Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Quantum dot wikipedia , lookup
Quantum dot cellular automaton wikipedia , lookup
Hydrogen atom wikipedia , lookup
Quantum fiction wikipedia , lookup
Coherent states wikipedia , lookup
Symmetry in quantum mechanics wikipedia , lookup
Quantum computing wikipedia , lookup
Copenhagen interpretation wikipedia , lookup
Bell test experiments wikipedia , lookup
Bell's theorem wikipedia , lookup
Many-worlds interpretation wikipedia , lookup
History of quantum field theory wikipedia , lookup
Quantum group wikipedia , lookup
Quantum machine learning wikipedia , lookup
Roger Penrose wikipedia , lookup
Delayed choice quantum eraser wikipedia , lookup
Canonical quantization wikipedia , lookup
EPR paradox wikipedia , lookup
Interpretations of quantum mechanics wikipedia , lookup
Quantum key distribution wikipedia , lookup
Quantum state wikipedia , lookup
Quantum teleportation wikipedia , lookup
Quantum cognition wikipedia , lookup
Hidden variable theory wikipedia , lookup
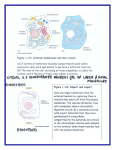
BME1450 – BIOENGINEERING SCIENCE (2003) 1 Quantum Collapse of Entangled Microtubules François Roy Abstract– It is almost inconceivable to think that the ordered firing patterns in neurons are the result of mere chance. For this reason, we propose that there is a higher order organisation which dictates the function of neurons in the brain. Recent evidence by Hameroff and Penrose [1] has revealed that microtubules found throughout the cytoskeleton are capable of information and memory processing. It is their periodic structure that makes them particularly well-suited for encoding information. Their ability to collapse a wavefunction (information) further reveals that microtubules may support micro sites of consciousness, and may therefore; provide the basement level of consciousness in the cognitive hierarchy. Hence, the entire interaction of entangled microtubules scattered throughout the brain may be responsible for initiating the cascade of reactions leading to the organized firing patterns in neurons. As a consequence, we further propose that quantum mechanics, along side systems biology, can be used to describe the biological interactions within the network of microtubules throughout the brain. Index Terms–Microtubules, quantum brain, entanglement, information processing, wave-function collapse I. INTRODUCTION We propose that biology is entangled in a web of wavefunctions and information. Just as chemistry can be used to describe interactions of enzymes and substrates, or membrane channels and ligands, we put forward that quantum mechanics can be used to explain biological interactions at the molecular level. Although these biological interactions are clearly not related through Aristotle’s idea of direct causality (stating that if A→B and B→C, then A→C), these interactions still exist through the phenomenon of entanglement. At the given time, we can think of entanglement as an interaction between elements that have at some time interacted with each other. We can treat entanglement as a dynamic relationship between different components of a system. Although these relationships are often not clearly defined in terms of direct causality, it is nonetheless important to keep their relationships in mind. For example, we can think of entanglement as the relationship between a boy and his dog. In this case, the boy’s future actions and interactions with other dogs, for example, are influenced by this relationship. I for one have never been particularly fond of dogs, but perhaps this is a Manuscript received October 21st, 2003. François Roy is with the Rehabilitation Engineering Laboratory of the Institute of Biomaterials and Biomedical Engineering at the University of Toronto, Toronto, ON, CANADA (email: [email protected]) consequence of my past experience as a young child when I was attacked by one. In essence, my current thoughts of dogs are the result of the entanglement of feelings of discontent and fear in the presence of dogs. Albeit this example may seem a little trivial at this time, the connection to system dynamics will become clearer as the central argument is developed. We will now delve into scientific research. Recent developments initially made by Hameroff and Penrose [1], have revealed that microtubules (MT) too are entangled with each other. In biological terms, microtubules are sub cellular protein polymers found throughout the cytoskeleton which participate in a wide variety of dynamic cellular processes. More importantly, microtubules are responsible for cellular architecture and modulate many dynamic functions throughout the cell. Of special interest is that microtubules have been found to participate in bioinformation processing such as learning and memory [2]. In fact, both microtubules along with DNA are unique cellular structures that exhibit a robust code system which dictates the function of processes throughout a cell [3]. Just as the arrangement of nucleotide bases on DNA forms the very basis for RNA transcription, and hence, protein synthesis throughout a cell, the periodic