progenitor cells (260, 338) or BMCs (336) results in myocardial regeneration after infarction and whether tissue reconstitution is mediated by cell fusion. Myocyte formation was demonstrated with Sca1-positive cardiac progenitors while BMCs apparently failed to repair the damaged heart. In the former case, nearly 50% of new myocytes were considered the product of fusion while the remaining 50% were derived from differentiation of cardiac precursors (338). Moreover, evidence of fusion of BMCs with resident cardiomyocytes has been found in healthy hearts (336). However, the cre-lox system is not perfect. It is surprising that the possibility of metabolic cooperation was not considered, since this phenomenon may account for some of these observations (455). By metabolic cooperation, a cell may acquire the Cre recombinase from a neighboring cell and undergo excision of the floxflanked DNA segment in the absence of cell fusion. This exchange of the enzyme between the donor cell and the recipient cell occurs through intercellular junctions (366, 478, 503, 504). Metabolic cooperation is important in a tissue that is functionally a syncytium like the myocardium. The creation of myocytes by fusion of a cardiac progenitor with a fully mature myocyte cannot trigger the division of the recipient terminally differentiated cell. The hybrid cell loses the ability to proliferate abrogating the fundamental 1401 ADULT STEM CELLS female mice resulted in the generation of male myocytes with at most one X and one Y chromosome with a 2C DNA content (228, 356). Moreover, at 5 days, the regenerating band contained myocytes, which varied in size from 80 to 1,200 m3, with an average cell volume of 360 m3. Together, 2.7 million new myocytes were formed in 5 days after infarction. At 10 days, the regenerating band contained myocytes, which varied in size from 110 to 2,000 m3 with an average cell volume of 610 m3. Together, 4.3 million new Physiol Rev • VOL 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 14. Myocyte regeneration without cell fusion. A–C: formation of new myocytes following the injection of CSCs in an infarcted heart is identified by the expression of cardiac myosin heavy chain (A, red). Additionally, the number of chromosome 12 in the nuclei is determined by FISH (B; yellow dots). In all cases, at most two chromosomes 12 are identified in each nucleus, excluding cell fusion. C: the merge of A and B. The regenerated myocytes express ␣-sarcomeric actin and have one or two chromosomes 12 in their nuclei. Nuclei are labeled by PI (blue). The localization of laminin at the periphery of the developing myocytes is illustrated by white lines. myocytes were formed in 10 days after infarction. If cell fusion were the mechanism of myocyte regeneration, the formed cells should have a volume of at least 20,000 m3 or larger because the volume of resident myocytes is ⬃20,000 m3. Moreover, fusion of a primitive cell with a terminally differentiated myocyte would not be able to activate the cell cycle. In this regard, the number of myocytes formed markedly exceeded the number of myocytes remaining after infarction, further excluding cell fusion. Finally, the short interval between the injection of BMCs and the generation of diploid male cells makes reductive mitosis an unlikely possibility (190, 228, 356). When myocardial regeneration was induced by injection of clonogenic CSCs in infarcted mice or male BMCs were implanted in female infarcted mice, a question persisted. Permanent coronary artery occlusion in rodents leads to transmural infarcts leaving no viable partner cells for fusion (51). For this reason, a model of ischemiareperfusion injury in the rat was implemented, and EGFPpositive clonogenic CSCs were delivered through the coronary circulation (115). Again, a diploid DNA content was found and the new myocytes expressed at most two chromosomes 12. Thus cell fusion can be considered a negligible mechanism of cardiac repair whether CSCs or BMCs are employed as therapeutic tools (51, 115, 228, 258, 356). The occurrence of cell fusion in vivo appears to be extremely limited. Consistent with the observations in the heart, cell fusion in other organs including the skin, the lung, epithelial lining (190), and the brain (14, 204, 515) is restricted to a few cells, which, by inference, have no physiological consequences on baseline function or on tissue regeneration in pathological states. The discovery that resident progenitor cells exist in the adult myocardium overcomes the need for the complex and time-consuming process of nuclear reprogramming (384) and cell fusion (135). This is because primitive cells nested in the cardiac microenvironment are predetermined to evolve into cardiac cell lineages, i.e., myocytes and vascular smooth muscle and endothelial cells. Moreover, the heart possesses an intrinsic growth reserve capable of responding to the physiological and pathological demands of the myocardium. Importantly, resident stem cells may be more efficient and powerful for tissue reconstitution. These results introduce a new strategy for myocardial reconstitution expanding the notion that injection of exogenous cells into the damaged heart is required for the replacement of lost myocytes. The demonstration that the heart harbors stem cells capable of creating functional myocardium explains earlier observations of a robust regenerative response in the acute postinfarcted heart in humans (54), but raises the question of why this regenerative response stops before the repair process is completed. However, cardiac stem cells may be coaxed in vivo to home to the damaged region of the heart and promote the formation of functionally com- 1402 LERI, KAJSTURA, AND ANVERSA petent myocardium (277). This possibility offers an alternative or complementary therapeutic approach to exogenous cells. V. FUTURE DIRECTIONS Several fundamental areas of stem cell research have to be addressed in the near future. An important question involves whether distinct classes of human stem cells condition the efficacy of myocardial regeneration. Whether the expression of a single stem cell antigen or a combination of epitopes is linked to the formation of a preferential cardiac cell progeny is of paramount relevance for management of human disease. This recognition may allow us to develop powerful and novel strategies for the regeneration of myocytes and coronary vasculature and the restoration of cardiac performance in the failing heart. According to need, a new tool may become available for the predominant formation of myocytes and/or coronary vessels. Similarly, the impact of age, gender, type, and duration of the overload on stem cell proliferation and lineage commitment is unknown. Aging effects on stem cells may be comparable to those induced by a prolonged and sustained overload on the myocardium, and strategies may have to be developed to modify genetically the cells before treatment. A similar relevant question is whether stem cells in women are less susceptible to senescence and death signals, possessing an inherent ability to preserve their growth potential and to counteract the activation of the cell death pathway. In fact, myocyte death with aging (347, 350) and heart failure (179, 498) is reduced in women who have a higher nuclear Physiol Rev • VOL accumulation of the survival kinase phospho-Akt473 (81). These observations strengthen the notion that age, gender, and type and duration of the cardiac disease have a common effect on stem cells, i.e., senescence and death. Of relevance, culture conditions may promote aging of primitive and progenitor/precursor cells (424, 425, 428), pointing to the significance of the problem at hand for cellular therapy in humans. The same phenomenon may be operative in committed cells impinging on their regenerative growth. However, these negative modulators of the stem cell pool may affect only a subset of stem cells, while a fraction of cells retains its youth and, thereby, an intact ability to grow and give rise to a highly dividing committed progeny. Identical questions apply to CSCs and their potential future clinical application. In this regard, there is a relevant issue to be acknowledged, i.e., whether the use of undifferentiated CSCs is superior, inferior, or equal to the utilization of CSCs together with highly dividing early committed cells. These cells already express nuclear and/or cytoplasmic markers of the myocyte, endothelial, and smooth muscle cell lineages (51, 277, 497, 498). A pool of CSCs and partially differentiated cells may engender a greater and faster regenerative response. A mixture of cells, with clearly defined properties and with high potential for proliferation, may be the most effective in reconstituting dead myocardium after infarction. Additionally, this composite cell population may actively replace poorly contracting myocytes in the chronically failing senescent heart, with younger better functioning cells and vascular supply. Under this setting, coronary reserve is impaired, and minimal coronary vascular resistance is 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 FIG. 15. Tissue engineering and stem cells. The injection of a self-assembling bioengineered scaffold loaded with adult CSCs and selective growth factors may enhance myocardial regeneration and orderly organization of parenchymal cells and vascular structures. Moreover, the presence of growth factors may favor the recruitment of endogenous progenitor cells in the area of injury. 1403 ADULT STEM CELLS Physiol Rev • VOL 324). It might be relevant to reiterate that the current treatment of choice in heart failure consists of angiotensin converting enzyme (ACE) inhibitors and -blockers. These drugs improve quality of life and significantly decrease mortality. ACE inhibitors have been used for ⬃20 years (378 –380), and -blockers have been around for ⬃40 years (494). -Blockers were considered detrimental for the decompensated heart. Contrary to expectation, -blockers are the most powerful agent available to patients with congestive heart failure (468). But, how ACE inhibitors and -blockers interfere with the progression of heart failure remains to be identified. ACKNOWLEDGMENTS Address for reprint requests and other correspondence: P. Anversa, Cardiovascular Research Institute, New York Medical College, Vosburgh Pavilion, Rm. 303, Valhalla, NY 10595 (E-mail: [email protected]). GRANTS This work was supported by National Institutes of Health Grants HL-38132, AG-15756, HL-65577, HL-66923, HL-65573, AG17042, AG-023071, HL-43023, HL-50142, AG-026107, and HL-081737. REFERENCES 1. Abbott JD, Huang Y, Liu D, Hickey R, Krause DS, and Giordano FJ. Stromal cell-derived factor-1 alpha plays a critical role in stem cell recruitment to the heart after myocardial infarction but is not sufficient to induce homing in the absence of injury. Circulation 110: 3300 –3305, 2004. 2. Abdallah BM, Haack-Sorensen M, Burns JS, Elsnab B, Jakob F, Hokland P, and Kassem M. Maintenance of differentiation potential of human bone marrow mesenchymal stem cells immortalized by human telomerase reverse transcriptase gene despite extensive proliferation. Biochem Biophys Res Commun 326: 527– 538, 2005. 3. Abdullah I, Lepore JJ, Epstein JA, Parmacek MS, and Gruber PJ. MRL mice fail to heal the heart in response to ischemiareperfusion injury. Wound Rep Reg 13: 205–208, 2005. 4. Abedi M, Greer DA, Colvin DA, Demers MS, Dooner MS, Harpel JA, Pimentel J, Menon MK, and Quesenberry JP. Tissue injury in bone marrow transdifferentiation. Blood Cells Mol Dis 32: 42– 46, 2004. 5. Adzick NS and Lorenz HP. Cells, matrix, growth factors, and the surgeon. The biology of scarless fetal wound repair. Ann Surg 220: 10 –18, 1994. 6. Agah R, Frenkel PA, French BA, Michael LH, Overbeek PA, and Schneider MD. Gene recombination in postmitotic cells. Targeted expression of Cre recombinase provokes cardiac-restricted, site-specific rearrangement in adult ventricular muscle in vivo J Clin Invest 100: 169 –179, 1997. 7. Agah R, Kirshenbaum LA, Abdellatif M, Truong LD, Chakraborty S, Michael LH, and Schneider MD. Adenoviral delivery of E2F-1 directs cell cycle reentry and p53-independent apoptosis in postmitotic adult myocardium in vivo. J Clin Invest 100: 2722–2728, 1997. 8. Alhadlaq A and Mao JJ. Mesenchymal stem cells: isolation and therapeutics. Stem Cells Dev 13: 436 – 448, 2004. 9. Alison MR, Poulsom R, Otto WR, Vig P, Brittan M, Direkze NC, Preston SL, and Wright NA. Plastic adult stem cells: will they graduate from the school of hard knocks? J Cell Sci 116: 599 – 603, 2003. 10. Alison MR, Poulsom R, Otto WR, Vig P, Brittan M, Direkze NC, Lovell M, Fang TC, Preston SL, and Wright NA. Recipes 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 increased (29, 252). Thus new myocytes, arterioles, and capillaries may be rapidly developed to improve tissue oxygenation and cardiac pump function. Data in the infarcted rat heart suggest that clonogenic CSCs (derived from a single founder cell) and a mixture of CSCs and rapidly growing amplifying cells (derived from several founder cells) both result in a robust reconstitution of damaged myocardium (51). But the latter cell population appears to give rise to more mature myocytes and coronary vessels and to result in a faster recovery of wall thickness in the infarcted region. The committed cells may have a reduced capacity to proliferate but may acquire the adult phenotype more rapidly. Myocardial regeneration after infarction, whether promoted by BMCs, CSCs, or CSCs together with early committed cells, has only partly reconstituted large portions of dead myocardium. Moreover, the structural organization of the newly formed tissue resembles fetal-neonatal myocardium with areas of more advanced differentiation. The created myocytes are oriented longitudinally to the tangential stress lacking the more complex architecture of cardiomyocyte bundles within the ventricular wall. Perhaps, disruption of the extracellular network and/or the dramatic increase in diastolic and systolic stress at the site of the infarct opposes the restoration of the adult pattern of the myocardium. This limitation may be overcome by the use of bioengineered scaffolds that can drive an orderly growth of myocytes and coronary vessels (330). These self-assembling environments can be loaded with peptides enhancing progenitor cell homing and differentiation (245). Thus bioengineered biological devices and CSCs may have to be combined for a successful repair of the injured heart (Fig. 15). The crisis in federal funding may become severe, imposing difficult decisions. The dramatic clinical manifestations of heart failure together with the enormous increase in the number of patients affected by this devastating disease have inevitably influenced the role that clinicians have taken in the rapid implementation of cell therapy for the decompensated heart. A profound dichotomy exists between our limited understanding of the basic mechanisms that regulate stem cell growth, differentiation, and death during myocardial repair and the number of patients who received BMCs for restoration of the damaged heart. Double-blind clinical trials are ongoing, and new patients are enrolled daily in spite of the uncertainty about the pathways implicated in the beneficial effects of stem cell therapy. This attitude has been questioned, and strong statements against the application of cell therapy for heart diseases have been made (45). Understanding the science underlying the efficacy of stem cell therapy is important, but criticizing world-renowned cardiologists for offering a safe, new treatment to their severely ill patients is not justified. This is particularly true since negative findings were reported in mice (45, 1404 11. 