Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Two-hybrid screening wikipedia , lookup
Ancestral sequence reconstruction wikipedia , lookup
Drug design wikipedia , lookup
Catalytic triad wikipedia , lookup
Point mutation wikipedia , lookup
Genetic code wikipedia , lookup
Ligand binding assay wikipedia , lookup
Metalloprotein wikipedia , lookup
Biochemistry wikipedia , lookup
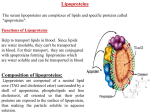
758 Comparison of Ligand-Binding Sites of Modeled Apo[a] Kringle-like Sequences in Human Lipoprotein [a] Juan Guevara Jr., Amy Y. Jan, Roger Knapp, Alexander Tulinsky, and Joel D. Morrisett Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 Human lipoprotein [a] contains at least two high-molecular-weight, disulfide-linked apolipoproteins, apo[a] and apo B-100. Apo[a] is a highly glycosylated, hydrophilic apoprotein that somewhat resembles plasminogen by containing an extended kringle domain and a carboxyl-terminal serine protease domain. The apo [a] kringle domain is composed of 11 distinct kringle types. Ten of these display high sequence homology to plasminogen kringle 4 (PGK4). The crystallographic coordinates for PGK4 were used to generate three-dimensional molecular models of the apo[a] kringle types, and the lysine-binding region of PGK4 was used to compare the different potential receptor-ligand and ligand-binding sites contained in each different PGK4-like kringle of apo [a]. A receptor-ligand site can be proposed for each kringle type. Potential serine protease cleavage sites, containing arginine-threonine and threonine-arginine, are located on the surface of the kringles. The ligand-binding site of one apo [a] kringle model is almost identical to that of PGK4 and may be a lysine-binding site of apo [a]. Four other apo [a] kringle models appear to have structurally similar lysine-binding sites, but with differences that may influence ligand-polypeptide specificity. Five apofa] kringle models have ligand-binding sites that probably do not bind lysine; one of these is the highly repeated kringle in the known apo [a] polymorph. (Arteriosclerosis and Thrombosis 1993;13:758-770) KEYWORDS • apolipoprotein[a] • plasminogen kringle 4 • ligand-binding sites E levated plasma levels of lipoprotein [a] (Lp[a]) in humans correlate with the incidence of coronary and cerebral artery disease.1"4 However, little is known about the mechanism(s) by which Lp[a] leads to atherosclerosis or whether an elevated level of plasma Lp[a] is a cause or a result of atherosclerosis. Lp[a] contains two major and distinctly different highmolecular-weight apoproteins.5-7 Within Lp[a], apoprotein[a], apo[a], a highly glycosylated, hydrophilic protein with little apparent affinity for lipid, is disulfide-linked to one molecule of apo B-100,68-13 a highly hydrophobic apolipoprotein. Apo B-100 is partly embedded in the lipid-rich, pseudomicellar Lp[a] particle. The primary structure of a single apo[a] polymorph inferred from cDNA sequencing14 suggests a polypeptide mass of about 530 kD. Apo[a] contains two distinct domains: a kringle-containing part and a serine protease domain. For the 530-kD polymorph, there are 38 repeated From the Departments of Medicine (J.G.Jr., A.Y.J., R.K., J.D.M.) and Biochemistry (J.D.M.), Baylor College of Medicine, Houston, Tex., and the Department of Chemistry (A.T.), Michigan State University, East Lansing, Mich. Supported in part by National Institutes of Health (NIH) grants HL-32971 to J.D.M., 2S07RR-05425-29 to J.G.Jr, and HL-25942 to A.T. and The W.M. Keck Center for Computational Biology. Some molecular modeling was performed using facilities of the Molecular Biology Information Resource, Department of Cell Biology, and 3DEM Resource Center (NIH grant RR-02250), Department of Biochemistry, Baylor College of Medicine. Address for correspondence: Dr. Joel D. Morrisett, The Methodist Hospital, MS A601, 6565 Fannin Street, Houston, TX 77030. Received September 14, 1992; revision accepted February 12, 1993. sequences in the kringle domain, and these occur as 11 different kringle types.15 The segments between kringles have short sequences in common with intercellular adhesion molecules16-17 and with the class of active sites of serine protease inhibitors known as serpins.1819 In apo[a], kringle types 1-10 are similar but not identical in sequence to plasminogen kringle 4 (PGK4); primary structural homologies range between 78% and 88% (Figure 1). The apo[a] kringle type 2 sequence (LPaK2) is serially repeated 28 times. A kringle is a highly conserved, triloop polypeptide structure stabilized by three disulfide bridges; its function is not catalytic but rather involves recognition and ligand binding.2021 Other proteins such as prothrombin, tissuetype plasminogen activator (t-PA), urokinase, factor XII, and hepatocyte growth factor also contain kringlelike sequences.22-23 There has been much speculation about the potential of apo[a] kringles to confer on Lp[a] the capability to compete with plasminogen for binding the lysine residues of fibrin, thereby interfering with the fibrinolytic process and leading to thrombosis and atherosclerosis.24-29 We have studied the structure of apo[a] kringle sequences to gain new insight into the possible positive physiological role(s) played by the Lp[a] particle and to help identify the mechanism(s) by which this lipoprotein may express its atherogenicity. The PGK4 and prothrombin kringle 1 structures elucidated by x-ray crystallography show conservation of the overall molecular structure,30"36 while kringle 2 of t-PA shows some small but significant differences.37-38 These observations provide a sound basis for using the three-dimensional FIGURE 2. (Left) Comparison of the amino acid residues composing the ligand-bi of PGK4 and apo[a] kringle sequences (LPaKl-LPaKlO). Conserved amino a cyan; conservative substitutions are in green; nonconservative substitutions are in Arg35, shown in purple, is a conservative change that is consistent in all PGK4-like in apo[a]. Residues 31-35 and 69 define the cationic region of the PGK4 ligand-b The residue is conserved in position 35. The anionic region in PGK4 is defined 53-57, with Asp56 as the important negatively charged side group. This feature is in LPaKlO, and a conservative substitution to glutamate maintains the chara LPaKS, LPaK6, LPaK7, and LPaK8. In PGK4, Trp60 and Trp70 form the h trough region of the ligand-binding site. LPaKZ is the highly repeated kringle s apo[a]. FIGURE 1. (Above) Comparison of apo[a] kringle-like sequences (LPaKl-LPa the 78-amino acid sequence of PGK4. This comparison is based on overal homology between the apo[a] sequences and PGK4. LPaKlO has a high degr sequence similarity (88%) and a high degree of amino acid homology at i ligand-binding site. Residues