Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Mitogen-activated protein kinase wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
MTOR inhibitors wikipedia , lookup
Lipid signaling wikipedia , lookup
Signal transduction wikipedia , lookup
Biochemical cascade wikipedia , lookup
Blood sugar level wikipedia , lookup
Paracrine signalling wikipedia , lookup
Secreted frizzled-related protein 1 wikipedia , lookup
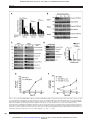
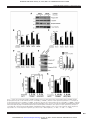
Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Cancer Research Molecular and Cellular Pathobiology Proliferation-Independent Control of Tumor Glycolysis by PDGFR-Mediated AKT Activation Cong Ran1,2, Huan Liu1,2, Yasuyuki Hitoshi1,2,3, and Mark A. Israel1,2 Abstract The differences in glucose metabolism that distinguish most malignant and normal tissues have called attention to the importance of understanding the molecular mechanisms by which tumor energy metabolism is regulated. Receptor tyrosine kinase (RTK) pathways that are implicated in proliferation and transformation have been linked to several aspects of tumor glucose metabolism. However, the regulation of glycolysis has invariably been examined under conditions in which proliferation is concomitantly altered. To determine whether RTKs directly regulate glycolysis without prerequisite growth modulation, we first identified a specific RTK signaling pathway, platelet-derived growth factor (PDGF)/PDGF receptor (PDGFR) that regulates glycolysis in glioma-derived tumor stem-like cells from a novel mouse model. We determined that PDGFregulated glycolysis occurs independent of PDGF-regulated proliferation but requires the activation of AKT, a known metabolic regulator in tumor. Our findings identifying a key characteristic of brain tumors, aerobic glycolysis, mediated by a pathway with multiple therapeutic targets suggests the possibility of inhibiting tumor energy metabolism while also treating with agents that target other pathways of pathologic significance. Cancer Res; 73(6); 1–13. 2013 AACR. Introduction Recent studies of glucose metabolism in malignant tissues have suggested that metabolic changes are key alterations contributing to tumorigenesis and tumor progression. Ongoing work seeks to link these changes to the genetic alterations underlying oncogenesis (1–4). Normal tissues use aerobic respiration to metabolize glucose in the presence of physiologic oxygen and glycolysis to metabolize glucose at times of oxygen deprivation. The Warburg effect refers to the finding that most malignant tissues, in contrast to normal tissues, used glycolysis as the main pathway to metabolize glucose even in the presence of sufficient oxygen (5). A hallmark of altered metabolism in cancer is increased glucose uptake. Virtually all high-grade glioma can be shown by clinical positron emission tomography (PET) to have enhanced glucose uptake, an aspect of aerobic glycolysis that has been widely studied in these tumors (6, 7). Growth factor receptor tyrosine kinases (RTK) implicated in tumorigenesis have been investigated for their role in tumor glucose metabolism due to Authors' Affiliations: 1Departments of Pediatrics and Genetics, Norris Cotton Cancer Center; 2Geisel School of Medicine at Dartmouth, Hanover, New Hampshire; and 3Department of Neurosurgery Rosai Hospital, Yatsushiro, Kumamoto, Japan Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Corresponding Author: Mark A. Israel, Dartmouth Medical School, One Medical Center Drive, Lebanon, NH 03756. Phone: 603-653-3611; Fax: 603-653-9003; E-mail: [email protected] doi: 10.1158/0008-5472.CAN-12-2460 2013 American Association for Cancer Research. their structural similarities to the insulin receptor, another RTK that has a well-defined role in regulating glucose homeostasis (8, 9). The use of common effector molecules by the insulin receptor and other RTKs suggests that RTKs may regulate tumor metabolism and thereby contribute to oncogenesis (4, 10). Although the EGF receptor (EGFR) activating a rarely expressed glucose receptor has been identified in prostate cancer cells and phosphorylation of pyruvate kinase M2 by the fibroblast growth factor receptor (FGFR) has been reported (11, 12), it is not known whether any RTKs directly regulate aerobic glycolysis in tumors. The typically synchronous changes in proliferation and metabolism that occur in tumors raises the possibility that RTK-mediated alterations in proliferation results in changes in glucose metabolism. Understanding whether regulation of proliferation and glycolysis by RTKs are independently mediated by RTK signaling cascades is essential for deciphering the mechanisms underlying the reprogramming of glucose metabolism that occurs so frequently in most tumor types. Considerable evidence points to an important role for platelet-derived growth factor (PDGF) signaling through the PDGF receptor (PDGFR), an RTK in the pathogenesis of glioma, particularly in mediating tumor cell proliferation (13–15). The Cancer Genome Atlas Network (TCGA) recently proposed a molecular classification of glioblastoma multiforme (GBM), the highest pathologic grade of glioma, and the most common brain tumor of adults (16, 17). While alterations in the PDGF/ PDGFR-mediated growth pathway were observed in all 4 TCGA-described subtypes of glioma, such alterations were identified at a much higher rate in the "proneural" subtype. Amplification of DNA encoding the PDGFRa and high levels www.aacrjournals.org Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF1 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. of PDGFRA gene expression were seen almost exclusively in this proneural subtype (17). A second signature genetic alteration of the proneural subtype of glioma was mutation of the IDH1 gene, which encodes isocitrate dehydrogenase 1, a mitochondrial enzyme of the Krebs cycle (17). Importantly, while mutation of IDH1 is found almost exclusively in GBM that can be classified as proneural, very few such GBMs had alterations of the PDGF/PDGFR