Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Cell growth wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell culture wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cellular differentiation wikipedia , lookup
List of types of proteins wikipedia , lookup
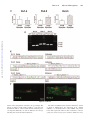
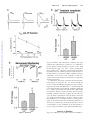
The Young Mouse Heart Is Composed of Myocytes Heterogeneous in Age and Function Marcello Rota, Toru Hosoda, Antonella De Angelis, Michael L. Arcarese, Grazia Esposito, Roberto Rizzi, Jochen Tillmanns, Derin Tugal, Ezio Musso, Ornella Rimoldi, Claudia Bearzi, Konrad Urbanek, Piero Anversa, Annarosa Leri, Jan Kajstura Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Abstract—The recognition that the adult heart continuously renews its myocyte compartment raises the possibility that the age and lifespan of myocytes does not coincide with the age and lifespan of the organ and organism. If this were the case, myocyte turnover would result at any age in a myocardium composed by a heterogeneous population of parenchymal cells which are structurally integrated but may contribute differently to myocardial performance. To test this hypothesis, left ventricular myocytes were isolated from mice at 3 months of age and the contractile, electrical, and calcium cycling characteristics of these cells were determined together with the expression of the senescence-associated protein p16INK4a and telomere length. The heart was characterized by the coexistence of young, aged, and senescent myocytes. Old nonreplicating, p16INK4a-positive, hypertrophied myocytes with severe telomeric shortening were present together with young, dividing, p16INK4a-negative, small myocytes with long telomeres. A class of myocytes with intermediate properties was also found. Physiologically, evidence was obtained in favor of the critical role that action potential (AP) duration and ICaL play in potentiating Ca2⫹ cycling and the mechanical behavior of young myocytes or in decreasing Ca2⫹ transients and the performance of senescent hypertrophied cells. The characteristics of the AP appeared to be modulated by the transient outward K⫹ current Ito which was influenced by the different expression of the K⫹ channels subunits. Collectively, these observations at the physiological and structural cellular level document that by necessity the heart has to constantly repopulate its myocyte compartment to replace senescent poorly contracting myocytes with younger more efficient cells. Thus, cardiac homeostasis and myocyte turnover regulate cardiac function. (Circ Res. 2007;101:387-399.) Key Words: action potential profile 䡲 excitation-contraction coupling 䡲 myocyte volume 䡲 telomere length 䡲 senescence-associated proteins T turnover of myocytes results in a heterogeneous population of parenchymal cells which are structurally integrated but may contribute differently to myocardial performance. If this were the case, senescent cells with alterations in the mechanical, electrical, and calcium cycling properties4 may coexist with newly formed young myocytes with enhanced contractile behavior.5 Data in humans and animals suggest that myocyte maturation and aging6 – 8 are characterized by loss of replicative potential, telomeric shortening, and the expression of the age-associated protein p16INK4a. The acquisition of the senescent phenotype involves a progressive increase in the size of the cell that reaches a critical volume beyond which cellular hypertrophy is no longer possible.6 In this study we have addressed the question whether myocyte aging is only partly related to the age of the organ and organism by testing whether young and old cells are present in the mouse heart at he recent identification of a population of resident progenitor cells that controls cardiomyogenesis imposes a reinterpretation of the fundamental mechanisms of growth and senescence of the heart.1 Traditionally, the heart was viewed as an organ characterized by a predetermined number of myocytes which is defined shortly after birth and is preserved throughout life. However, BrdU administration has documented that myocyte renewal occurs in the adult heart and that the rate of cell regeneration is slow, heterogeneous, and involves only a small percentage of cells at any given time.2,3 In mammals, ventricular myocytes are replaced several times throughout life.2,3 These findings were obtained by pulse-chase BrdU labeling assay2 and by the rate of growth and commitment of cardiac progenitor cells.3 Collectively, they challenge the notion that the age and lifespan of myocytes coincide with the age and lifespan of the organ and organism. According to the new paradigm, the continuous Original received February 28, 2007; revision received June 13, 2007; accepted June 20, 2007. From the Cardiovascular Research Institute (M.R., T.H., A.D.A., M.L.A., G.E., R.R., J.T., D.T., E.M., C.B., K.U., P.A., A.L., J.K.), Department of Medicine, New York Medical College, Valhalla; and the MRC Clinical Sciences Centre (O.R.), Faculty of Medicine, Imperial College School of Medicine, London, UK. Correspondence to Marcello Rota, PhD, Cardiovascular Research Institute, Vosburgh Pavilion, Room 302, New York Medical College, Valhalla, New York 10595. E-mail [email protected] © 2007 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.107.151449 387 388 Circulation Research August 17, 2007 3 months of age and whether this nonuniform cell population is functionally heterogeneous. This possibility would strengthen the notion of the dynamism of the heart that is regulated by a pool of progenitor cells which dictate cell turnover, organ homeostasis, and aging. Materials and Methods Myocytes were enzymatically dissociated from the heart of C57Bl6 mice. The expression of the senescence-associated protein p16INK4a and telomere length were determined together with the electrical and mechanical properties of the isolated myocytes. Additionally, the expression of Kv1.2, Kv4.2 and Kv4.3 in myocytes of different size was determined by real-time RT-PCR and immunocytochemistry. An expanded Materials and Methods section can be found in the online data supplement available at http://circres.ahajournals.org. Results INK4a Myocyte Aging: p16 Cell Volume , Telomere Length, and Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Left ventricular (LV) myocytes were enzymatically dissociated and the expression of p16INK4a was determined by immunocytochemistry (Figure 1A), the length of telomeres was evaluated by Q-FISH (Figure 1B), and myocyte volume was assessed by confocal microscopy (Figure 1C). In mice at 3 months of age, mononucleated, binucleated and multinucleated myocytes represented 6.0⫾0.74%, 91⫾0.62%, 3.0⫾0.71%, respectively. The increase in number of nuclei was paralleled by an increase in average cell volume (supplemental Figure IA) and a shift to the right in the myocyte volume distribution (supplemental Figure IB). Myocytes of different size coexisted in distinct anatomical regions (supplemental Figure IC). These data are consistent with previous results supporting the notion that under physiological conditions local stresses do not affect significantly myocyte volume across the LV wall.9 The analysis of myocytes was restricted to the LV base and midregion; the apex was excluded. Mononucleated myocytes included a subset of replicating Ki67- and BrdU-positive cells together with dividing myocytes (Figure 2A and 2B). The recognition that small, mononucleated myocytes proliferate and differentiate until the acquisition of the terminally differentiated phenotype3 is in agreement with recent results in the postnatal and adult overloaded heart.6,10 The cell cycle inhibitor p16INK4a is a reliable marker of irreversible growth arrest and cellular senescence.8 Apoptosis