12. 13. 14. 15. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. for adult stem cell plasticity: fusion cuisine or readymade? J Clin Pathol 57: 113–120, 2004. Alivisatos P. The use of nanocrystals in biological detection. Nat Biotechnol 22: 47–52, 2004. Allman D, Aster JC, and Pear WS. Notch signaling in hematopoiesis and early lymphocyte development. Immunol Rev 187: 75– 86, 2002. Almeida-Porada G, El Shabrawy D, Porada C, and Zanjani D. Differentiative potential of human metanephric mesenchymal stem cells. Exp Hematol 30: 1454 –1462, 2002. Alvarez-Dolado M, Pardal R, Garcia-Verdugo JM, Fike JR, Lee HO, Pfeffer K, Lois C, Morrison SJ, and Alvarez-Buylla A. Fusion of bone-marrow-derived cells with Purkinje neurons, cardiomyocytes and hematocytes. Nature 425: 968 –973, 2003. Andrades JA, Han B, Becerra J, Sorgente N, Hall FL, and Nimni ME. A recombinant human TGF-beta1 fusion protein with collagen-binding domain promotes migration, growth, and differentiation of bone marrow mesenchymal cells. Exp Cell Res 250: 485– 498, 1999. Andrews PW. From teratocarcinomas to embryonic stem cells. Philos Trans R Soc Lond B Biol Sci 357: 405– 417, 2002. Angoulvant D, Fazel S, and Li RK. Neovascularization derived from cell transplantation in ischemic myocardium. Mol Cell Biochem 264: 133–142, 2004. Anversa P. Myocyte death in the pathological heart. Circ Res 86: 121–124, 2000. Anversa P and Capasso JM. Loss of intermediate-sized coronary arteries and capillary proliferation after left ventricular failure in rats. Am J Physiol Heart Circ Physiol 260: H1552–H1560, 1991. Anversa P and Kajstura J. Myocyte cell death in the diseased heart. Circ Res 82: 1231–1233, 1998. Anversa P and Kajstura J. Ventricular myocytes are not terminally differentiated in the adult mammalian heart. Circ Res 83: 1–14, 1998. Anversa P, Kajstura J, Nadal-Ginard B, and Leri A. Primitive cells and tissue regeneration. Circ Res 92: 579 –582, 2003. Anversa P, Kajstura J, and Leri A. Circulating progenitor cells: search for an identity. Circulation 110: 3158 –3160, 2004. Anversa P and Leri A. Myocardial Regeneration. Braunwald’s Heart Disease. New York: Elsevier, 2005. Anversa P, Leri A, Kajstura J, and Nadal-Ginard B. Myocyte growth and cardiac repair. J Mol Cell Cardiol 34: 91–105, 2002. Anversa P, Leri A, Nadal-Ginard B, and Kajstura J. Stem Cells and Heart Disease. Handbook of Stem Cells: Adult and Fetal. New York: Elsevier, 2004, vol. 2. Anversa P and Nadal-Ginard B. Myocyte renewal and ventricular remodelling. Nature 415: 240 –243, 2002. Anversa P and Nadal-Ginard B. Cardiac chimerism: methods matter. Circulation 106: 129 –131, 2002. Anversa P and Olivetti G. Cellular basis of physiological and pathological myocardial growth. In: Handbook of Physiology. The Cardiovascular System. The Heart. Bethesda, MD: Am. Physiol. Soc., 2002, sect. 2 chapt. 2, p. 75–144. Anversa P, Palackal T, Sonnenblick EH, Olivetti G, and Capasso JM. Hypertensive cardiomyopathy: myocyte nuclei hyperplasia in the mammalian rat heart. J Clin Invest 85: 994 –997, 1990. Anversa P, Ricci R, and Olivetti G. Quantitative structural analysis of the myocardium during physiologic growth and induced cardiac hypertrophy: a review. J Am Coll Cardiol 7: 1140 –1149, 1986. Anversa P, Ricci R, and Olivetti G. Effects of exercise on the capillary vasculature of the rat heart. Circulation 75: I12–I8, 1987. Anversa P and Sonnenblick EH. Ischemic cardiomyopathy: pathophysiologic mechanisms. Prog Cardiovasc Dis 33: 49 –70, 1990. Anversa P, Sussman MA, and Bolli R. Molecular genetic advances in cardiovascular medicine: focus on the myocyte. Circulation 109: 2832–2838, 2004. Arai F, Hirao A, Ohmura M, Sato H, Matsuoka S, Takubo K, Ito K, Koh GY, and Suda T. Tie2/Angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell 118: 149 –161, 2004. Physiol Rev • VOL 36. Asakura A, Komaki M, and Rudnicki M. Muscle satellite cells are multipotential stem cells that exhibit myogenic, osteogenic, and adipogenic differentiation. Differentiation 68: 245–253, 2001. 37. Asakura A, Seale P, Girgis-Gabardo A, and Rudnicki MA. Myogenic specification of side population cells in skeletal muscle. J Cell Biol 159: 123–134, 2002. 38. Askari AT, Unzek S, Popovic ZB, Goldman CK, Forudi F, Kiedrowski M, Rovner A, Ellis SG, Thomas JD, DiCorleto PE, Topol EJ, and Penn MS. Effect of stromal-cell-derived factor 1 on stem-cell homing and tissue regeneration in ischaemic cardiomyopathy. Lancet 362: 697–703, 2003. 39. Assmus B, Schachinger V, Teupe C, Britten M, Lehmann R, Dobert N, Grunwald F, Aicher A, Urbich C, Martin H, Hoelzer D, Dimmeler S, and Zeiher AM. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction (TOPCARE-AMI). Circulation 106: 3009 –3017, 2002. 40. Awad HA, Butler DL, Boivin GP, Smith FN, Malaviya P, Huibregtse B, and Caplan AI. Autologous mesenchymal stem cell-mediated repair of tendon. Tissue Eng 5: 267–277, 1999. 41. Azouz A, Razzaque MS, El-Hallak M, and Taguchi T. Immunoinflammatory responses and fibrogenesis. Med Electron Microsc 37: 141–148, 2004. 42. Bache RJ. Effects of hypertrophy on the coronary circulation. Prog Cardiovasc Dis 30: 403– 440, 1988. 43. Baddoo M, Hill K, Wilkinson R, Gaupp D, Hughes C, Kopen GC, and Phinney DG. Characterization of mesenchymal stem cells isolated from murine bone marrow by negative selection. J Cell Biochem 89: 1235–1249, 2003. 44. Bader A, Gruss A, Hollrigl A, Al-Dubai H, Capetanaki Y, and Weitzer G. Paracrine promotion of cardiomyogenesis in embryoid bodies by LIF modulated endoderm. Differentiation 68: 31– 43, 2001. 45. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, and Robbins RC. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature 428: 668 – 673, 2004. 46. Barry FP. Mesenchymal stem cells therapy in joint disease. Novartis Found Symp 249: 86 –96, 2003. 47. Bearzi C, Cascapera S, Nascimbene A, Casarsa C, Rastaldo R, Hosoda T, de Angelis A, Rota M, Quaini F, Urbanek K, Leri A, Anversa P, Bolli R, and Kajstura J. Characterization and growth of human cardiac stem cells. Late-breaking developments in stem cell biology and cardiac growth regulation. Circulation 111: 1720, 2005. 48. Becker AJ, McCulloch EA, and Till JE. Cytological demonstration of the clonal nature of spleen colonies derived from transplanted mouse marrow cells. Nature 197: 452– 454, 1963. 49. Beddington RS and Robertson EJ. An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo. Development 105: 733–737, 1989. 50. Behfar A, Zingman LV, Hodgson DM, Rauzier JM, Kane GC, Terzic A, and Puceat M. Stem cell differentiation requires a paracrine pathway in the heart. FASEB J 16: 1558 –1566, 2002. 51. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, and Anversa P. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 114: 763–776, 2003. 52. Beltrami CA, Finato N, Rocco M, Feruglio GA, Puricelli C, Cigola E, Quaini F, Sonnenblick EH, Olivetti G, and Anversa P. Structural basis of end-stage failure in ischemic cardiomyopathy in humans. Circulation 89: 151–163, 1994. 53. Beltrami CA, Finato N, Rocco M, Feruglio GA, Puricelli C, Cigola E, Sonnenblick EH, Olivetti G, and Anversa P. The cellular basis of dilated cardiomyopathy in humans. J Mol Cell Cardiol 27: 291–305, 1995. 54. Beltrami AP, Urbanek K, Kajstura J, Yan SM, Finato N, Bussani R, Nadal-Ginard B, Silvestri F, Leri A, Beltrami CA, and Anversa P. Evidence that human cardiac myocytes divide after myocardial infarction. N Engl J Med 344: 1750 –1757, 2001. 55. Berdnik D, Torok T, Gonzalez-Gaitan M, and Knoblich JA. The endocytic protein ␣-adaptin is required for numb-mediated asymmetric cell division in Drosophila. Dev Cell 3: 221–231, 2002. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 16. LERI, KAJSTURA, AND ANVERSA 1405 ADULT STEM CELLS Physiol Rev • VOL 79. 80. 81. 82. 83. 84. 85. 86. 87. 88. 89. 90. 91. 92. 93. 94. 95. 96. 97. 98. 99. 100. 101. FR. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 425: 841– 846, 2003. Camargo FD, Finegold M, and Goodell MA. Hematopoietic myelomonocytic cells are the major source of hepatocyte fusion partners. J Clin Invest 113: 1266 –1270, 2004. Camargo FD, Green R, Capetanaki Y, Jackson KA, and Goodell MA. Stem cell plasticity: from transdifferentiation to macrophage fusion. Cell Prolif 37: 55– 65, 2004. Camper-Kirby D, Welch S, Walker A, Shiraishi I, Setchell KD, Schaefer E, Kajstura J, Anversa P, and Sussman MA. Myocardial Akt activation and gender: increased nuclear activity in females versus males. Circ Res 88: 1020 –1027, 2001. Campos LS. Neurospheres: insights into neural stem cell biology. J Neurosci Res 78: 761–769, 2004. Campos LS, Leone DP, Relvas JB, Brakebusch C, Fassler R, Suter U, and ffrench-Constant C. 1 Integrins activate a MAPK signaling pathway in neural stem cells that contributes to their maintenance. Development 131: 3433–3444, 2004. Caplan AI. Mesenchymal stem cells. J Orthop Res 9: 641– 650, 1991. Caplan AI and Bruder SP. Mesenchymal stem cells: building blocks for molecular medicine in the 21st century. Trends Mol Med 7: 259 –267, 2001. Castro-Malaspina H, Ebell W, and Wang S. Human bone marrow fibroblast colony-forming units (CFU-F). Prog Clin Biol Res 154: 209 –236, 1984. Castro-Malaspina H, Gay RE, Resnick G, Kapoor N, Meyers P, Chiarieri D, McKenzie S, Broxmeyer HE, and Moore MA. Characterization of human bone marrow fibroblast colony-forming cells (CFU-F) and their progeny. Blood 56: 289 –301, 1980. Chambers I and Smith A. Self-renewal of teratocarcinoma and embryonic stem cells. Oncogene 23: 7150 –7160, 2004. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, and Smith A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 113: 643– 655, 2003. Charlton CA, Mohler WA, Radice GL, Hynes RO, and Blau HM. Fusion competence of myoblasts rendered genetically null for N-cadherin in culture. J Cell Biol 138: 331–336, 1997. Chen A, Ekman JM, and Heathcote RD. Reduction in cell size during development of spinal cord. J Comp Neurol 409: 592– 602, 1999. Chen W, Fu X, Ge S, Sun T, Zhou G, Jiang D, and Sheng Z. Ontogeny of expression of transforming growth factor- and its receptors and their possible relationship with scarless healing in human fetal skin. Wound Repair Regen 13: 68 –75, 2005. Cheng W, Li B, Kajstura J, Li P, Wolin MS, Sonnenblick EH, Hintze TH, Olivetti G, and Anversa P. Stretch-induced programmed myocyte cell death. J Clin Invest 96: 2247–2259, 1995. Chi MM and Powley TL. c-kit mutant mouse behavioral phenotype: altered meal patterns and CCK sensitivity but normal daily food intake and body weight. Am J Physiol Regul Integr Comp Physiol 285: R1170 –R1183, 2003. Chien KR. Stem cells: lost in translation. Nature 428: 607– 608, 2004. Chien KR. To Cre or not to Cre: the next generation of mouse models of human cardiac diseases. Circ Res 88: 546 –549, 2001. Chien KR and Olson EN. Converging pathways and principles in heart development and disease. Cell 110: 153–162, 2002. Chimenti C, Kajstura J, Torella D, Urbanek K, Heleniak H, Colussi C, Di Meglio F, Nadal-Ginard B, Frustaci A, Leri A, Maseri A, and Anversa P. Senescence and death of primitive cells and myocytes lead to premature cardiac aging and heart failure. Circ Res 93: 604 – 613, 2003. Choi KS, Shin JS, Lee JJ, Kim YS, Kim SB, and Kim CW. In vitro trans-differentiation of rat mesenchymal cells into insulinproducing cells by rat pancreatic extract. Biochem Biophys Res Commun 330: 1299 –1305, 2005. Christoffels VM, Burch JB, and Moorman AF. Architectural plan for the heart: early patterning and delineation of the chambers and the nodes. Trends Cardiovasc Med 14: 301–307, 2004. Civin CI and Loken MR. Cell surface antigens on human marrow cells: dissection of hematopoietic development using monoclonal 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 56. Beresford WA. Direct transdifferentiation: can cells change their phenotype without dividing? Cell Differ Dev 29: 81–93, 1990. 57. Bhatia M. AC133 expression in human stem cells. Leukemia 15: 1685–1688, 2001. 58. Blau HM. How fixed is the differentiated state? Lessons from heterokaryons. Trends Genet 5: 268 –272, 1989. 59. Blau HM, Brazelton TR, and Weimann JM. The evolving concept of a stem cell: entity or function? Cell 105: 829 – 841, 2001. 60. Blau HM, Chiu CP, and Webster C. Cytoplasmic activation of human nuclear genes in stable heterocaryons. Cell 32: 1171–1180, 1983. 61. Blau HM, Pavlath GK, Hardeman EC, Chiu CP, Silberstein L, Webster SG, Miller SC, and Webster C. Plasticity of the differentiated state. Science 230: 758 –766, 1985. 62. Boheler KR, Czyz J, Tweedie D, Yang HT, Anisimov SV, and Wobus AM. Differentiation of pluripotent embryonic stem cells into cardiomyocytes. Circ Res 91: 189 –201, 2002. 63. Bolli R. Regeneration of the human heart: no chimera? N Engl J Med 346: 5–15, 2002. 64. Borue X, Lee S, Grove J, Herzog EL, Harris R, Diflo T, Glusac E, Hyman K, Theise ND, and Krause DS. Bone marrow-derived cells contribute to epithelial engraftment during wound healing. Am J Pathol 165: 1767–1772, 2004. 