depicted in green are conservative substitutions, while t in white are nonconservative amino acid changes. Six changes are shown in pur which are conserved in all apo[a] kringle-like sequences, i.e., Vall7,Arg20, Thr21,A Thr76. Twenty-eight copies ofLPaK2 are present in the single apo[a] polymorph inf cDNA sequencing.u This apo[a] polymorph contains 37 PGK4-like repeat sequenc PGK5-like sequence. CVH6HGQSYRGTFSTTVTGRTCQSUSSMTPHRHQRTPENYPNDGLTMNVCRNPDADTGPUCFTHDPS1RUEVCNL CYHQDGRSYRGISSTTyTGRTCQSUSSHXPHUHQRTPENYPNAGLTENYCRNPDSaKOPUCVTTDPCVRUEVCNL CVHQDGQSYRG FSTTWTG RTCQSUSSMTPHUHQRTTEVYPN8GLTRNYCRNPDA EIRPUC VTHDPSVRUE YC CVgggGQSVRGTLSTT I TGRTCQSUSSMTPHUHRRIPLVYPNAGLTRNYCRNPDA EIRPUC THDPSVRWEYC CYH8DGQSVRG FSTTWTG RTCQSUSSHTPHUHQRTTEVVPN8GLTRNVCRNPDA »"ISPUC - TMDPHVRUEYC CVVHVGQSVRGTVSTTWTGRTCQAUSSMTPHQHSRTPENYPNAGLTRNYCRNPDt IR UC THDPSVRUEYCN CYM6HGQSVRGTVSTTWTGRTCQAUSSMTPHSHSRTPEVYPNAGLIMNYCRNPDI VAA VC TRDP8VRUEVCN CYH8MGQSYRGTVSTTWTGRTCQAUSSHTPHQHNRTTENYPNAGLIMNVCRNPD1 VAA VCVTRDP8VRUEVCN CYH6NGQSVRGTVSTTVTGRTCQAUSSMTPHSHSRTPEVVPNAGLIMNVCRNPDPVAA VC "TRDPSURUEVC CYH6NGQSVOGTVFITVTGRTCQAUSSMTPHSHSRTPAVYPNA6LIKNYCRNPDPVAA UCVTTDPSVRUEVCH Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 760 Arteriosclerosis and Thrombosis TABLE 1. Kringle model PGK4 LPaKl LPaK2 LPaK3 LPaK4 LPaK5 LPaK6 LPaK7 LPaK8 LPaK9 LPaKlO Vol 13, No 5 May 1993 Summary of Potential Energy Values (kcal) for Models of Apoprotein[a] Kringles Bond Angle Dihedral Improper 24 25 27 29 29 29 29 27 28 32 29 124 122 125 135 121 128 121 123 127 125 132 226 240 236 249 246 248 225 236 230 236 251 23 23 22 23 23 24 22 23 25 22 25 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 crystallographic structure of PGK4 in evaluating the putative ligand-binding sites of the apo[a] kringle-like sequences. Thus, the coordinates of PGK435 were used to generate molecular graphic models of the apo[a] PGK4-like kringles. In PGK4, the ligand-binding site, which has both dipolar and hydrophobic characteristics, appears to accommodate lysine and analogous molecules such as e-aminocaproic acid.3133'36'39"41 X-ray crystallographic data show that in PGK4, Asp54 and Asp56 form the anionic region and interact with the e-amino group of the ligand. The cationic region is composed of Lys35 and Arg69, which are located at the opposite side of the ligand-binding site. Wu et al3S have reported a crystallographic structure of PGK4 complexed with e-aminocaproic acid, which shows that in the cationic region Lys35 and Arg69 are involved in the interaction with the carboxyl moiety of e-aminocaproic acid and hence may also be important in the interaction between the kringle-binding site and the carbonyl oxygens of the polypeptide main chain of the ligand protein. The intermediate region, or hydrophobic trough of the ligand-binding site, which interacts with the methylene backbone of the lysine residue, is composed of Trp60 and Trp70. Therefore, segments of PGK4 that are important in forming the ligand-binding site that binds to lysine are located within segments 31-35, 52-57, 59-62, and 69-72 (Figure 2). Subtle amino acid substitutions in these regions of the apo[a] PGK4-like sequences suggest possible differences in the specificity of the ligand-binding sites and their adjacent microdomains. The different ligand-binding sites of modeled apo[a] kringle types 1-10 are described in this report. Molecular Modeling Systems and Methods Modeling was performed on molecular graphics workstations from Silicon Graphics Inc., using the crystallographic modeling programs CHAIN,42 Pplygen Corp.'s QUANTA 3.0, the Adopted Basis Newton-Raphson energy minimization procedure in CHARMm (version 21), and Molecular Simulations, Inc., BIOGRAF 3.0 program. There are amino acid substitutions in critical regions of Lp[a] kringles that distinguish them from PGK4 (Figure 1). The effect of these changes on the kringle three-dimensional structure was studied by using two approaches to the molecular modeling of each apo[a] kringle type. One method built each amino acid onto the appropriate a-carbon coordinates from the van der Waals Electrostatic Total -391 -3,692 -3,686 -379 -361 -367 -3,544 -3,493 -3,549 -3,513 -3,444 -366 -3,419 -3,480 -3,366 -399 -3,477 -3,447 -375 -3,611 -3,622 -3,589 -3,420 -370 -3,435 -3,770 -390 -3,501 -3,454 -377 -395 -3,590 -3,725 x-ray crystallographic structure of PGK4 by adding the amino acid side groups without positional bias. The a-carbon atoms were then held fixed as the model was subjected to energy minimization. Then all the atoms were allowed to move and the molecule was refined further. Although total energy values for these models were similar to those of PGK4, Ramachandran plots showed the dihedral angles of many nonglycine amino acids to be in energetically unfavorable regions. Therefore, these models were abandoned and were not used. The second approach was to use the complete PGK4 crystallographic structure and to make the amino acid substitutions required to convert the molecule to the appropriate apo[a] kringle type. Each model structure was refined as described above, except that amino acids such as tryptophan, tyrosine, and phenylalanine were also regularized to maintain strict aromatic ring planarity within CHAIN42 before the final energy minimization was applied. Results Amino Acid Differences Between Non-Ligand-Binding Regions ofApo[a] Kringle Sequences and PGK4 Sequence similarities between apo[a] kringles and PGK4 are shown in Figure 1. Homologies range between 78% (LPaK4) and 88% (LPaKlO). Five amino acid replacements, Vall7, Arg20, Thr21, Arg35, and Thr76, are conserved in the 10 apo[a] kringle types. Interestingly, Lys20 and Lys21 of PGK4 are replaced with arginine and threonine in all apo[a] kringles. A second analogous occurrence of this type of change is the replacement of Lys76 and Lys77 of PGK4 with threonine and arginine in LPaK4, LPaK8, and LPaKlO. (In Glu-plasminogen, Lys76-Lys77 is the cleavage site for fibrinolytic serine proteases in the plasmin activation process43-47.) In the remaining apo[a] kringles this Lys76-Lys77 sequence is replaced with threonine-glutamine. Another site where a similar change occurs is PGK4 Lys35-Thr36, which becomes Arg35-Thr36 in all apo[a] PGK4-like kringles except LPaK8. Thus, in all apo[a] PGK4-like kringles except LPaK8, there are two copies of argininethreonine. Furthermore, all apo[a] PGK4-like kringles except LPaK9 have one copy of threonine-arginine, although at different positions in the kringle. All apo[a] kringles except LPaK4 have one copy of argi- Guevara et al Modeling of Apo[a] Kringle Sequences 761 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 3. Stereoview of the ligand-binding site of modeled LPaKlO. LPaKlO is about 88% homologous with PGK4, including high sequence homology and structural fidelity with respect to the ligand-binding site ofPGK4. FIGURE 4. Stereoview of the ligand-binding site of modeled LPaK5. The molecular model of LPaKS shows a ligand-binding site that is similar to that of LPaKlO and PGK4. There are five amino acid changes that may affect the ligand and ligand-polypeptide specificity in this apoprotein[a] kringle. 