of myocytes and cardiac progenitor cells (CPCs) in animals and humans is restricted to p16INK4a-positive cells which do not express telomerase and typically show severe telomere shortening.6,7,11 The percentage of senescent myocytes increased dramatically in the larger binucleated and multinucleated cells (Figure 2C). Lineage commitment of CPCs to cardiomyocytes involves the generation of progenitors, precursors, and amplifying cells6 which divide until the adult phenotype is reached. Telomere shortening occurs in developing and mature myocytes; it reflects the replicative history of the cell together with a time-dependent accumulation of oxidative DNA damage.12,13 Telomere attrition promotes the expression of p16INK4a providing a mechanistic link between these 2 indices of cellular senescence which are both related to myocyte volume.6,8 A distribution plot showed an inverse correlation between telomere length and cell volume (Figure 2D). Telomere length varied from 14 to 58 kbp and large old myocytes possessed very short telomeres consistent with their senescent phenotype. When cells were separated in 3 classes, ⬍20 000, 20 000 to 40 000, and ⬎40 000 m3, telomere length was 40⫾0.73, 33⫾0.82, and 25⫾0.36 kbp, respectively (Figure 2E). In comparison with myocytes ⬍20 000 m3, the fraction of apoptotic myocytes increased ⬇5-fold and ⬇15-fold in cells 20 to 40 000 and ⬎40 000 m3, respectively (Figure 2F). Thus, the young adult mouse heart contains a heterogeneous population of myocytes of different age and cellular aging involves an increase in cell volume, telomeric shortening, p16INK4a expression, and apoptosis. Myocyte Aging: Cell Mechanics and Ca2ⴙ Handling The documentation that senescent and nonsenescent myocytes are present in the young adult mouse heart raised the question whether structural heterogeneity is paralleled by a similar variability in mechanical performance. Comparable results at the physiological level would support the notion that the heart is constantly repopulating its myocyte compartment to preserve cardiac function. Therefore, myocytes were field-stimulated at 1 Hz and cell and sarcomere shortening were measured. Increases in myocyte volume were coupled with decline in contractile behavior. Bivariate distribution of parameters of cell contractility and volume were fitted with straight lines. Cell shortening, maximal rate of contraction (dL/dt), and relaxation (⫹dL/dt) decreased, respectively, 0.32⫾0.058 m, 5.91⫾1.20 m/sec and 5.43⫾0.92 m/sec per 10 000 m3 increase in volume (Figure 3A and 3B). Relaxation was also prolonged in large myocytes but timeto-peak-shortening was similar in all cells. The question was whether myocyte shortening, p16INK4a expression, and telomere length could be concurrently evaluated in the same cells to provide a cause-and-effect relationship among these variables. To facilitate our task, mononucleated, binucleated, and multinucleated myocytes were studied. Large multinucleated myocytes were p16INK4apositive, had short telomeres, and showed a severe depression in myocyte shortening and relengthening (Figure 3C). This complex protocol requires measurements in living cells and subsequently in fixed preparations; 16 myocytes were evaluated in this manner. These results support the notion that cellular aging dictated by p16INK4a and shortened telomeres is characterized by attenuation in myocyte mechanical behavior. To define the mechanisms of depressed myocyte performance with cellular hypertrophy-aging, intracellular Ca2⫹ and sarcomere shortening were analyzed. Both parameters decreased with increases in myocyte volume (Figure 3D and 3E). Diastolic sarcomeric length was not affected by cell size and averaged 1.720⫾0.003 m (Figure 3E). The relative increase of Ca2⫹ amplitude in small-young myocytes was associated with a prolongation of time to 50% Ca2⫹ transients. However, the timing of Ca2⫹ decay was comparable in all cells (not shown). The Ca2⫹ present in the SR was then measured by the rapid addition of caffeine. The amount of Ca2⫹ in the SR was similar in cells of different size (Figure Rota et al Myocyte Heterogeneity 389 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 1. Myocyte characteristics. A, p16INK4a (yellow, arrows) in a binucleated and tetranucleated myocyte (␣ -sarcomeric-actin, red). Nuclei (PI, blue). B, Telomeres (white) in a mononucleated, binucleated, and trinucleatedmyocyte. C, Three-dimensional optical section reconstruction by confocal-microscopy of mononucleated, binucleated, trinucleated, and tetranucleated myocytes. 390 Circulation Research August 17, 2007 Myocyte Aging: ICaL and Excitation-Contraction (EC) Coupling Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 The recognition that increases in myocyte volume were associated with a decrease in Ca2⫹ transients in the absence of changes in Ca2⫹ load of the SR raised the possibility that Ca2⫹ current (ICaL) or its efficacy to activate the ryanodine receptors (RyRs) was impaired in senescent myocytes.14 The release of Ca2⫹ from the SR is triggered by the influx of Ca2⫹ from the extracellular space through L-type channels. However, the ability of ICaL to promote Ca2⫹ release from the SR, ie, excitation-contraction coupling gain (ECC gain), may be altered by spatial remodeling of ECC elements and/or open probability of the RyRs. Patch-clamp in voltage-clamp mode was used to measure Ca2⫹ current in myocytes5,7,15 ⬍20 000, 20 000 to 40 000, and ⬎40 000 m3. These myocytes had membrane capacitance (Cm) ⬍98, 98 to 164, and ⬎164 pF, respectively (supplemental Figure II). Depolarizing steps showed that ICaL density and current-voltage relations did not differ in small-young and hypertrophied-old myocytes (Figure 4A and 4B). ICaL activation-curve and half-maximal-activation-potential were comparable in all myocytes. However, in small-young myocytes, ICaL inactivation curve was shifted to the right (Figure 4B). Fast and slow time constants of ICaL inactivation time course were similar in the myocyte categories. Subsequently, ICaL and Ca2⫹ transients were recorded simultaneously in voltage-clamped cells. Depolarizing steps activated similar Ca2⫹ currents and led to comparable Ca2⫹ transients in all myocytes (Figure 4C and 4D). The ratio of Ca2⫹ transient and ICaL amplitude, ie, ECC gain, was similar in the 3 myocyte classes (Figure 4E). Thus, ICaL and ECC gain are not affected by myocyte volume or aging and thereby do not account for the decrease in SR Ca2⫹ release seen in enlarged-old myocytes. Myocyte Aging: Action Potential Figure 2. Myocyte formation and senescence. A, Ki67 (green) and BrdU (white) in mononucleated myocytes. B, Metaphase chromosomes (upper-panels, DAPI, blue) in mononucleated myocytes (lower-panels; phospho-H3⫽green). C, p16INK4apositive myocytes (MONO, n⫽115; BI, n⫽1501; MULTI, n⫽46). D, Distribution of telomere length vs myocyte volume (n⫽512) was fitted with a straight line (r2⫽0.387; P⬍0.0001). E, Telomere length in myocytes ⬍20 000 (n⫽182), 20 to 40 000 (n⫽109), and ⬎40 000 m3 (n⫽221). F, Apoptosis in myocytes ⬍20 000 (n⫽297⫻103), 20 to 40 000 (n⫽259⫻103), and ⬎40 000 m3 (n⫽242⫻103). *P⬍0.05 vs ⬍20 000; **P⬍0.05 vs 20 to 40 000. 