65. Braun KM, Niemann C, Jensen UB, Sundberg JP, Silva-Vargas V, and Watt FM. Manipulation of stem cell proliferation and lineage commitment: visualization of label-retaining cells in wholemounts of mouse epidermis. Development 130: 5241–5255, 2003. 66. Braun KM and Watt FM. Epidermal label-retaining cells: background and recent applications. J Invest Dermatol Symp Proc 9: 196 –201, 2004. 67. Braz JC, Gregory K, Pathak A, Zhao W, Sahin B, Klevitsky R, Kimball TF, Lorenz JN, Nairn AC, Liggett SB, Bodi I, Wang S, Schwartz A, Lakatta EG, DePaoli-Roach AA, Robbins J, Hewett TE, Bibb JA, Westfall MV, Kranias EG, and Molkentin JD. PKC-alpha regulates cardiac contractility and propensity toward heart failure. Nat Med 10: 248 –254, 2004. 68. Brittan M, Braun KM, Reynolds LE, Conti FJ, Reynolds AR, Poulsom R, Alison MR, Wright NA, and Hodivala-Dilke KM. Bone marrow cells engraft within the epidermis and proliferate in vivo with no evidence of cell fusion. J Pathol 205: 1–13, 2005. 69. Britten MB, Abolmaali ND, Assmus B, Lehmann R, Honold J, Schmitt J, Vogl TJ, Martin H, Schachinger V, Dimmeler S, and Zeiher AM. Infarct remodeling after intracoronary progenitor cell treatment in patients with acute myocardial infarction (TOPCAREAMI): mechanistic insights from serial contrast-enhanced magnetic resonance imaging. Circulation 108: 2212–2218, 2003. 70. Brodsky WY and Uryvaeva IV. Cell polyploidy: its relation to tissue growth and function. Int Rev Cytol 50: 275–332, 1977. 71. Bruchez M Jr, Moronne M, Gin P, Weiss S, and Alivisatos AP. Semiconductor nanocrystals as fluorescent biological labels. Science 281: 2013–2013, 1998. 72. Bruder SP, Jaiswal N, and Haynesworth SE. Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. J Cell Biochem 64: 278 –294, 1997. 73. Bunting KD. ABC transporters as phenotypic markers and functional regulators of stem cells. Stem Cells 20: 11–20, 2002. 74. Bunting HD, Zhou S, Lu T, and Sorrentino BP. Enforced Pglycoprotein pump function in murine bone marrow cells results in expansion of side population stem cells in vitro and repopulating cells in vivo. Blood 96: 902–909, 2000. 75. Cable J, Jackson IJ, and Steel KP. Mutations at the W locus affect survival of neural crest-derived melanocytes in the mouse. Mech Dev 50: 139 –150, 1995. 76. Cai J, Weiss ML, and Rao MS. In search of “stemness.” Exp Hematol 32: 585–598, 2004. 77. Cai CL, Liang X, Shi Y, Chu PH, Pfaff SL, Chen J, and Evans S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev Cell 5: 877– 889, 2003. 78. Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, Martin RP, Schipani E, Divieti P, and Bringhurst 1406 102. 103. 104. 105. 106. 108. 109. 110. 111. 112. 113. 114. 115. 116. 117. 118. 119. 120. 121. antibodies and multiparameter fow cytometry. Int J Cell Cloning 5: 267–288, 1987. Clark EB. Functional Aspects of Cardiac Development. Growth of the Heart in Health and Disease. New York: Raven, 1984. Clark RAF. The Molecular and Cellular Biology of Wound Repair. New York: Plenum, 1996. Cohn JN, Ferrari R, and Sharpe N. Cardiac remodeling— concepts and clinical implications: a consensus paper from an international forum on cardiac remodeling. Behalf of an International Forum on Cardiac Remodeling. J Am Coll Cardiol 35: 569 –582, 2000. Colter DC, Class R, DiGirolamo CM, and Prockop DJ. Rapid expansion of recycling stem cells in cultures of plastic-adherent cells from human bone marrow. Proc Natl Acad Sci USA 97: 3213–3218, 2000. Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL, and Rando T. Rejuvenation of aged progenitor cells by exposure to a young systematic environment. Nature 433: 760 –764, 2005. Conboy IM, Conboy MJ, Smythe GM, and Rando T. A Notchmediated restoration of regenerative potential to aged muscle. Science 302: 1575–1577, 2003. Conboy I and Rando T. Aging, stem cells and tissue regeneration: lessons from muscle. Cell Cycle. In press. Condorelli G, Borello U, De Angelis L, Latronico M, Sirabella D, Coletta M, Galli R, Balconi G, Follenzi A, Frati G, Cusella De Angelis MG, Gioglio L, Amuchastegui S, Adorini L, Naldini L, Vescovi A, Dejana E, and Cossu G. Cardiomyocytes induce endothelial cells to trans-differentiate into cardiac muscle: implication for myocardium regeneration. Proc Natl Acad Sci USA 98: 10733–10738, 2001. Corti S, Locatelli F, Donadoni C, Guglieri M, Papadimitriou D, Strazzer S, Del Bo R, and Comi GP. Wild-type bone marrow cells ameliorate the phenotype of SOD1–G93A ALS mice and contribute to CNS, heart and skeletal muscle tissues. Brain 127: 2518 – 2532, 2004. Dahlke MH, Popp FC, Larsen S, Schlitt HJ, and Rasko JE. Stem cell therapy of the liver—fusion or fiction? Liver Transplant 10: 471– 479, 2004. Davani S, Deschaseaux F, Chalmers D, Tiberghien P, and Kantelip JP. Can stem cells mend a broken heart? Cardiovasc Res 65: 305–316, 2005. Davey RE and Zandstra PW. Signal processing underlying extrinsic control of stem cell fate. Curr Opin Hematol 11: 95–101, 2004. Davidoff MS, Middendorf R, Enikopolov G, Rietmacher D, Holstein AF, and Muller D. Progenitor cells of the testoroneproducing Leydig cells revealed. J Cell Biol 167: 935–944, 2004. Dawn B, Stein AB, Urbanek K, Rota M, Whang B, Rastaldo R, Torella I, Tang XL, Rezazadeh A, Kajstura J, Leri A, Hunt G, Varma J, Prabhu S, Anversa P, and Bolli R. Cardiac stem cells delivered intravascularly traverse the vessel barrier, regenerate infarcted myocardium, and improve cardiac function. Proc Natl Acad Sci USA 102: 3766 –3771, 2005. Deb A, Wang S, Skelding KA, Miller D, Simper D, and Caplice NM. Bone marrow-derived cardiomyocytes are present in adult human heart: a study of gender-mismatched bone marrow transplantation patients. Circulation 107: 1247–1249, 2003. Dennis JE and Charbord P. Origin and differentiation of human and murine stroma. Stem Cells 20: 205–214, 2002. Desbaillets I, Ziegler U, Groscurth P, and Gassmann M. Embryoid bodies: an in vitro model of mouse embryogenesis. Exp Physiol 85: 645– 651, 2000. Digirolamo CM, Stokes D, Colter D, Phinney DG, Class R, and Prockop DJ. Propagation and senescence of human marrow stromal cells in culture: a simple colony-forming assay identifies samples with the greatest potential to propagate and differentiate. Br J Haematol 107: 275–281, 1999. Dimmeler S, Zeiher AM, and Schneider MD. Unchain my heart: the scientific foundations of cardiac repair. J Clin Invest 115: 572–583, 2005. Dodou E, Verzi MP, Anderson JP, Xu SM, and Black BL. Mef2c is a direct transcriptional target of ISL1 and GATA factors in the anterior heart field during mouse embryonic development. Development 131: 3931–3942, 2004. Physiol Rev • VOL 122. Doetsch F. A niche for adult neural stem cells. Curr Opin Genet Dev 13: 543–550, 2003. 123. Donnelly DS and Krause DS. Hematopoietic stem cells can be CD34⫹ or CD34⫺. Leuk Lymphoma 40: 221–234, 2001. 124. Doss MX, Koehler CI, Gissel C, Hescheler J, and Sachinidis A. Embryonic stem cells: a promising tool for cell replacement therapy. J Cell Mol Med 8: 465– 473, 2004. 125. Doyonnas R, LaBarge MA, Sacco A, Charlton C, and Blau HM. Hematopoietic contribution to skeletal muscle regeneration by myelomonocytic precursors. Proc Natl Acad Sci USA 101: 13507– 13512, 2004. 126. Draper JS and Fox V. Human embryonic stem cells: multilineage differentiation and mechanisms of self-renewal. Arch Med Res 34: 558 –564, 2003. 127. Draper JS, Pigott C, Thomson JA, and Andrews PW. Surface antigens of human embryonic stem cells: changes upon differentiation in culture. J Anat 200: 249 –258, 2002. 128. Drukker M and Benvenisty N. The immunogenicity of human embryonic stem-derived cells. Trends Biotechnol 22: 136 –141, 2004. 129. Eberhard D and Jockusch H. Patterns of myocardial histogenesis as revealed by mouse chimeras. Dev Biol 278: 336 –346, 2005. 130. Edgar BA and Orr-Weaver TL. Endoreplication cell cycles: more for less. Cell 105: 297–306, 2001. 131. Eglitis MA and Mezey E. Hematopoietic cells differentiate into both microglia and macroglia in the brains of adult mice. Proc Natl Acad Sci USA l94: 4080 – 4085, 1997. 132. Eguchi G and Kodama R. Transdifferentiation. Curr Opin Cell Biol 5: 1023–1028, 1993. 133. Eisenberg CA and Eisenberg LM. WNT11 promotes cardiac tissue formation of early mesoderm. Dev Dyn 216: 45–58, 1999. 134. Eisenberg LM, Burns L, and Eisenberg CA. Hematopoietic cells from bone marrow have the potential to differentiate into cardiomyocytes in vitro. Anat Rec A Discov Mol Cell Evol Biol 274: 870 – 882, 2003. 135. Eisenberg LM and Eisenberg CA. Stem cell plasticity, cell fusion, and transdifferentiation. Birth Defects Res C Embryo Today 69: 209 –218, 2003. 136. Eisenberg LM and Eisenberg CA. An in vitro analysis of myocardial potential indicates that phenotypic plasticity is an innate property of early embryonic tissue. Stem Cells Dev 13: 614 – 624, 2004. 137. Eisenberg LM and Eisenberg CA. Adult stem cells and their cardiac potential. Anat Rec A Discov Mol Cell Evol Biol 276: 103– 112, 2004. 138. Eisenberg LM, Kubalak SW, and Eisenberg CA. Stem cells and the formation of the myocardium in the vertebrate embryo. Anat Rec A Discov Mol Cell Evol Biol 276: 2–12, 2004. 139. Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, and Gage FH. Neurogenesis in the adult human hippocampus. Nat Med 4: 1313–1317, 1998. 140. Estes BT, Gimble JM, and Guilak F. Mechanical signals as regulators of stem cell fate. Curr Top Dev Biol 60: 91–126, 2004. 141. Estivill-Torrus G, Pearson H, van Heyningen V, Price DJ, and Rashbass P. Pax 6 is required to regulate the cell cycle and the rate of progression from symmetrical to asymmetrical division in mammalian cortical progenitors. Development 129: 455– 466, 2002. 142. Etzion S, Battler A, Barbash IM, Cagnano E, Zarin P, Granot Y, Kedes LH, Kloner RA, and Leor J. Influence of embryonic cardiomyocyte transplantation on the progression of heart failure in a rat model of extensive myocardial infarction. J Mol Cell Cardiol 33: 1321–1330, 2001. 143. Ferguson MW and O’Kane S. Scar-free healing: from embryonic mechanisms to adult therapeutic intervention. Philos Trans R Soc Lond B Biol Sci 359: 839 – 850, 2004. 144. Fernandez-Aviles F, San Roman JA, Garcia-Frade J, Fernandez ME, Penarrubia MJ, de la Fuente L, Gomez-Bueno M, Cantalapiedra A, Fernandez J, Gutierrez O, Sanchez PL, Hernandez C, Sanz R, Garcia-Sancho J, and Sanchez A. Experimental and clinical regenerative capability of human bone marrow cells after myocardial infarction. Circ Res 95: 742–748, 2004. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 107. LERI, KAJSTURA, AND ANVERSA 1407 ADULT STEM CELLS Physiol Rev • VOL 166. 167. 168. 169. 170. 171. 172. 173. 174. 175. 176. 177. 178. 179. 180. 181. 182. 183. 184. 185. 186. 187. bone marrow progenitors, fibroblasts, and endothelial cells. Arterioscler Thromb Vasc Biol 25: 692– 697, 2005. Geissler EN, Ryan MA, and Housman DE. The dominant-white spotting (W) locus of the mouse encodes the c-kit proto-oncogene. Cell 55: 185–192, 1988. Gilbert DM. The future of embryonic stem cell research: addressing ethical conflict with responsible scientific research. Med Sci Monit 10: 99 –103, 2004. Glaser R, Lu MM, Narula N, and Epstein JA. Smooth muscle cells, but not myocytes, of host origin in transplanted human hearts. Circulation 106: 17–19, 2002. Gnecchi M, He H, Liang OD, Melo LG, Morello F, Mu H, Noiseux N, Zhang L, Pratt RE, Ingwall JS, and Dzau VJ. Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nat Med 11: 367–368, 2005. Goldberg GS, Valiunas V, and Brink PR. Selective permeability of gap junction channels. Biochim Biophys Acta 23: 96 –101, 2004. Gonzalez-Reyes A. Stem cells, niches and cadherins: a view from Drosophila. J Cell Sci 116: 949 –954, 2003. Goodell MA, McKinney-Freeman S, and Camargo FD. Isolation and characterization of side population cells. Methods Mol Biol 290: 343–352, 2005. Gorla GR, Malhi H, and Gupta S. Polyploidy associated with oxidative injury attenuates proliferative potential of cells. J Cell Sci 114: 2943–2951, 2001. Groeneveld K, te Marvelde JG, van den Beemd MW, Hooijkaas H, and van Dongen JJ. Flow cytometric detection of intracellular antigens for immunophenotyping of normal and malignant leukocytes. Leukemia 10: 1383–1389, 1996. Gronthos S, Graves SE, Ohta S, and Simmons PJ. The STRO-1⫹ fraction of adult human bone marrow contains the osteogenic precursors. Blood 84: 4164 – 4173, 1994. Grossman W, Carbello BA, Gunther S, and Fifer MA. Ventricular wall stress and the development of cardiac hypertrophy and failure. In: Myocardial Hypertrophy and Failure. New York: Raven, 1983, p. 1– 8. Grossman W, Jones D, and McLaurin LP. Wall stress and patterns of hypertrophy in the human left ventricle. J Clin Invest 56: 56 – 64, 1975. Grove JE, Bruscia E, and Krause DS. Plasticity of bone marrowderived stem cells. Stem Cells 22: 487–500, 2004. Guerra S, Leri A, Wang X, Finato N, Di Loreto C, Beltrami CA, Kajstura J, and Anversa P. Myocyte death in the failing human heart is gender dependent. Circ Res 85: 856 – 866, 1999. Guidotti JE, Bregerie O, Robert A, Debey P, Brechot C, and Desdouets C. Liver cell polyploidization: a pivotal role for binuclear hepatocytes. J Biol Chem 278: 19095–19101, 2003. Guo Y, Lubbert M, and Engelhardt M. CD34- hematopoietic stem cells: current concepts and controversies. Stem Cells 21: 15–20, 2003. Gupta S. Hepatic polyploidy and liver growth control. Semin Cancer Biol 10: 161–171, 2000. Gupta S, Bhargava KK, and Novikoff PM. Mechanisms of cell engraftment during liver repopulation with hepatocyte transplantation. Semin Liver Dis 19: 15–26, 1999. Gussoni E, Soneoka Y, Strickland CD, Buzney EA, Khan MK, Flint AF, Kunkel LM, and Mulligan RC. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 401: 390 –394, 1999. Hakuno D, Fukuda K, Makino S, Konishi F, Tomita Y, Manabe T, Suzuki Y, Umezawa A, and Ogawa S. Bone marrow-derived regenerated cardiomyocytes (CMG Cells) express functional adrenergic and muscarinic receptors. Circulation 105: 380 –386, 2002. Hamano K, Nishida M, Hirata K, Mikamo A, Li TS, Harada M, Miura T, Matsuzaki M, and Esato K. Local implantation of autologous bone marrow cells for therapeutic angiogenesis in patients with ischaemic heart disease: clinical trial and preliminary results. Jpn Circ J 65: 845– 847, 2001. Hamazaki T, Oka M, Yamanaka S, and Terada N. Aggregation of embryonic stem cells induces Nanog repression and primitive endoderm differentiation. J Cell Sci 117: 5681–5686, 2004. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 145. Ferrari G, Cusella-De Angelis G, Coletta M, Paolucci E, Stornaiuolo Cossu G, and Mavilio F. Muscle regeneration by bone marrow-derived myogenic progenitors. Science 279: 528 –530, 1998. 