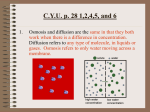
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Metagenomics wikipedia , lookup
Bioinformatics wikipedia , lookup
Gene therapy wikipedia , lookup
Gene expression profiling wikipedia , lookup
Gene nomenclature wikipedia , lookup
Genome editing wikipedia , lookup
Promoter (genetics) wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Gene regulatory network wikipedia , lookup
Real-time polymerase chain reaction wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Silencer (genetics) wikipedia , lookup
Designer baby wikipedia , lookup
Gene prediction wikipedia , lookup
SUPPLEMENTARY INFORMATION Details of Vacuole Purification and Homotypic Vacuole Fusion Methods Freshly purified vacuoles were used in all experiments. The purification procedure that we implemented is described in the literature (Haas, 1995); however, we made a few minor changes. In short, overnight 1.5 liter YPD cultures of OD600 1-1.5 were centrifuged. The pellet was then re-suspended in 35 ml wash buffer (0.1 M PIPES-KOH [pH 9.4], 10 mM DTT), and subsequently incubated at 30oC for 10 minutes. Finally, each cell pellet (+ 5 ml volume) was re-suspended in 10 ml spheroplasting buffer (50 mM KH2PO4 [pH 7.5], 600 mM sorbitol, made up in YEP+0.2% dextrose) containing 10 mg zymolyase 100T (Seikagaku Corporation), and incubated at 30oC for 30 minutes. Osmolarity during spheroplasting influences fusion efficiency of vacuoles after purification. In most cases, the standard high-osmolarity spheroplasting buffer shown above was used (Haas, 1995). However, in the case of the Rpt1-K256S mutant, the effect of the mutation on fusion was only observed using a low-osmolarity spheroplasting buffer (50 mM KH2PO4 [pH 7.5], 300 mM sorbitol, 0.2% glucose, made up in H2O). The spheroplasts were pelleted by centrifugation, and the pellet (typical volume + 4ml) was then re-suspended in an equal volume of 15% Ficoll solution (in 10 mM PIPES-KOH [pH 6.8], 200 mM sorbitol). To make lysis occur, 200 l DEAE- dextran solution (0.4 mg/ml in 15% Ficoll solution, freshly made) was added to the solution, and heat-shock was performed for 90 s at 30oC, after which the solution was chilled again on ice. Subsequently, 4.8 ml of lysate was deposited in the bottom of an SW40 Ti ultracentrifuge 1 tube (Beckman Ultra-Clear, 14x95 mm), and overlayed sequentially with 3 ml 8% Ficoll solution, 3 ml 4% Ficoll solution and 2 ml 0% Ficoll solution. The tubes were centrifuged in an SW40 swinging bucket rotor at 30,000 rpm for 90 minutes at 4oC. The white wafer of membraneous material at the interface between the 4% and 0% Ficoll solution was retrieved and found to be strongly enriched with the Pho8 vacuolar membrane marker (see Figure 1B, bottom panel). The overall protein concentration was determined with a Coomassie-based detection reagent (Pierce), and the vacuolar membrane fractions were adjusted to an overall protein concentration of 0.3 to 0.4 mg/ml with the 0% Ficoll solution. Multiubiquitin chains can be sensitive to degradation during the standard vacuole purification procedure (see also results section Figure 3D). In such cases, protease inhibitors were added to the solutions during vacuole purification (COMPLETE-EDTA tablet; Roche) and complemented with EDTA (EDTA concentration in the spheroplasting buffer of 4 mM and in the Ficoll solutions of 1 mM). For Figure 5D, the high-grade zymolyase 100T (Seikagaku Corporation) concentration was kept as low as possible. High concentrations of zymolyase led to partial Ypt7 deubiquitination during vacuole purification (data not shown). So did lower-grade zymolase. When performing a fusion reaction, 10 l of pho8 vacuoles were mixed with 10 l of pep4 prb1 vacuoles, and added to 10 l of buffer, consisting of 3 l of an ATPase regenerating system (10 mM PIPES-KOH [pH 6.8], 10 mM MgCl2, 10 mg/ml creatine kinase, 95 mg/ml creatine phosphate, 6 mg/ml ATP, 200 mM sorbitol, 10 mM KOH), 3 2 l of a salt-adjustment buffer (1.5 M KCl, 5 mM MgCl2, 10 mM PIPES-KOH [pH 6.8], 200 mM sorbitol), and the remaining 4 l made up with either 0% Ficoll solution (10 mM PIPES-KOH [pH 6.8], 200 mM sorbitol) or specific inhibitors. To make the negative control, we added the solvent that is used to generate the stock solutions of these inhibitors (DMSO) to the fusion reaction. Our procedure in each fusion reaction was as follows. We placed two fusion reaction samples on ice, and incubated three reaction samples at the fusion temperature (27oC). We calculated the mean alkaline phosphatase value of the two ice samples, and subtracted this value from each of the three alkaline phosphatase values of the three samples incubated at the fusion temperature. The error bars shown in the figures represent the standard deviations of the three resulting values. Each figure shows a representative experiment. To generate the data