3F) suggesting that defects in the fractional release of Ca2⫹ from the SR developed in enlarged-old myocytes. Thus, the young adult mouse heart contains a population of hypertrophied-senescent myocytes with depressed contractile behavior possibly mediated by alterations in Ca2⫹ handling. Lack of alterations in ICaL and ECC gain pointed to changes in action potential (AP) duration as a mechanism for the decrease in Ca 2 ⫹ transients and shortening in hypertrophied-old myocytes.16,17 The inotropic effect induced by AP prolongation is mediated by changes in transmembrane Ca2⫹ influx enhancing Ca2⫹-induced Ca2⫹ release from the SR.14,16 At 1 Hz, an inverse relationship was found between AP duration and myocyte volume (Figure 5A). In comparison with AP in myocytes ⬍20 000 m3, 90% repolarization time was 30% and 55% shorter in myocytes 20 000 to 40 000 and ⬎40 000 m3, respectively (Figure 5B). Similar results were obtained at 10%, 30%, 50%, and 70% repolarization. The electromechanical properties of endocardial and epicardial myocytes were evaluated in a cell subset to determine whether the distinct anatomical localization of these cells was partly responsible for the results discussed above. Shorter APs and declined contractility were detected in myocytes of increasing volume independently from their regional distribution (supplemental Figure III). Thus, the heterogeneity in myocyte performance and electrophysiology appears to affect all areas of the young-adult heart. Rota et al Myocyte Heterogeneity 391 To confirm that decreases in AP duration of large myocytes were a consequence of cellular aging, the electrical properties of 1-day-old neonatal mouse myocytes were determined (Figure 5C). The AP was significantly prolonged in these myocytes supporting the notion that cellular aging is coupled with faster repolarization. Therefore, changes in the profile of the AP may promote alterations in Ca2⫹ handling and contractile performance of hypertrophied-senescent myocytes. Myocyte Aging: Transient Outward Kⴙ Current Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 The documentation that shorter APs are present in old myocytes suggested that changes in transmembrane ionic currents may occur with cellular aging. The transient outward K⫹ current Ito is responsible for the early repolarization phase of the AP.18 Ito expression varies with ventricular loading and hypertrophy,17,19 raising the possibility that changes in Ito alter the electrical properties of enlarged-senescent myocytes. Ito density was positively correlated with myocyte volume (Figure 6A and 6B). To confirm that the expression of K⫹ channel mRNAs changed as a function of cell volume, the K⫹ channel subunits Kv1.4, Kv4.2, and Kv4.3 were measured by real-time RTPCR in small and large myocytes.10 The expression of Kv1.4 was lower in large than in small cells while Kv4.2 and Kv4.3 were more abundant in hypertrophied myocytes (Figure 6C through 6E). These results at the mRNA level were accompanied by a similar difference in K⫹ channel protein for Kv4.3 detected by immunocytochemistry (Figure 6F). Thus, the expression of K⫹ channel subunits in combination with Ito current density conditions AP duration in small-young and hypertrophied-old myocytes. Myocyte Aging: Action Potential and Developed Ca2ⴙ To establish whether the AP profile was the major determinant of depressed developed-Ca2⫹ in hypertrophied-senescent cells, AP-clamp studies were performed together with measurements of Ca2⫹ transients. Action potentials with slow (APslow), intermediate (APintermediate), and fast (APfast) repolarizing phase were recorded from myocytes with Cm 25, 112, and 207 pF, which corresponded to cells of ⬍20 000, 20 000 to 40 000, and ⬎40 000 m3, respectively (Figure 7A). Subsequently, myocytes from adult and neonatal hearts were loaded with Fluo-3 and APslow, APintermediate, and APfast waveforms were imposed as voltage-clamp command (AP-clamp mode) to evaluate their effects on Ca2⫹ transients. Ca2⫹ amplitude varied as a function of the waveforms and these changes were Figure 3. Myocyte mechanics and Ca2⫹ cycling. A, Shorteningtracings. B-F, Bivariate distribution of physiological-parameters versus myocyte-volume (B, E, F). B: cell-shortening (r2⫽0.0973; P⬍0.0001), maximal rate of contraction (r2⫽0.0802; P⬍0.0001) and relaxation (r2⫽0.1104; P⬍0.0001), time-to-peak shortening (r2⫽0.00912; P⫽0.11) and time to 90% relaxation (r2⫽0.11954; P⬍0.0001); n⫽281. C, Shortening-tracings in MONO, BI, and MULTI myocytes; BI and MULTI myocytes are p16INK4a-positive (yellow) and have short telomeres. D, Ca2⫹ transients and sarcomere shortening. E, Sarcomere shortening (r2⫽0.0311; P⬍0.05; n⫽133), diastolic sarcomere length (r2⫽0.004; P⫽0.26; n⫽271), Ca2⫹ amplitude (r2⫽0.1452; P⬍0.0001; n⫽133) and timing of Ca2⫹ peak (r2⫽0.0410; P⬍0.05; n⫽133). F, Ca2⫹ transient without (r2⫽0.1347; P⬍0.05) and with (r2⫽0.0267; P⫽0.36) caffeine and computed Ca2⫹ fractional release (r2⫽0.2212; P⬍0.05); n⫽34. 392 Circulation Research August 17, 2007 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 5. AP-duration. A, APs of myocytes. B, AP-repolarization in myocytes ⬍20 000 (n⫽14), 20 to 40 000 (n⫽46), and ⬎40 000 m3 (n⫽20). *P⬍0.05 vs ⬍20 000; **P⬍0.05 vs 20 to 40 000. C, AP in neonatal mouse myocyte and AP-repolarization in neonatal (n⫽18) and adult (n⫽80) myocytes. *P⬍0.05 vs neonatal-myocytes. independent from myocyte volume (Figure 7A). In comparison with APslow waveforms, APintermediate and APfast waveforms led to a 33% and 42% decrease in developed Ca2⫹, respectively (Figure 7B). Consistent with results in rodents,17,20 the decrease in Ca2⫹ amplitude mediated by fast repolarization was paralleled by attenuation in Ca2⫹ influx (Figure 7C). To strengthen the role that repolarization has in myocyte function, cells of different size were perfused with 4-aminopyridine which blocks Ito and prolongs the AP21; 4-aminopyridine had a profound effect on large myocytes and a smaller impact on small cells. In the former, 4-aminopyridine showed a marked increase in Ca2⫹ transients together with enhanced sarcomere shortening (Figure 7D and 7E). Blockade of Ito reestablished the physiological properties of old-hypertrophied myocytes by restoring in part Ca2⫹ cycling and contractility (supplemental Figure IV). Thus, the AP profile modulates Ca2⫹ transients and is implicated in the decrease of Ca2⫹ release from the SR with cellular hypertrophy and aging. This defect is reversed by prolongation of the AP. Myocyte Properties in the Senescent Mouse Heart Figure 4. L-type Ca2⫹ current and ECC gain. A, ICaL in myocytes. B, ICaL in myocytes ⬍20 000 (n⫽26 and 28), 20 to 40 000 (n⫽22 and 25), and ⬎40 000 m3 (n⫽10 and 10). Results are mean⫾SEM. *P⬍0.05 vs ⬍20 000. C, ICaL and Ca2⫹ transients. D, Voltage relations for ICaL and Ca2⫹-transients in myocytes ⬍20 000 (n⫽24), 20 to 40 000 (n⫽21), and ⬎40 000 m3 (n⫽30). E, ECC-gain. The recognition that young and old myocytes with distinct mechanical and electrical properties are present in the developing heart raised the question whether chronological myocardial aging is characterized similarly by a pool of heterogeneous myocytes with different functional behavior. LV myocytes were obtained from mice at 22 months of age and analyzed. The senescent mouse heart contained myocytes Rota et al Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 which varied in volume from 1600 to 120 000 m3 (supplemental Figure V). Therefore, small and large myocytes were present in the old heart mimicking the observations in young animals. With respect to the young heart, however, chronological age resulted in a shift to the right in the distribution of myocyte volume. Consistent with previous results,7 the percentage of cells positive for p16INK4a and with markedly shortened telomeres increased with chronological age (Figure 8A through 8C). Clusters of myocytes at both extremes of the spectrum were present in the young and old heart (Figures 2C through 2E and 8A through 8C; supplemental Figure V). Heterogeneity in the myocyte characteristics persisted with age although a change in the relative proportion of young and old cells varied in the developing and senescent heart. Measurements of sarcomere shortening, Ca2⫹ transients and AP duration in myocytes collected from old hearts decreased progressively with the enlargement of the cells (Figure 8D and 8E). Conversely, transient outward K⫹ current Ito was