structure of microtubules makes them particularly well-suited to support the encoding of information [2]. More particularly, microtubules can be used for the superposition of coherent quantum states [3]. What this means is that microtubules can be used to extract information from many different signals, called wave-functions. For example, we can think of a MT as some type of measuring device similar to a measuring ruler. When a stick, in this case the wave-function, is presented to the microtubule, the microtubule measures and records the length of the stick, say 1 meter. Hence, the microtubule can support and extract information from the wave-function of the stick, and collapse the stick into its appropriate quantum state, length. The superposition of quantum states means that other properties of the stick can be measured and extracted at the same time. For example, we can measure the length, the diameter and circumference of the stick and superimpose the results simultaneously. However, if we try to use microtubules to support the quantum state of water, for example, then the microtubule would be a little perplexed at trying to measure the length of water. Of course, in a strict sense, length is really not a property of water! Hence, microtubules are not able to support this quantum state for water, and therefore will not encode any information. This analogy is of course the very basis which determines what information can be encoded. What is interesting about information and memory in microtubules is that the microtubules’ code system is strongly related to a “mental code” or quantum computing [3]. When a BME1450 – BIOENGINEERING SCIENCE (2003) state of an object, for example, the length of the stick, is recorded, the wave-function of the stick is collapsed into its quantum state, length, and yields a particular value of 1 meter. Hence, the value of 1 meter can be encoded and retrieved by the microtubules through the collapse of the wave-function. Moreover, at the moment of collapse, an organised quantum exocytosis occurs. i.e. the simultaneous emission of neurotransmitter molecules at the synaptic junction in neurons [4]. When the superposition of quantum states occurs, more states (or quantum states) related to the initial signal are collapsed and further enhance the probability of release of neurotransmitters at the synaptic junction. Importantly, according to Norwich [5], the increase in the firing rate of neurons is described as an increase in information transfer. As a consequence, this may be how a “mental order” may be translated into a physical reality through means of physiological action. This is certainly convenient, since Hebb [6] also found that an increase in the activity at the synaptic junction in neurons is responsible for improved neural communication and the emergence of learning in cells. We will now introduce the idea of non-locality to this framework. Non-locality refers to the idea of entanglement; which again was the simple analogy of the boy and his dog. Here, the idea of non-locality refers to the spatial relationship or better yet, the entangled relationship of different objects in space. At this time, we will use a more concrete example to describe the ideas of non-locality and entanglement. The idea of non-locality, debated by Einstein, Podolski, and Rosen (EPR-Paradox) [7] can be described using two chess-pieces. Suppose we were to place a black chess-piece in one hand and a white chess-piece in the other. With both hands behind our back, what if we were to ask someone to try to guess which hand holds the black piece. Of course, we can infer that the information in each hand is entangled with each other. Since we know that if someone picks the black piece in the right hand, we will be certain that the left hand contains the white piece. Now suppose that each piece was hidden again in our hands, and each of the hands was taken to different ends of the Universe. Could we still say that the chess-pieces are entangled? Of course we can, since we still don’t know where the black or white pieces are located. Therefore, the states are still completely entangled, and as a consequence, non-locality emerges. Einstein found it very disturbing to consider that a measurement of one of the entangled states in one location had an instantaneous effect on the other distant state. However, experiments in the 1980’s have confirmed that the non-local property of quantum mechanics exists [8]. Once again, non-locality refers to the presence of entangled states even in completely distant localities. This of course forms the very basis of this argument. As a wave-function is simultaneously collapsed by multiple microtubules, the information content of the signal is encoded by the entangled states of the microtubules, and hence, information can be encoded non-locally by the microtubules. Since wavefunctions interpreted by our senses can be collapsed by microtubules in many cells throughout the brain, we propose that information can be encoded and decoded by means of this 2 enormous biological network or system of microtubules. Moreover, the