pathway and most proneural glioma that had evidence of altered PDGF/PDGFR signaling did not bear mutated IDH1 (17). This dichotomy suggested to us that studying the role of an RTK, namely PDGFR, in regulating glioma glucose metabolism might be productively pursued in proneural glioma with normal IDH1 and activation of the PDGF/PDGFR growth-stimulatory pathway. Our laboratory has described a mouse model of human glioma based on overexpression, human PDGFB (hPDGFB) in tissues such as neural stem cells (NSC) that express glial fibrillary acidic protein (GFAP; ref. 18). In this mouse model, GFAP-mediated expression of hPDGFB is required for gliomagenesis and tumor progression and can be regulated by the administration of tetracycline, which inhibits hPDGFB expression. Using this unique mouse model, we examined the role of PDGF in regulating glycolysis in cultures enriched for gliomaderived tumor stem-like cells (TSC; ref. 19), a cellular subpopulation thought to be critical for tumor growth and progression. We found that in glioma-derived TSCs, PDGF signaling regulated the Warburg effect, aerobic glycolysis. We also observed that such regulation was independent of the effect of PDGF signaling on cellular proliferation. The phosphoinositide 3-kinase (PI3K)/AKT complex, a PDGFR downstream effector that has been recognized to mediate multiple activities of RTKs was found to be a key mediator of PDGF-regulated glycolysis in these glioma-derived TSCs. Materials and Methods OF2 lentivirus encoding short hairpin RNAs (shRNAs) complementary to PDGFRA (Santa Cruz Biotechnology) and Sh-scramble (ShScram), a lentivirus encoding a random set of 20 nucleotides, were used to generate cultures stably expressing these transgenes. Recombinant plasmids encoding hemaglutinnin (HA)-tagged AKT-DN (pCMV-HA-AKT-DN), myristolated AKT1 (HA-myr-AKT1), and control constructs (pCMV and PCDNA3; Addgene) were used to establish TSCs expressing these transgenes (20). Cell growth and viability TSCs (100,000 cells/6-well dish) were seeded, harvested, and counted by trypan blue exclusion. Cell viability was monitored by fluorescence-activated cell-sorting (FACS) analysis of propidium iodide (PI). Lactate production, glucose consumption, and glucose uptake measurement To examine lactate production and glucose consumption, cells were seeded in Dulbecco's' Modified Eagle's Media (DMEM):F12 without glutamine and 1B27 without insulin supplementation (5,000 cells/24-well dish). Total lactate secreted and glucose remaining in the conditioned media was assayed (Point Scientific, L7596 and G7521) on a plate reader (Molecular Devices Emax). Glucose consumption was determined by subtracting glucose in conditioned media from glucose in the plating media. Lactate and glucose measurements were normalized to cell numbers. To measure 2-NBDG uptake, TSCs were treated with doxycycline at 100 pg/mL for 18 hours, incubated DMEM containing 1 mmol/L 2-NBDG and 1 mmol/L of glucose for 1 hours, and evaluated by FACS (21) Glycolytic rate measurement TSCs (1.5 106 cells/10-cm dish) were incubated for 48 hours and the glycolytic rate measured (22). Cell cultures TSCs were isolated as previously described (18). Plasmid DNA with LipoD293 (SignaGen) was used for transfection. 0.4 mg/mL puromycin (Sigma) or 250 mg/mL G418 (Invitrogen) was added for selection after transfection and for a minimum of 10 days. Infection with lentiviruses and selection in 0.4 mg/ mL puromycin for a minimum of 7 days was used to prepared polyclonal shPDGFRA-expressing cultures. TSC-R cultures were established recurrent gliomas after 2 months of continuous subcutaneous doxycycline treatment. To establish doxycycline-resistant tumors from doxycyclinesensitive TSC xenografts, we injected 10,000 doxycycline-sensitive TSCs into nu/nu or syngeneic mice. Following tumor formation, animals were treated with oral doxycycline and tumors regressed. After 3 months of treatment, recurrent flank tumors were excised and cultured. All animal usage was in accordance with IACUC-approved protocols. Mitochondrial potential, intracellular ATP assay, and ADP/ATP ratio To determine mitochondrial membrane potential, 500,000 cells were collected, washed with PBS, and incubated in PBS [1% bovine serum albumin (BSA)/150 nmol/L tetramethylrhodamine, methyl ester (TMRM)] for 10 minutes before FACS analysis (23). To examine ATP, TSCs (25,000 cells/24-well dish) were seeded and incubated for 12 to 18 hours. Bioluminescence was used to determine ATP (PerkinElmer) and ADP/ATP ratio (Abcam, Ab65313). Materials PDGFR inhibitor AG1295 (Sigma), MEK1/2 inhibitor UO126 (Qiagen), and PI3K inhibitor LY294002 (Sigma) were diluted in dimethyl sulfoxide (DMSO) as indicated. Pooled recombinant Immunoblotting Cells lysed by radioimmunoprecipitation assay (RIPA) buffer were fractionated by 8% SDS-PAGE, transferred to polyvinylidene fluoride (PVDF; Millipore), and evaluated by Cancer Res; 73(6) March 15, 2013 Real-time PCR RNA was isolated (RNeasy Kit; Qiagen) and reverse transcribed using iScript (Bio-Rad). RNA was examined using iQ SYBR Green Supermix (Bio-Rad) and the MyIQ Real-Time PCR system (Bio-Rad). Expression was quantified after normalization using cyclophilin and 18S as controls. Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis Immunoblots using antibodies: phospho-PATHSCAN at 1:200 and anti-PDGFR, phospho-PDGFRa, AKT, phospho-AKT, ERK1/2, and phospho-ERK1/2 at 1:2,500 in 5% BSA/TBST (Cell Signaling); anti-Glut1 antibody at 1:200 in 5% milk/TBST (Abcam); anti-b-actin at 1:2,000 in 5% milk/TBST (Sigma). Densitometric measurement of band density was completed using ImagePro software on scanned blots. Statistics Data are representative of 3 or more independent experiments, presented as mean SEM. We used the 2-tailed Student t test to evaluate differences between 2 groups. P < 0.05 was designated statistically significant. For experiments involving multiple comparisons, we conducted one-way ANOVA and Tukey post hoc test to evaluate differences. Statistical analyses were completed using GraphPad Prism software. Results Modulation of PDGF signaling in glioma-derived TSCs To model human glioma, we described a GFAP-tTA:TREhPDGFB mouse model overexpressing hPDGFB that can be regulated by doxycycline (18). To extend these studies, we established cultures of glioma-derived TSCs from central nervous system (CNS) glioma arising in these mice (14, 18). To characterize the in vitro regulation of the hPDGFB transgene by doxycycline, we incubated cultures from 3 individual tumors and found that hPDGFB mRNA levels decreased in a dose-dependent manner (Fig. 1A). Because PDGFR is activated by phosphorylation, we examined whether inhibition of hPDGFB expression by doxycycline was associated with a decrease in PDGFR phosphorylation (Fig. 1B). We found inhibition of PDGFR phosphorylation and examined the effect of doxycycline on activation of other kinases downstream of PDGFR. Levels of activated, phosphorylated SHP2, AKT, and ERK1/2, and total ERK1/2 expression were decreased when hPDGFB expression was inhibited (Fig. 1C). To alternatively modulate PDGFR, we treated these TSCs with AG1295, a pharmacologic inhibitor of PDGFR (24). We found decreased PDGFR phosphorylation and decreased activation of AKT and ERK1/2 (Fig. 1D). As predicted from our previously published work examining doxycycline treatment of GFAP-tTA:TRE-hPDGFB mice (18), we observed growth inhibition with doxycycline at concentrations that did not significantly alter cell survival (Fig. 1E, Supplementary Fig. S1). Also, proliferation of TSCs was inhibited by AG1295 (Fig. 1F and data not shown). We named these TSCs as doxycycline-sensitive: TSC-S1, TSC-S2, and TSC-S3. GFAP-tTA:TRE-hPDGFB TSCs exhibit altered glucose metabolism Amongst glioma of the proneural subtype, alterations of PDGF signaling and IDH1 mutations affecting glucose metabolism rarely occur in the same tumor (17). Glioma-derived TSCs did not have alterations in IDH1 function (personal communications, Dr. Hai Yan, Duke University, Durham, NC), and so we investigated PDGF regulation of glucose metabolism. We found that in addition to slightly enhanced growth compared with www.aacrjournals.org NSCs from wild-type animals (WT NSC; Fig. 2A), TSCs exhibited increased lactate production (Fig. 2B) and glucose consumption (Fig. 2C). These changes are emblematic in tumor cells exhibiting the Warburg effect. We then used 5-[3H]-glucose to measure their glycolytic rate (22). We found that the glycolytic rate was higher in TSCs than in WT NSCs (Fig. 2D). To exclude the possibility that enhanced glycolysis was secondary to defective mitochondria, we examined mitochondrial membrane potential in TSCs using TMRM, a cationic tetramethylrhodamine methyl ether (25). We found that mitochondrial membrane potential was not decreased in TSCs (Fig. 2E), indicating that increased glycolysis was not a result of defective mitochondria. To determine the contributions of mitochondrial activity and glycolysis to intracellular ATP, we inhibited mitochondrial phosphorylation in TSCs and WT NSCs using carbonyl cyanide m-chlorophenyl hydrozone (CCCP), an uncoupling agent, as well as, antimycin A, an electron transport inhibitor (12). We found that ATP in TSCs following inhibition of mitochondrial respiration remained significantly above the levels observed in treated NSCs (Fig. 2F), indicating that increased glycolysis contributed to the increased ATP in TSCs. The observation that decreased ATP levels resulting from treatment with these agents was similar in WT NSCs and TSCs suggests a similar contribution of mitochondrial respiration to ATP levels. We also surveyed mRNA expression of multiple enzymes in glucose metabolism and found that mRNA levels of hexokinase (HK) 1 and 2, lactate dehydrogenase A (LDHA), and Glut1 were elevated in TSCs. PDGF regulates glucose metabolism in glioma-derived TSCs In vascular smooth muscle cells, enhanced glycolytic flux and increased mitochondrial reserve are associated with PDGFR activation (26–28). Activation of PDGFR is a key oncogenic alteration leading to tumorigenesis, and we hypothesized that PDGFR activation contributed to the alterated glucose metabolism in TSCs (Fig. 2). To examine this, we first determined whether modulating hPDGFB expression and PDGFR signaling altered glucose metabolism. Doxycycline treatment diminished hPDGFB expression and decreased lactate production and glucose consumption (Fig. 3A and B). AG1295 inhibition of PDGFR caused a decrease in lactate production and glucose consumption (Fig. 3C and D). Findings in all 3 TSC cultures examined gave similar results (data not shown). We examined the glycolytic rate of TSCs following PDGF/ PDGFR inhibition. We observed approximately a 40% decrease in glycolytic rate in 2 independent TSC cultures following inhibition of PDGF/PDGFR by doxycycline (Fig. 3E), indicating PDGF-induced glucose uptake contributed to enhanced glycolysis. We then evaluated glucose uptake using the fluorescent glucose analogue, 2-[N-(7-nitrobenz-2-oxa-1,3-diaxol4-yl)amino]-2-deoxyglucose (2-NBDG), in TSCs (21). A decrease in glucose uptake was detectable by FACS following inhibition of PDGF/PDGFR (Fig. 3F). Also, the expression of HK1, HK2, and Glut1, molecules contributing glucose uptake (29), was enhanced by PDGF (Fig. 3G, Supplementary Fig. S2A Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF3 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. Figure 1. Doxycycline and PDGFR inhibitor, AG1295, effectively inhibit PDGF signaling in primary glioma-derived TSCs. TSC cultures, TSC-S1, S2, and S3, were derived from 3 independent mouse CNS gliomas (18). A, steady-state hPDGFB mRNA levels in TSCs following 36 hours of doxycycline treatments. B, immunoblot analysis of PDGFR phosphorylation in TSCs following 48 hours of doxycycline treatments. C, immunoblot analysis of PDGFRa, PDGFRb, SHP2, AKT, and ERK1/2 phosphorylation and AKT and ERK1/2 total expression in TSCs following 48 hours of doxycycline treatments. D, left, immunoblot analysis of PDGFR, AKT, and ERK1/2 activation in lysates of TSC-S1 