linearly related to myocyte volume (Figure 8F). As anticipated, small myocytes with prolonged AP showed enhanced contractile performance, paralleling the observations in the young heart (Figures 3A through 3E, 5A through 5C, 6A, and 6B; supplemental Figure IIIB). When myocytes from young and old hearts were compared, chronological age was characterized by a consistent prolongation of the AP in cells of all classes although this difference was not statistically significant. This phenomenon suggests that a change in the AP of myocytes in the senescent heart tends to compensate for the attenuation in the biophysical parameters late in life in this model. Discussion The myocyte compartment of the young adult mouse heart is heterogeneous and characterized by the coexistence of cells of different age. Old myocytes are unable to replicate, express aging-associated proteins, and show a significant degree of cellular hypertrophy. Cellular aging is accompanied by a progressive increase in volume of myocytes which together with the expression of p16INK4a and telomeric shortening document the complex dynamism of the adult heart. The recognition that senescent and nonsenescent myocytes are present in the mouse heart at 3 months of age supports the notion that these cells do not have a common origin and are the product of a time-dependent activation and differentiation of distinct populations of progenitor cells. Functionally, cellular aging leads to alterations in the electromechanical properties of myocytes which in combination with abnormalities in Ca2⫹ handling negatively affect contractile performance. Collectively, these data document that by necessity the heart constantly repopulates its myocyte compartment to replace senescent poorly-contracting dying myocytes with younger more efficient cells. Thus, cardiac homeostasis and myocyte turnover regulate cardiac function and organ aging. Although similarities have been found between agedmyocytes in the young and old mouse heart, in the latter case senescent cells are exposed to additional stresses dictated by defects in large conductive arteries together with humoral changes and cardiovascular diseases which Myocyte Heterogeneity 393 occur with chronological age. These variables point to the complexity of organ and organism aging and the difficulty to discern the causative role of individual factors in myocyte aging. Myocyte Aging So far, chronological age has formed the premise of multiple studies in rodents and large mammals including humans.4 Comparisons have been made among groups of animals or individuals of different age and aging effects have been identified and interpreted on this basis. With this approach, however, the consequences of aging of other organs and organism cannot be separated from intrinsic aging effects on the heart. The underlying assumption is that chronological age and myocyte age coincide and the heart is composed of cells of identical age with life expectancy which coincides with that of the organism.22 Findings in the current study do not support this traditional view of the biology of the heart but strongly suggest that myocardial aging is mediated by alterations in cell turnover and physiological balance between new and old myocytes. This imbalance leads, as demonstrated here, to a disproportionate accumulation of senescent cells in combination with negative ventricular remodeling and heart failure.7,11 The mechanisms of cellular senescence are difficult to identify. Mutations of mitochondrial DNA, oxidative stress with damage to proteins, lipids and nucleic acids, telomeric shortening, and genetic modifications are all implicated in the onset of cellular aging and the pace by which this process reaches the final stage of senescence and impaired cell function.12,23,24 We have elected to measure telomere length and the expression of p16INK4a because telomere attrition occurs in amplifying myocytes during proliferation, differentiation, and the acquisition of the adult phenotype.6,7,11 Thereafter, telomeric shortening is mediated by cumulative oxidative stress which is a time-dependent process.13 Severe telomere dysfunction upregulates p16INK4a and p53 and ultimately activates the apoptotic pathway.12 Myocyte volume was found to represent a critical variable of cellular senescence together with the inevitable outcome of depressed mechanical behavior. Therefore, myocyte aging is a selfautonomous process only partly related to chronological age. Myocyte Size and Number In the young-adult mouse heart, 40% of myocytes were ⬍20 000 m3 in volume and comprised 20% of the LV-mass whereas 41% of myocytes were 20 000 to 40 000 m3 and occupied 42% of the LV. Old myocytes, ⬎40 000 m3 in volume, represented 19% of the cell pool and formed 38% of the muscle mass. In the old mouse heart, cells with volume ⬍20 000 m3 accounted for 31% of myocytes and 11% of the organ. Myocytes 20 000 to 40 000 m3 were 35% of the cells and constituted 30% of the myocardium. Myocytes ⬎40 000 m3 corresponded to 34% of cells and 59% of the tissue. Therefore, myocyte heterogeneity is present early in life and is potentiated in the old heart. Although the relative contribution of myocyte classes to ventricular function could not be determined in the young or old heart, in both cases small young myocytes have superior contractile ability and 394 Circulation Research August 17, 2007 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 6. Outward transient K⫹ current. A, Ito in myocytes. B, Ito-V relations in myocytes ⬍20 000 (n⫽9), 20 to 40 000 (n⫽10), and ⬎40 000 m3 (n⫽10). *P⬍0.05 vs ⬍20 000; **P⬍0.05 vs 20 to 40 000. C, Kv1.4, Kv4.2, and Kv4.3 mRNA in small and large myocytes (n⫽5). D and E, RT-PCR products had the expected MW and nucleotide sequences. F, Kv4.3 in small and large myocytes. may counteract partly the depressed mechanical behavior of hypertrophied-senescent myocytes. The electromechanical properties of these cells classes were independent from their anatomical location and differently affected large proportion of the myocyte population in the young and senescent heart. Studies in transgenic mice with cardiac restricted overexpression of IGF-1 and Akt support this possibility.5,7 In both cases, the genetic modifications result in an increase in the number of myocytes which are smaller in volume, possess enhanced mechanical performance, potentiate LV function, and delay cardiac aging.5,7,25 In a manner similar to that observed here in small myocytes, telomeres are longer and the fraction of p16INK4a-positive cells is markedly reduced in IGF-1 and Akt transgenic myocytes. These cellular properties potentiate the myocardial response of the heart to acute and chronic ischemic damage, genetic myopathy, and diabetes.25,26 Myocyte Physiology Excitation-contraction coupling is modulated by changes in the expression and phosphorylation of proteins27,28 that regulate transmembrane Ca2⫹ influx and extrusion, release, and uptake of Ca2⫹ from the SR, and responsiveness and binding affinity of the myofilaments to Ca2⫹. AP profile has profound effects on calcium cycling and myocyte mechanics. A prolongation of the repolarization phase may involve an increase in ICaL which in turn enhances Ca2⫹ release from the SR and, thereby, myocyte contractility.17 The results here provide strong evidence in favor of the critical role that AP duration and ICaL play in potentiating Ca2⫹ cycling and mechanical behavior of young-small myocytes or in decreasing Ca2⫹ transients and the performance of senescent-hypertrophied cells. Three statements of caution have to be made. Myocyte mechanics were assessed in unloaded cells, which has limi- Rota et al Myocyte Heterogeneity 395 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 6. Continued tations in the extrapolation of results