interaction of entangled microtubules dictates and governs the subsequent biological cascades in an organism, and hence, the firing patterns in neurons throughout the brain. II. FINDINGS A. Microtubule: structure and function The dynamic rearrangements of cytoplasmic structures, including the cytoskeleton, account for the changing shape, interaction, and motility of various cells [9]. The cytoskeleton composed of microtubules, actin filaments, and intermediate filaments are of fundamental importance for cellular function and survival. Parallel-arrayed microtubules are interconnected by cross-bridging proteins called microtubule-associated proteins (MAPs) [3]. Importantly, the MT-MAP complexes are responsible for regulating many dynamic functions of cells including mitosis, cell differentiation, and synaptic formation and function. It is usually understood that microtubules are ubiquitous throughout all of cellular biology [10]. Importantly, as DNA is the genetic library for hereditary information, microtubules are the real-time executives of the dynamic activities within living cells [3]. Fig. 1. (A) Structure of a microtubule. The tubulin dimers arranged throughout the the microtubule are comprised of α and β monomers [1]. (B) Model of the two state configurations of the tubulin protein. The states|α> (left) and |β> (right) are determined by the position of the electron on the tubulin subunit [1]. Microtubules are hollow crystalline cylindrical structures whose walls are made up of hexagonal lattices of tubulin dimers (Fig.1). Moreover, microtubules self-assemble to extend the axons and dendrites at synaptic connections of neurons [11]. Of importance, each tubulin dimer found on the microtubules can exist in one of two different geometrical conformations |α> or |β> (Fig. 2), [12]. Each of the |α> and |β> configurations corresponds to a different state of the dimer’s electric polarization. This occurs since an electron placed at the α-tubulin/β-tubulin junction may shift between the two different positions. This of course embodies the textbook case of a two-state quantum-mechanical system. BME1450 – BIOENGINEERING SCIENCE (2003) More careful study of the microtubules’ structure by Koruga [2] has revealed that microtubules possess a coding system. Importantly, it was found that microtubules possess the best known binary error-correcting codes called the 6-binary-dimer K1[13,26,5] [13]. It was also found that the conformations of the two microtubule states or code systems are strongly linked to protein function [3]. Theoretical models further suggest that conformational states of tubulin within microtubule lattices may interact with the six neighbouring tubulins to process, represent, and propagate information throughout the cells [14]. This mechanism has often been called a molecular computing system. B. Entanglement in single-celled organisms We can infer that the phenomenon of quantum entanglement of microtubules started thousands of years ago when prokaryotic cells became entangled through spirochetes, prokaryotes possessing a whip-like tail composed of microtubules [15]. Today, single-celled eukaryotic organism, like the amoeba and the paramecium appear to have adaptive or almost cognitive abilities; amoeba can search for food whereas paramecium can avoid obstacles [3]. If these singlecelled organisms completely lack a nervous system, then how can these adaptive or so-called cognitive abilities be possible? A logical explanation is that microtubules provide communication and information processing through the cell. According to experiments by Sherrington [16] almost half a century ago, the cytoskeleton is used for coordinated action in the form of metachronal waves, vibrations in the cytoskeleton. This can clearly be seen through movement or beating of the cilia (tail-like structure). Coincidently, this corresponds to a signal transduction in sensory cilia due to the propagation of changes in the conformations of microtubules [17]. C. Experiments on microtubules Many other experiments have revealed that microtubules are related to so-called cognitive function or information processing in the brain. Experiments with trained goldfish have revealed that the drug colchicine produces retrograde amnesia, by interfering with microtubules at the synaptic junctions [18]. Other experiments on baby chick brains have revealed that peak information processing through memory, learning, and experience occurs simultaneously with high levels of tubulin production and, hence, microtubule activity [19]. Similarly, baby rats have also shown a significant increase in tubulin production in the visual cortex when they first opened their eyes [20]. Furthermore, experiments on humans with neurological disorders including Alzheimer’s disease and schizophrenia have also revealed impaired functions in the microtubules and their associated proteins [21]. Together, these experiments are good indicators that microtubules are important in information processing in the brain. D. Effect of Anaesthesia on microtubules There also seems to be a connection between coherent