following 48 hours of AG1295 treatments. Right, bar graph of immunoblot band density measured in triplicate normalized to actin. B–D, derived from multiple gels independently blotted and examined with indicated antibodies. E, growth curves of TSC-S1 following doxycycline treatment. F, growth curves of TSC-S1 following AG1295 treatments. Data in all panels are representative of 3 independent experiments and presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01). OF4 Cancer Res; 73(6) March 15, 2013 Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis Figure 2. Glioma-derived TSCs exhibit Warburg-like glucose metabolism. A, growth curves of WT NSCs and TSC-S1. Lactate production (B) and glucose consumption (C) of WT NSCs and TSC-S1. D, glycolytic rate of WT NSCs and TSCs. E, FACS analysis to evaluate mitochondrial membrane potential of WT NSCs and TSCs by TMRM. Left, histogram representation of TMRM intensity in WT NSC and TSCs (20,000 cells). Right, average median fluorescence of WT NSCs and TSCs. F, intracellular ATP in WT NSCs and TSC-S1 following treatment with CCCP (40 minutes) or antimycin A (40 minutes). G, steady-state mRNA levels of enzymes involved in glucose and lactate metabolism in WT NSCs and TSC-S1. Data in all panels are representative of 3 or more independent experiments, presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01; , P < 0.005). and S2B). These findings indicate that glucose influx is enhanced in TSCs by activation of PDGFR. The expression of LDHA and pyruvate dehydrogenase kinase 1 (PDK1), enzymes affecting the conversion of pyruvate to lactate, decreased following hPDGFB inhibition (Fig. 3G, Supplementary Fig. S2C), suggesting that PDGF signaling is associated with other changes enhancing glycolysis. We observed in TSCs intracellular ATP levels above those in corresponding normal cells (Fig. 2F), suggesting that PDGFB www.aacrjournals.org may regulate ATP in glioma-derived TSCs. We evaluated ATP following inhibition of PDGFB and found that following doxycycline treatment intracellular ATP was reduced (Fig. 4A, Supplementary Fig. S3A). This dramatic decrease in ATP was also observed in TSCs treated with AG1295 (Fig. 4B, Supplementary Fig. S3B). We then examined the ADP/ATP ratio in TSCs. Diminished PDGF signaling resulted in an increased ADP/ATP ratio (Fig. 4C). In addition, we examined activation of AMP-activated protein kinase (AMPK), an energy sensor Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF5 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. Figure 3. PDGF pathway regulation of glucose metabolism in glioma-derived TSCs. Lactate production (A) and glucose consumption (B) of TSCs following 96 hours of doxycycline treatments. Lactate production (C) and glucose consumption (D) of TSC-S1 following 96 hours of AG1295 treatments. E, glycolytic rate of TSCs in the presence (þ) or absence () of doxycycline treatment. F, FACS analysis of glucose uptake in TSC-S1 after 18 hours of doxycycline treatment (100 pg/mL) using 2-NBDG. Left, histogram representation of 2-NBDG intensity (20,000 cells). Right, quantification of FACS analysis. G, steady-state mRNA levels of TSC-S1 following 48 hours of doxycycline treatment (100 pg/mL). Data in all panels are representative of 3 or more independent experiments, presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01; , P < 0.005). activated by increased intracellular AMP:ATP ratio (30). We detected increasing levels of phosphorylated, activated AMPK when hPDGFB expression was inhibited (Fig. 4D) consistent with the decreased ATP following inhibition of PDGF/PDGFR (Fig. 4A and B, Supplementary Fig. S3). Decreased ATP following PDGF/PDGFR inhibition (Fig. 4A and B) could be due to decreased glycolysis or decreased OF6 Cancer Res; 73(6) March 15, 2013 mitochondrial function (27, 31). To characterize the contribution of these mechanisms to decreased ATP, we measured mitochondrial membrane potential before and after hPDGFB inhibition. No changes in membrane potential were observed following doxycycline (Fig. 4E), indicating that PDGF/PDGFR signaling does not inhibit mitochondrial function. Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis Figure 4. Intracellular ATP levels in doxycycline-sensitive TSCs are maintained by PDGF signaling. Intracellular ATP in TSCs following 18 hours of doxycycline treatments (A) or AG1295 treatments (B). C, ADP/ ATP ratio in TSCs following 18 hours of dox treatment. D, immunoblot analysis of AMPK a-subunit phosphorylation in TSCs following 24 hours of doxycycline treatments. E, FACS analysis of TSC-S1 following 24 hours of doxycycline treatment to evaluate mitochondrial membrane potential by TMRM. Left, histogram representation of TMRM intensity in TSCs (20,000 cells). Right, quantification of FACS analysis. All data are representative of 3 or more independent experiments and presented as mean SEM of triplicate ( , P < 0.05; , P < 0.01; , P < 0.005). PDGF regulation of glycolysis in TSCs is independent of its effect on proliferation To determine whether PDGF-regulated glycolysis was a consequence of changes in proliferation, we examined proliferation-arrested cells for changes in glucose metabolism. If hPDGFB regulates glycolysis, its effects on glucose metabolism would be independent of proliferation. We induced cell-cycle arrest of glioma TSCs in M phase with mitomycin C (MMC; ref. 32) and evaluated glycolysis in these cells (Supplementary Fig. S4A). We found that doxycycline-mediated decreases in hPDGFB expression resulted in decreased lactate production and glucose consumption (Supplementary Fig. S4B and S4C), www.aacrjournals.org suggesting that PDGFR signaling modulated glucose metabolism independent of cellular proliferation. To extend this observation (Supplementary Fig. S4), we determined whether PDGF-regulated glycolysis in TSCs isolated from GFAP-tTA:TRE-hPDGFB tumors resistant to the growth-inhibitory effects of decreased PDGF signaling. We previously observed that a small percentage of GFAP-tTA: TRE-hPDGFB glioma recurred in vivo