146. Ferrer D, Gabbianelli M, Peschle C, Lange B, and Rovera G. Surface phenotypes of human hematopoietic progenitor cells defined by monoclonal antibodies. Blood 66: 496 –502, 1985. 147. Fijvandraat AC, vanGinneken AC, de Boer PA, Ruijter JM, Christoff VM, Moorman AF, and Lekanne Deprez RH. Cardiomyocytes derived from embryonic stem cells resemble cardiomyocytes of the embryonic heart tube. Cardiovasc Res 58: 399 – 409, 2003. 148. Fiordaliso F, Leri A, Cesselli D, Limana F, Safai B, NadalGinard B, Anversa P, and Kajstura J. Hyperglycemia activates p53 and p53-regulated genes leading to myocyte cell death. Diabetes 50: 2363–2375, 2001. 149. Fishbach GD and Fishbach RL. Stem cells: science, policy, and ethics. J Clin Invest 114: 1364 –1370, 2004. 150. Forbes SJ, Vig P, Poulsom R, Wright NA, and Alison MR. Adult stem cell plasticity: new pathways of tissue regeneration become visible. Clin Sci 103: 355–369, 2002. 151. Forrester JS, Price MJ, and Makkar RR. Stem cell repair of infarcted myocardium: an overview for clinicians. Circulation 108: 1139 –1145, 2003. 152. Frangogiannis NG, Perrard JL, Mendoza LH, Burns AR, Lindsey ML, Ballantyne CM, Micheal LH, Smith CW, and Entman ML. Stem cell factor induction is associated with mast cell accumulation after canine myocardial ischemia and reperfusion. Circulation 98: 687– 698, 1998. 153. French SW, Hoyer KK, Shen RR, and Teitell MA. Transdifferentiation and nuclear reprogramming in hematopoietic development and neoplasia. Immunol Rev 187: 22–39, 2002. 154. Friedenstein AJ. Osteogenetic activity of transplanted transitional epithelium. Acta Anat 45: 31–59, 1961. 155. Friedenstein AJ, Chailakhjan RK, and Lalykina KS. The development of fibroblast colonies in monolayer cultures of guineapig bone marrow and spleen cells. Cell Tissue Kinet 3: 393– 403, 1970. 156. Frustaci A, Kajstura J, Chimenti C, Jakoniuk I, Leri A, Maseri A, Nadal-Ginard B, and Anversa P. Myocardial cell death in human diabetes. Circ Res 87: 1123–1132, 2000. 157. Fuchs E, Tumbar T, and Guasch G. Socializing with the neighbors: stem cells and their niche. Cell 116: 769 –778, 2004. 158. Fuchs S, Satler LF, Kornowski R, Okubagzi P, Weisz G, Baffour R, Waksman R, Weissman NJ, Cerqueira M, Leon MB, and Epstein SE. Catheter-based autologous bone marrow myocardial injection in no-option patients with advanced coronary artery disease: a feasibility study. J Am Coll Cardiol 41: 1721–1724, 2003. 159. Fukuda K. Molecular characterization of regenerated cardiomyocytes derived from adult mesenchymal stem cells. Congenit Anom 42: 1–9, 2002. 160. Fukuda K. Use of adult marrow mesenchymal stem cells for regeneration of cardiomyocytes. Bone Marrow Transplant 32: S25–S27, 2003. 161. Fukuda S and Taga T. Cell fate determination regulated by a transcriptional signal network in the developing mouse brain. Anat Sci Int 80: 12–18, 2005. 162. Fukuhara S, Tomita S, Nakatani T, Ohtsu Y, Ishida M, Yutani C, and Kitamura S. G-CSF promotes bone marrow cells to migrate into infarcted mice heart, and differentiate into cardiomyocytes. Cell Transplant 13: 741–748, 2004. 163. Galinanes M, Loubani M, Davies J, Chin D, Pasi J, and Bell PR. Autotransplantation of unmanipulated bone marrow into scarred myocardium is safe and enhances cardiac function in humans. Cell Transplant 13: 7–13, 2004. 164. Gallagher KL and Benfey PN. Not just another hole in the wall: understanding intercellular protein trafficking. Genes Dev 19: 89 – 95, 2005. 165. Galli D, Innocenzi A, Staszewsky L, Zanetta L, Sampaolesi M, Bai A, Martinoli E, Carlo E, Balconi G, Fiordaliso F, Chimenti S, Cusella G, Dejana E, Cossu G, and Latini R. Mesoangioblasts, vessel-associated multipotent stem cells, repair the infarcted heart by multiple cellular mechanisms. A comparison with 1408 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL 208. 209. 210. 211. 212. 213. 214. 215. 216. 217. 218. 219. 220. 221. 222. 223. 224. 225. 226. 227. dren with osteogenesis imperfecta: implications for cell therapy of bone. Proc Natl Acad Sci USA 99: 8932– 8937, 2002. Horwitz EM, Prockop DJ, Fitzpatrick LA, Koo WW, Gordon PL, Neel M, Sussman M, Orchard P, Marx JC, Pyeritz RE, and Brenner MK. Transplantability and therapeutic effects of bone marrow-derived mesenchymal cells in children with osteogenesis imperfecta. Nat Med 5: 309 –313, 1999. Horwitz EM, Prockop DJ, Gordon PL, Koo WW, Fitzpatrick LA, Neel MD, McCarville ME, Orchard PJ, Pyeritz RE, and Brenner MK. Clinical responses to bone marrow transplantation in children with severe osteogenesis imperfecta. Blood 97: 1227– 1231, 2001. Hsieh J and Gage FH. Epigenetic control of neural stem cell fate. Curr Opin Genet Dev 14: 1–9, 2004. Ianus A, Holz GG, Theise ND, and Hussain MA. In vivo derivation of glucose-competent pancreatic endocrine cells from bone marrow without evidence of cell fusion. J Clin Invest 111: 843– 850, 2003. Imitola J, Park KI, Teng YD, Nisim S, Lachyankar M, Ourednik J, Mueller FJ, Yiou R, Atala A, Sidman RL, Tuszynski M, Khoury SJ, and Snyder EY. Stem cells: cross-talk and developmental programs. Philos Trans R Soc Lond B Biol Sci 359: 823– 837, 2004. In ’t Anker PS, Scherjon SA, Kleijburg-van der Keur C, Noort WA, Claas FH, Willemze R, Fibbe WE, and Kanhai HH. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 102: 1548 –1549, 2003. In ’t Anker PS, Scherjon SA, Kleijburg-van der Keur C, de Groot-Swings GM, Claas FH, Fibbe WE, and Kanhai HH. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta. Stem Cells 22: 1338 –1345, 2004. Ito CY, Li CY, Bernstein A, Dick JE, and Stanford WL. Hematopoietic stem cell and progenitor defects in Sca-1/Ly-6A-null mice. Blood 101: 517–523, 2003. Itskovitz-Eldor JI, Schuldiner M, Karsenti D, Eden A, Yanuka O, Amit M, Soreq H, and Benvenisty N. Differentiation of human embryonic stem cells into embryoid bodies comprising the three embryonic germ layers. Mol Med 6: 88 –95, 2000. Jackson KA, Majka SM, Wang H, Pocius J, Hartley CJ, Majesky MW, Entman ML, Michael LH, Hirschi KK, and Goodell MA. Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells. J Clin Invest 107: 1395–1402, 2001. Jackson KA, Snyder DS, and Goodell MA. Skeletal muscle fiber-specific green autofluorescence: potential for stem cell engraftment artifacts. Stem Cells 22: 180 –187, 2004. Jackson KA, Majka SM, Wulf GG, and Goodell MA. Stem cells: a minireview. J Cell Immun Biochem 38: 1– 6, 2002. Jafar-Nejad H, Norga K, and Bellen H. Numb: “Adapting” Notch for endocytosis. Dev Cell 3: 155–156, 2002. Jaiswal N, Haynesworth SE, Caplan AI, and Bruder SP. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J Cell Biochem 64: 295–301, 1997. Jalali S, del Pozo MA, Chen KD, Miao H, Li YS, Schwartz MA, Shyy JYJ, and Chien S. Integrin-mediated mechanotransduction requires its dynamic interaction with specific extracellular matrix (ECM) ligands. Proc Natl Acad Sci USA 98: 1042–1046, 2001. Jan YN and Jan LY. Asymmetric cell division. Nat Rev Neurosci 392: 775–778, 2001. Jessup M and Brozena S. Heart failure. N Engl J Med 348: 2007–2018, 2003. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, and Verfaillie CM. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 418: 41– 49, 2002. Kawada H, Fujita J, Kinjo K, Matsuzaki Y, Tsuma M, Miyatake H, Muguruma Y, Tsuboi K, Itabashi Y, Ikeda Y, Ogawa S, Okano H, Hotta T, Ando K, and Fukuda K. Nonhematopoietic mesenchymal stem cells can be mobilized and differentiate into cardiomyocytes after myocardial infarction. Blood 104: 3581–3587, 2004. Kajstura J, Cheng W, Sarangarajan R, Li P, Li B, Nitahara JA, Chapnick S, Reiss K, Olivetti G, and Anversa P. Necrotic and 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 188. Harada M, Qin Y, Takano H, Minamino T, Zou Y, Toko H, Ohtsuka M, Matsuura K, Sano M, Nishi J, Iwanaga K, Akazawa H, Kunieda T, Zhu W, Hasegawa H, Kunisada K, Nagai T, Nakaya H, Yamauchi-Takihara K, and Komuro I. G-CSF prevents cardiac remodeling after myocardial infarction by activating the Jak-Stat pathway in cardiomyocytes. Nat Med 11: 305–311, 2005. 189. Hardy K, Spanos S, Becker D, Iannelli P, Winston RM, and Stark J. From cell death to embryo arrest: mathematical models of human preimplantation embryo development. Proc Natl Acad Sci USA 98: 1655–1660, 2001. 190. Harris RG, Herzog EL, Bruscia EM, Grove JE, Van Arnam JS, and Krause DS. Lack of a fusion requirement for development of bone marrow-derived epithelia. Science 305: 90 –93, 2004. 191. Hart AH, Hartley L, Ibrahim M, and Robb L. Identification, cloning and expression analysis of the pluripotency promoting Nanog genes in mouse and human. Dev Dyn 230: 187–198, 2004. 192. Harvey KA, Siddiqui RA, Jansen J, Akard LP, Thompson JM, Cui Y, Chang Q, and English D. Growth factor induction of cytosolic protein tyrosine kinase activity in human haemopoietic progenitor cells isolated by flow cytometry. Br J Haematol 93: 515–526, 1996. 193. Hatano SY, Tada M, Kimura H, Yamaguchi S, Kono T, Nakano T, Suemori H, Nakatsuji N, and Tada T. Pluripotential competence of cells associated with Nanog activity. Mech Dev 122: 67–79, 2005. 194. Hattan N, Kawaguchi H, Ando K, Kuwabara E, Fujita J, Murata M, Suematsu M, Mori H, and Fukuda K. Purified cardiomyocytes from bone marrow mesenchymal stem cells produce stable intracardiac grafts in mice. Cardiovasc Res 65: 334 –344, 2005. 195. Haynesworth SE, Baber MA, and Caplan AI. Cell surface antigens on human marrow-derived mesenchymal cells are detected by monoclonal antibodies. Bone 13: 69 – 80, 1992. 196. He JQ, Ma Y, Lee Y, Thomson JA, and Kamp TJ. Human embryonic stem cells develop into multiple types of cardiac myocytes: action potential characterization. Circ Res 93: 32–39, 2003. 197. Heber-Katz E, Leferovich J, Bedelbaeva K, Gourevitch D, and Clark L. The scarless heart and the MRL mouse. Philos Trans R Soc Lond B Biol Sci 359: 785–793, 2004. 198. Heng BC, Haider HK, Sim EK, Cao T, Tong GQ, and Ng SC. Comments about possible use of human embryonic stem cellderived cardiomyocytes to direct autologous adult stem cells into the cardiomyogenic lineage. Acta Cardiol 60: 7–12, 2005. 199. Heeschen C, Lehmann R, Honold J, Assmus B, Aicher A, Walter DH, Martin H, Zeiher AM, and Dimmeler S. Profoundly reduced neovascularization capacity of bone marrow mononuclear cells derived from patients with chronic ischemic heart disease. Circulation 109: 1615–1622, 2004. 200. Heil M, Ziegelhoeffer T, Mees B, and Schaper W. A different outlook on the role of bone marrow stem cells in vascular growth: bone marrow delivers software not hardware. Circ Res 94: 573–574, 2004. 201. Hierlihy AM, Seale P, Lobe CG, Rudnicki MA, and Megeney LA. The post-natal heart contains a myocardial stem cell population. FEBS Lett 530: 239 –243, 2002. 202. Hirao A, Arai F, and Suda T. Regulation of cell cycle in hematopoietic stem cells by the niche. Cell Cycle 3: 1481–1483, 2004. 203. Hodgson DM, Behar A, Zingman LV, Kane GC, Perez-Terzic C, Alekseev AE, Puceat M, and Terzic A. Stable benefit of embryonic stem cell therapy in myocardial infarction. Am J Physiol Heart Circ Physiol 287: H471–H479, 2004. 204. Holden C and Vogel G. Stem cells plasticity: time for a reappraisal? Science 29: 2126 –2129, 2000. 205. Horsley V and Pavlath GK. Forming a multinucleated cell: molecules that regulate myoblast fusion. Cells Tissues Organs 176: 67–78, 2004. 206. Horwitz EM. Stem cell plasticity: the growing potential of cellular therapy. Arch Med Res 34: 600 – 606, 2003. 207. Horwitz EM, Gordon PL, Koo WK, Marx JC, Neel MD, McNall RY, Muul L, and Hofmann T. Isolated allogeneic bone marrowderived mesenchymal cells engraft and stimulate growth in chil- 1409 ADULT STEM CELLS 228. 229. 230. 231. 232. 234. 235. 236. 237. 238. 239. 240. 241. 242. 243. 244. 245. 246. 247. Physiol Rev • VOL 248. Kozorovitskiy Y and Gould E. Stem cell fusion in the brain. Nat Cell Biol 5: 952–954, 2003. 249. Kunisada T, Yoshida H, Yamakazi H, Miyamoto A, Hemmi H, Nishimura E, Shultz LD, Nishikawa S, and Hayashi S. Transgene expression of steel factor in the basal layer of epidermis promotes survival, proliferation, differentiation and migration of melanocyte precursors. Development 125: 2915–2923, 1998. 250. Laflamme MA, Myerson D, Saffitz JE, and Murry CE. Evidence for cardiomyocyte repopulation by extracardiac progenitors in transplanted human hearts. Circ Res 90: 634 – 640, 2002. 251. Lai EC. Notch signaling: control of cell communication and cell fate. Development 131: 965–973, 2004. 252. Lakatta EG and Levy D. Arterial and cardiac aging: major shareholders in cardiovascular disease enterprises. Part I: aging arteries: a “set up” for vascular disease. Circulation 107: 139 –146, 2003. 253. Lakshmipathy U and Verfaillie C. Stem cell plasticity. Blood Rev 19: 29 –38, 2005. 254. Landry DW and Zucker HA. Embryonic death and the creation of human embryonic stem cells. J Clin Invest 114: 1184 –1186, 2004. 255. Lanza F, Latorraca A, Moretti S, Castagnari B, Ferrari L, and Castoldi G. Comparative analysis of different permeabilization methods for the flow cytometry measurement of cytoplasmic myeloperoxidase and lysozyme in normal and leukemic cells. Cytometry 30: 134 –144, 1997. 256. Lanza RP, Cibelli JB, and West MD. Prospects for the use of nuclear transfer in human transplantation. Nat Biotechnol 17: 1171–1174, 1999. 257. Lanza RP, Cibelli JB, and West MD. Human therapeutic cloning. Nat Med 5: 975–977, 1999. 258. Lanza R, Moore MA, Wakayama T, Perry AC, Shieh JH, Hendrikx J, Leri A, Chimenti S, Monsen A, Nurzynska D, West MD, Kajstura J, and Anversa P. Regeneration of the infarcted heart with stem cells derived by nuclear transplantation. Circ Res 94: 820 – 827, 2004. 259. Lanza R and Rosenthal N. The stem cell challenge. Sci Am 290: 92–99, 2004. 260. Laugwitz KL, Moretti A, Lam J, Gruber P, Chen Y, Woodard S, Lin LZ, Cai CL, Lu MM, Reth M, Platoshyn O, Yuan JX, Evans S, and Chien KR. Postnatal isl1⫹cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 433: 585–587, 2005. 261. Laverge H, Van der Elst J, De Sutter P, Verschraegen-Spae MR, De Paepe A, and Dhont M. Fluorescent in-situ hybridization on human embryos showing cleavage arrest after freezing and thawing. Hum Reprod 13: 425– 429, 1998. 262. Leferovich JM, Bedelbaeva K, Samulewicz S, Zhang XM, Zwas D, Lankford EB, and Heber-Katz E. Heart regeneration in adult MRL mice. Proc Natl Acad Sci USA 98: 9830 –9835, 2001. 