oscillations in the microtubule conformational states and the emergence of consciousness [1]. It is a well-known phenomenon that the EEG of patients under general 3 anaesthesia persists despite that absence of consciousness. Nonetheless, it is rather remarkable that general anaesthesia can be induced by a number of different substances from chloroform to ether to xenon [22]. Studies on the mechanisms of anaesthetics have revealed that they act on the conformational functions of proteins, thereby blocking the changing states of microtubules from occurring [23]. The anaesthetic molecules disrupt the Val der Waals’ forces and prevent the normal configuration switching of tubulin from occurring. Furthermore, as the concentration of anaesthetics is decreased, the temporal Van der Waals forces blocking the electron flow between the tubulin proteins are removed, and the conformational changes within the microtubules are free to occur. This simple explanation of the puzzling features of anaesthetics further provides evidence that microtubules are perhaps micro sites of consciousness [24]. There is of course a cognitive hierarchy, and microtubules may provide the basement level of consciousness. III. DISCUSSION We have proposed that the wave-functions interpreted by the senses can be collapsed by non-local microtubules. The collapse of the wave-functions in the entangled states of the microtubules forms a so-called system of perceived information. By looking at the ElectroEncephaloGram (EEG), we would certainly expect that the asynchronous firing of multiple randomly distributed neurons would produce a zero net effect on the scalp electrodes. Instead, it has been found that the evoked potentials from the firing of particular non-local neural groups are radically different from the results from spontaneous random cortical activity [25]. This is certainly quite interesting, since it seems that the temporal rearrangement within neural groups can characterize the evoked potentials. Amazingly enough, by imposing the phase characteristics of the invoked potential on the spontaneous waveform, it is possible to reproduce the characteristic shape of the observed waveform i.e. regenerate the initial wavefunction [26]. As a consequence, it seems that the external stimulus, interpreted by the senses, does not only add energy to the brain, but yet it organises the signal in a coherent way. Hence, it provides a coherent superposition of the quantum states of the signal. This is certainly similar to the case of the stick in which we were able to superimpose the quantum states of length, radius, and circumference simultaneously. MRI scans can localize the spontaneous firing of distant neurons through the brain. It is intriguing to note that distant, nonadjacent regions of the brain can be activated during different types of thoughts. It is almost inconceivable to think of this as a result of pure chance, since our thoughts clearly make sense to us. For this reason, we propose that entanglement of microtubules during the collapse of wavefunctions forms the very basis of conscious thought. Indeed, conscious thought seems to correspond to temporal states in the brain with particular patterns of neural excitations. Conscious thoughts are integrated by the collapse of quantum states, described by the entangled microtubule networks. This of course is quite important since it would reveal that non- BME1450 – BIOENGINEERING SCIENCE (2003) local neural groups are in fact related and interact with each other. Hence, the firing pattern may be the result of the entangled states of the microtubules in the different neurons throughout the brain. It is, however, the entire interaction of the entangled microtubules which may activate neuronal synapses and initiate the cascade of reactions leading to consciousness [3]. The hierarchical structure is susceptible to quantum treatment, since it can be used to describe the particular dynamics of microtubule interactions. We may consider the two conformation states |α> and |β> of the tubulin dimers in the microtubules as the basic unit for this quantum biological system. Perhaps, we can even consider the entire brain as a single measuring device where all the cells and neurons are intrinsically related and constantly interacting through their entangled states. The collapse of a wave-function transmitted through the senses into an observable quantity, for example, the length of the stick, allows us to consciously “feel” that the stick has a length of 1 meter. It is an organized series of fluctuations in the conformations of microtubules which dictate our conscious thoughts. Microtubules provide the database or the storage of information from our senses, whereas quantum mechanical principles extract the information into an observable whole. The selection process for conscious thought is still completely unknown; nonetheless, we propose that the entangled states of the microtubules are the very mechanisms responsible for bookkeeping and storing information. IV. PROBLEMS AND ISSUES We must keep in mind that the physical evidence supporting the quantum collapse in entangled microtubules may sometimes look circumstantial. In theoretical applications, the hydrogen atom or even the electron in a vacuum is as far as we have been able to push the understanding of quantum mechanics. As a consequence, the explanation of quantum collapse in microtubules is a great leap from an isolated simplistic system to a completely in vivo process. Nonetheless, the manner in which evidence from completely disconnected fields fits together in such a coherent fashion may be more than simply mere chance. 