to in vivo settings. The effects of changes in AP profile on SR Ca2⫹ load were not determined. And we cannot exclude that the isolation protocol impacted differently on cells of different size and age affecting their electromechanical behavior. The AP is modulated by the transient outward K⫹ current Ito which is influenced by the expression of K⫹ channel subunits29 as documented here at the mRNA and protein levels. The expression of Kv1.4, Kv4.2 and Kv4.3 varied in young and senescent myocytes and these differences may 396 Circulation Research August 17, 2007 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 7. AP and Ca2⫹ transients. A, APs with slow, intermediate, and fast repolarization (upper panel) were used as voltage clamp commands and Ca2⫹ transients recorded. B, Ca2⫹ transients with different APs (n⫽11). Neonatal myocytes (n⫽4). Traces and values were normalized for Ca2⫹ transients after APslow. C, ICaL in AP-clamp. Ca2⫹ influx plotted vs 70% repolarization time of the respective APs (r2⫽0.9999; P⬍0.05, n⫽4). D and E, Differential effect of 4-aminopyridine on Ca2⫹ transients and cell shortening in myocytes⬍20 000 (n⫽8) and ⬎20 000 m3 (n⫽12). Traces are normalized to baseline value. represent the molecular mechanisms responsible for the alterations in transient outward K⫹ current and AP with myocyte aging and hypertrophy. Changes in Ito have been reported in myocytes as a function of chronological age.30,31 However, they are at variance with the current results because AP prolongations in rats have been considered a successful adaptation of old cells.20,32 Additionally, gender differences in myocyte behavior have been reported in mice.33 Our observations suggest that these potential differences may be accounted for by the type of cells sampled. A large variability has been found in relation to the size of the cells and expression of aging-associated proteins and telomeric length. Our results point to the need of distribution analysis of the myocyte electromechanical properties to avoid sampling bias and partial characterization of the physiological behavior of aging myocytes. Myocyte heterogeneity is not restricted to the young and old mouse heart. Similar findings have been reported in rats, Rota et al Myocyte Heterogeneity 397 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 dogs and humans, and pathological conditions and aging potentiate this phenomenon.6,34 However, it is difficult to predict whether small developing myocytes in the adult stressed human heart6 may have a longer repolarization phase, enhanced Ca2⫹ transients, and increased cell shortening. Human myocytes exhibit a pronounced plateau phase of the AP which is not present in rodents.35 Whether Ito plays a comparable or lesser role in the regulation of AP, Ca2⫹ cycling and mechanical behavior in humans is an important unanswered question. A comment has to be made concerning the electromechanical properties of aging myocytes. Ca2⫹ load of the SR and ICaL are not altered. Similarly, the ability of L-type current to trigger the activation of the RyR and Ca2⫹ release from the SR is not impaired. This suggests that the machinery underlying Ca2⫹ cycling is preserved in old cells and that cellular aging is dictated by changes in electrical activity which negatively modulate excitation-contraction coupling. In senescent myocytes, the faster repolarization of the AP leads to decreased Ca2⫹ transients and faster time to peak Ca2⫹. However, these abnormalities in Ca2⫹ metabolism can be reversed by prolongation of AP which restores the mechanical performance of old myocytes. Collectively, our observations point to electrical remodeling as a key modulator of the heterogeneity of myocyte aging and function in the adult mammalian heart. Sources of Funding Figure 7. Continued Supported by NIH grants and by MRC Strategic Grant G0300395. 398 Circulation Research August 17, 2007 Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 Figure 8. Myocyte aging in the old heart. A, p16INK4a-positive MONO (n⫽1744), BI (n⫽608), and MULTI (n⫽420) myocytes. *P⬍0.05 vs MONO; **P⬍0.05 vs BI. B, Distribution of telomere length vs myocyte volume (n⫽180) was fitted with a straight line (r2⫽0.312; P⬍0.0001). C, Telomere length in myocytes ⬍20 000 (n⫽34), 20 to 40 000 (n⫽79), and ⬎40 000 m3 (n⫽67). *P⬍0.05 vs ⬍20 000. D, Bivariate distribution of Ca2⫹ transients (r2⫽0.0534; P⬍0.05) and cell shortening (r2⫽0.0861; P⬍0.05) vs myocyte volume (n⫽89). E, AP-repolarization in myocytes ⬍20 000 (n⫽8) and ⬎20 000 m3 (n⫽10). F, Ito-V relations in myocytes ⬍20 000 (n⫽6) and ⬎20 000 m3 (n⫽10). *P⬍0.05 vs ⬍20 000. Disclosures None. References 1. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, NadalGinard B, Anversa P. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003;114:763–776. 2. Urbanek K, Cesselli D, Rota M, Nascimbene A, De Angelis A, Hosoda T, Bearzi C, Boni A, Bolli R, Kajstura J, Anversa P, Leri A. Stem cell niches in the adult mouse heart. Proc Natl Acad Sci U S A. 2006;103: 9226 –9231. 3. Anversa P, Kajstura J, Leri A, Bolli R. Life and death of cardiac stem cells: a paradigm shift in cardiac biology. Circulation. 2006;113: 1451–1463. 4. Lakatta EG. Arterial and cardiac aging: major shareholders in cardiovascular disease enterprises: Part III: cellular and molecular clues to heart and arterial aging. Circulation. 2003;107:490 – 497. 5. Rota M, Boni A, Urbanek K, Padin-Iruegas ME, Kajstura TJ, Fiore G, Kubo H, Sonnenblick EH, Musso E, Houser SR, Leri A, Sussman MA, Anversa P. Nuclear targeting of Akt enhances ventricular function and myocyte contractility. Circ Res. 2005;97:1332–1341. Rota et al Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 6. Urbanek K, Quaini F, Tasca G, Torella D, Castaldo C, Nadal-Ginard B, Leri A, Kajstura J, Quaini E, Anversa P. Intense myocyte formation from cardiac stem cells in human cardiac hypertrophy. Proc Natl Acad Sci U S A. 2003;100:10440 –10445. 7. Torella D, Rota M, Nurzynska D, Musso E, Monsen A, Shiraishi I, Zias E, Walsh K, Rosenzweig A, Sussman MA, Urbanek K, Nadal-Ginard B, Kajstura J, Anversa P, Leri A. Cardiac stem cell and myocyte aging, heart failure, and insulin-like growth factor-1 overexpression. Circ Res. 2004; 94:514 –524. 8. Beusejour CM, Campisi J. Ageing: balancing regeneration and cancer. Nature. 2006;443:404 – 405. 9. Anversa P, Loud AV, Giacomelli F, Wiener J. Absolute morphometric study of myocardial hypertrophy in experimental hypertension. II. Ultrastructure of myocytes and interstitium. Lab Invest. 1978;38:597– 609. 10. Chen X, Wilson RM, Kubo H, Berretta RM, Harris DM, Zhang X, Jaleel N, Macdonnell SM, Bearzi C, Tillmanns J, Trofimova I, Hosoda T, Mosna F, Cribbs L, Leri A, Kajstura J, Anversa P, Houser SR. Adolescent feline heart contains a population of small, proliferative ventricular myocytes with immature physiological properties. Circ Res. 2007;100: 536 –544. 11. Chimenti C, Kajstura J, Torella D, Urbanek K, Heleniak H, Colussi C, Di Meglio F, Nadal-Ginard B, Frustaci A, Leri A, Maseri A, Anversa P. Senescence and death of primitive cells and myocytes leads to premature cardiac aging and heart failure. Circ Res. 2003;93:604 – 613. 12. Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet. 2005;6:611– 622. 13. von Zglinicki T. Role of oxidative stress in telomere length regulation and replicative senescence. Ann N Y Acad Sci. 2000;908:99 –110. 14. Fabiato A. Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am J Physiol. 1983;245:C1–C14. 15. Rota M, Vassalle M. Patch-clamp analysis in canine cardiac Purkinje cells of a novel sodium component in the pacemaker range. J Physiol. 2003;548:147–165. 16. Bouchard RA, Clark RB, Giles WR. Effects of action potential duration on excitation-contraction coupling in rat ventricular myocytes. Action potential voltage-clamp measurements. Circ Res. 1995;76:790 – 801. 