263. Leong FT and Freeman LJ. Acute renal infarction. J R Soc Med 98: 121–122, 2005. 264. Leor J, Patterson M, Quinones MJ, Kedes LH, and Kloner RA. Transplantation of fetal myocardial tissue into the infarcted myocardium of rat. A potential method for repair of infarcted myocardium? Circulation 94: II332–II336, 1996. 265. Leri A, Claudio PP, Li Q, Wang X, Reiss K, Wang S, Malhotra A, Kajstura J, and Anversa P. Stretch-mediated release of angiotensin II induces myocyte apoptosis by activating p53 that enhances the local RAS and decreases the Bcl-2 to Bax protein ratio in the cell. J Clin Invest 101: 1326 –1342, 1998. 266. Leri A, Kajstura J, Nadal-Ginard B, and Anversa P. Some like it plastic. Circ Res 94: 132–134, 2004. 267. Leri A, Liu Y, Li B, Fiordaliso F, Malhotra A, Latini R, Kajstura J, and Anversa P. Up-regulation of AT(1) and AT(2) receptors in postinfarcted hypertrophied myocytes and stretch-mediated apoptotic cell death. Am J Pathol 156: 1663–1672, 2000. 268. Levenberg S, Golub JS, Amit M, Itskovitz-Eldor J, and Langer R. Endothelial cells derived from human embryonic stem cells. Proc Natl Acad Sci USA 99: 4391– 4396, 2002. 269. Levinson H, Peled Z, Liu W, Longaker MT, Allison GM, and Ehrlich HP. Fetal rat amniotic fluid: TGF- and fibroblast collagen lattice formation. J Surg Res 100: 205–210, 2001. 270. Li E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet 3: 662– 673, 2002. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 233. apoptotic myocyte cell death in the aging heart of Fischer 344 rats. Am J Physiol Heart Circ Physiol 271: H1215–H228, 1996. Kajstura J, Rota M, Whang B, Cascapera S, Hosoda T, Bearzi C, Nurzynska D, Kasahara H, Zias E, Bonafe M, Nadal-Ginard B, Torella D, Nascimbene A, Quaini F, Urbanek K, Leri A, and Anversa P. Bone marrow cells differentiate in cardiac cell lineages after infarction independently of cell fusion. Circ Res 96: 127–137, 2005. Kajstura J, Leri A, Finato N, Di Loretto C, Beltrami CA, and Anversa P. Myocyte proliferation in end-stage cardiac failure in humans. Proc Natl Acad Sci USA 95: 8801– 8805, 1998. Karam R, Healy BP, and Wicker P. Coronary reserve is depressed in postmyocardial infarction reactive cardiac hypertrophy. Circulation 81: 238 –246, 1990. Karbanova J and Mokry J. Histological and histochemical analysis of embryoid bodies. Acta Histochem 104: 361–365, 2002. Keegan BR, Feldman JL, Begemann G, Ingham PW, and Yelon D. Retinoic acid signaling restricts the cardiac progenitor pool. Science 307: 247–249, 2005. Kehat I and Gepstein L. Human embryonic stem cells for myocardial regeneration. Heart Fail Rev 8: 229 –236, 2003. Kehat I, Kenyagin KD, Snir M, Segev H, Amit M, Gepstein A, Livne E, Binah O, Eldor JI, and Gepstein L. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest 108: 407– 414, 2001. Kehat I, Khimovich L, Caspi O, Gepstein A, Shofti R, Arbel G, Huber I, Satin J, Itskovitz-Eldor J, and Gepstein L. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Nat Biotech 10: 1– 8, 2004. Kerkis AY and Zhdanova NS. Formation and ultrastructure of somatic cell hybrids. Electron Microsc Rev 5: 1–24, 1992. Kikyo N and Wolffe AP. Reprogramming nuclei: insights from cloning, nuclear transfer and heterokaryons. J Cell Sci 113: 11–20, 2000. Kim KM and Shibata D. Methylation reveals a niche: stem cell succession in human colon crypts. Oncogene 21: 5441–5449, 2002. Kinnaird T, Stabile E, Burnett MS, and Epstein SE. Bonemarrow-derived cells for enhancing collateral development: mechanisms, animal data, and initial clinical experiences. Circ Res 95: 354 –363, 2004. Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S, and Epstein SE. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ Res 94: 678 – 685, 2004. Klinge O. Disturbances of karyokinesis and its relation to the nuclear pattern. Beitr Pathol 155: 79 – 83, 1975. Koc ON, Gerson SL, Cooper BW, Dyhouse SM, Haynesworth SE, Caplan AI, and Lazarus HM. Rapid haematopoietic recovery after co-infusion of autologous blood stem cells and culture-expanded marrow mesenchymal stem cells in advanced breast cancer patients receiving high dose chemotherapy. J Clin Oncol 18: 307– 316, 2000. Kofidis T, deBruin JL, Yamane T, Balsam LB, Lebl DR, Swijnenburg R, Tanaka M, Weissman IL, and Robbins RC. IGF-1 promotes engraftment, differentiation and functional improvement after transfer of embryonic stem cells for myocardial restoration. Stem Cells 22: 1239 –1245, 2004. Kofidis T, deBruin JL, Yamane T, Tanaka M, Lebl DR, Swijnenburg R, Weissman IL, and Robbins RC. Stimulation of paracrine pathways with growth factors enhances embryonic stem cell engraftement and host-specific differentiation in the heart after ischemic myocardial injury. Circulation 111: 2486 –2493, 2005. Koh CJ and Atala A. Tissue engineering, stem cells, and cloning: opportunities for regenerative medicine. J Am Soc Nephrol 15: 1113–1125, 2004. Kondo M, Wagers AJ, Manz MG, Prohaska SS, Scherer DC, Beilhach GF, Shizuru JA, and Weissman IL. Biology of hematopoietic stem cells and progenitors: implications for clinical application. Annu Rev Immunol 21: 759 – 806, 2003. Korbling M and Estrov Z. Adult stem cells and tissue repair. Bone Marrow Transplant 32: S23–S24, 2003. 1410 LERI, KAJSTURA, AND ANVERSA Physiol Rev • VOL 294. 295. 296. 297. 298. 299. 300. 301. 302. 303. 304. 305. 306. 307. 308. 309. 310. 311. 312. 313. 314. 315. Abcg2, identifies cardiac SP cells in the developing and adult heart. Dev Biol 265: 262–275, 2004. Martin MJ, Muotri A, Gage F, and Varki A. Human embryonic stem cells express an immunogenic nonhuman sialic acid. Nat Med 11: 228 –232, 2005. Mathur A and Martin JF. Stem cells and repair of the heart. Lancet 364: 183–192, 2004. Matsuura K, Nagai T, Nishigaki N, Oyama T, Nishi J, Wada H, Sano M, Toko H, Akazawa H, Sato H, Nakaya H, Kasanuki H, and Komuro I. Adult cardiac Sca-1-positive cells differentiate into beating cardiomyocytes. J Biol Chem 279: 11384 –11391, 2004. McBride WH, Chiang CS, Olson JL, Wang CC, Hong JH, Pajonk F, Dougherty GJ, Iwamoto KS, Pervan M, and Liao YP. A sense of danger from radiation. Radiat Res 162: 1–19, 2004. McCay CM, Pope F, Lunsford W, Sperling G, and Sambhavaphol P. Parabiosis between young and old rats. Gerontologia 1: 7–17, 1957. McKinney-Freeman SL, Jackson KA, Camargo FD, Ferrari G, Mavilio F, and Goodell MA. Muscle-derived hematopoietic stem cells are hematopoietic in origin. Proc Natl Acad Sci USA 99: 1341–1346, 2002. McKinney-Freeman S and Goodell MA. Circulating hematopoietic stem cells do not efficiently home to bone marrow during homeostasis. Exp Hematol 32: 868 – 876, 2004. Medvinsky A and Smith A. Stem cells: fusion brings down barriers. Nature 422: 823– 825, 2003. Meilhac SM, Esner M, Kerszberg M, Moss JE, and Buckingham ME. Oriented clonal cell growth in the developing mouse myocardium underlies cardiac morphogenesis. J Cell Biol 164: 97–109, 2004. Meilhac SM, Esner M, Kelly RG, Nicolas JF, and Buckingham ME. The clonal origin of myocardial cells in different regions of the embryonic mouse heart. Dev Cell 6: 685– 698, 2004. Meilhac SM, Kelly RG, Rocancourt D, Trinquet SE, Nicolas JF, and Buckingham ME. A retrospective clonal analysis of the myocardium reveals two phases of clonal growth in the developing mouse heart. The Company of Biologists. Development 130: 3877– 3889, 2003. Melan MA. Overview of cell fixatives and cell membrane permeants. Methods Mol Biol 115: 45–55, 1999. Menasche P, Hagege AA, Scorsin M, Pouzet B, Desnos M, Duboc D, Schwartz K, Vilquin JT, and Marolleau JP. Myoblast transplantation for heart failure. Lancet 347: 279 –280, 2001. Mendes SC, Robin C, and Dzierzak E. Mesenchymal progenitor cells localize within hematopoietic sites throughout ontogeny. Development 132: 1127–1136, 2005. Messina E, De Angelis L, Frati G, Morrone S, Chimenti S, Fiordaliso F, Salio M, Battaglia M, Latronico MVG, Coletta M, Vivarelli E, Frati L, Cossu G, and Giacomello A. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ Res 95: 911–921, 2004. Mezey E. Commentary: on bone marrow stem cells and openmindedness. Stem Cells Dev 13: 147–152, 2004. Mikawa T and Fischman DA. The polyclonal origin of myocyte lineages. Annu Rev Physiol 58: 509 –521, 1996. Min JY, Sullivan MF, Yang Y, Zhang JP, Converso KL, Morgan JP, and Xiao YF. Significant improvement of heart function by cotransplantation of human mesenchymal stem cells and fetal cardiomyocytes in postinfarcted pigs. Ann Thorac Surg 74: 1568 –1575, 2002. Min JY, Yang Y, Converso KL, Liu L, Huang Q, Morgan JP, and Xiao YF. Transplantation of embryonic stem cells improves cardiac function in post-infarcted rats. J Appl Physiol 92: 288 –296, 2002. Min JY, Yang Y, Sullivan MF, Ke Q, Converso KL, Chen Y, Morgan JP, and Xiao YF. Long-term improvement of cardiac function in rats after infarction by transplantation of embryonic stem cells. J Thorac Cardiovasc Surg 125: 361–369, 2003. Minguell JJ, Erices A, and Conget P. Mesenchymal stem cells. Exp Biol Med 226: 507–520, 2001. Mironov V, Visconti RP, and Markwald RR. What is regenerative medicine? Emergence of applied stem cell and developmental biology. Exp Opin Biol Ther 4: 773–781, 2004. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 271. Li J, Santoro R, Koberna K, and Grummt I. The chromatin remodeling complex NoRC controls replication timing of rRNA genes. EMBO J 24: 120 –127, 2004. 272. Li RK, Jia ZQ, Weisel RD, Merante F, and Mickle DA. Smooth muscle cell transplantation into myocardial scar tissue improves heart function. J Mol Cell Cardiol 31: 513–522, 1999. 273. Li TS, Hamano K, Hirata K, Kobayashi T, and Nishida M. The safety and feasibility of the local implantation of autologous bone marrow cells for ischemic heart disease. J Card Surg 18: S69 –S75, 2003. 274. Liew CC and Dzau VJ. Molecular genetics and genomics of heart failure. Nat Rev Genet 5: 811– 825, 2004. 275. Lin F, Cordes K, Li L, Hood L, Couser WG, Shankland SJ, and Igarashi P. Bone marrow stem cells contribute to healing of the kidney. J Am Soc Nephrol 14: S48 –S54, 2003. 276. Lin H. The stem cell niche theory: lessons from flies. Nat Rev Sci 3: 931–940, 2002. 277. Linke A, Müller P, Nurzynska D, Casarsa C, Torella D, Nascimbene A, Castaldo C, Cascapera S, Böhm M, Quaini F, Urbanek K, Leri A, Hintze TH, Kajstura J, and Anversa P. Cardiac stem cells in the dog heart regenerate infarcted myocardium improving cardiac performance. Proc Natl Acad Sci USA 102: 8966 – 8971, 2005. 278. Liu Y, Leri A, Li B, Wang X, Cheng W, Kajstura J, and Anversa P. Angiotensin II stimulation in vitro induces hypertrophy of normal and postinfarcted ventricular myocytes. Circ Res 82: 1145– 1159, 1998. 279. Liu Y and Rao MS. Transdifferentiation—fact or artifact. J Cell Biochem 88: 29 – 40, 2003. 280. Lopez LR, Shocket AL, Stanford RE, Claman HN, and Kohler PF. Gastro-intestinal involvement in leukocytoclastic vasculitis and polyarteritis nodosa. J Rheumatol 7: 677– 684, 1980. 281. Losordo DW and Dimmeler S. Therapeutic angiogenesis and vasculogenesis for ischemic disease: part II: cell-based therapies. Circulation 109: 2692–2697, 2004. 282. Losordo DW and Dimmeler S. Therapeutic angiogenesis and vasculogenesis for ischemic disease. Part I: angiogenic cytokines. Circulation 109: 2487–2491, 2004. 283. Loud AV and Anversa P. Morphometric analysis of biologic processes. Lab Invest 50: 250 –261, 1984. 284. Lum LG, Padbury JF, Davol PA, and Lee RJ. Virtual reality of stem cell transplantation to repair injured myocardium. J Cell Biochem. In press. 285. Lysaght MJ and Hazlehurst AL. Private sector development of stem cell technology and therapeutic cloning. Tissue Eng 9: 555– 561, 2003. 286. MacKay AM, Beck SC, Murphy JM, Barry FP, Chichester CO, and Pettenger MF. Chondrogenic differentiation of cultured human mesenchymal stem cells from marrow. Tissue Eng 4: 415– 428, 1998. 287. Mackool RJ, Gittes GK, and Longaker MT. Scarless healing: the fetal wound. Clin Plast Surg 25: 357–361, 1998. 288. MacLellan WR and Schneider MD. Genetic dissection of cardiac growth control pathways. Annu Rev Physiol 62: 289 –319, 2000. 289. Majka SM, Jackson KA, Kienstra KA, Majesky MW, Goodell MA, and Hirschi KK. Distinct progenitor populations in skeletal muscle are bone marrow-derived and exhibit different cell fates during vascular regeneration. J Clin Invest 111: 71–79, 2003. 290. Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, Pan J, Sano M, Takahashi T, Hori S, Abe H, Hata J, Umezawa A, and Ogawa S. Cardiomyocytes can be generated from marrow stromal cells in vitro. J Clin Invest 103: 697–705, 1999. 291. Mangi AA, Noiseux N, Kong D, He H, Rezvani M, Ingwall JS, and Dzau VJ. Mesenchymal stem cells modified with Akt prevent remodeling and restore performance of infarcted hearts. Nat Med 9: 1195–1201, 2003. 292. Margolis RL, Lohez OD, and Andreassen PR. G1 tetraploidy check point and the suppression of tumorigenesis. J Cell Biochem 88: 673– 683, 2003. 293. Martin CM, Meeson AP, Robertson SM, Hawke TJ, Richardson JA, Bates S, Goetsch SC, Gallardo TD, and Garry DJ. Persistent expression of the ATP-binding cassette transporter, 1411 ADULT STEM CELLS Physiol Rev • VOL 335. 336. 337. 338. 339. 340. 341. 342. 343. 344. 345. 346. 347. 348. 349. 350. 351. 352. 353. F. Influence of mobilized stem cells on myocardial infarct repair in a nonhuman primate model. Blood 102: 436 – 468, 2003. Nurzynska D, Whang B, Rota M, Baker M, Torella D, Sheikh F, Quaini F, Urbanek K, Kajstura J, Leri A, Bolli R, and Anversa P. Cardiac stem cell activation and myocardial regeneration reverse heart failure and prolong the lifespan of senescent Fischer 344 rats. Circulation 110: III-171, 2004. Nygren JM, Jovinge S, Breitbach M, Sawen P, Roll W, Hescheler J, Taneera J, Fleischmann BK, and Jacobsen SE. Bone marrow-derived hematopoietic cells generate cardiomyocytes at a low frequency through cell fusion, but not transdifferentiation. Nat Med 10: 494 –501, 2004. Odorico JS, Kaufman DS, and Thomson JA. Multilineage differentiation from human embryonic stem cell lines. Stem Cells 19: 193–204, 2001. Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, Mishina Y, Pocius J, Michael LH, Behringer RR, Garry DJ, and Schneider MD. Cardiac progenitor cells from adult myocardium: homing, differentiation, and fusion after infarction. Proc Natl Acad Sci USA 100: 12313–12318, 2003. Oh H and Schneider MD. The emerging role of telomerase in cardiac muscle cell growth and survival. J Mol Cell Cardiol 34: 717–724, 2002. Oh H, Wang SC, Prahash A, Sano M, Moravec CS, Taffet GE, Michael LH, Youker KA, Entman ML, and Schneider MD. Telomere attrition and Chk2 activation in human heart failure. Proc Natl Acad Sci USA 100: 5378 –5383, 2003. Ohlstein B, Kai T, Decotto E, and Spradling A. The stem cell niche: theme and variations. Curr Opin Cell Biol 16: 693– 699, 2004. Okumura-Nakanishi S, Saito M, Niwa H, and Ishikawa F. Oct-3/4 and Sox2 regulate Oct-3/4 gene in embryonic stem cells. J Biol Chem 280: 5307–5317, 2005. Oliver JA, Maarouf O, Cheema FH, Martens TP, and AlAwqati Q. The renal papilla is a niche for adult kidney stem cells. J Clin Invest 114: 795– 804, 2004. Olivetti G, Abbi R, Quaini F, Kajstura J, Cheng W, Nitahara JA, Quaini E, Di Loreto C, Beltrami CA, Krajewski S, Reed JC, and Anversa P. Apoptosis in the failing human heart. N Engl J Med 336: 1131–1141, 1997. Olivetti G, Capasso JM, Meggs LG, Sonnenblick EH, and Anversa P. Cellular basis of chronic ventricular remodeling after myocardial infarction in rats. Circ Res 68: 856 – 869, 1991. Olivetti G, Cigola E, Maestri R, Corradi D, Lagrasta C, Gambert SR, and Anversa P. Aging, cardiac hypertrophy and ischemic cardiomyopathy do not affect the proportion of mononucleated and multinucleated myocytes in the human heart. J Mol Cell Cardiol l28: 1463–1477, 1996. Olivetti G, Giordano G, Corradi D, Melissari M, Lagrasta C, Gambert SR, and Anversa P. Gender differences and aging: effects on the human heart. J Am Coll Cardiol 26: 1068 –1079, 1995. Olivetti G, Melissari M, Balbi T, Quaini F, Cigola E, Sonnenblick EH, and Anversa P. Myocyte cellular hypertrophy is responsible for ventricular remodeling in the hypertrophied heart of middle aged individuals in the absence of cardiac failure. Cardiovasc Res 28: 1199 –1208, 1994. Olivetti G, Melissari M, Balbi T, Quaini F, Sonnenblick EH, and Anversa P. Myocyte nuclear and possible cellular hyperplasia contribute to ventricular remodeling in the hypertrophic senescent heart in humans. J Am Coll Cardiol 24: 140 –149, 1994. Olivetti G, Melissari M, Capasso JM, and Anversa P. Cardiomyopathy of the aging human heart. Myocyte loss and reactive cellular hypertrophy. Circ Res 68: 1560 –1568, 1991. Olivetti G, Quaini F, Sala R, Lagrasta C, Corradi D, Bonacina E, Gambert SR, Cigola E, and Anversa P. Acute myocardial infarction in humans is associated with activation of programmed myocyte cell death in the surviving portion of the heart. J Mol Cell Cardiol 28: 2005–2016, 1996. Olivetti G, Ricci R, and Anversa P. Hyperplasia of myocyte nuclei in long-term cardiac hypertrophy in rats. J Clin Invest 80: 1818 –1821, 1987. Olivetti G, Ricci R, Lagrasta C, Maniga E, Sonnenblick EH, and Anversa P. Cellular basis of wall remodeling in long-term 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 316. Miyoshi K, Shillingford JM, Le Provost F, Gounari F, Bronson R, von Boehmer H, Taketo MM, Cardiff RD, Hennighausen L, and Khazaie K. Activation of beta-catenin signaling in differentiated mammary secretory cells induces transdifferentiation into epidermis and squamous metaplasias. Proc Natl Acad Sci USA 99: 219 –224, 2002. 317. Montecino-Rodriguez E and Dorshkind K. Regulation of hematopoiesis by gap junction-mediated intercellular communication. J Leukoc Biol 70: 341–347, 2001. 318. Montero C. The antigen-antibody reaction in immunohistochemistry. J Histochem Cytochem 51: 1– 4, 2003. 319. Morrison SJ, Uchida N, and Weissman IL. The biology of hematopoietic stem cells. Annu Rev Cell Dev Biol 11: 35–71, 1995. 320. Morrison SJ and Weissman IL. The long-term repopulating subset of hematopoietic stem cells is deterministic and isolatable by phenotype. Immunity 1: 661– 673, 1994. 321. Moscoso I, Centeno A, Lopez E, Rodriguez-Barbosa JI, Santamarina I, Filgueira P, Sanchez MJ, Dominguez-Perles R, Penuelas-Rivas G, and Domenech N. Differentiation “in vitro” of primary and immortalized porcine mesenchymal stem cells into cardiomyocytes for cell transplantation. Transplant Proc 37: 481– 482, 2005. 322. Muller P, Pfeiffer P, Koglin J, Schafers HJ, Seeland U, Janzen I, Urbschat S, and Bohm M. Cardiomyocytes of noncardiac origin in myocardial biopsies of human transplanted hearts. Circulation 106: 31–35, 2002. 323. Murry CE, Wiseman RW, Schwartz SM, and Hauschka SD. Skeletal myoblast transplantation for repair of myocardial necrosis. J Clin Invest 98: 2512–2523, 1996. 324. Murry CE, Soonpaa MH, Reinecke H, Nakajima H, Nakajima HO, Rubart M, Pasumarthi KB, Virag JI, Bartelmez SH, Poppa V, Bradford G, Dowell JD, Williams DA, and Field LJ. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Nature 428: 664 – 668, 2004. 325. Mutsaers SE, Bishop JE, McGrouther G, and Laurent GJ. Mechanisms of tissue repair: from wound healing to fibrosis. Int J Biochem Cell Biol 29: 5–17, 1997. 326. Nadal-Ginard B, Kajstura J, Anversa P, and Leri A. A matter of life and death: cardiac myocyte apoptosis and regeneration. J Clin Invest 111: 1457–1459, 2003. 327. Nadal-Ginard B, Kajstura J, Leri A, and Anversa P. Myocyte death, growth, and regeneration in cardiac hypertrophy and failure. Circ Res 92: 139 –150, 2003. 328. Nadin BM, Goodell MA, and Hirschi KK. Phenotype and hematopoietic potential of side population cells throughout embryonic development. Blood 102: 2436 –2443, 2003. 329. Nakamura T and Schneider MD. The way to a human’s heart is through the stomach: visceral endoderm-like cells drive human embryonic stem cells to a cardiac fate. Circulation 107: 2638 –2639, 2003. 330. Narmoneva DA, Vukmirovic R, Davis ME, Kamm RD, and Lee RT. Endothelial cells promote cardiac myocyte survival and spatial reorganization: implications for cardiac regeneration. Circulation 110: 962–968, 2004. 331. Negrin RS, Atkinson K, Leemhuis T, Hanania E, Juttner C, Tierney K, Hu WW, Johnston LJ, Shizurn JA, Stockerl-Goldstein KE, Blume KG, Weissman IL, Bower S, Baynes R, Dansey R, Karanes C, Peters W, and Klein J. Transplantation of highly purified CD34⫹Thy-1⫹ hematopoietic stem cells in patients with metastatic breast cancer. Biol Blood Marrow Transplant 6: 262–271, 2000. 332. Ng IO, Chan K, Shek W, Lee JM, Fong DY, Lo C, and Fan S. Stem cells and liver disease: promise laced with confusion and intrigue. Hepatology 38: 989 –998, 2003. 333. Nocka K, Majumder S, Chabot B, Ray P, Cervone M, Bernstein A, and Besmer P. Expression of c-kit gene products in known cellular targets of W mutations in normal and W mutant mice: evidence for an impaired c-kit kinase in mutant mice. Genes Dev 3: 816 – 826, 1989. 334. Norol F, Merlet P, Isnard R, Sebillon P, Bonnet N, Cailliot C, Carrion C, Ribeiro M, Charlotte F, Pradeau P, Mayol JF, Peinnequin A, Drouet M, Safsafi K, Vernant JP, and Herodin 1412 354. 355. 356. 357. 358. 360. 361. 362. 363. 364. 365. 366. 367. 368. 369. 370. 371. 372. 373. 374. pressure overload-induced right ventricular hypertrophy in rats. Circ Res 63: 648 – 657, 1988. Olson EN and Schneider MD. Sizing up the heart: development redux in disease. Genes Dev 17: 1937–1956, 2003. Onishchenko NA, Potapov IV, Bashkina LV, Krasheninnikov ME, Zaidenov VA, and Avramov PV. Recovery of contractile function of cryodamaged rat myocardium after transplantation of fetal cardiomyocytes and predifferentiation bone marrow stromal cells. Bull Exp Biol Med 138: 357–360, 2004. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, Pickel J, McKay R, Nadal-Ginard B, Bodine DM, Leri A, and Anversa P. Bone marrow cells regenerate infarcted myocardium. Nature 410: 701–705, 2001. Orlic D, Kajstura J, Chimenti S, Limana F, Jakoniuk I, Quaini F, Nadal-Ginard B, Bodine DM, Leri A, and Anversa P. Mobilized bone marrow cells repair in infarcted heart, improving function and survival. Proc Natl Acad Sci USA 98: 10344 –10349, 2001. Osawa M, Nakamura K, Nishi N, Takahasi N, Tokuomoto Y, Inoue H, and Nakauchi H. In vivo self-renewal of c Kit⫹ Sca-1⫹ Lin(low/-) hematopoietic stem cells. J Immunol 156: 3207–3214, 1996. Page DL, Caulfield JB, Kastor JA, and DeSanctis RW. Myocardial changes associated with cardiogenic shock. N Engl J Med 285: 133–137, 1971. Palermo AT, Labarge MA, Doyonnas R, Pomerantz J, and Blau HM. Bone marrow contribution to skeletal muscle: a physiological response to stress. Dev Biol 279: 336 –344, 2005. Palmer TD. Adult neurogenesis and the vascular Nietzsche. Neuron 34: 856 – 858, 2002. Palmer TD, Willhoite AR, and Gage FH. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol 425: 479 – 494, 2000. Pan GJ, Chang ZY, Scholer HR, and Pei D. Stem cell pluripotency and transcription factor Oct 4. Cell Res 12: 321–329, 2002. Pandur P. What does it take to make a heart? Biol Cell 97: 197–210, 2005. Paraguassu-Braga FH, Borojevic R, Bouzas LF, Barcinski MA, and Bonomo A. Bone marrow stroma inhibits proliferation and apoptosis in leukemic cells through gap junction-mediated cell communication. Cell Death Differ 10: 1101–1108, 2003. Park JY, Elshami AA, Amin K, Rizk N, Kaiser LR, and Albelda SM. Retinoids augment the bystander effect in vitro and in vivo in herpes simplex virus thymidine kinase/ganciclovir-mediated gene therapy. Gene Ther 4: 909 –917, 1997. Parsch D, Fellenberg J, Brummendorf TH, Eschlbeck AM, and Richter W. Telomere length and telomerase activity during expansion and differentiation of human mesenchymal stem cells and chondrocytes. J Mol Med 82: 49 –55, 2001. Pavlath GK and Blau HM. Expression of muscle genes in heterokaryons depends on gene dosage. J Cell Biol 102: 124 –130, 1986. Pera MF, Reubinoff B, and Trounson A. Human embryonic stem cells. J Cell Sci 113: 5–10, 2000. Pera MF and Trounson AO. Human embryonic stem cells: prospects for development. Development 131: 5515–5525, 2004. Perez-Moreno M, Jamora C, and Fuchs E. Sticky business: orchestrating cellular signals at adherens junctions. Cell 112: 535– 548, 2003. Perin EC, Dohmann HF, Borojevic R, Silva SA, Sousa AL, Mesquita CT, Rossi MI, Carvalho AC, Dutra HS, Dohmann HJ, Silva GV, Belem L, Vivacqua R, Rangel FO, Esporcatte R, Geng YJ, Vaughn WK, Assad JA, Mesquita ET, and Willerson JT. Transendocardial, autologous bone marrow cell transplantation for severe, chronic ischemic heart failure. Circulation 107: 2294 –2302, 2003. Perin EC, Dohmann HF, Borojevic R, Silva SA, Sousa AL, Silva GV, Mesquita CT, Belem L, Vaughn WK, Rangel FO, Assad JA, Carvalho AC, Branco RV, Rossi MI, Dohmann HJ, and Willerson JT. Improved exercise capacity and ischemia 6 and 12 mo after transendocardial injection of autologous bone marrow mononuclear cells for ischemic cardiomyopathy. Circulation 110: 213–218, 2004. Petersen BE, Goff JP, Greenberger JS, and Michalopoulos GK. Hepatic oval cells express the hematopoietic stem cell marker Thy-1 in the rat. Hepatology 27: 433– 445, 1998. Physiol Rev • VOL 375. Petrich BG, Molkentin JD, and Wang Y. Temporal activation of c-Jun N-terminal kinase in adult transgenic heart via cre-loxPmediated DNA recombination. FASEB J 17: 749 –751, 2003. 376. Petronczki M and Simos MF. Menagequatadre: molecular biology of chromosome segregation meiosis. Cell 112: 423– 440, 2003. 377. Pfeffer MA and Braunwald E. Ventricular remodeling after myocardial infarction. Experimental observations and clinical implications. Circulation 81: 1161–1172, 1990. 378. Pfeffer MA, Pfeffer JM, Steinberg C, and Finn P. Survival after an experimental myocardial infarction: beneficial effects of longterm therapy with captopril. Circulation 72: 406 – 412, 1985. 379. Pfeffer MA, Lamas GA, Vaughan DE, Parisi AF, and Braunwald E. Effect of captopril on progressive ventricular dilatation after anterior myocardial infarction. N Engl J Med 319: 80 – 86, 1988. 380. Pfeffer JM, Pfeffer MA, and Braunwald E. Influence of chronic captopril on the infarcted left ventricle of the rat. Circ Res 57: 84 –95, 1985. 381. Phinney DG, Kopen G, Righter W, Webster S, Tremain N, and Prockop DJ. Donor variation in the growth properties and osteogenic potential of human marrow stromal cells. J Cell Biochem 75: 424 – 436, 1999. 382. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, and Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science 284: 143–147, 1999. 383. Pittenger MF and Martin BJ. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ Res 95: 9 –20, 2004. 384. Pomerantz J and Blau HM. Nuclear reprogramming: a key to stem cell function in regenerative medicine. Nat Cell Biol 6: 810 – 816, 2004. 385. Poss KD, Wilson LG, and Keating MT. Heart regeneration in zebrafish. Science 298: 2188 –2190, 2002. 386. Potten CS, Booth C, Tudor GL, Booth D, Brady G, Hurley P, Ashton G, Clarke R, Sakakibara S, and Okano H. Identification of a putative intestinal stem cell and early lineage marker; musashi-1. Differentiation 71: 28 – 41, 2003. 387. Potten CS, Owen G, and Booth D. Intestinal stem cells protect their genome by selective segregation of template DNA strands. J Cell Sci 115: 2381–2388, 2003. 388. Poulsom R, Alison MR, Cook T, Jeffery R, Ryan E, Forbes SJ, Hunt T, Wyles S, and Wright NA. Bone marrow stem cells contribute to healing of the kidney. J Am Soc Nephrol 14: S48 –S54, 2003. 389. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 276: 71–74, 1997. 390. Prockop DJ, Gregory CA, and Spees JL. One strategy for cell and gene therapy: harnessing the power of adult stem cells to repair tissues. Proc Natl Acad Sci USA 100: 11917–11923, 2003. 391. Quaini F, Urbanek K, Beltrami AP, Finato N, Beltrami CA, Nadal-Ginard B, Kajstura J, Leri A, and Anversa P. Chimerism of the transplanted heart. N Engl J Med 346: 5–15, 2002. 392. Quesenberry PJ, Abedi M, Aliotta J, Colvin G, Demers D, Dooner M, Greer D, Hebert H, Menon MK, Pimentel J, and Paggioli D. Stem cell plasticity: an overview. Blood Cells Mol Dis 32: 1– 4, 2004. 393. Rakusan K. Assessment of Cardiac Growth. Growth of the Heart in Health and Disease. New York: Raven, 1984. 394. Reik W, Dean W, and Walter J. Epigenetic reprogramming in mammalian development. Science 293: 1089 –1093, 2001. 395. Reinecke H, Zhang M, Bartosek T, and Murry CE. Survival, integration, and differentiation of cardiomyocyte grafts: a study in normal and injured rat hearts. Circulation 100: 193–202, 1999. 396. Reith AD, Rottapel R, Giddens E, Brady C, Forrester L, and Bernstein A. W mutant mice with mild or severe developmental defects contain distinct point mutations in the kinase domain of the c-kit receptor. Genes Dev 4: 390 – 400, 1990. 397. Reubinoff BE, Pera M, Fong CY, Trounson A, and Bongso A. Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro. Nat Biotech 18: 399 – 404, 2000. 