Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Rosetta@home wikipedia , lookup
Homology modeling wikipedia , lookup
Circular dichroism wikipedia , lookup
Protein domain wikipedia , lookup
List of types of proteins wikipedia , lookup
Intrinsically disordered proteins wikipedia , lookup
Protein design wikipedia , lookup
Protein folding wikipedia , lookup
Protein structure prediction wikipedia , lookup
Bimolecular fluorescence complementation wikipedia , lookup
Protein moonlighting wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
Protein mass spectrometry wikipedia , lookup
Western blot wikipedia , lookup
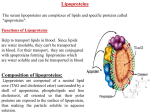
Biochemical SocietyTransactions ( 1 993) 21 Phosphorplation of apoproteins in VIADLand 1J)L by protein kinases in vitro. CLAIRE E HARRISON and MICHAEL R. MUNDAY Dept of Pharmaceutical Chemistry, School of Pharmacy, University of London, 29-39 Brunswick Square, London WClN IAX, UK. Elevated plasma levels of cholesterol-rich low density lipoprotein (LDL) are a major risk factor in the development of atherosclerosis and coronary heart disease 111. LDL is formed from intermediate density lipoprotein (IDL) derived from the peripheral catabolism of very low density lipoprotein (VLDL) originally assembled and secreted by liver 111. VLDL consists of a triacylglycerol and cholesterol-ester core which is surrounded by a monolayer of phospholipid interspersed with cholesterol. Embedded in this monolayer is apoprotein E, a 34 KDa protein whose function is to bind to LDL-receptorsand increase the plasma clearance of the IDL 121, and apoprotein BlOO a 514 KDa protein that provides a structuraJ framework for the partxle with alternating hydrophilic and hydrophobic domains that interface with the aqueous medium and the lipid core of the parr.de, respectively [3). Apo BlOO also forms the receptor recognition site in LDL particles since these have lost apoE during their formation from IDL [3]. Functional apo BlOO is essential for the hepatic assembly and secretion of VLDL which is under hormonal control, notably that of insulin and glucagon 141. There is evidence that inhibibon of VLDL production is not always matched by a decrease in apo B synthesis suggesting a possible post-translational regulation of apo B function 14). It has been reported that in newlysecreted VLDL from rat liver or hepatocytes apo B is a phosphoprotein 15, 61;that this phosphorylation is hormonally regulated by insulin 16,qand is important for apo B function [7]. We have observed that CAMP-dependent protein kinase (CAMP-PK),AMP-activated protein kinase (AMP-PK) and casein kinase 2 stoichiometrically phosphorylate apo BlOO in vitro in preparations of human LDL where it is the only apoprotein present (M R Munday, D Carling, S Takhar and C E Harrison, unpublished results). Incubation of human VLDL with [y-32P] ATP and the catalytic subunit of bovine heart CAMP-PK in virro results in the 32P-labelling of a number of protein bands separated directly by SDS PAGE (Figure la). One of these is apo BlOO in confirmation of the results obtained with LDL. The major phosphorylated band migmted with an apparent Mr of 35 KDa and minor phosphorylated bands ran at 19 and 17 KDa. Other 32P-labelledproteins are a result of phosphorylation of proteins within the protein kinase preparation parhcularly the autophosphorylated C-subunit of CAMP-PK at 40 KDa (Figure la). Figure la) Separation of VLDL apoproteins phosphorylated by CAMP-PKin vitm on 8-15% SDS PAGE. b) Cleveland V8 map on 15% SDS PAGE of apo B100 from I B L phosphorylated by CAMP-PKor AMP-PK in vitro. a) Rotein Autoradiograph blank VLDL blank VLDL apoB 514 KDa b) Autoradiograph CAMP-PK AMP-PK 1295 Cleveland mapping of these protein bands using Staphylococcus aurew V8 protease showed different patterns of 32P-labelled peptides for each suggesting that they were distinct