17. Sah R, Ramirez RJ, Oudit GY, Gidrewicz D, Trivieri MG, Zobel C, Backx PH. Regulation of cardiac excitation-contraction coupling by action potential repolarization: role of the transient outward potassium current (Ito). J Physiol. 2003;546:5–18. 18. Apkon M, Nerbonne JM. Characterization of two distinct depolarizationactivated K⫹ currents in isolated adult rat ventricular myocytes. J Gen Physiol. 1991;97:973–1011. 19. Kaab S, Nuss HB, Chiamvimonvat N, O’Rourke B, Pak PH, Kass DA. Marban E, Tomaselli GF. Ionic mechanism of action potential prolon- 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. Myocyte Heterogeneity 399 gation in ventricular myocytes from dogs with pacing-induced heart failure. Circ Res. 1996;78:262–273. Janczewski AM, Spurgeon HA, Lakatta EG. Action potential prolongation in cardiac myocytes of old rats is an adaptationt sustain youthful intracellular Ca2⫹ regulation. J Mol Cell Cardiol. 2002;34:641– 648. Volk T, Nguyen TH, Schultz JH, Ehmke H. Relationship between transient outward K⫹ current and Ca2⫹ influx in rat cardiac myocytes of endo- and epicardial origin. J Physiol. 1999;519:841– 850. Rubart M, Field LJ. Cardiac regeneration: repopulating the heart. Annu Rev Physiol. 2006;68:29 – 49. Balaban RS, Nemoto S, Finkel T. Mitochondria, oxidants, and aging. Cell. 2005;120:483– 495. Guarente L, Kenyon C. Genetic pathways that regulate ageing in model organisms. Nature. 2000;408:255–262. Webster KA. Aktion in the nucleus. Circ Res. 2004;94:856 – 859. Anversa P. Aging and longevity: the IGF-1 enigma. Circ Res. 2005;97: 411– 414. Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415: 198 –205. Fitzsimons DP, Moss RL. Cooperativity in the regulation of force and the kinetics of force development in the heart and skeletal muscles: crossbridge activation of force. Adv Exp Med Biol. 2007;592:177–189. Wickenden AD, Kaprielian R, Parker TG, Jones OT, Backx PH. Effects of development and thyroid hormone on K⫹ currents and K⫹ channel gene expression in rat ventricle. J Physiol. 1997;504:271–286. Liu SJ, Wyeth RP, Melchert RB, Kennedy RH. Aging-associated changes in whole cell K⫹ and L-type Ca2⫹ currents in rat ventricular myocytes. Am J Physiol. 2000;279:H889 –H900. Chouabe C, Ricci E, Amsellem J, Blaineau S, Dalmaz Y, Favier R, Pequignot JM, Bonvallet R. Effects of aging on the cardiac remodeling induced by chronic high altitude. Am J Physiol. 2004;287:H1246 –H1253. Xiao RP, Tomhave ED, Wang DJ, Ji X, Boluyt MO, Cheng H, Lakatta EG, Koch WJ. Age-associated reductions in cardiac beta1- and beta2adrenergic responses without changes in inhibitory G proteins or receptor kinases. J Clin Invest. 1998;101:1273–1282. Grandy SA, Howlett SE. Cardiac excitation-contraction coupling is altered in myocytes from aged male mice but not in cells from aged female mice. Am J Physiol. 2006;291:H2362–H2370. Fraticelli A, Josephson R, Dazinger R, Lakatta E, Spurgeon H. Morphological and contractile characteristics of rat cardiac myocytes from maturation to senescence. Am J Physiol. 1989;257:257:H259 –H265. Akar FG, Wu RC, Deschenes I, Armoundas AA, Piacentino V, Houser SR, Tomaselli GF. Phenotypic differences in transient outward K⫹ current of human and canine ventricular myocytes: insights into molecular composition of ventricular Ito. Am J Physiol. 2004;286: H602–H609. Downloaded from http://circres.ahajournals.org/ by guest on June 11, 2017 The Young Mouse Heart Is Composed of Myocytes Heterogeneous in Age and Function Marcello Rota, Toru Hosoda, Antonella De Angelis, Michael L. Arcarese, Grazia Esposito, Roberto Rizzi, Jochen Tillmanns, Derin Tugal, Ezio Musso, Ornella Rimoldi, Claudia Bearzi, Konrad Urbanek, Piero Anversa, Annarosa Leri and Jan Kajstura Circ Res. 2007;101:387-399; originally published online June 29, 2007; doi: 10.1161/CIRCRESAHA.107.151449 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 2007 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/101/4/387 Data Supplement (unedited) at: http://circres.ahajournals.org/content/suppl/2007/06/29/CIRCRESAHA.107.151449.DC1 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/ Online Supplement Rota et al., The Young Mouse Heart Is Composed… Materials and Methods Animals A total of 60 C57Bl/6 mice at 3 months of age, 10 mice at 22 months, and 3 litters of neonatal mice were used. In each study, a minimum of 5 mice were employed. In a group of 5 adult mice, BrdU, 50 mg per kg body weight, was injected twice a day for three days before sacrifice. Animals were maintained in accordance with the Guide for Care and Use of Laboratory Animals and the experiments were approved by New York Medical College. Myocyte Isolation Following chloral hydrate anesthesia (400 mg/kg body weight, i.p.), the heart was excised and left ventricular (LV) myocytes were enzymatically dissociated (1, 2). Briefly, the myocardium was perfused retrogradely through the aorta at 37°C with a Ca2+-free solution gassed with 85% O2 and 15% N2. After 5 minutes, 0.1 mmol/L CaCl2, 274 units/mL collagenase (type 2, Worthington Biochemical Corp, Lakewood, NJ) and 0.57 units/mL protease (type XIV, Sigma, St. Louis, MO) were added to the solution which contained (mmol/L): NaCl 126, KCl 4.4, MgCl2 5, HEPES 5, Glucose 22, Taurine 20, Creatine 5, Na Pyruvate 5 and NaH2PO4 5 (pH 7.4, adjusted with NaOH). At completion of digestion, the LV was cut in small pieces and re-suspended in Ca2+ 0.1 mmol/L solution. In 10 hearts, the epicardium, endocardium, base and apex of the left ventricle were dissected and myocytes obtained from these anatomical regions. Similarly, neonatal myocytes were enzymatically dissociated from the heart of C57Bl/6 mice at 1 day after birth. Mice were decapitated and the heart was quickly removed and placed in Ca/Mgfree Hanks’ balanced salt solution (HBSS, Sigma). After removal of the atria, the tissue was cut in small pieces which were transferred into 10 ml dissociation medium (0.1% trypsin, 1:250, and 0.01% DNase I in HBSS). After 25 minutes of gentle stirring at 37°C the supernatant was collected and the cells contained in this fraction were centrifuged at 1 Online Supplement Rota et al., The Young Mouse Heart Is Composed… 300 g for 4 min, resuspended in Eagle’s MEM (Sigma) supplemented with 10% FBS (Sigma) and stored on ice. Isolated cells were pre-plated in a 150-mm Petri dish for 1 hour at 37°C, and myocytes which did not attach to the dish during this time were collected and plated in 35-mm Petri dish at low density (500/mm2) and cultured in Dulbecco’s MEM (Sigma) with 10% FBS (3). Patch-clamp experiments were performed utilizing 24-48 hours cultured neonatal myocytes. Myocyte Volume For the determination of myocyte volume, cells were isolated from 20 and 5 mice at 3 and 22 months of age, respectively. Myocytes were washed in PBS and fixed for 20 minutes in 4% paraformaldehyde (1, 4). Myocytes were stained with α-sarcomeric actin mouse monoclonal antibody (Sigma) and connexin 43 rabbit polyclonal antibody (Sigma) to identify cell aggregates. Nuclei were recognized by 4'-6-diamidino-2-phenylindole (DAPI) staining (Sigma). In physiological studies, cell length and width were measured with a computerized image analysis system and the ratio of the minor to the major axis of the ellipse was obtained by confocal microscopy. Cell volume was then computed assuming an elliptical cross-section with the major axis equivalent to cell width; the minor axis was computed from the measured ratios. In morphological studies, the volume of freshly isolated paraformaldehyde–fixed myocytes in each cell class was obtained by three-dimensional optical section reconstruction by confocal microscopy (Bio-Rad Radiance 2100, Hercules, CA). Three-dimensional reconstruction of isolated myocytes was computed utilizing Imaris software (Bitplane AG, Zurich, Switzerland). These preparations were also used for the determination of apoptosis (see below). Immunocytochemistry Immunocytochemistry was performed on 4% paraformaldehyde-fixed isolated myocyte preparations. Nuclei were stained by DAPI or PI. p16INK4a (F-12 antibody, Santa Cruz Biotechnology, Santa Cruz, CA; 2, 5, 6), Kv4.3 (Chemicon, Temecula, CA), αsarcomeric actin (Sigma), BrdU (Roche Molecular Biochemistry, Indianapolis, IN), Ki67 2 Online Supplement Rota et al., The Young Mouse Heart Is Composed… (Vector, Burlingame, CA) and phospho-H3 (Upstate Biotechnology, Charlottesville, VA) antibodies were employed. Apoptosis was detected by the TUNEL assay (BD Pharmingen, San Jose, CA). Telomere Length in Isolated Myocytes Telomere length in nuclei was evaluated in 4% paraformaldehyde-fixed isolated myocyte preparations by quantitative fluorescence in situ hybridization (Q-FISH) and confocal microscopy. An FITC-peptide nucleic acid probe was used (2, 5-7). The fluorescent signal measured in lymphoma cells (L5178Y) with short (7 kbp) and long (48 kbp) telomeres (kindly provided to us by Dr. M.A. Blasco, Madrid, Spain) were utilized to compute absolute length of telomeres. Fluorescence intensity was measured utilizing ImagePro software (Media Cybernetics, Silver Spring, MD). Sarcomere Shortening and Ca2+ Transients Isolated LV myocytes were placed in a bath on the stage of an Axiovert Zeiss Microscope (Zeiss, Germany) and IX71 Olympus inverted microscope (Olympus, Melville, NY) for contractility, Ca2+ transients and patch-clamp measurements. Experiments were conducted at room temperature. Cells were bathed continuously with a Tyrode solution containing (mmol/L): NaCl 140, KCl 5.4, MgCl2 1, HEPES 5, Glucose 5.5 and CaCl2 1.0 (pH 7.4, adjusted with NaOH). Measurements were performed in fieldstimulated cells by using IonOptix fluorescence and contractility systems (IonOptix, Milton, MA). Contractions were elicited by rectangular depolarizing pulses, 2 ms in duration, and twice-diastolic threshold in intensity, by platinum electrodes (1, 2, 8, 9). Cell shortening was measured by edge-track detection system. Changes in mean sarcomere length were computed by determining the mean frequency of sarcomere spacing by fast Fourier transform and then frequency data were converted to length. Ca2+ transients were measured by epifluorescence after loading the myocytes with 10 µM Fluo-3 AM (Invitrogen, Carlsbad, CA). Excitation length was 480 nm with emission collected at 535 nm using a 40x oil objective. Fluo-3 signals were expressed as 3 Online Supplement Rota et al., The Young Mouse Heart Is Composed… normalized fluorescence (F/F0). Caffeine–induced Ca2+ transient was evoked by rapid application of 20 mmol/L caffeine to assess SR-Ca2+ load (1, 10). In field-stimulated cells, the prolongation of the action potential was achieved by perfusion with 0.5 mmol/L 4-aminopyridine (11). Patch-Clamp Experiments Data were acquired by means of the whole-cell patch-clamp technique in voltage-clamp mode using a Multiclamp 700A amplifier (Axon Instruments, Union City, CA). Electrical signals were digitized using a 500 kHz 16-bit resolution A/D converter (Digidata 1322, Axon Instruments) and recorded using pCLAMP 9.0 software (Axon Instruments) with low-pass filtering at 2 kHz (1, 2). L-type Ca2+ Current L-type Ca2+ Current (ICaL) properties were assessed using a Na+-K+ free perfusing solution (11) of the following composition (mmol/L): N-methyl-D-glucamine (NMDG) 140, CsCl 4, MgCl2 1, HEPES 5, glucose 5.5, CaCl2 1 and 4-aminopyridine 2 (pH 7.4 with CsOH). The composition of the pipette solution was (mmol/L): NMDG 10, CsCl 113, MgCl2 0.5, Tris-ATP 5, glucose 5.5, HEPES 10, EGTA 5 and TEA-Cl 20 (pH 7.2 with CsOH). The pipettes were pulled by means of a Narishige PB-7 glass microelectrode puller (Narishige, Tokyo, Japan) and when filled had a resistance of 1-2 MΩ. ICaL current-voltage (I-V) relation and activation properties were determined applying depolarizing steps 300 ms in duration from holding potential (Vh) -70 mV in 10 mV increments. For inactivation properties a 300 ms preconditioning step was applied within the range -70 to +60 mV in 10 mV increments prior to the 300 ms test step to 0 mV. The half maximal activation potential and the potential giving 50% inactivation for ICaL was obtained by fitting the activation and inactivation curves with a Boltzmann equation (1, 12). Inactivation time course of ICaL was described by fitting the current traces between the inward peak and the end of the 300 ms pulse to 0 mV (from Vh -70 mV) with a biexponential function (1). Membrane capacitance (Cm) was calculated using a 5 mV voltage step. 4 Online Supplement Rota et al., The Young Mouse Heart Is Composed… Excitation-Contraction Coupling Gain For excitation-contraction (EC) coupling gain measurements (13, 14), Ca2+ current and Ca2+ transient were measured simultaneously in voltage-clamped myocyte. Myocytes were loaded with Fluo-3 AM and bathed with a modified Tyrode solution in which KCl was replaced with CsCl and 2 mmol/L 4-aminopyridine was added to the solution. The composition of the pipette solution was (mmol/L): NaCl 10, CsCl 113, MgCl2 0.5, TrisATP 5, Glucose 5.5, HEPES 10 and tetraethylammonium chloride 20 (pH 7.2 with CsOH). Fluorescence signal intensity was collected using a photomultiplier and a photon to voltage converter (IonOptix) connected to the patch-clamp A/D converter. Cells were depolarized for 300 ms from Vh -40 mV to a family of test potentials ranging from -30 to +70. Each voltage-clamp was preceded by a train of 4 conditioning pulses (100 ms, 1Hz) form -40 to 0 mV. Action Potentials, K+ current and Action Potential-Clamp For action potential (AP) measurements, current-clamp mode was utilized. Cells were stimulated at 1 Hz with current pulses 1.5 times threshold. Myocyte were bathed with Tyrode solution as described above. The composition of the pipette solution was (mmol/L): NaCl 10, KCl 113, MgCl2 0.5, K2-ATP 5, Glucose 5.5, EGTA 5, HEPES 10 (pH 7.2 with KOH). For action potential clamp (AP-clamp) experiments a set of action potential waveforms were utilized as voltage-clamp command in myocytes loaded with Fluo-3 AM. Solution were similar to the one utilized for current-clamp experiments. To assess Ca2+ influx during AP-clamp studies, specific solutions were employed to record ICaL. Ca2+ influx was measured as the integral of the inward current over time normalized by membrane capacitance (11). Transient outward K+ current (Ito) was assessed in voltage-clamp mode with Tyrode solution containing 0.3 mmol/L cadmium chloride to block ICaL. Pipette solution was similar to that utilized for AP measurements (15, 16). Current-voltage (I-V) relations were determined applying depolarizing steps 500 ms in duration from holding potential (Vh) -40 mV in 10 mV increments. Ito amplitude was 5 Online Supplement Rota et al., The Young Mouse Heart Is Composed… measured as the difference between the peak outward current at the beginning of the step and Isus, the current at the end of the 500 ms step (15, 16). Real-Time RT-PCR Isolated myocytes were separated according to their size utilizing a 20 µm mesh filter (CellTrics, Partec, Munster, Germany) (17). After centrifugation, cell pellets were snapfrozen in liquid nitrogen for real time RT-PCR analysis of the transcripts of Kv1.2, Kv4.2 and Kv4.3 (18). RNA was extracted utilizing Trizol reagent (Sigma) and treated with DNase I (Sigma). Reverse transcription reaction was performed with 2 µg of total RNA, oligo(dT) primer and 200 units of SuperScript III (Invitrogen) at 50°C for 3.5 hours. Samples were then treated with RNase H (Ambion, Austin, TX). RT-PCR was performed in the Real Time PCR System 7300 (Applied Biosystems, Warrington, UK) utilizing 