398. Reyes M, Lund T, Lenvik T, Aguiar D, Koodie L, and Verfaillie CM. Purification and ex vivo expansion of postnatal human marrow mesodermal progenitor cells. Blood 98: 2615–2625, 2001. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 359. LERI, KAJSTURA, AND ANVERSA 1413 ADULT STEM CELLS Physiol Rev • VOL 419. 420. 421. 422. 423. 424. 425. 426. 427. 428. 429. 430. 431. 432. 433. 434. 435. 436. 437. 438. 439. growth responses in isolated adult rat hearts. Evidence for loadindependent induction of cardiac protein synthesis by angiotensin II. Circ Res 76: 489 – 497, 1995. Schwartz RE, Reyes M, Koodie L, Jiang Y, Blackstad M, Lund T, Lenvik T, Johnson S, Hu WS, and Verfaillie CM. Multipotent adult progenitor cells from bone marrow differentiate into functional hepatocyte-like cells. J Clin Invest 109: 1291–1302, 2002. Schwartz RS and Curfman GD. Can the heart repair itself? N Engl J Med 346: 2– 4, 2002. Seiler C, Pohl T, Wustmann K, Hutter D, Nicolet PA, Windecker S, Eberli FR, and Meier B. Promotion of collateral growth by GM-CSF in patients with coronary artery disease: a randomised double blind placebo-controlled study. Circulation 104: 2012–2015, 2001. Sekiya I, Colter DC, and Prockop DJ. BMP-6 enhances chondrogenesis in a subpopulation of human marrow stromal cells. Biochem Biophys Res Commun 284: 411– 418, 2001. Sekiya I, Larson BL, Smith JR, Pochampally R, Cui JG, and Prockop DJ. Expansion of human adult stem cells from bone marrow stroma: conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells 20: 530 –541, 2002. Shake JG, Gruber PJ, Baumgartner WA, Senechal G, Meyers J, Redmond JM, Pittenger MF, and Martin BJ. Mesenchymal stem cell implantation in a swine myocardial infarcted model: engraftment and functional effects. Ann Thorac Surg 73: 1919 – 1925, 2002. Shay JW and Wright WE. Hayflick, his limit, and cellular ageing. Nat Rev Mol Cell Biol 1: 72–76, 2000. Shen CN, Horb ME, Slack JM, and Tosh D. Transdifferentiation of pancreas to liver. Mech Dev 120: 107–116, 2003. Shen Q, Goderie S, Jin L, Karanth N, Sun Y, Abramova N, Vincent P, Pumiglia K, and Temple S. Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science. In press. Sherr CJ and DePinho RA. Cellular senescence: mitotic clock or culture shock? Cell 102: 407– 410, 2000. Shinohara T, Orwig KE, Avarbock MR, and Brinster RL. Remodeling of the postnatal mouse testis is accompanied by dramatic changes in stem cell number and niche accessibility. Proc Natl Acad Sci USA 98: 6186 – 6191, 2001. Shiraishi I, Melendez J, Ahn Y, Skavdahl M, Murphy E, Welch S, Schaefer E, Walsh K, Rosenzweig A, Torella D, Nurzynska D, Kajstura J, Leri A, Anversa P, and Sussman MA. Nuclear targeting of Akt enhances kinase activity and survival of cardiomyocytes. Circ Res 94: 884 – 891, 2004. Shizuru JA, Negrin RS, and Weissman IL. Hematopoietic stem and progenitor cells: clinical and preclinical regeneration of the hematolymphoid system. Annu Rev Med 56: 509 –538, 2005. Sieber-Blum M. Cardiac neural crest stem cells. Anat Rec 276: 34 – 42, 2004. Sigal SH, Rajvanshi P, Gorla GR, Sokhi RP, Saxena R, Gebhard DR Jr, Reid LM, and Gupta S. Partial hepatectomy-induced polyploidy attenuates hepatocyte replication and activates cell aging events. Am J Physiol Gastrointest Liver Physiol 276: G1260 – G1272, 1999. Silva GV, Litovsky S, Assad JAR, Sousa ALS, Martin BJ, Vela D, Coulter SC, Lin J, Ober J, Vaughn WK, Branco RVC, Oliveira EM, He R, Geng YJ, Willerson JT, and Perin EC. Mesenchymal stem cells differentiate into an endothelial phenotype, enhance vascular density, and improve heart function in a canine chronic ischemia model. Circulation 111: 150 –156, 2005. Silver J and Miller JH. Regeneration beyond the glial scar. Nat Rev Neurosci 5: 146 –156, 2004. Siminovitch L, McCulloch EA, and Till JE. The distribution of colony-forming cells among spleen colonies. J Cell Physiol 62: 327–336, 1963. Skibbens RV and Hieter P. Kinetochores and the checkpoint mechanism that monitors for defects in the chromosome segregation machinery. Annu Rev Genet 32: 307–337, 1998. Slack JM and Tosh D. Transdifferentiation and metaplasia— switching cell types. Curr Opin Genet Dev 11: 581–586, 2001. Smith JR, Pochampally R, Perry A, Hsu SC, and Prockop DJ. Isolation of a highly clonogenic and multipotential subfraction of 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 399. Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ, and Collins AT. CD133, a novel marker for human prostatic epithelial stem cells. J Cell Sci 117: 3539 –3545, 2004. 400. Romanov YA, Svintsitskaya VA, and Smirnov VN. Searching for alternative sources of postnatal human mesenchymal stem cells: candidate MSC-like cells from umbilical cord. Stem Cells 21: 105– 110, 2003. 401. Rosenthal N. Prometheus’s vulture and the stem-cell promise. N Engl J Med 349: 267–274, 2003. 402. Roufosse CA, Direkze NC, Otto WR, and Wright NA. Circulating mesenchymal stem cells. Int J Biochem Cell Biol 36: 585–597, 2004. 403. Ruiz-Lozano P and Chien KR. Cre-constructing the heart. Nat Genet 33: 8 –9, 2003. 404. Rustom A, Saffrich R, Markovic I, Walther P, and Gerdes HH. Nanotubular highways for intercellular organelle transport. Science 303: 1007–1010, 2004. 405. Ryden M, Dicker A, Gotherstrom C, Astrom G, Tammik C, Arner P, and Le Blanc K. Functional characterisation of human mesenchymal stem cell-derived adipocytes. Biochem Biophys Res Commun 311: 391–397, 2003. 406. Ryu JK, Choi HB, Hatori K, Heisel RL, Pelech SL, McLarnon JG, and Kim SU. Adenosine triphosphate induces proliferation of human neural stem cells: role of calcium and p70 ribosomal protein S6 kinase. J Neurosci Res 72: 352–362, 2003. 407. Sadoshima J and Izumo S. The cellular and molecular response of cardiac myocytes to mechanical stress. Annu Rev Physiol 59: 551–571, 1997. 408. Sadoshima J, Qiu Z, Morgan JP, and Izumo S. Angiotensin II and other hypertrophic stimuli mediated by G protein-coupled receptors activate tyrosine kinase, mitogen-activated protein kinase, and 90-kD S6 kinase in cardiac myocytes. The critical role of Ca(2⫹)-dependent signaling. Circ Res 76: 1–15, 1995. 409. Sadoshima J, Malhotra R, and Izumo S. The role of the cardiac renin-angiotensin system in load-induced cardiac hypertrophy. J Card Fail 2: S1–S6, 1996. 410. Saegusa M, Takano Y, and Okudaira M. Human hepatic infarction: histopathological and postmortem angiological studies. Liver 13: 239 –245, 1993. 411. Saito T, Kuang JQ, Lin CC, and Chiu RC. Transcoronary implantation of bone marrow stromal cells ameliorates cardiac function after myocardial infarction. J Thorac Cardiovasc Surg 126: 114 –123, 2003. 412. Sammons J, Ahmed N, El-Sheemy M, and Hassan HT. The role of BMP-6, IL-6 and BMP-4 in mesenchymal stem cells-dependent bone development: effects on osteoblastic differentiation induced by parathyroid hormone and vitamin D3. Stem Cells Dev 13: 273– 280, 2004. 413. Sanchez-Ramos J, Song S, Cardozo-Pelaez F, Hazzi C, Stedeford T, Willing A, Freeman TB, Saporta S, Janssen W, Patel N, Cooper DR, and Sanberg PR. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp Neurol 174: 11–20, 2000. 414. Sarjeant JM, Butany J, and Cusimano RJ. Cancer of the heart: epidemiology and management of primary tumors and metastases. Am J Cardiovasc Drugs 3: 407– 421, 2003. 415. Schachinger V, Assmus B, Britten MB, Honold J, Lehmann R, Teupe C, Abolmaali ND, Vogl TJ, Hofmann WK, Martin H, Dimmeler S, and Zeiher AM. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction: final one-year results of the TOPCARE-AMI Trial. J Am Coll Cardiol 44: 1690 –1699, 2004. 416. Schinkel AH, Mayer U, Wagenaar E, Mol CA, van Deemter L, Smit JJ, van der Valk MA, Voordouw AC, Spits H, van Tellingen O, Zijlimans JM, Fibbe WE, and Borst P. Normal viability and altered pharmacokinetics in mice lacking mdr1-type (drug transporting) P-glycoproteins. Proc Natl Acad Sci USA 94: 4028 – 4033, 1997. 417. Schulinder M, Yanuka O, Itskovitz-Elder J, Melton D, and Benvenisty N. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc Natl Acad Sci USA 97: 11307–11312, 2000. 418. Schunkert H, Sadoshima J, Cornelius T, Kagaya Y, Weinberg EO, Izumo S, Riegger G, and Lorell BH. Angiotensin II-induced 1414 440. 441. 442. 443. 444. 445. 447. 448. 449. 450. 451. 452. 453. 454. 455. 456. 457. 458. 459. 460. 461. adult stem cells from bone marrow stroma. Stem Cells 22: 823– 831, 2004. Solloway MJ and Harvey RP. Molecular pathways in myocardial development: a stem cell perspective. Cardiovasc Res 58: 264 –277, 2003. Song X and Xie T. DE-cadherin-mediated cell adhesion is essential for maintaining somatic stem cells in the Drosophila ovary. Proc Natl Acad Sci USA 99: 14813–14818, 2002. Song X, Zhu CH, Doan C, and Xie T. Germline stem cells anchored by adherens junctions in the Drosophila ovary niches. Science 296: 1855–1857, 2002. Soonpaa MH and Field LJ. Survey of studies examining mammalian cardiomyocyte DNA synthesis. Circ Res 83: 15–26, 1998. Soonpaa MH, Koh GY, Klug MG, and Field LJ. Formation of nascent intercalated disks between grafted fetal cardiomyocytes and host myocardium. Science 264: 98 –101, 1994. Spangrude GJ, Heimfeld S, and Weissman IL. Purification and characterization of mouse hematopoietic stem cells. Science 241: 58 – 62, 1988. Spradling A, Drummond-Barbosa D, and Kai T. Stem cells find their niche. Nature 414: 98 –104, 2001. Srinivasan M, Sedmak D, and Jewell S. Effects of fixatives and tissue processing on the content and integrity of nucleic acids. Am J Pathol 161: 1961–1971, 2002. Stamm C, Kleine HD, Westphal B, Petzsch M, Kittner C, Nienaber CA, Freund M, and Steinhoff G. CABG and bone marrow stem cell transplantation after myocardial infarction. J Thorac Cardiovasc Surg 52: 152–158, 2004. Stamm C, Westphal B, Kleine HD, Petsch M, Kittner C, Klinge H, Schumichen C, Nienaber CA, Freund M, and Steinhoff G. Autologous bone marrow stem cell transplantation for myocardial regeneration. Lancet 361: 45– 46, 2003. Stenderup K, Justesen J, Clausen C, and Kassem M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone 33: 919 –926, 2003. Stocum DL. Limb regeneration: re-entering the cell cycle. Curr Biol 9: R644 –R646, 1999. Strauer BE, Brehm M, Zeus T, Kostering M, Hernandez A, Sorg RV, Kogler G, and Wernet P. Repair of infarcted myocardium by autologous intracoronary mononuclear bone marrow cell transplantation in humans. Circulation 106: 1913–1918, 2002. Strobel ES, Gay RE, and Greenberg PL. Characterization of the in vitro stromal microenvironment of human bone marrow. Int J Cell Cloning 4: 341–356, 1986. Styles JA. Measurement of ploidy and cell proliferation in the rodent liver. Enviorn Health Perspect 101 Suppl 5: 67–71, 1993. Subak-Sharpe H, Burk RR, and Pitts JD. Metabolic co-operation between biochemically marked mammalian cells in tissue culture. Rev Med Virol 12: 69 –78, 1969. Sun S, Guo Z, Xiao X, Liu B, Liu X, Tang PH, and Mao N. Isolation of mouse marrow mesenchymal progenitors by a novel and reliable method. Stem Cells 21: 527–535, 2003. Suva D, Garavaglia G, Menetrey J, Chapuis B, Hoffmeyer P, Bernheim L, and Kindler V. Non-haematopoietic human bone marrow contains long-lasting pluripotential mesenchymal stem cells. J Cell Physiol 198: 110 –118, 2004. Tai MH, Chang CC, Olson LK, and Trosko JE. Oct4 expression in adult human stem cells: evidence in support of the stem cell theory of carcinogenesis. Carcinogenesis 26: 495–502, 2005. Takada T, Suzuki Y, Kondo Y, Kadota N, Kobayashi K, Nito S, Kimura H, and Torii R. Monkey embryonic stem cell lines expressing green fluorescent protein. Cell Transplant 11: 631– 635, 2002. Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, Magner M, Isner J, and Asahara T. Ischemia and cytokineinduced mobilization of bone marrow-derived progenitor cells for neovascularization. Nat Med 5: 434 – 438, 1999. Takeda Y, Mori T, Imabayashi H, Kiyono T, Gojo S, Miyoshi S, Hida N, Ita M, Segawa K, Ogawa S, Sakamoto M, Nakamura S, and Umezawa A. Can the life span of human marrow stromal cells be prolonged by bmi-1, E6, E7, and/or telomerase without affecting cardiomyogenic differentiation? J Gene Med 6: 833– 845, 2004. Physiol Rev • VOL 462. Tang YL, Qian K, Zhang YC, Shen L, and Phillips MI. Mobilization of hematopoietic stem cells to ischemic myocardium by plasmid mediated stromal-cell-derived factor-1 alpha (SDF-1 alpha) treatment. Regul Pept 125: 1– 8, 2005. 463. Tauchi H, Sato T, and Ito Y. Morphological aspects of aging liver: half a century of progress. Arch Gerontol Geriatr 19: 135–144, 1994. 464. Tauchi H and Sato T. Changes in hepatic cell mitochondria during parabiosis between old and young rats. Mech Aging Dev 12: 7–14, 1980. 465. Tavassoli M and Takahashi K. Morphological studies on longterm culture of marrow cells: characterization of the adherent stromal cells and their interactions in maintaining the proliferation of hemopoietic stem cells. Am J Anat 164: 91–111, 1982. 466. Taylor DA, Atkins BZ, Hungspreugs P, Jones TR, Reedy MC, Hutcheson KA, Glower DD, and Kraus WE. Regenerating functional myocardium: improved performance after skeletal myoblast transplantation. Nat Med 4: 929 –933, 1998. 467. Taylor DA, Hruban R, Rodriguez ER, and GoldSchmidt-Clermont PJ. Cardiac chimerism as a mechanism for self-repair Does it happen and if so to what degree? Circulation 106: 2– 4, 2002. 468. Taylor MR and Bristow MR. The emerging pharmacogenomics of the beta-adrenergic receptors. Congest Heart Fail 10: 281–288, 2004. 469. Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y, Meyer EM, Morel L, Petersen BE, and Scott EW. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature 416: 542–545, 2002. 470. Teyssier-Le Discorde M, Prost S, Nandrot E, and Kirzenbaum M. Spatial and temporal mapping of c-kit and its ligand, stem cell factor expression during human embryonic haemopoiesis. Br J Haematol 107: 247–253, 1999. 471. Thiele J, Varus E, Wickenhauser C, Kvasnicka HM, Metz KA, and Beelan DW. Regeneration of heart muscle tissue: quantification of chimeric cardiomyocytes and endothelial cells following transplantation. Histol Histopathol 19: 201–209, 2004. 472. Thiele J, Varus E, Wickenhauser C, Kvasnicka HM, Lorenzen J, Gramley F, Metz KA, Rivero F, and Beelan DW. Mixed chimerism of cardiomyocytes and vessels after allogenic bone marrow and stem-cell transplantation in comparison with cardiac allografts. Transplantation 77: 1902–1905, 2004. 