762 Arteriosclerosis and Thrombosis Vol 13, No 5 May 1993 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 5. The ligand-binding sites of modeled LPaK6 and LPaK7. The ligand-binding sites of LPaK6 (panel A) and LPaK7 (panel B) are almost identical. FIGURE 6. Stereoview of the ligand-binding site of modeled LPaK8. The structural arrangement of the LPaK8 ligand-binding site suggests that it would bind lysine. Guevara et al Modeling of Apo[a] Kringle Sequences 763 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 7. Stereoview of the ligand-binding site of modeled LPaK9. Significant amino acid differences occur in the anionic region. FIGURE 8. Stereoview of the ligand-binding site of modeled LPaKA. LPaK4 has the least sequence homology with PGK4 and has several important amino acid substitutions at its putative ligand-binding site. 764 Arteriosclerosis and Thrombosis Vol 13, No 5 May 1993 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 9. Stereoviews of the ligand-binding sites of modeled LPaKl (panel A), LPaK2 (panel B), and LPaK3 (panel C). The ligand-binding sites of these apoprotein[a] kringles are very similar, quite hydrophobic, and different from that of PGK4 and the other apoproteinfa] kringle models. Guevara et al Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 9. Modeling of Apo[a] Kringle Sequences 765 Continued nine-glycine in positions 10-11. These may be potential cleavage sites for both plasmin and a-thrombin.47-49 Except for ArglO, all other arginine residues mentioned are located on the surface of the apo[a] kringle models. In LPaK8, Arg3 is part of the only "RGD" adhesion sequence present in this polymorph of apo[a]. This sequence is a /3-turn on the surface of the kringle in the LPaK8 model. Amino Acid Differences in the Ligand-Binding-Site Regions ofApofaJ Kringle Sequences General. The effects of amino acid differences in critical sequence positions of the ligand-binding sites of apo[a] kringles were evaluated by molecular modeling. The dipolar and hydrophobic ligand-binding region of PGK4 is composed of kringle segments 31-35, 54-57, 59-62, and 69-72 (Figure 2).31-33-36 Lysine and analogous molecules such as e-aminocaproic acid occupy most of the available space in the ligand-binding site of PGK4.36 This can be described as a three-sided pocket, with segments 31-35, 54-57, and 69-72 forming the sides and segment 59-62 forming the base of the pocket. The anionic region, segment 54-57, is formed by Asp54 and Asp56, which interact with the e-amino moiety of the ligand. The cationic region is composed of Lys35 and Arg69, which are located opposite to the anionic region of the ligand-binding site. The guanidino group of Arg69 and the e-amino group of Lys35 have been shown to interact with the carboxyl terminus of e-aminocaproic acid ligand.36 This suggests that either or both cationic moieties may interact with the appropriate carbonyls on a ligand polypeptide main chain. The hydrophobic region, segment 69-72, is formed by Trp70, which forms the trough, or cradle, of the ligandbinding site together with Trp60 of segment 59-62. This region interacts with the methylene backbone of the lysine residue. One residue, Trp70, is partially exposed to the aqueous solvent on the surface of the kringle. Its indole ring forms about a 90° angle with the ring of Trp60. Also integral to the binding site are Phe62 and Tyr72, which are located at opposite ends of the trough and perhaps enhance its hydrophobicity. The ligandbinding environment of PGK4 is therefore created by Lys35, Asp54, Asp56, Trp60, Arg69, Trp70, and perhaps Phe62 and Tyr72. Amino acid changes in any of these critical positions will most likely alter the binding environment as well as ligand specificity. The models of PGK4-like kringle sequences in apo[a] were examined to determine how amino acid replacements in these positions would influence the ligand-binding environment and to evaluate their lysine-binding capacities. A comparison of the total potential energies for these models and of PGK4 is given in Table 1. Ramachandran plots for each LPaK model showed that the dihedral angles of only a few nonglycine amino acids were just outside energetically favorable regions. Ligand-binding site ofLPaKlO. The ligand-binding site of LPaKlO (Figure 3) appears to retain many of the electrostatic and hydrophobic characteristics of the lysine-binding site present in PGK4. Amino acid differences occur in positions 35 and 57: Lys35 to arginine and Lys57 to threonine, respectively. The Arg35 is a mild substitution that still maintains the electropositive center of the lysine-binding site. The replacement of Lys57 with threonine could influence the positioning of Tyr72, but such an effect is not evident in the model of 766 Arteriosclerosis and Thrombosis Vol 13, No 5 May 1993 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 LPaKlO. Of all apo[a] kringle types, LPaKlO would appear to have the highest probability of binding lysine or a lysine-type ligand by a mechanism similar to the one used by PGK4. The relatively nonpolar, cradle- or trough-like environment formed by Trp60 and Trp70 is conserved between the anionic region formed by the carboxylic moieties of Asp54 and Asp56 and the cationic region formed by the guanidino groups of Arg35 and Arg69. The Asp54, Arg35, and Arg69 residues, conserved in all apo[a] kringle sequences, are prominent features of LPaKlO. The guanidino group of Arg35 forms a hydrogen bond with the carboxyl group of Asp54, and the guanidino moiety of Arg69 may be important in the interaction of the apo[a] kringle with the carbonyl moieties of ligand polypeptides. Ligand-binding site of LPaK5. Although amino acid differences between PGK4 and LPaK5 occur at positions 32, 34, 35, 56, 57, 58, and 62 (Figure 2), the electrostatic and hydrophobic characteristics of the ligand-binding site environment of PGK4 are retained in LPaK5 (Figure 4). In the anionic region, Asp56 is replaced with glutamate, but the carboxyl moiety is positioned at the top of the Trp60/Trp70 hydrophobic trough. At the base of the PGK4 trough near the cationic region formed by Arg69, Phe62 has been replaced by a tyrosine in LPaK5. The hydroxyl of Tyr62 appears to be in a polar interaction with the guanidino group of Arg69, while the Tyr62 phenolic ring interacts with the indole ring of Trp60. The cationic region is unchanged, since Lys35 of PGK4 is replaced