despite continuous doxycycline treatment (Supplementary Fig. S5A and data not shown). Because in vivo recurrence was rare, we sought to develop a system in which recurrent tumors occur more frequently. We therefore established flank xenografts of Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF7 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. doxycycline-sensitive TSCs in syngeneic or nude mice (Supplementary Fig. S5B). Despite continuous doxycycline treatment, approximately 70% of these animals developed recurrent tumors. Three in vivo recurrent tumors and 3 xenografts that recurred after continuous doxycycline treatment, a total of 6 recurrent tumors, which were each resistant to doxycycline inhibition on tumor growth, were excised and propagated in sphere culture as TSCs. We found that these TSCs were not growth inhibited in vitro by doxycycline (Supplementary Fig. S5C and data not shown). We named these TSCs that are resistant to the antiproliferative effects of doxycycline TSC-R. TSC-R1, 2, and 3 were derived from recurrences of primary tumors and the TSC-R4, 5, and 6 were from the xenograft model. We evaluated TSC-R cells from these 6 independent tumors and found that PDGF expression remained responsive to doxycycline (data not shown), although the cells were not growth inhibited by doxycycline. Similarly, AG1295 did not affect proliferation in any of these cultures (Supplementary Fig. S5D). As in doxycycline-sensitive TSCs, inactivation of PDGF/ PDGFR signaling in doxycycline-resistant TSCs decreased lactate production and glucose consumption (Fig. 5A and B), without detectable changes in proliferation (Supplementary Fig. S5D). Also, we detected a decrease in intracellular ATP following PDGFR inhibition in these doxycycline-resistant TSCs (Fig. 5C) comparable with the decreases in doxycycline-sensitive TSCs (Fig. 4B). Finally, we observed a decrease in the glycolytic rate of doxycycline-resistant TSCs when PDGFR was inhibited (Fig. 5D), confirming that glycolysis was regulated by PDGF/PDGFR. To provide further evidence that these changes in glycolysis were mediated specifically by PDGFR, we inhibited the expression of surface receptor PDGFRA using a short hairpin (Sh) to diminish PDGFR. Two concentrations of recombinant retrovirus encoding shPDGFRA, 0.5 and 2 multiplicity of infection (moi), were used. In both instances phosphorylated and total PDGFRA decreased (Fig. 5E). Also, glioma-derived doxycycline-resistant TSCs infected with a lentivirus encoding shPDGFRA (0.5 and 2 moi) exhibited reduced lactate production (Fig. 5F), decreased glucose consumption (Fig. 5G), and decreased intracellular ATP (Fig. 5H). AKT is a key mediator of PDGF-regulated tumor glycolysis To pursue the mechanism by which PDGF regulates glycolysis in glioma TSCs, we evaluated downstream pathways including the mitogen-activated protein kinase (MAPK) and PI3K/AKT pathways that are modulated by PDGF/PDGFR signaling (10, 33). To evaluate the MAPK pathway, we used UO126 to inhibit MEK1/2 kinase activity (34). Treatment decreased phosphorylation of the MEK target, ERK1/2, although other downstream targets of PDGFR were unaffected (Fig. 6A). While MEK1/2 inhibition resulted in a decrease in proliferation (Fig. 6B), lactate production and glucose consumption at multiple time points after treatment exhibited no appreciable difference (Fig. 6C and D). We interpreted these data to indicate that OF8 Cancer Res; 73(6) March 15, 2013 MEK signaling did not mediate PDGF/PDGFR regulation of glycolysis. We then examined the possible role of PI3K/AKT signaling in glycolysis. We observed 2 classes of doxycycline-resistant TSCs: those in which the AG1295 inhibited AKT phosphorylation (Fig. 6E: TSC-R1, TSC-R5, and data not shown) and those in which AKT phosphorylation was not inhibited (Fig. 6E: TSCR4 and TSC-R6). Importantly, when AKT phosphorylation remained responsive to PDGFR activity as in TSC-R1, TSCR2, and TSC-R5, lactate production (Fig. 6F) and glucose consumption (Fig. 6G) were modulated by PDGFR; however, when AKT was unaffected by inhibition of PDGFR as it was in TSC-R4 and TSC-R6, there were no reproducible or significant changes in lactate production (Fig. 6F) or glucose consumption (Fig. 6G) following PDGF/PDGFR inhibition. This apparent association of AKT modulation and glycolysis suggests that AKT may mediate PDGF-regulated glucose metabolism. We therefore inhibited AKT activation in both doxycycline-sensitive and -resistant TSCs with a PI3K inhibitor, LY294002 (Fig. 7A). Following treatment, we observed significant decreases in glycolysis as determined by lactate production (Fig. 7B), glucose consumption (Fig. 7C), and the glycolytic rate of (Fig. 7D) This finding was representative of all other TSC cultures examined (data not shown). We then overexpressed in TSC-S1 cells a HA-tagged dominant-negative form of AKT (AKT-DN), which prevents AKT activation by blocking endogenous AKT from binding to PI3K (20). Cultures in which approximately 40% of the population was transfected with AKT-DN as estimated from the cytologic evaluation of cells transfected with a comparable GFP-expressing control vector exhibited decreased phosphorylation of a kinase downstream of AKT, S6-kinase (Supplementary Fig. S6A). In these cells, we observed a 30% to 40% decrease in lactate production (Supplementary Fig. S6B) and glucose consumption (Supplementary Fig. S6C). We then expressed a constitutively activated form of AKT1 in TSC-S3 (Fig. 7E–G) (35). We transiently expressed HA-tagged myr-AKT1 (HA-myrAKT1) in TSCs. Activated AKT can be evaluated by assaying the phosphorylation of downstream kinase p70-S6K (Fig. 7E; ref. 36). In cells transfected with constitutively activated AKT1, no increase in glycolysis was seen presumably because in the PDGFB-secreting TSCs, AKT is extensively activated (Fig. 1C). Adding doxycycline did not inhibit glycolysis, although it did inhibit glycolysis in cells lacking constitutively activated AKT1 (Fig. 7F and G). These findings indicate that activated AKT1 can rescue PDGF inhibition of glycolysis. Consistent with this interpretation is our finding that modulation of hPDGFB by doxycycline alters expression of AKT target genes important for glucose metabolism (Fig. 3G, Supplementary Fig. S2; refs. 21, 37). Discussion Altered glucose metabolism is a hallmark of cancer underlying PET imaging, which depends upon quantification of 2deoxy-2(18F)fluoro-D-glucose in tumors (6, 37). Such imaging distinguishes most tumors from corresponding normal tissues. Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis Figure 5. The PDGF pathway regulates glucose metabolism independent of proliferation. Lactate production (A) and glucose consumption (B) of TSCs following 96 hours of AG1295 (þ) or DMSO () treatment. C, intracellular ATP in TSCs after 24 hours of AG1295 (þ) or DMSO () treatment. D, glycolytic rate following 48 hours of AG1295 (þ) or DMSO () treatment. E, immunoblot analysis of PDGFR activation in TSC-R1 infected with sh-scramble (ShScram) and sh-PDGFRa (ShPDGFRA) lentiviruses. Lactate production (F) and glucose consumption (G) of TSC-S1 expressing ShScram or ShPDGFRA. H, intracellular ATP in TSC-R1 expressing shScram or ShPDGFRA. Data in all panels are representative of 3 or more independent experiments and presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01). Alterations in glucose metabolism are important for tumor progression (38). Mutations altering enzymatic activity of IDH1, succinate dehydrogenase, or fumarate hydratase, enzymes important for glucose metabolism, occur in a variety of tumors (39, 40). Also, ectopic expression of alternative isoforms of glycolytic enzymes that increase glycolysis, such as PKM2, HK2, and phosphofructose kinase 4, promote tumor www.aacrjournals.org cell proliferation, survival, and invasion affecting patient outcome (29, 39, 41). Further evidence suggesting an association between glucose metabolism and malignancy is the observation that oncogenic signaling molecules modulate expression of glycolytic enzymes enhancing glycolysis and proliferation (38). For example, stimulation of the MAPK pathway, a mediator of proliferation, Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF9 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. Figure 6. PI3K/AKT but not the MAPK pathway modulates tumor glycolysis in glioma-derived TSCs. A, immunoblot analysis of ERK1/2, AKT, and PDGFR phosphorylation in lysates of TSC-S1 following 48 hours of MEK1/2 inhibitor UO126 treatments. B, growth curves of TSC-S2 following UO126 treatments. Lactate production (C) and glucose consumption (D) of TSC-S2 after 96-hour UO126 treatments. E, immunoblot analysis of PDGFR and AKT phosphorylation in TSCs following 48 hours of AG1295 treatments. Lactate production (F) and glucose consumption (G) of TSC-Rs following 96 hours of 10 mmol/L AG1295 (þ) or DMSO () treatment. Results are presented as a ratio normalized to DMSO-treated cells. Data in all panels are representative of 3 or more independent experiments and presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01). alters pyruvate dehydrogenase kinase 4 expression increasing pyruvate metabolism (34). Activated FGFR phosphorylates PDK1 and PKM2 enhancing glycolysis (12, 34, 40). These observations document an association between glucose metabolism and proliferation. Some have proposed that proliferation is hard-wired to changes in glycolysis to fulfill increased requirements for macromolecular synthesis (1, 2, 38), but modulation of cell-cycle progression alone is not OF10 Cancer Res; 73(6) March 15, 2013 sufficient to produce changes that resemble the Warburg effect (42–44), raising the possibility of proliferation and glycolysis being independently regulated in tumor cells. The PDGF/PDGFR pathway is known to be deregulated in multiple tumor types and it is frequently altered in glioma (13, 16, 17). We found evidence of altered glucose metabolism by PDGF and observed that glioma-derived TSCs exhibited enhanced glycolysis indistinguishable from the Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis Figure 7. AKT is a key downstream mediator of PDGF-modulated glycolysis in glioma-derived TSCs. A, immunoblot analysis of PDGFR and AKT phosphorylation in lysates of TSCs following 48 hours of LY294002 (þ) or DMSO () treatment. Lactate production (B) and glucose consumption (C) of TSCs following 96 hours of LY294002 (þ) or DMSO () treatment. D, glycolytic rate of TSCs following 48 hours of LY294002 (þ) or DMSO () treatment. E, left, immunoblot analysis of HA tag, pAKT, and phosphorylated p70-S6K in lysates of TSC-S3 overexpressing PCDNA3 and PCDNA3-HA-myrAKT1 transfectant. Right, bar graph of immunoblot band density measured in triplicate normalized to actin expression. Lactate production (F) and glucose consumption (G) of TSC-S3 transfected with PCDNA3 or PCDNA3-HA-myrAKT1 following 72 hours of doxycycline treatments. Data in all panels are representative of 3 or more independent experiments, presented as mean SEM of triplicates ( , P < 0.05; , P < 0.01; , P < 0.005). www.aacrjournals.org Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF11 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Ran et al. Warburg effect. In these TSCs, glucose uptake and lactate production are coordinately increased (Figs. 2 and 3) and were dependent on PDGF/PDGFR (Fig. 3). The PDGFinduced glucose uptake we observed was consistent with previous reports showing PDGF signaling associated with increased glucose uptake and vascular smooth muscle cell growth (27, 45). We found that elevated levels of glucose consumption and lactate production as well as an increased cellular glycolytic rate (Fig. 5) in proliferating TSCs were responsive to PDGF/PDGFR signaling. Several lines of investigation, however, indicated this regulation was independent of proliferation (Figs. 5 and 6). Seeking to identify the mediators of PDGF regulation of glioma glycolysis, we investigated 2 pathways downstream of PDGFR, PI3K/AKT and MAPK. We found that inhibition of AKT resulted in decreased glycolysis, whereas modulation