apoproteins and not proteolytic degradation products of apoB I00 (Figure 2). The 35 KDa band is suggestive of apoE whereas the apparent 19 and 17 KDa bands fall midway between the quoted Mr for apo A and apo C classes. Preliminary experiments suggest that phosphorylations of VLDL by AMP-PK and casein kinase 2 result in 32P-labelling of apo BlOO but not the other apoproteins (data not shown). Figure 2 Cleveland V8 map of 32P labelled apoprotein bands from VLDL shown in Figure la. apo BlOO 34 KDa (apo E) 17 KDa doublet Cleveland V8 mapping of the apo BlOO band from the SDS PAGE of LDL showed a different pattern of 32P-labelled peptides for apo BlOO phosphorylated by CAMP-PKcompared with AMPPK (Figure lb). This indicates that these two kinases phosphorylate different sites. This has also been suggested by reversed phase HPLC separation of tryptic peptides from apo BlOO phosphorylated by either kinase and will be confmed when these phosphorylation sites are identified by amino acid sequencing. The physiological significance of these phosphorylations is unclear. Wr have observed that 32P-labelled apoproteins in VLDL or LDL phosphorylated by CAMP-PKare rapidly dephosphorylatedon subsequent incubation with plasma (known to be rich in phosphakx activity). Therefore, if they are to be physiologically relevant, these phosphorylationsare likely to be intracellular. Apo A, C and E are all functionally important in the circulation, but apo B is most important in intracellular VLDL assembly. Investigations of the topography of VLDL assembly poses questions as to where apo BlOO may be exposed to protein kinases 181. However, we have observed the presence of up to 0.5 mol of alkali labile phosphate per mol of apo B in freshly isolated human LDL. Phosphorylation of apo BlOO could inhibit or augment VLDL assembly and secretion. The phosphorylation of a number of proteins such as casein 191 and vitellogenin 1101 is essential for their successful secretion. Furthermore, hepatic CAMP-PK,AMP-PK and casein kinase 2 have all been observed to be under the hormonal control of glucagon andor insulin I11,12,131. Identification of apoprotein kinases, sites of phosphorylation and their presence in the lipoproteins in vivo in different physiological situations will reveal the importance of this phenomenon to VLDL assembly, secretion and metabolism. Claire Harrison is an SERC PhD student. We are grateful to the British Heart Foundation for the financial support of this project. [ I I Brown, M.S. & Goldstein J.L. (1987) Sci Am 256 ( I ) 52-60. [2] Mahley, R.W. (19 ) Science 240 622-630. 131 Knott,T.J., Pease, R.J., Powell, L.M., Wallis, S.C., Rall Jr, S.C., CAMP-PH 40 KDa 35 KDa 19 KDa 17 KDa Innerarity, T.L., Blackhart, B., Taylor, W.H., Marcell, Y.,Milne, R., Johnson, D., Fuller, M., Lusis, A.J., McCarthy, B.J., Mahley, R.W., Levy-Wilson, B. & Scott, J. (1986) Nature 323,734-738. I41 Gihhons, G.F. (1990) Biochem. J. 268 1-13. [51 Davis, R.A., Clinton, G.M., Borchardt, R.A., Malone-McNeil, M., Tan, T. & Lattier, G.R. (1984) J. B i d . Chem. 259 3383-3386. 161 Sparks, J.D., Sparks, C.E., Roncone, A.M. & Amatruda, J.M. (1988) J. Bid. Chem. 263 5001-5004. 171 Jackson, T.K., Salhanick, A.I., Elovson, J., Deichman, M.L. & Amatruda, J.M. (1990) J. Clin. Invest. 86 1746-1751. [8] Pease, RJ., Harrison, G. & Scott, J. (1991) Nature 353 448-450. [9] Bingham, E. & Farrell, H.M. (1977) J. Dairy Sci. 60 1199-1207. [lo] Wang, S. & Williams, D. (1982) J. Bid. Chem. 257 3837-3846. [ I 11 Hardie, D.G. (1989) Prog. Lipid Res. 28 117-146. 1121 Munday, M.R., Milic, M.R., Takhar, S., Holness, M J . & Sugden, M.C. (1991) Biochem. J. 280 733-737. 1131 Klarlund, J.K. & Czech, M.P. (1988) I. Biol. Chem. 263 1587215875.








![Comparison of Ligand-Binding Sites of Modeled Apo[a] Kringle](http://s1.studyres.com/store/data/022146975_1-ba8d6ebf00813a811b124aa3a02c9785-150x150.png)