with arginine. Lysine in position 57 of PGK4 is replaced with isoleucine in LPaK5, and its alkyl side group is oriented away from the putative binding site in the model of this kringle (Figure 4). Major residue changes are seen at positions 32 and 34, where PGK4 Arg32 and Gln34 are replaced with glutamine and serine, respectively. Since positions 32 and 34 do not appear to be part of the cationic site, this apo[a] kringle appears to retain the necessary environment for binding of lysine or lysinelike molecules. Ligand-binding sites ofLPaK6 and LPaK7. The models of LPaK6 and LPaK7 show that many of the ligandbinding site characteristics of LPaK5 are again conserved. The putative ligand-binding site of these apo[a] kringles is shown in Figure 5. The anionic and hydrophobic characteristics are retained, since the required orientations of the critical amino acid side groups are conserved in both models. The major changes occur in positions 32 and 34, where tryptophan and glutamine of LPaK6 replace glutamine and serine, respectively, of LPaK5, and Arg32 and Lys35 in PGK4 are replaced with tryptophan and arginine, respectively. These two kringles differ at positions 58 and 67, where LPaK6 has serine and asparagine, respectively, while LPaK7 has arginine and serine. These two positions are located peripheral to the putative ligand-binding site and do not appear to influence its polarity or geometry in the models. Therefore, both LPaK6 and LPaK7 should have the potential for binding lysine or lysine-like molecules. Ligand-binding site of LPaK8. The model for the ligand-binding site of LPaK8 (Figure 6) is similar to those sites described above for LPaK5, LPaK6, and LPaK7 with one notable exception. The cationic region of the LPaK8 model is composed of Arg34, Arg35, and Arg69. Significant differences between the models of LPaK8 and PGK4 occur at six positions (Arg32, Gln34, Lys35, Asp56, Lys57, and Phe62 of PGK4 are replaced with tryptophan, arginine, arginine, glutamate, isoleucine, and tyrosine, respectively). Notwithstanding these amino acid differences, the LPaK8 model also appears to have the potential for binding lysine or lysine-like ligands. Ligand-binding site of LPaK9. Unlike other apo[a] kringle types, which contain six cysteine residues, LPaK9 contains a seventh cysteine that occurs at position 67. This position is peripheral to the ligandbinding site in the LPaK9 kringle model. The kringle is a prime candidate for the site of disulfide linkage to apo B-100 because of this extra cysteine residue. Covalent and noncovalent interactions between LPaK9 and apo B-100 have been discussed elsewhere.50 In the model of LPaK9, the ligand-binding site appears to have remained intact (Figure 7) but is different from that of PGK4 and the other apo[a] kringle models described above. Major differences occur in the anionic region, where Ala55 and Asp56 of PGK4 are altered to serine and glycine, respectively, in LPaK9, which not only reduce the polarity of the anionic center of the ligand-binding site but also increase its binding volume capacity as well. The hydrophobic trough formed by Trp60 and Trp70 remains intact. Therefore, the ligand-binding site of LPaK9 appears to be less specific for lysine analogues but capable of binding larger, hydrophobic ligands, such as tyrosine, phenylalanine, and methionine. Ligand-binding site ofLPaK4. Although the hydrophobic trough formed by Trp60 and Trp70 is retained by this apo[a] kringle model, amino acid substitutions at positions 32, 34, 35, 55, 56, 57, and 62 significantly change the characteristics of the ligand-binding environment (Figure 8). In the anionic region of LPaK4, Pro55, Val56, and Ala57 replace alanine, aspartate, and lysine of PGK4, thus changing the electrostatic properties of the trough and thereby making it more hydrophobic. In the LPaK4 model there is a slight shift in the position of Trp70 and Tyr72, which also changes the orientation of the trough somewhat. Other amino acid substitutions that modify the nature of the ligand-binding site for LPaK4 occur at positions 32 and 34, which are both serine instead of the arginine and glutamine that are present in the cationic region of PGK4. Lysine in position 35 of PGK4 is altered to arginine in LPaK4, as with the other apo[a] kringles. The extensive primary and tertiary structural differences between PGK4 and LPaK4 do not favor the latter kringle for binding lysine-type ligands. However, an interaction with the ligand polypeptide main-chain carbonyl groups is suggested by the conservation of the cationic nature of the binding site with Arg69. The LPaK4 model is compatible with the binding of bulkier hydrophobic ligands. Ligand-binding sites of LPaK3, LPaK2, and LPaKl. The models of LPaKl, LPaK2, and LPaK3 suggest that alteration of Trp60 in PGK4 to Tyr60 in the apo[a] kringles alters their ligand-binding sites substantially (Figures 9A-9C). The ligand-binding site now contains three tyrosines, Tyr60, Tyr62, and Tyr72. In these kringle models, Trp70 assumes a different orientation from that in PGK4 or in the other apo[a] kringles. The hydrophobic trough is not formed, and the anionic region is likewise changed by the presence of Val56 in Guevara et al Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 FIGURE 10. The ligand-binding region of LPaKlO (yellow, cylindrical) superimposed on the analogous region of PGK4 (green). Two replacements occur at positions 35 and 57, where arginine in LPaKlO replaces lysine in PGK4 and threonine replaces lysine, respectively. An e-aminocaproate molecule (blue) is located in the ligand-binding site. place of Asp56 in PGK4. Although a ligand-binding site appears to be partially formed by residues Arg35, Asp54, Arg69, and Trp70, its size is now considerably reduced. Side-group differences at positions 32, 34, and 55 are peripheral to the ligand-binding site and do not appear to alter its geometry or polarity. Based on these models, it would appear that kringles LPaKl, LPaK2, and LPaK3 will not bind lysine or lysine-like ligands. Discussion Several assumptions were made in developing the molecular graphic models of the apo[a] kringle-like sequences that are based on the amino acid sequence of apo[a] inferred from the cDNA sequence.14 This sequence indicates that the apo[a] polymorph contains 11 different types of kringle-like repeats and that 10 of them are similar but not identical to PGK4.15 Therefore, because there exists a high degree of sequence homology among all kringles, especially between apo[a] kringle-like sequences and PGK4, we have assumed that the apo[a] sequences also exist in a kringle conformation. However, there is no direct structural evidence at present to indicate that these repeated sequences do in fact occur in the typical tri-loop kringle