of MAPK activation did not (Fig. 6). AKT, a key mediator of insulin-induced glucose uptake, coordinates changes in glucose uptake and lactate production by altering the phosphorylation state and expression of several different glycolytic enzymes (9, 26). We found that the mRNA or protein expression of these enzymes correlates with PDGFB signaling (Fig. 2G and Supplementary Fig. S2), suggesting the possibility of AKTmediating glycolysis in these cells. Overexpression of constitutively active AKT rescued decreased glycolysis associated with inhibited PDGF/PDGFR consistent with AKT acting as a key mediator of PDGF-regulated glycolysis (Fig. 7F and G). PDGFR inhibition has not yet been shown clinically to impact significantly on glioma progression as measured by tumor growth (46). We observed among the cell cultures growth resistant to doxycycline that some proliferated in the presence of AKT inhibition, suggesting that these cells acquired doxycycline resistance by activation of downstream proliferative pathways. Our finding that a PDGFR/PI3K/AKT axis regulates glioma glucose metabolism by a pathway distinct from proliferation suggests that PDGFR inhibition altering glucose metabolism might have therapeutic effects that cannot be recognized in single-agent trials monitoring tumor cell proliferation. Consistent with this idea is the finding that tumor cells may have enhanced sensitivities to apoptosis following nutriment restriction (11, 47) and the work of others to treat cancer by inhibiting glucose metabolism (2). Other evidence shows that alterations in glucose metabolism alone can influence key characteristics of malignancy (48, 49) provides a strong rationale to examine the potential of PDGF/PDGFR inhibition to contribute to tumor treatment by complementing effective, targeted growth inhibitors. Disclosure of Potential Conflicts of Interest No potential conflicts of interest were disclosed. Authors' Contributions Conception and design: C. Ran, M.A. Israel Development of methodology: C. Ran, H. Liu, Y. Hitoshi Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): C. Ran, H. Liu, Y. Hitoshi Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): C. Ran, H. Liu, M.A. Israel Writing, review, and/or revision of the manuscript: C. Ran, M.A. Israel Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): C. Ran, H. Liu Study supervision: C. Ran, M.A. Israel Acknowledgments The authors thank Z. Reitman, Drs. D. Compton, L. Witters, and H. Yan for suggestions and assistance and thank Dr. T. Tosteson for statistical consultation. Grant Support Support was generously provided by the Theodora B. Betz Foundation, Kyra and Jordon Memorial Foundation and Hitchcock Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Received June 27, 2012; revised December 20, 2012; accepted January 4, 2013; published OnlineFirst January 15, 2013. References 1. Hsu PP, Sabatini DM. Cancer cell metabolism: Warburg and beyond. Cell 2008;134:703–7. 2. Kroemer G, Pouyssegur J. Tumor cell metabolism: cancer's Achilles' heel. Cancer Cell 2008;13:472–82. 3. Jones RG, Thompson CB. Tumor suppressors and cell metabolism: a recipe for cancer growth. Genes Dev 2009;23:537–48. 4. Israel M, Schwartz L. The metabolic advantage of tumor cells. Mol Cancer 2011;10:70. 5. Warburg O. On the origin of cancer cells. Science 1956;123:309–14. 6. Hayat MA. Methods of cancer diagnosis, therapy, and prognosis. 1st ed. Union, NJ: Springer; 2010. 7. Wolf A, Agnihotri S, Guha A. Targeting metabolic remodeling in glioblastoma multiforme. Oncotarget 2011;1:552–62. 8. Thong FSL, Bilan PJ, Klip A. The Rab GTPase-activating protein AS160 integrates Akt, protein kinase C, and AMP-activated protein kinase signals regulating GLUT4 traffic. Diabetes 2007;56: 414–23. 9. Whiteman EL, Cho H, Birnbaum MJ. Role of Akt/protein kinase B in metabolism. Trends Endocrinol Metab 2002;13: 444–51. 10. Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell 2000;103:211–25. OF12 Cancer Res; 73(6) March 15, 2013 11. Weihua Z, Tsan R, Huang WC, Wu Q, Chiu CH, Fidler IJ, et al. Survival of cancer cells is maintained by EGFR independent of its kinase activity. Cancer Cell 2008;13:385–93. 12. Hitosugi T, Kang S, Vander Heiden MG, Chung TW, Elf S, Lythgoe K, et al. Tyrosine phosphorylation inhibits PKM2 to promote the Warburg effect and tumor growth. Sci signal 2009;2:ra73. 13. Dai C, Celestino JC, Okada Y, Louis DN, Fuller GN, Holland EC. PDGF autocrine stimulation dedifferentiates cultured astrocytes and induces oligodendrogliomas and oligoastrocytomas from neural progenitors and astrocytes in vivo. Genes Dev 2001;15:1913–25. 14. Hambardzumyan D, Squatrito M, Carbajal E, Holland EC. Glioma formation, cancer stem cells, and akt signaling. Stem Cell Rev 2008;4: 203–10. 15. Shih AH, Dai C, Hu X, Rosenblum MK, Koutcher JA, Holland EC. Dosedependent effects of platelet-derived growth factor-B on glial tumorigenesis. Cancer Res 2004;64:4783–9. 16. TCGA. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008;455: 1061–8. 17. Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009;462:739–44. Cancer Research Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 PDGFR-Mediated Regulation of Glioma Glycolysis 18. Hitoshi Y, Harris BT, Liu H, Popko B, Israel MA. Spinal glioma: plateletderived growth factor B-mediated oncogenesis in the spinal cord. Cancer Res 2008;68:8507–15. 19. Gottschalk S, Anderson N, Hainz C, Eckhardt SG, Serkova NJ. Imatinib (STI571)-mediated changes in glucose metabolism in human leukemia BCR-ABL-positive cells. Clin Cancer Res 2004;10:6661–8. 20. Zhou BP, Hu MC, Miller SA, Yu Z, Xia W, Lin SY, et al. HER-2/neu blocks tumor necrosis factor-induced apoptosis via the Akt/NF-kappaB pathway. J Biol Chem 2000;275:8027–31. 21. O'Neil RG, Wu L, Mullani N. Uptake of a fluorescent deoxyglucose analog (2-NBDG) in tumor cells. Mol Imaging Biol 2005;7:388–92. 22. Elstrome RL, Bauer DE, Buzzai MKR, Karnauskas R, Harris MH, Plas DR, et al. Akt Stimulates Glycolysis in Cancer Cells. Cancer Res 2004;64:3892–99. 