presented in Figure 6B and 6C, equal numbers of cells from the pho8 and pep4 prb1 strains were mixed and then pelleted by centrifugation. We used the mixed cell pellet to purify vacuoles. The purified vacuoles that were generated was consequently a mixture of pho8 and pep4 prb1 vacuoles. This procedure thus enabled us to minimize the variation between vacuolar preparations. The pho8 and pep4 prb1 vacuoles used in these fusion reactions can be considered pre-tethered. In the experiment that led to Figure 6, we used vacuoles that were purified from a strain in which Ypt7 is over-expressed. By contrast, in the experiment that led to Figure 2, we used vacuoles that contained endogenous Ypt7. Because Ypt7 is an ubiquitinated proteasomal substrate with a major role in fusion, it is likely that Ypt7 over-expression influences the proteasome inhibitor’s effect on fusion. 3 In vitro vacuolar fusion requires both ATP and GTP. ATP is exogenously added as a standard component of the fusion reaction buffer. As far as GTP is concerned, in vitro fusion can proceed without supplementing exogenous GTP. Because GTPase function is required for in vitro fusion, the fact that fusion occurs even when exogenous GTP has not been not added, suggests that a sufficient number of the GTPases that co-purify with vacuoles are GTP-bound. Cloning strategies for general strain construction and Ypt7 ubiquitination mutants Standard PCR-based gene deletion and modification cassettes were used to disrupt genes and tag genes (described in Yeast 14, 953-961, 1998). We replaced the endogenous YPT7 gene with a modified ypt7 gene (which was tagged and either with or without ubiquitination site mutations); the gene’s expression is controlled by a GAL promoter. To replace the endogenous YPT7 gene, we used a replacement cassette which had been cloned into the pCR2.1 plasmid vector (Invitrogen). This cassette consist of four adjacent sequences: (1) a stretch of the upstream YPT7 promoter sequence (2) a KAN marker cassette (3) a PGAL promoter that regulates a 3HA-tagged ypt7 ORF (4) a stretch of sequence that is downstream from the YPT7 ORF. We sequenced the ypt7 sequence of this replacement cassette to make sure the sequence was correct. We performed mutagenesis of the HA-tagged ypt7 gene on this plasmid-based cloning cassette using Quickchange (Stratagene). Every time we made a point mutation, we sequenced the plasmid-based ypt7 gene to confirm that the particular mutation had occurred, and no other mutations were introduced. We subjected a cassette that carried the required ypt7 4 point mutations to PCR, and the PCR DNA was transformed into yeast. The flanking YPT7 gene sequences resulted in replacement of the endogenous YPT7 gene with the PGAL-driven HA-tagged wild-type or mutant version. We performed colony PCR in order to screen for both absence of the endogenous wild-type, and presence of the replacement HA-tagged ypt7 gene. Next, DNA obtained by colony PCR of the newly introduced, integrated ypt7 gene was again sequenced to ensure correct introduction of the desired mutations and the absence of mutations resulting from the gene-swap procedure. Immunoprecipitation of HA-tagged and untagged Ypt7/Rho1 Equilibrium-flotated vacuoles were diluted in 0% Ficoll buffer (10 mM PIPES-KOH [pH 6.8], 200 mM sorbitol) and collected by centrifugation. The pelleted vacuoles were lysed in NP-40 lysis buffer (50 mM Tris-HCl [pH 8], 0.5% NP-40, 150 mM NaCl, 5 mM MgCl2) containing the COMPLETE protease inhibitor cocktail (1 tablet/10 ml; Roche). In order to immunoprecipitate HA-tagged proteins, HA affinity matrix (immobilized clone 3F10 rat monoclonal antibody; Roche) was added (50 l per ml lysate). After incubating the mixture at 4oC for one hour while it was rotating, the beads were pelleted by centrifugation and then washed four times in the lysis buffer; the washing process included a change of tubes. We analyzed some immunoprecipitates by mass spectrometry and some by SDS/PAGE (10 to 12.5% acrylamide). We captured untagged Ypt7 protein by adding 2 l of an -Ypt7 rabbit serum to one ml of vacuolar lysate; in the control, an unrelated serum was added to the vacuolar lysate. After we incubated the lysate at 4 oC for an hour, we added 50 l of protein-A agarose beads (Invitrogen). Subsequently, we incubated the lysate for another 30 min at 4 oC. In the experiments that yielded the data represented in Figure 3C and 3D, lysates were first subjected to immunoprecipitation 5 using an unrelatedcontrol serum. Then, the cleared lysate was subjected to the-Ypt7 serum immunoprecipitation. Sequential precipitation, first with the control serum and then with the -Ypt7 serum, resulted in a cleaner precipitate. This is because the first immunoprecipitation with the control serum reduced the amount of background material in the subsequent immunoprecipitation of Ypt7. 6