conformation. Sequences such as those of prothrombin kringle 1, plasminogen kringles 1 and 4, and t-PA kringle 2, with which sequence homology is about 55%, have been shown to exist in the kringle structure. To gain a clearer understanding of the apo[a] protein and how its kringle domain may influence the physiological role of the Lp[a] particle, we have developed molecular models of the apo[a] kringle-like sequences. These sequences share >78% sequence homology with PGK4. Another consideration supporting the view that these kringle structures Modeling of Apo[a] Kringle Sequences 767 may also be conserved in apo[a] is that the side-chain structures of corresponding amino acid residues appear to be the same for conserved residues in PGK1, PGK4, t-PA kringle 2, and prothrombin kringle I.35 Other important structural aspects of PGK4 that appear to be retained in the apo[a] PGK4-like sequences are the two hydrophobic cores. In PGK4, these cores are formed by Trp25, His33, Pro37, Tyr40, Leu45, Pro53, Trp60, and Phe62 and by Tyr2, Tyr9, Pro59, and Leu75. Interestingly, except for LPaKl, LPaK6, and LPaK7, apo[a] kringle models contain all residues of the hydrophobic cores of PGK4. The latter hydrophobic cluster is conserved and is evident in all apo[a] kringle models. In the model of LPaKl, substitutions in positions 37 and 60 at which threonine and tyrosine replace proline and tryptophan, respectively, the cluster is partially but not significantly modified. In LPaK6 and LPaK7, the replacement of Pro37 by threonine does not appear to alter the cluster. These observations are corroborated theoretically by the low potential energies for van der Waals forces shown in Table 1. Nevertheless, it is understood that although refinement of molecular structures by nonhydrated, static energy minimization can yield secondary and tertiary structures that are energetically favorable (Table 1), the modeled structures may be different from the true structure(s). Suffice it to add that computer molecular modeling has been shown to be a reliable method for evaluating structures that have not been elucidated by x-ray crystallographic or multidimensional nuclear magnetic resonance methods, especially when the homology is high.31 All of the apo[a] kringle models are based on coordinates of the crystallographic structure for PGK4 and are presented here as predicted structures. We then used these models to evaluate and compare their putative ligandbinding sites with that of PGK4. In human plasma, plasminogen is present at a concentration range of 1.3-2.3 /xM29 and exists in at least four molecular forms.51 Two forms are considered to be the complete molecules and have glutamate at the amino terminus of 790 amino acids in identical sequences but have different glycosylation levels. These are referred to as the Glu-I and Glu-II forms of plasminogen. The other two forms are identical but lack the first 76 residues and have lysine as the amino terminus. These are termed Lys-I and Lys-II forms of plasminogen. All forms can be readily purified from crude human plasma containing Lp[a] by affinity chromatography by a single pass through a lysine-Sepharose column in high-phosphate buffers.52-53 Plasma contains potentially 2.0-4.0 FIM plasminogen lysine-binding kringles in PGK1 and PGK4. Typically, few other proteins are retained by lysine-Sepharose, and they represent probably less than 5% of the total protein bound. Lp[a] is not one of the species that binds or competes with plasminogen in this form of affinity chromatography. Theoretically, if the PGK4-like apo[a] kringles have a binding affinity for lysine similar to that of either PGK4 or PGK1, then a small fraction of Lp[a] from crude plasma, along with plasminogen, should bind to a lysine-Sepharose affinity gel. Potential Lysine-Binding Apofa] Kringles The plasma levels of Lp[a] vary from 10 mg/L to 2,000 mg/L.29-54 Approximately 17% of these values yield an 768 TABLE 2. Arteriosclerosis and Thrombosis Comparison of Lysine-Binding Kringles, Plasminogen Kringles 1 and 5, to LPaKll 1 PGK1 LPaKll PGKS Vol 13, No 5 May 1993 30 70 20 40 50 80 10 60 CKTGDOKNYROTMSKTKNOITCQKWSSTSPHRPR-FSPATHPSEQLBENYCRNPDNDPQOPWCYITDPEKRYDYCDILBC CMFQNQKGYRaKKATTVTGTPCQEWAAQEPHRHSTPIPGTNJCWAGLBKNYCRNPDGDINGPWCYTMNPRKLFDYCDIPLC CMFGHGKGYRGiaUTTVTGTPCQDWAAQBPHRHSIFTPETNPRAGLBKNYCRNPDGDVGGPWCYTTNPRKLYDYCDVPQC Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 estimate of the concentration of apo[a] of approximately 1.7-350 mg/L. The plasma concentration of apo[a] for the 530-kD polymorph would then range between 3.0 nM and 660 nM. At the high end of these plasma levels of Lp[a], the concentration of a 280-kD apo[a] polymorph could be about 1.25 /xM. If only one of the PGK4-like apo[a] kringles binds to lysine in each apo[a] polymorph, then at 1.25 /AM apo[a], a significant fraction of the protein retained by the lysine-Sepharose affinity column would be expected to be Lpjaj. Comparison of the molecular model of LPaKlO and PGK4 with e-aminocaproic acid in the ligand pocket of the kringle reveals that this is the only apo[a] kringle type with a ligand-binding site capable of binding lysine (Figure 10). This apo[a] kringle would, therefore, be expected to bind lysine moieties in a protein such as fibrinogen or to lysine-Sepharose. Typically, plasminogen represents the bulk of crude plasma protein that binds to lysine-Sepharose under the conditions reported by others,51-53 which would suggest that LPaKlO has a different, much lower affinity than PGK4 and PGK1 for lysine-Sepharose under the conditions used for plasminogen binding. This change in specificity may be dependent on changes that occur at locations outside the ligand-binding site, perhaps at positions 35 and 68. The Lys35-to-Arg35 change seems innocuous, but the side group of Ile68 in LPaKlO may interact with amino acids that precede the lysine ligand. Alternatively, and more probable, the ligand-binding sites of apo[a] kringles may already be occupied with lysine residues of apo B-100 and would not recognize other lysine moieties.55"57 Precedence for an extraordinary attraction of the lysine-binding site for lysine residues has been set by the intermolecular interactions observed in the crystal structure of PGK43335 and the trimer formation of t-PA kringle 2, in which three intermolecular lysyl side chains bind in respective binding sites.37 Molecular models of LPaK5, LPaK6, LPaK7, and LPaK8 also show ligand-binding sites that differ slightly from those of LPaKlO and PGK4 with respect to both electrostatic and hydrophobic properties. Although the hydrophobic trough formed by Trp60 and Trp70 remains intact in these kringles, glutamate at position 56 and Tyr62 bring a negatively charged characteristic to the surface of the ligand trough. Amino acid substitutions in the cationic region, residues 31-35, may also influence the ligand-binding site formed in the apo[a] kringles, and it is possible that the residues lend specificity to each kringle type for the residues that precede lysine. These results suggest a difference in ligandaffinity constants for each kringle, perhaps as different as those reported for PGK1 and PGK4. 