23. Wlodkowic D, Skommer J, Darzynkiewicz Z. Flow cytometry-based apoptosis detection. Methods Mol Biol 2009;559:19–32. 24. Fishbein I, Waltenberger J, Banai S, Rabinovich L, Chorny M, Levitzki A, et al. Local delivery of platelet-derived growth factor receptorspecific tyrphostin inhibits neointimal formation in rats. Arterioscler Thromb Vasc Biol 2000;20:667–76. 25. Zamzami N, Marchetti P, Castedo M, Decaudin D, Macho A, Hirsch T, et al. Sequential reduction of mitochondrial transmembrane potential and generation of reactive oxygen species in early programmed cell death. J Exp Med 1995;182:367–77. 26. Hoehn KL, Hohnen-Behrens C, Cederberg A, Wu LE, Turner N, Yuasa T, et al. IRS1-independent defects define major nodes of insulin resistance. Cell Metab 2008;7:421–33. 27. Perez J, Hill BG, Benavides GA, Dranka BP, Darley-Usmar VM. Role of cellular bioenergetics in smooth muscle cell proliferation induced by platelet-derived growth factor. Biochem J 2010;428: 255–67. 28. Umahara M, Okada S, Yamada E, Saito T, Ohshima K, Hashimoto K, et al. Tyrosine phosphorylation of Munc18c regulates platelet-derived growth factor-stimulated glucose transporter 4 translocation in 3T3L1 adipocytes. Endocrinology 2008;149:40–9. 29. Wolf A, Agnihotri S, Micallef J, Mukherjee J, Sabha N, Cairns R, et al. Hexokinase 2 is a key mediator of aerobic glycolysis and promotes tumor growth in human glioblastoma multiforme. J Exp Med 2011;208: 313–26. 30. Guo D, Hildebrandt IJ, Prins RM, Soto H, Mazzotta MM, Dang J, et al. The AMPK agonist AICAR inhibits the growth of EGFRvIII-expressing glioblastomas by inhibiting lipogenesis. Proc Natl Acad Sci U S A 2009;106:12932–7. 31. Ko YH, Pan W, Inoue C, Pedersen PL. Signal transduction to mitochondrial ATP synthase: Evidence that PDGF-dependent phosphorylation of the delta-subunit occurs in several cell lines, involves tyrosine, and is modulated by LPA. Mitochondrion 2002; 1:339–48. 32. Kang SG, Chung H, Yoo YD, Lee JG, Choi YI, Yu YS. Mechanism of growth inhibitory effect of Mitomycin-C on cultured human retinal pigment epithelial cells: apoptosis and cell cycle arrest. Curr Eye Res 2001;22:174–81. 33. Liu K-W, Feng H, Bachoo R, Kazlauskas A, Smith EM, Symes K, et al. SHP-2/PTPN11 mediates gliomagenesis driven by PDGFRA and INK4A/ARF aberrations in mice and humans. J Clin Invest 2011;121: 905–17. www.aacrjournals.org 34. Grassian AR, Metallo CM, Coloff JL, Stephanopoulos G, Brugge JS. Erk regulation of pyruvate dehydrogenase flux through PDK4 modulates cell proliferation. Genes Dev 2011;25:1716–33. 35. Ramaswamy S, Nakamura N, Vazquez F, Batt DB, Perera S, Roberts TM, et al. Regulation of G1 progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc Natl Acad Sci U S A 1999;96:2110–5. 36. Yecies Jessica, Zhang HuiH, Menon S, Liu S, Yecies D, Lipovsky AI, et al. Akt stimulates hepatic SREBP1c and lipogenesis through parallel mTORC1-dependent and independent pathways. Cell Metabolism 2011;14:21–32. 37. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011;144:646–74. 38. Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 2009;324:1029–33. 39. Goidts V, Bageritz J, Puccio L, Nakata S, Zapatka M, Barbus S, et al. RNAi screening in glioma stem-like cells identifies PFKFB4 as a key molecule important for cancer cell survival. Oncogene 2012;31: 3235–43. 40. Hitosugi T, Fan J, Chung TW, Lythgoe K, Wang X, Xie J, et al. Tyrosine phosphorylation of mitochondrial pyruvate dehydrogenase kinase 1 is important for cancer metabolism. Mol Cell 2011;44:864–77. 41. Christofk HR, Vander Heiden MG, Harris MH, Ramanathan A, Gerszten RE, Wei R, et al. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature 2008;452:230–3. 42. Lemons JMS, Feng X-J, Bennett BD, Legesse-Miller A, Johnson EL, Raitman I, et al. Quiescent fibroblasts exhibit high metabolic activity. PLoS Biol 2010;8:e1000514. 43. Funes JM, Quintero M, Henderson S, Martinez D, Qureshi U, Westwood C, et al. Transformation of human mesenchymal stem cells increases their dependency on oxidative phosphorylation for energy production. Proc Natl Acad Sci U S A 2007; 104:6223–8. 44. Weinberg F, Hamanaka R, Wheaton WW, Weinberg S, Joseph J, Lopez M, et al. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc Natl Acad Sci U S A 2010;107: 8788–93. 45. Livnat T, Chen-Zion M, Beitner R. Platelet-derived growth factor (PDGF) rapidly stimulates binding of glycolytic enzymes to muscle cytoskeleton, prevented by calmodulin antagonists. Biochem Med Metab Biol 1994;53:28–33. 46. Razis E, Selviaridis P, Labropoulos S, Norris JL, Zhu MJ, Song DD, et al. Phase II study of neoadjuvant imatinib in glioblastoma: evaluation of clinical and molecular effects of the treatment. Clin Cancer Res 2009;15:6258–66. 47. Sengupta S, Peterson TR, Sabatini DM. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Molecular Cell 2010;40:310–22. 48. Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis?Nat Rev Cancer 2004;4:891–9. 49. Ward PS, Thompson CB. Metabolic reprogramming: a cancer hallmark even Warburg did not anticipate. Cancer Cell 2012;21: 297–308. Cancer Res; 73(6) March 15, 2013 Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research. OF13 Published OnlineFirst January 15, 2013; DOI: 10.1158/0008-5472.CAN-12-2460 Proliferation-Independent Control of Tumor Glycolysis by PDGFR-Mediated AKT Activation Cong Ran, Huan Liu, Yasuyuki Hitoshi, et al. Cancer Res Published OnlineFirst January 15, 2013. Updated version Supplementary Material E-mail alerts Reprints and Subscriptions Permissions Access the most recent version of this article at: doi:10.1158/0008-5472.CAN-12-2460 Access the most recent supplemental material at: http://cancerres.aacrjournals.org/content/suppl/2013/01/15/0008-5472.CAN-12-2460.DC1 Sign up to receive free email-alerts related to this article or journal. To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at [email protected]. To request permission to re-use all or part of this article, contact the AACR Publications Department at [email protected]. Downloaded from cancerres.aacrjournals.org on June 15, 2017. © 2013 American Association for Cancer Research.