1/20th of the synthesized cDNA together with 0.625 units of AmpliTaq Gold DNA polymerase (Applied Biosystems), 0.5 µM each of forward and reverse primers, and 0.1 µM of FAM-labeled probe for each target gene; 0.4 µg of total RNA was included as a non-RT control sample. PCR conditions were as follows: 95°C for 10 min, 40 cycles of 95°C for 15 sec and 60°C for 1 min. FAM fluorescence was measured during the second step at the end of each cycle. Primers and probe of each gene were designed on the same exon. Sequences of primers and probes were: mKv1.4 F: 5'- AGA GGC GGA TGA ACC CAC TAC -3' (59°C) P: 5'- CAT TTC CAA AGC ATT CCA GAT GCG TTT TG -3' (70°C) R: 5'- CGA CTG TGA TGG GCT TCA TG -3' (59°C) amplicon size: 110 bp mKv4.2 F: 5'- AAT GTG TCG GGA AGC CAT AGA -3' (58°C) P: 5'- CCA ACA GCC GAT CCA GCT TAA ATG CC -3' (70°C) R: 5'- ACA GTT TAG TTT AAC ACA CTC TTC CAT TTT -3' (58°C) 6 Online Supplement Rota et al., The Young Mouse Heart Is Composed… amplicon size: 135 bp mKv4.3 F: 5'- CAG CAC GCT CCA CAT CCA -3' (59°C) P: 5'- CAG TCG CTC CAG CCT TAA TTT GAA AGC AGA -3' (70°C) R: 5'- TGG TAA TCT GGG ATG TTT TGC A -3' (59°C) amplicon size: 116 bp The housekeeping gene β-actin was used to normalize PCR conditions: mActb F: 5'- AGA AGG AGA TTA CTG CTC TGG CTC -3' (58°C) P: 5'- CTA GCA CCA TGA AGA TCA AGA -3' (72°C) R: 5'- ACA TCT GCT GGA AGG TGG ACA -3' (59°C) amplicon size: 126bp The expression profile of each target gene was analyzed using Automatic Baseline of Sequence Detection Software version 1.2.2 (Applied Biosystems). The threshold was fixed manually at 0.05 to determine the cycle of threshold (Ct), which was then normalized by β-actin; the Ct value of β-actin was subtracted from the Ct value of the target genes (∆Ct). The levels of expression were presented as percentage relative to the quantity of β-actin transcript. Data Analysis Data are presented as mean±S.E.M. Significance was determined by Student’s t test and Bonferroni method (19, 20). Bivariate distribution plots were fitted by straight line function (19). P<0.05 was considered significant. 7 Online Supplement Rota et al., The Young Mouse Heart Is Composed… References 1. Rota M, Boni A, Urbanek K, Padin-Iruegas ME, Kajstura TJ, Fiore G, Kubo H, Sonnenblick EH, Musso E, Houser SR, Leri A, Sussman MA, Anversa P. Nuclear targeting of Akt enhances ventricular function and myocyte contractility. Circ Res. 2005; 97:1332-1341. 2. Torella D, Rota M, Nurzynska D, Musso E, Monsen A, Shiraishi I, Zias E, Walsh K, Rosenzweig A, Sussman MA, Urbanek K, Nadal-Ginard B, Kajstura J, Anversa P, Leri A. Cardiac stem cell and myocyte aging, heart failure, and insulin-like growth factor-1 overexpression. Circ Res. 2004; 94: 514-524. 3. Kajstura J, Pertoldi B, Leri A, Beltrami CA, Deptala A, Darzynkiewicz Z, Anversa P. Telomere shortening is an in vivo marker of myocyte replication and aging. Am J Pathol. 2000;156:813-819. 4. Limana F, Urbanek K, Chimenti S, Quaini F, Leri A, Kajstura J, Nadal-Ginard B, Izumo S, Anversa P. bcl-2 overexpression promotes myocyte proliferation. Proc Natl Acad Sci U S A. 2002; 99: 6257-6262. 5. Chimenti C, Kajstura J, Torella D, Urbanek K, Heleniak H, Colussi C, Di Meglio F, Nadal-Ginard B, Frustaci A, Leri A, Maseri A, Anversa P. Senescence and death of primitive cells and myocytes leads to premature cardiac aging and heart failure. Circ Res. 2003; 93: 604–613. 6. Rota M, LeCapitaine N, Hosoda T, Boni A, De Angelis A, Padin-Iruegas ME, Esposito G, Vitale S, Urbanek K, Casarsa C, Giorgio M, Luscher TF, Pelicci PG, Anversa P, Leri A, Kajstura J. Diabetes promotes cardiac stem cell aging and heart failure, which are prevented by deletion of the p66shc gene. Circ Res. 2006;99:42-52. 7. Leri A, Franco S, Zacheo A, Barlucchi L, Chimenti S, Limana F, Nadal-Ginard B, Kajstura J, Anversa P, Blasco MA. Ablation of telomerase and telomere loss leads to cardiac dilatation and heart failure associated with p53 upregulation. EMBO J. 2003;22:131-139. 8 Online Supplement Rota et al., The Young Mouse Heart Is Composed… 8. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, Anversa P. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003; 114: 763-776. 9. Urbanek K, Rota M, Cascapera S, Bearzi C, Nascimbene A, De Angelis A, Hosoda T, Chimenti S, Baker M, Limana F, Nurzynska D, Torella D, Rotatori F, Rastaldo R, Musso E, Quaini F, Leri A, Kajstura J, Anversa P: Cardiac stem cells possess growth factorreceptor systems that after activation regenerate the infarcted myocardium, improving ventricular function and long-term survival. Circ Res. 2005;97:663-673. 10. DuBell WH, Houser SR. Rest decay of calcium transients and contractility in feline ventricular myocytes. Am J Physiol. 1990;259:H395-H402. 11. Volk T, Nguyen TH, Schultz JH, Ehmke H. Relationship between transient outward K+ current and Ca2+ influx in rat cardiac myocytes of endo- and epicardial origin. J Physiol. 519:841-850, 1999. 12. Chen X, Piacentino V 3rd, Furukawa S, Goldman B, Margulies KB, Houser SR. Ltype Ca2+ channel density and regulation are altered in failing human ventricular myocytes and recover after support with mechanical assist devices. Circ Res. 2002;91: 517-524. 13. Gomez AM, Guatimosim S, Dilly KW, Vassort G, Lederer WJ. Heart failure after myocardial infarction: altered excitation-contraction coupling. Circulation 2001; 104:688-93. 14. Pott C, Philipson KD, Goldhaber JI. Excitation-contraction coupling in Na+-Ca2+ exchanger knockout mice: reduced transsarcolemmal Ca2+ flux. Circ Res. 2005; 97:12881295. 15. Kaprielian R, Wickenden A D, Kassiri Z, Parker TG, Liu PP, Backx PH. Relationship between K+ channel down-regulation and [Ca2+]i in rat ventricular myocytes following myocardial infarction. J Physiol. 1999;517:229-245. 9 Online Supplement Rota et al., The Young Mouse Heart Is Composed… 16. Wang L, Duff HJ. Developmental changes in transient outward current in mouse ventricle. Circ Res. 1997;81:120-127. 17. Chen X, Wilson RM, Kubo H, Berretta RM, Harris DM, Zhang X, Jaleel N, Macdonnell SM, Bearzi C, Tillmanns J, Trofimova I, Hosoda T, Mosna F, Cribbs L, Leri A, Kajstura J, Anversa P, Houser SR. Adolescent feline heart contains a population of small, proliferative ventricular myocytes with immature physiological properties. Circ Res. 2007; Epub ahead of print. 18. Gong N, Bodi I, Zobel C, Schwartz A, Molkentin JD, Backx PH. Calcineurin increases cardiac transient outward K+ currents via transcriptional up-regulation of Kv4.2 channel subunits. J Biol Chem. 2006;281:38498-38506. 19. Wallenstein S, Zucker CL, Fleiss JL. Some statistical methods useful in circulation research. Circ Res. 1980;47:1-9. 20. Berenson ML, Levine DM, Rindskopf D, eds. Applied statistics, a first course. Englewood Cliffs, New Jersey: Prentice-Hall; l988. 10 Online Supplement Rota et al., The Young Mouse Heart Is Composed… Supplemental Figures Supplemental Figure I. Volume distribution of myocytes. A, Volume of mononucleated (MONO; n=84), binucleated (BI; n=522) and multinucleated (MULTI; n=88) myocytes. *P<0.05 vs MONO; **P<0.05 vs BI. B and C, Volume distribution of myocytes (B, n=694; C, n=2,819). Supplemental Figure II. Cell capacitance. Bivariate distribution of Cm and myocyte volume (r2=0.961; P<0.0001); n=54. Supplemental Figure III. Regional distribution of AP duration and sarcomere shortening. A, AP-repolarization in myocytes <20,000, and >20,000 µm3 from the endocardium (n=11 and 10) and epicardium (n=9 and 12). *P<0.05 vs small myocytes. B, Bivariate distribution of sarcomere-shortening vs cell volume in myocytes from endocardium (r2=0.0915; P<0.05, n=80) and epicardium (r2=0.1135; P<0.05, n=78). Supplemental Figure IV. Ca2+-transients and shortening with 4-aminopyridine. Supplemental Figure V. Volume distribution of myocytes at 3-months (n=2,819) and 22-months (n=613) of age. 11