58 ^ Non-Lysine-Binding Apo [a] Kringles The ligand-binding sites in LPaKl, LPaK2, LPaK3, LPaK4, and LPaK9 differ significantly from the sites of apo[a] kringles with respect to their potential for bind- ing lysine. The ligand specifically bound by LPaK9 may be related to the association of this apo[a] kringle with a site on apo B-100. The Cys4,057 residue in apo[a] (Cys67 in the LPaK9 sequence) is believed to form an intermolecular disulfide bond with a cysteine of apo B-100. Molecular models of LPaK9 suggest that Cys67 is located on the surface of the kringle near the ligandbinding site. Ligands such as phenylalanine and methionine appear to fit well into the ligand-binding site of the modeled LPaK9. LPaK9 is probably an important region of apo[a] for understanding the interaction of apo[a] with apo B-100 and perhaps for identifying other regions of the kringle that are involved in the binding interaction. LPaK9 is approximately 80% homologous to PGK4 and contains a modified ligand-binding site with some nonhomologous replacements in and adjacent to the ligand-binding region. For example, Asp56, an essential amino acid for the binding of lysine to PGK4, is replaced with glycine in LPaK9. In the cationic region, Trp32 and Arg35 replace Arg32 and Lys35, respectively, in PGK4, thereby changing the surface characteristics peripheral to the ligand-binding region. These changes appear to influence the interaction of apo B-100 segments with this apo[a] kringle. In a separate study, selected segments of the low density lipoprotein apo B-100 sequence, which contain free sulfhydryl cysteines, have been subjected to energy minimization and docking with the ligand-binding site and adjacent regions of the LPaK9 model.50 In the docking experiments, apo B-100 segment 3,732-3,745 (PSCKLDFREIQIYK) displayed the best fit and the largest number of van der Waals contacts with models of LPaK9. The models showed that Phe3,738 fit and minimized well in the ligand-binding site of LPaK9 with numerous atomic contacts with Trp60 and Trp70 of the kringle. Other interactions between apo[a] and apo B-100 are likely and may include noncovalent binding through one or more of the apo[a] kringles.55-56 These interactions may involve ligands such as L-proline and L-lysine, but with lysine having a lesser role and/or affinity.57 Proline, an amino acid that is thought to be more important than lysine in the binding of apo[a] and Lp[a] to biotinylated low density lipoprotein,57 may bind to LPaKl, LPaK2, LPaK3, and/or LPaK4. The models of these apo[a] kringles demonstrate ligand-binding pockets that are significantly different from the lysinebinding site of PGK4. Proline may be a specific ligand for these apo[a] kringles. Previously, proline was modeled with LPaK9 alone and as part of an apo B-100 fragment. In the models, proline occupies only a fraction of the apparent space in the binding site of LPaK9. As part of apo B-100 fragment 1,476-1,490, proline appeared to be minimally compatible with LPaK9. Our experience with modeling the interaction of small ligands, such as single amino acids with kringles, has produced inconclusive results, suggesting that addi- Guevara et al Modeling of Apo [a] Kringle Sequences tional amino acids (e.g., those preceding and succeeding the residue of interest) most likely play a role in the overall interaction. However, it is possible that proline alone, when added in excess, can interfere with the interactions between apo[a] and biotinylated low density lipoprotein by competing with one or more proline residues on apo B-100 that bind to the apo[a] kringles, especially the highly repeated LPaK2. The critical anionic residue, Asp56 in PGK4, is replaced by valine in LPaK2, a change that influences the polarity and size of the kringle lysine-binding site. The trypsin-releasable fractions of apo B-100 from low density lipoprotein contain 78 proline residues60; 56% of these prolines are located in the ^/-terminal region. Perhaps some of these prolines might occur in exposed structural /3-turns and are sterically accessible to interact with the many repeats of LPaK2. Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 LPaKll as a Lysine-Binding Kringle Little attention has been devoted to the PGK5-like sequence contained in this apo[a] polymorph, which we have designated as LPaKll (Table 2). LPaKll comprises 80 residues and shares high homology with the non-lysinebinding kringle of plasminogen, PGK5 (about 93%), and the high-affinity, lysine-binding kringle PGK1 (about 69%). A molecular model for LPaKll was not developed, since we considered this highly speculative. However, several laboratories have shown that Lp[a] and recombinant apo[a] bind to intact fibrin as well as to digested fibrin.27'28'61 Those results suggest that LPaKlO recognizes lysines located within a polypeptide chain as well as at a carboxyl terminus, a functional feature not generally attributed to PGK4. An alternative explanation would be that LPaKll is sufficiently similar to the high-affinity, lysine-binding kringle PGK1 to also bind to lysine residues within a polypeptide chain. Other Potential Apo[a] Ligand Sites There are other sites on the apo[a] kringle models that may be ligands that interact with specific receptors. The structure of these sites was suggested by molecular modeling of the kringle sequences. For example, there is a single copy of the RGD adhesion sequence that occurs at positions 3-5 (Figure 1) in LPaK8 or positions 3,8923,894 in the complete apo[a] polymorph sequence. This motif occupies the first J3-turn on the surface of LPaK8. The sequence STTVTG at positions 14-19 in most apo[a] kringle sequences shares some homology with the receptor-ligand site of thrombospondin that is contained in the sequence SCSVTC.6263 Binding of plasminogen to specific receptors may also involve these short sequences, three copies of which are similar but not identical to this sequence and are present in plasminogen at positions 16-18, 270-272, and 476-478. Whether these sites on both apo[a] and plasminogen are receptor-binding sites remains to be shown. A third ligand type present in the apo[a] kringle sequence is the arginine-threonine and threonine-arginine sequence; it is repeated at least twice in every kringle sequence and, therefore, occurs many times throughout the entire apo[a] molecule. These sequences may be cleavage sites for