473. Theise ND and d’Inverno M. Understanding cell lineages as complex adaptive systems. Blood Cells Mol Dis 32: 17–20, 2004. 474. Theise ND, Krause DS, and Sharkis S. Comment on “Little evidence for developmental plasticity of adult hematopoietic stem cells.” Science 299: 1317, 2003. 475. Theise ND, Nimmakayalu M, Gardner R, Illei PB, Morgan G, Teperman L, Henegariu O, and Krause DS. Liver from bone marrow in humans. Hepatology 32: 11–16, 2000. 476. Theoharides TC, el-Mansoury M, Letourneau R, Boucher W, and Rozniecki JJ. Dermatitis characterized by mastocytosis at immunization sites in mast-cell-deficient W/Wv mice. Int Arch Allergy Immunol 102: 352–361, 1993. 477. Thomson JA, Eldor JI, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, and Jones JM. Embryonic stem cell lines derived from human blastocysts. Science 282: 1145–1147, 1998. 478. Thust R, Tomicic MT, Grabner R, Friedrichs C, Wutzler P, and Kaina B. Cytogenetic detection of a trans-species bystander effect: induction of sister chromatid exchanges in murine 3T3 cells by ganciclovir metabolized in HSV thymidine kinase gene-transfected Chinese hamster ovary cells. Mutagenesis 19: 27–33, 2004. 479. Till JE, McCulloch EA, and Siminovitch L. A stochastic model of stem cell proliferation, based on the growth of spleen colonyforming cells. Proc Natl Acad Sci USA 51: 29 –36, 1964. 480. Till JE, McCulloch EA, and Siminovitch L. Isolation of variant cell lines during serial transplantation of hematopoietic cells derived from fetal liver. J Natl Cancer Inst 33: 707–720, 1964. 481. Tjonnfjord GE, Steen R, Veiby OP, Morkid L, and Egeland T. Haemopoietic progenitor cell differentiation: flow cytometric assessment in bone marrow and thymus. Br J Haematol 91: 1006 – 1016, 1995. 482. Toda H, Tsuji M, Nakano I, Kobuke K, Hayashi T, Kasahara H, Takahashi J, Mizoguchi A, Houtani T, Sugimoto T, Hashimoto N, Palmer TD, Honjo T, and Tashiro K. Stem cell-derived neural 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 446. LERI, KAJSTURA, AND ANVERSA 1415 ADULT STEM CELLS 483. 484. 485. 487. 488. 489. 490. 491. 492. 493. 494. 495. 496. 497. 498. 499. 500. 501. 502. 503. Physiol Rev • VOL 504. Vitek JA. The behavior of cells during the evaluation of gap junctional intercellular communication by metabolic cooperation assay. Neoplasma 43: 119 –123, 1996. 505. Vogel W, Grunebach F, Messam CA, Kanz L, Brugger W, and Buhring HJ. Heterogeneity among human bone marrow-derived mesenchymal stem cells and neural progenitor cells. Haematologica 88: 126 –133, 2003. 506. Vulliet PR, Greeley M, Halloran SM, MacDonald KA, and Kittleson MD. Intra-coronary arterial injection of the mesenchymal stromal cells and microinfarction in dogs. Lancet 363: 783–784, 2004. 507. Wagers AJ, Sherwood RI, Christensen JL, and Weissman IL. Little evidence for developmental plasticity of adult hematopoietic stem cells. Science 297: 2256 –2259, 2002. 508. Wagers AJ and Weissman IL. Plasticity of adult stem cells. Cell 116: 639 – 648, 2004. 509. Wang JF, Yang Y, Wang G, Min J, Sullivan MF, Ping P, Xiao YF, and Morgan JP. Embryonic stem cells attenuate viral myocarditis in murine models. Cell Transplant 11: 753–758, 2002. 510. Wang X, Willenbring H, Akkari Y, Torimaru Y, Foster M, Al-Dhalimy M, Lagasse E, Finegold M, Olson S, and Grompe M. Cell fusion is the principal source of bone-marrow-derived hepatocytes. Nature 422: 897–901, 2003. 511. Ward J. Complexity of damage produced by ionizing radiation. Cold Spring Harb Symp Quant Biol 65: 377–382, 2000. 512. Watanabe K, Abe H, Mishima T, Ogura G, and Suzuki T. Polyangitis overlap syndrome: a fatal case combined with adult Henoch-Schonlein purpura and polyarteritis nodosa. Pathol Int 53: 569 –573, 2003. 513. Watt FM and Hogan BLM. Out of Eden: stem cells and their niches. Science 287: 1427–1438, 2000. 514. Weber KT. Aldosterone in congestive heart failure. N Engl J Med 345: 1689 –1697, 2001. 515. Weimann JM, Johansson CB, Trejo A, and Blau HM. Stable reprogrammed heterokaryons form spontaneously in Purkinje neurons after bone marrow transplant. Nat Cell Biol 5: 959 –966, 2003. 516. Weissman IL, Anderson DJ, and Gage F. Stem and progenitor cells: origins, phenotypes, lineage commitments, and transdifferentiations. Annu Rev Cell Dev Biol 17: 387– 403, 2001. 517. Weissman IL. The road ended up at stem cells. Immunol Rev 185: 159 –174, 2002. 518. Wells WA. Is transdifferentiation in trouble? J Cell Biol 157: 15–18, 2002. 519. Wenzel F, Dittrich M, Hescheler J, and Grote J. Hypoxia influences generation and propagation of electrical activity in embryonic cardiomyocyte clusters. Comp Biochem Physiol A Mol Integr Physiol 132: 111–115, 2002. 520. Wessels A and Perez-Pomares JM. The epicardium and epicardially derived cells (EPDC’s) as cardiac stem cells. Anat Rec 276A: 43–57, 2004. 521. White JG, Amos WB, and Fordham M. An evaluation of confocal versus conventional imaging of biological structures by fluorescent light microscopy. J Cell Biol 105: 41– 48, 1987. 522. Willenbring H and Grompe M. Delineating the hepatocyte’s hematopoietic fusion partner. Cell Cycle 3: 1489 –1491, 2004. 523. Wills SE. Federal funding of human embryonic stem cell research—illegal, unethical and unnecessary. J Contemp Health Law Policy 18: 95–145, 2001. 524. Winn N, Paul A, Musaro A, and Rosenthal N. Insulin-like growth factor isoforms in skeletal muscle aging, regeneration, and disease. Cold Spring Harb Symp Quant Biol 67: 507–518, 2002. 525. Wobus AM, Guan K, and Pich U. In vitro differentiation of embryonic stem cells and analysis of cellular phenotypes. Methods Mol Biol 158: 263–286, 2002. 526. Wolgemuth DJ, Laurion E, and Leile KM. Regulation of the mitoic and meiotic cell cycles in male germ life. Rec Prog Horm Res 57: 75–101, 2002. 527. Wollert KC and Drexler H. Clinical applications of stem cells in the heart. Circ Res 96: 151–136, 2005. 528. Wollert KC, Meyer GP, Lotz J, Ringes-Lichtenberg S, Lippolt P, Breidenbach C, Fichtner S, Korte T, Hornig B, Messinger D, Arseniev L, Hertenstein B, Ganser A, and Drexler H. Intracoronary autologous bone-marrow cell transfer after myocardial 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 486. stem/progenitor cell supporting factor is an autocrine/paracrine survival factor for adult neural stem/progenitor cells. J Biol Chem 278: 35491–35500, 2003. Tomita S, Ishida M, Nakatani T, Fukuhara S, Hisashi Y, Ohtsu Y, Suga Yutani C, Yagihara T, Yamada K, and Kitimura S. Bone marrow is a source of regenerated cardiomyocytes in doxorubicininduced cardiomyopathy and granulocyte colony-stimulating factor enhances migration of bone marrow cells and attenuates cardiotoxity of doxorubicin under electron microscopy. J Heart Lung Transplant 23: 577–584, 2004. Torella D, Rota M, Nurzynska D, Musso E, Monsen A, Shiraishi I, Zias E, Walsh K, Rosenzweig A, Sussman MA, Urbanek K, Nadal-Ginard B, Kajstura J, Anversa P, and Leri A. Cardiac stem cell and myocyte aging, heart failure, and insulin-like growth factor-1 overexpression. Circ Res 94: 514 –524, 2004. Tosh D, Shen CN, and Slack JM. Differentiated properties of hepatocytes induced from pancreatic cells. Hepatology 36: 534 – 543, 2002. Tosh D, Shen CN, and Slack JM. Conversion of pancreatic cells to hepatocytes. Biochem Soc Trans 30: 51–55, 2002. Tosh D and Slack JM. How cells change their phenotype. Nat Rev Mol Cell Biol 3: 187–194, 2002. Tosh D and Strain A. Liver stem cells: prospects for clinical use. J Hepatol 42: S75–S84, 2005. Tsai MS, Lee JL, Chang YJ, and Hwang SM. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum Reprod 19: 1450 –1456, 2004. Tsonis PA. Regenerative biology: the emerging field of tissue repair and restoration. Differentiation 70: 397– 409, 2002. Tumbar T, Guasch G, Greco V, Blanpain C, Lowry WE, Rendl M, and Fuchs E. Defining the epithelial stem cell niche in skin. Science 303: 359 –363, 2004. Tzukerman M, Rosenberg T, Ravel Y, Reiter I, Coleman R, and Skorecki K. An experimental platform for studying growth and invasiveness of tumor cells within teratomas derived from human embryonic stem cells. Proc Natl Acad Sci USA 100: 13507–13512, 2003. Uchida N and Weissman IL. Searching for hematopoietic stem cells: evidence that Thy-1.1lo Lin-Sca-1⫹ cells are the only stem cells in C57BL/Ka-Thy-11 bone marrow. J Exp Med 175: 175–184, 1992. Udelson JE, Patten RD, and Konstam MA. New concepts in post-infarction ventricular remodeling. Rev Cardiovasc Med 4 Suppl 3: S3–S12, 2003. Unlisted authors. No consensus on stem cells. Nature 428: 587, 2004. Unlisted authors. Statistically significant. Nat Med 11: 1, 2005. Urbanek K, Quaini F, Tasca G, Torella D, Castaldo C, NadalGinard B, Leri A, Kajstura J, Quaini E, and Anversa P. Intense myocyte formation from cardiac stem cells in human cardiac hypertrophy. Proc Natl Acad Sci USA 100: 10440 –10445, 2003. Urbanek K, Torella D, Sheikh F, Nurzynska D, Silvestri F, Beltrami CA, Beltrami AP, Bussani R, Quaini F, Bolli R, Leri A, Kajstura J, and Anversa P. Myocardial regeneration by activation of multipotent cardiac stem cells in ischemic heart failure. Proc Natl Acad Sci USA 102: 8692– 8697, 2005. Van Bekkum DW. Phylogenetic aspects of tissue regeneration: role of stem cells: a concise overview. Blood Cells Mol Dis 32: 11–16, 2004. Van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, and Gage FH. Functional neurogenesis in the adult hippocampus. Nature 415: 1030 –1034, 2002. Vassilopoulous G, Wang PR, and Russell DW. Transplanted bone marrow regenerates liver by cell fusion. Nature 422: 901–904, 2003. Vescovi A, Gritti A, Cossu G, and Galli R. Neural stem cells: plasticity and their transdifferentiation potential. Cells Tissues Organs 171: 64 –76, 2002. Vitek JA. The biological conditions of assessment of ethylene glycol-induced inhibition of gap junctional intercellular communication by metabolic cooperation assay. Folia Biol 43: 109 –114, 1997. 1416 529. 530. 531. 532. 533. 534. 536. 537. 538. 539. 540. infarction: the BOOST randomized controlled clinical trial. Lancet 364: 141–148, 2004. Woodbury D, Schwartz EJ, Prockop DJ, and Black IB. Adult rat and human bone marrow stromal cells differentiate into neurones. J Neurosci Res 61: 364 –370, 2000. Wurmser AE and Gage FH. Stem cells: cell fusion causes confusion. Nature 416: 485– 487, 2002. Xie T, Kawase E, Kirilly D, and Wong MD. Intimate relationships with their neighbors: tales of stem cells in Drosophila reproductive systems. Dev Dyn 232: 775–790, 2005. Xie T and Spradling AC. A niche maintaining germ line stem cells in the Drosophila ovary. Science 290: 328 –330, 2000. Xie T and Spradling AC. Decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell 94: 251–260, 1998. Xu YJ, Chapman D, Dixon IM, Sethi R, Guo X, and Dhalla NS. Differential gene expression in infarct scar and viable myocardium from rat heart following coronary ligation. J Cell Mol Med 8: 85–92, 2004. Yang Y, Min JY, Rana JS, Ke Q, Cai J, Chen Y, Morgan JP, and Xiao YF. VEGF enhances functional improvement of post-infarcted hearts by transplantation of ESC-differentiated cells. J Appl Physiol 93: 1140 –1151, 2002. Yap AS and Kovacs EM. Direct cadherin-activated cell signaling: a view from the plasma membrane. J Cell Biol 160: 11–16, 2003. Ying QL, Nichols J, Evans EP, and Smith AG. Changing potency by spontaneous fusion. Nature 416: 545–548, 2002. Yoo JU, Barthel TS, Nishimura K, Solchaga L, Caplan AI, and Goldberg VM. The chondrogenic potential of human bone marrow-derived mesenchymal progenitor cells. J Bone Joint Surg 80: 1745–1767, 1998. Yoon YS, Wecker A, Heyd L, Park JS, Tkebuchava T, Kusano K, Hanley A, Scadova H, Qin G, Cha DH, Johnson KL, Aikawa R, Asahara T, and Losordo DW. Clonally expanded novel multipotent stem cells from human bone marrow regenerate myocardium after myocardial infarction. J Clin Invest 115: 326 –338, 2005. Yoshimoto M, Shinohara T, Heike T, Shiota M, Kanatsu-Shinohara M, and Nakahata T. Direct visualization of transplanted hematopoietic cell reconstruction in intact mouse organs indicates the presence of a niche. Exp Hematol 31: 733–740, 2003. Physiol Rev • VOL 541. Yuan S and Schoenwolf GC. Islet-1 marks the early heart rudiments and is asymmetrically expressed during early rotation of the foregut in the chick embryo. Anat Rec 260: 204 –207, 2000. 542. Yuasa S, Itabashi Y, Koshimizu U, Tanaka T, Sugimura K, Kinoshita M, Hattori F, Fukami S, Shimazaki T, Okano H, Ogawa S, and Fukuda K. Transient inhibition of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic stem cells. Nat Biotechnol 23: 607– 611, 2005. 543. Zammit P and Beauchamp J. The skeletal muscle satellite cell: stem cell or son of stem cell? Differentiation 68: 193–204, 2001. 544. Zhang M, Methot D, Poppa V, Fujio Y, Walsh K, and Murry CE. Cardiomyocyte grafting for cardiac repair: graft cell death and anti-death strategies. J Mol Cell Cardiol 33: 907–921, 2001. 545. Zhang S, Wang D, Estrov Z, Raj S, Willerson JT, and Yeh ETH. Both cell fusion and transdifferentiation account for the transformation of human peripheral blood CD34-positive cells into cardiomyocytes in vivo. Circulation 110: 3803–3807, 2004. 546. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M, Nakauchi H, and Sorrentino BP. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med 7: 1028 –1034, 2001. 547. Zhou S, Zong Y, Lu T, and Sorrentino BP. Hematopoietic cells from mice that are deficient in both Bcrp1/Abcg2 and Mdr1a/b develop normally but are sensitized to mitoxantrone. Biotechniques 35: 1248 –1252, 2003. 548. Zhou X, Quann E, and Gallicano GI. Differentiation of nonbeating embryonic stem cells into beating cardiomyocytes is dependent on downregulation of PKC beta and zeta in concert with upregulation of PKC epsilon. Dev Biol 255: 407– 422, 2003. 549. Zhu CH and Xie T. Clonal expansion of ovarian germline stem cells during niche formation in Drosophila. Development 130: 2579 –2588, 2003. 550. Zicha D, Dobbie IM, Holt MR, Monypenny J, Soong DY, Gray C, and Dunn GA. Rapid actin transport during cell protrusion. Science 300: 142–145, 2003. 551. Zile MR, Baicu CF, and Gaasch WH. Diastolic heart failure— abnormalities in active relaxation and passive stiffness of the left ventricle. N Engl J Med 350: 1953–1959, 2004. 552. Zuk PA, Zhu M, Ashjian P, Benhaim P, Henrick MH, and Fraser JK. Human adipose tissue as source of multipotent stem cells. Mol Cell Biol 13: 4279 – 4295, 2002. 85 • OCTOBER 2005 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.33.6 on May 2, 2017 535. LERI, KAJSTURA, AND ANVERSA