4 understand how cellular processes occur and how elements of biology are related. Here, we propose that cellular components, more precisely microtubules, are intrinsically connected with each other through the presence of entanglement. In this case, we can assume that their dynamic relationships are beyond any classical or Newtonian dynamics. Nonetheless, this is a realistic assumption since cellular processes occur at the molecular level, and therefore, as these classical or Newtonian mechanics breakdown, tools at the atomic and subatomic levels must be introduced. Their relationships, as we have described, are certainly not as clearly defined as in more standard systems biology approaches. Nevertheless, they are equally and fundamentally important for understanding how biology truly works. REFERENCES [1] [2] [3] [4] [5] [6] [7] [8] [9] [10] [11] [12] [13] [14] V. CONCLUSION We propose that wave-functions interpreted by the senses are encoded by the microtubules in neurons throughout the brain. Information or memory intrinsically links microtubules with each other through the process of entanglement. The collapse of the wave-function by the microtubules entangles them in space and time, and hence, activation of the non-local entangled microtubules is the mechanism responsible for nonlocal firing patterns of neurons in the brain. Although the argument certainly walks a fine line between systems biology and quantum computing, we are convinced that the argument is an intrinsic part of both trains of thought. Systems biology is itself a framework for understanding and modelling the dynamics between the components of the system. The tools and approaches are constantly evolving, but the underlying principle remains the same; we wish to [15] [16] [17] [18] [19] [20] [21] [22] [23] [24] [25] [26] S. Hameroff and R. Penrose, ”Orchestrated reduction of quantum coherence in brain microtubules: A model for consciousness,” Mathematics and Computers in Simulation, Volume 40, Issues 3-4 (1996) 453-480. D. L. Koruga, Annals of the N.Y. Academy of Sciences, 466 (1986) 953. D.V. Nanopoulos, “THEORY OF BRAIN FUNCTION, QUANTUM MECHANICS AND SUPERSTRINGS,” presented at 15th Brazilian National Meeting on Particles and Fields, Angra dos Reis, Brazil, 5-8 Oct 1994 and at Physics Without Frontiers Four Seas Conf., Trieste, Italy, Jun 25 - Jul 1, 1995. Available: http://xxx.lanl.gov/PS_cache/hepph/pdf/9505/9505374.pdf J. C. Eccles, Proc. R. Soc. Lond. B227 (1986) 411. K. H. Norwich, Information, sensation, and perception, San Diego, CA: Academic Press, c1993. D. O. Hebb, The organization of behaviour, Wiley, New York, 1949. A. Einstein, B. Podolsky, and N. Rosen, Phys. Rev. 47 (1935) 777. A. Aspect and P. Grangier, “Experiments on EPR-type correlations with pairs of visible photons,” Quantum Concepts in Space and Time, edited by R. Penrose and C. J. Isham, Oxford University Press, 1986. S. R. Hameroff, Ultimate Computing: Biomolecular Consciousness and Nanotechnology, North-Holland, Amsterdam, 1987. P. Dustin, Microtubules (second edition), Springer-Verlag, New York, 1984. S. Hameroff, A. Nip, M. Porter, J. Tuszynski, “Conduction pathways in microtubules, biological quantum computation, and consciousness,” Biosystems. Jan;64(1-3): (2002),149-68. S. Hagan S, S. Hameroff, and J. A. Tuszynski JA, “Quantum computation in brain microtubules: decoherence and biological feasibility,” Phys Rev E Stat Nonlin Soft Matter Phys. June 65, (2002). N. J. Sloane, Scient. Amer. 250 (1984) 116. S.R. Hameroff, J.E. Dayhoff, R. Lahoz-Beltra, A. Samsonovich and S. Rasmussen, “Conformational automata in the cytoskeleton: Models for molecular computation,” IEEE Comput. (1992). L. Sagan, J. Theor. Biol. 14 (1967) 225. C. S. Sherrington, Man on his nature, Cambridge University Press, Cambridge (1951). J. Atema, J. Theor. Biol. 38 (1973/4) 181. J. Cronly-Dillon, D. Carden, and C. Birks, J. Exper. Biol. 61 (1974) 44. R. Mileusnic, S. P. Rose, and P. Tillson, Neuro. Chem. 34 (1980) 1007. D. Moshkov, et. al., Acta Histochemica Supp-Band. XLI (1992) 241. S. E. Arnold, V. M. Y. Lee, R. E. Gur and J. Q. Trojanowski, Proc. Nat. Acad. Sci. USA 88 (1991) 10850. A. Mershin, D. V. Nanopoulos, and E. Skoulakis. “QUANTUM BRAIN?,”ACT-08-00, CTP-TAMU-18-00, Jun 2000. Available: http://xxx.lanl.gov/PS_cache/quant-ph/pdf/0007/0007088.pdf N. P. Franks, and W. R. Lieb, Nature 316 (1985) 349-351. J. Ellis, N.E. Mavromatos and D.V. Nanopoulos, Phys. Lett. B293 (1992), 37. E. R. John, Science 177 (1972) 850. B. Sayers, H. A. Beagley, and W. R. Henshall, Nature 247 (1974) 481.