other serine proteases, providing a mechanism for release of the Lp[a] particle by altering the kringle structure. Molecular modeling has shown these 769 short motifs to be located on the surface of the kringles, facilitating steric access to the motifs. Conclusions The kringle-like sequences in apo[a] readily minimize into the kringle conformation of PGK4 with very similar potential energy values and dihedral angle characteristics. The models reveal subtle amino acid differences between LPaK models and PGK4 that influence the microenvironments of the kringle structure, which could suggest differences in specificity for the ligand moiety alone and also for the ligand within a polypeptide. The model of the highly repeated apo[a] kringle type LPaK2 displays a ligandbinding site that is very different from that of PGK4 and most likely does not bind lysine, lysine analogues, or lysine adducts. Our models of apo[a] kringles and the complete primary structure of apo[a] suggest that apo[a] imparts to Lp[a] the capacity to recognize and bind a wide variety of ligand-receptor proteins. The varied ligand-binding sites contained by the apo[a] kringles as well as the adhesionlike regions of the kringle and interkringle domains strongly suggest that Lp[a] may bind a wide variety of different ligands that involve the lipoprotein in different physiological roles. References 1. Dahlen GH, Guyton JR, Attar M, Farmer JA, Kautz JA, Gotto AM Jr: Association of levels of lipoprotein Lp[a], plasma lipids, and other lipoproteins with coronary artery disease documented by angiography. Circulation 1986;74:758-765 2. Hoefler G, Harnoncourt F, Paschke E, Mirtl W, Pfeiffer KH, Kostner GM: Lipoprotein Lp(a): A risk factor for myocardial infarction. Arteriosclerosis 1988;8:398-401 3. Zenker G, Koltringer P, Bone G, Niederkorn K, Pfeiffer K, Jurgens G: Lipoprotein(a) as a strong indicator for cerebrovascular disease. Stroke 1986;17:942-945 4. Murai A, Miyahara T, Fujimoto N, Matsuda M, Kameyama M: Lp(a) lipoprotein as a risk factor for coronary heart disease and cerebral infarction. Atherosclerosis 1986;59:199-204 5. Ehnholm C, Garoff H, Renkonen O, Simons K: Protein and carbohydrate composition of lp(a) lipoprotein from human plasma. Biochemistry 1972;ll:3229-3232 6. Gaubatz JW, Heideman C, Gotto AM Jr, Morrisett JD, Dahlen GH: Human plasma lipoprotein [a] structural properties. J Biol Chem 1983;258:4582-4589 7. Utermann G, Weber W: Protein composition of Lp[a] lipoprotein from human plasma. FEBS Lett 1983;154:357-361 8. Gaubatz JW, Chari MV, Nava ML, Guyton JR, Morrisett JD: Isolation and characterization of the two major apoproteins in human lipoprotein[a]. J Lipid Res 1987;28:69-79 9. Armstrong VW, Walli AK, Seidel D: Isolation, characterization, and uptake in human fibroblasts of an apo(a)-free lipoprotein obtained on reduction of lipoprotein(a). J Lipid Res 1985 ;26: 1314-1323 10. Fless GM, ZumMallen ME, Scanu AM: Physico-chemical properties of apolipoprotein(a) and lipoprotein(a-) derived from the dissociation of human plasma lipoprotein(a). J Biol Chem 1986; 261:8712-8718 11. Seman LJ, Breckenridge WC: Isolation and partial characterization of apolipoprotein(a) from human lipoprotein (a). Biochem Cell Biol 1986;64:999-1009 12. Eaton DL, Fless GM, Kohr WJ, McLean JW, Xu Q-T, Miller CG, Lawn RM, Scanu AM: Partial amino acid sequence of apolipoprotein(a) shows that it is homologous to plasminogen. Proc NatlAcad SciUSA 1987;84:3224-3228 13. Kratzin H, Armstrong VW, Niehaus M, Hilschmann N, Seidel D: Structural relationship of an apolipoprotein (a) phenotype (570 kDa) to plasminogen: Homologous kringle domains are linked by carbohydrate-rich regions. Biol Chem Hoppe-Seyler 1987;368: 1533-1544 14. McLean JW, Tomlinson JE, Kuang W-J, Eaton DL, Chen EY, Fless GM, Scanu AM, Lawn RM: cDNA sequence of human apolipoprotein(a) is homologous to plasminogen. Nature 1987;330: 132-137 770 Arteriosclerosis and Thrombosis Vol 13, No 5 May 1993 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 15. Morrisett JD, Gaubatz JW, Knapp RD, Guevara JG Jr: Structural properties of apo(a): A major apoprotein of human lipoprotein(a), chapter 4, in Scanu A (ed): Lipoprotein(a). San Diego, Calif, Academic Press Inc, 1990, pp 53-74 16. Staunton DE, Marlin SD, Stratowa C, Dustin ML, Springer TA: Primary structure of ICAM-1 demonstrates interaction between members of the immunoglobulin and integrin supergene families. Cell 1988;52:925-933 17. Staunton DE, Dustin ML, Springer TA: Functional cloning of ICAM-2, a cell adhesion ligand for LFA-1 homologous to ICAM-1. Nature 1989;339:61-64 18. Huber R, Carrell RW: Implications of the three-dimensional structure of a,-antitrypsin for structure and function of serpins. Biochemistry 1989;28:8951-8966 19. Potempa J, Shieh B-H, Travis J: Alpha-2-antiplasmin: A serpin with two separate but overlapping reactive sites. Science 1988;241: 699-700 20. Magnusson S, Sottrup-Jensen L, Petersen TE, Claeys H: The primary structure of prothrombin, the role of vitamin K in blood coagulation, and a thrombin-catah/zed "negative-feedback" control mechanism for limiting activation of prothrombin, in Hemker HC, Veltkamp J (eds): Prothrombin and Related Coagulation Factors: Boerhaave Symposium, May 1974. Leiden, University Press, 1975, p 25 21. Magnusson S, Sottrup-Jensen L, Petersen TE, Dudek-Wojciechowska G, Claeys H: Homologous "kringle" structures common to plasminogen and prothrombin: Substrate specificity of enzymes activating prothrombin and plasminogen, in Ribbons DW, Brew K (eds): Proteofysis and Physiological Regulation. New York, Academic Press Inc, 1976, pp 203-238 22. Castellino FJ, Beals JM: The genetic relationships between the kringle domains of human plasminogen, prothrombin, tissue plasminogen activator, urokinase, and coagulation factor XII. J Mol Evol 1976;26:358-369 23. Ikeo K, Takahashi K, Gojobori T: Evolutionary origin of numerous kringles in human and simian apolipoproteins(a). FEBS Lett 1991;287:146-148 24. Gonzalez-Gronow M, Edelberg JM, Pizzo SV: Further characterization of the cellular plasminogen binding site: Evidence that plasminogen 2 and lipoprotein a compete for the same site. Biochemistry 1989;28:2374-2377 25. Hajjar KA, Gavish D, Breslow JL, Nachman RL: Lipoprotein(a) modulation of endothelial cell surface fibrinolysis and its potential role in atherosclerosis. Nature 1989;339:303-305 26. Miles LA, Fless GM, Levin EG, Scanu AM, Plow EF: A potential basis for the thrombotic risks associated with lipoprotein(a). Nature 1989;339:301-303 27. Harpel PC, Gordon BR, Parker TS: Plasmin catalyzes binding of lipoprotein(a) to immobilized fibrinogen and fibrin. Proc NatlAcad SciUSA 1989;86:3847-3851 28. Loscalzo J, Weinfeld M, Fless GM, Scanu AM: Lipoprotein(a), fibrin binding, and plasminogen activation. Arteriosclerosis 1990;10: 240-245 29. Rouy D, Grailhe P, Nigon F, Chapman J, Angles-Cano E: Lipoprotein(a) impairs generation of plasmin by fibrin-bound tissue-type plasminogen activator. Arterioscler Thromb 1991;ll:629-638 30. Park CH, Tulinsky A: Three-dimensional structure of the kringle sequence: Structure of prothrombin fragment 1. Biochemistry 1986; 25:3977-3982 31. Tulinsky A, Park CH, Mao B, Llinas M: Lysine/fibrin binding sites of kringles modeled after the structure of kringle 1 of prothrombin. Proteins Struct Fund Genet 1988;3:85-96 32. Tulinsky A, Park CH, Skrzypczak-Jankun E: Structure of prothrombin fragment 1 refined at 2.8 A resolution. J Mol Biol 1988; 202:885-901 33. Mulichak AM, Tulinsky A: Structure of the lysine-fibrin binding site of human plasminogen kringle 4. Blood Coagul Fibrinolysis 1990;l:673-679 34. Seshadri TP, Tulinsky A, Skrzypczak-Jankun E, Park CH: Structure of bovine prothrombin fragment 1 refined at 2.25 A resolution. J Mol Biol 1991;220:481-494 35. Mulichak AM, Tulinsky A, Ravichandran KG: Crystal and molecular structure of human plasminogen kringle 4 refined at 1.9 A resolution. Biochemistry 1991;30:10576-10588 36. Wu T-P, Padmanabhan K, Tulinsky A, Mulichak AM: The refined structure of the e-aminocaproic acid complex of human plasminogen kringle 4. Biochemistry 1991;30:10589-10594 37. de Vos AM, Ultsch MH, Kelley RF, Padmanabhan K, Tulinsky A, Westbrook ML, Kossiakoff AA: Crystal structure of kringle 2 of tissue plasminogen activator at 2.4 A resolution. Biochemistry 1992; 31:270-279 38. Byeon I-JL, LlinSs M: Solution structure of the tissue-type plasminogen activator kringle 2 domain complexed to 6-aminohexanoic acid an antifibrinolytic drug. J Mol Biol 1991;222:1035-1051 39. Vali Z, Patthy L: Location of the intermediate and high affinity omega-aminocarboxylic acid-binding sites in human plasminogen. JBiolChem 1982;257:2104-2110 40. Trexler M, Vali Z, Patthy L: Structure of the omega-aminocarboxylic acid-binding sites of human plasminogen. / Biol Chem 1982; 257:7401-7406 41. Petros AM, Ramesh V, Llinas M: 'H NMR studies of aliphatic ligand binding to human plasminogen kringle 4. Biochemistry 1989; 28:1368-1376 42. Sack JS: CHAIN: A crystallographic modeling program. J Mol Graph 1988;6:224-225 43. Allen RA, Pepper DS: Isolation and properties of human vascular plasminogen activator. Thromb Haemost 1981;45:43-50 44. Robbins KC, Summaria L, Wohl RC: Human plasmin. Methods Enzymol 1981;80:379-387 45. Holmes WE, Pennica D, Blaber M, Rey MW, Guenzler WA, Steffens GJ, Heyneker HL: Cloning and expression of the gene for pro-urokinase in Escherichia coli. Biotechniques 1985;3:923-929 46. Ichinose A, Takio K, Fujikawa K: Localization of the binding site of tissue-type plasminogen activator to fibrin. J Clin Invest 1986; 78:163-169 47. Furlan M: Structure of fibrinogen and fibrin, in Francis JL (ed): Fibrinogen, Fibrin Stabilization, and Fibrinolysis: Clinical, Biochemical and Laboratory Aspects. New York, VCH Publishers, 1988, chap 1, pp 17-64 48. Hedner V, Davie EW: Introduction to hemostasis and the vitamin K-dependent coagulation factors: Part 14. Blood and bloodforming tissue, in Scriver CR, Beaudet AL, Sly WS, Valle D (eds): The Metabolic Basis of Inherited Disease. New York, McGraw-Hill Info Serv Co, 1989, pp 2107-2134 49. Machovich R (ed): The Thrombins, Volume I. Boca Raton, Fla, CRC Press, 1984, pp 1-166 50. Guevara J Jr, Spurlino JC, Jan AY, Yang C-Y, Tulinsky A, Prasad BVV, Gaubatz JW, Morrisett JD: Proposed mechanisms for binding of apo[a] kringle type 9 to apo B-100 in human lipoprotein[a]. Biophys J 1993;64:686-700 51. Nieuwenhuizen W, Traas DW: A rapid and simple method for the separation of four molecular forms of human plasminogen. Thromb Haemost 1989;61:208-210 52. Deutsch DG, Mertz ET: Plasminogen: Purification from human plasma by affinity chromatography. Science 1970;170:1095-1096 53. Summaria L, Spitz F, Arzadon L, Boreisha IG, Robbins KC: Isolation and characterization of the affinity chromatography forms of human glu- and lys-plasminogens and plasmins. J Biol Chem 1976; 251:3693-3699 54. Kamboh ML, Ferrell RE, Kottke BA: Expressed hypervariable polymorphism of apolipoprotein (a). Am J Hum Genet 1991;49: 1063-1074 55. Ye SQ, Trieu VN, Stiers DL, McConathy WJ: Interactions of low density lipoprotein2 and other apolipoprotein B-containing lipoproteins with lipoprotein(a). J Biol Chem 1988;263:6337-6343 56. Trieu VN, McConathy WJ: Lipoprotein(a) binding to other apolipoprotein B containing lipoproteins. Biochemistry 1990;29:5919-5924 57. Trieu VN, Zioncheck TF, Lawn RM, McConathy WJ: Interaction of apolipoprotein(a) with apolipoprotein B-containing lipoproteins. J Biol Chem 1991;266:5480-5485 58. Lerch PG, Rickli EE, Lergier W, Gillessen D: Localization of individual lysine binding regions in human plasminogen and investigations on their complex forming properties. Eur J Biochem 1980;107:7-13 59. Cole KR, Castellino FJ: The binding of antifibrinolytic amino acids to kringle 4-containing fragments of plasminogen. Arch Biochem Biophys 1984;229:568-575 60. Yang C-Y, Gu Z-W, Weng S-a, Kim TW, Chen S-H, Pownall HJ, Sharp PM, Liu S-W, Li W-H, Gotto AM Jr, Chan L: Structure of apolipoprotein B-100 of human low density lipoproteins. Arteriosclerosis 1989;9:96-108 61. Rouy D, Koschinsky ML, Fleury V, Chapman J, Angl6s-Cano E: Apolipoprotein(a) and plasminogen interaction with fibrin: A study with recombinant apolipoprotein(a) and isolated plasminogen fragments. Biochemistry 1992;31:6333-6339 62. Tuszynski GP, Rothman VL, Deutch AH, Hamilton BK, Eyal J: Biological activities of peptides and peptide analogues derived from common sequences present in thrombospondin, properdin, and malarial proteins. J Cell Biol 1992; 116:209-217 63. Asch AS, Silbiger S, Heimer E, Nachman RL: Thrombospondin sequence motif (CSVTCG) is responsible for CD36 binding. Biochem Biophys Res Commun 1992;182:1208-1217 Downloaded from http://atvb.ahajournals.org/ by guest on August 3, 2017 Comparison of ligand-binding sites of modeled apo[a] kringle-like sequences in human lipoprotein[a]. J Guevara, Jr, A Y Jan, R Knapp, A Tulinsky and J D Morrisett Arterioscler Thromb Vasc Biol. 1993;13:758-770 doi: 10.1161/01.ATV.13.5.758 Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1993 American Heart Association, Inc. All rights reserved. Print ISSN: 1079-5642. Online ISSN: 1524-4636 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://atvb.ahajournals.org/content/13/5/758 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Arteriosclerosis, Thrombosis, and Vascular Biology can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answerdocument. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular Biology is online at: http://atvb.ahajournals.org//subscriptions/