Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Basal metabolic rate wikipedia , lookup
Proteolysis wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Biosynthesis wikipedia , lookup
Lactate dehydrogenase wikipedia , lookup
Citric acid cycle wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Phosphorylation wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
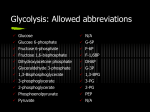
Liver glycogen constitutes a reserve of glucose for the body as a whole. The reserve is not large. In fact, the reserve is virtually exhausted in 24 hours. In such an animal the continuing requirement for glucose is satisfied by gluconeogenesis, which is the synthesis of glucose from non-carbohydrate precursors: lactate, propionate, glycerol, pyruvate, gluconeogenic amino acids. The contribution of gluconeogensis to meeting glucose requirements clearly depends on the amount of carbohydrates absorbed from the intestine, which in many species is not the same as the amount of carbohydrates ingested. Herbivores have a diet that is rich in cellulose, but microbial action in the intestine converts this carbohydrate to acetate, butyrate and propionate, of which only the last can yield glucose by gluconeogenesis. In general, only root and seed eaters, which include man, may have a diet in which a large amount of carbohydrates is both ingested and absorbed. Carnivores must derive a large part of their essential glucose from the gluconeogenic amino acids. The constitution of the diet affects not only the rate of gluconeogenesis but also the total capacity for gluconeogenesis. The amounts of certain key enzymes in the gluconeogenesis pathway vary inversely with the amount of carbohydrates in the diet. These enzymes include those which distinguish from the reversal of glucolysis and are namely: pyruvate carboxylase, glucose-6-pohsphatase, and fructose-1-6-biphosphatase and phospohenolpyruvate carboxykinase. The secretion of insulin and cortisol depends directly or indirectly on the concentration of blood glucose. The concentration of these hormones in turn influences the synthesis of some of these enzymes, and hence the gluconeogenesis. In this practical the effect of the previous diet on the gluconeogenic capacity of an organ will be investigates. Only the liver and kidney are capable of gluconeogenesis and the greater size of the liver makes its contribution predominant. However, it is technically more convenient to investigate the gluconeogenic capacity in the kidney. The kidney has no store of glycogen whose breakdown might obscure the synthesis of glucose from noncarbohydrate precursors, or into which the newly synthesized glucose might be incorporated. Because the body’s metabolism can change rapidly, killing must be quick and avoid chemicals, which affect it. One method is to stun the rat and break its neck. The experiment is illustrative and to obtain reliable data in a practical of this nature would require a number of rats starved and fed rats. The technique will be the same for all experiments, but the treatment of the animals (rats) and the nature of the precursor will be varied. The experiment will study the effect on the rate of gluconeogensis in starvation and the nature of the gluconeogenic precursor. We will determine the rate of gluconeogenesis from five precursors by kidney slices from rats which have been on a normal digest or starved for 24 hours. The incubation media includes one of the five precursors: lactate, alanine, propionate, acetate or glycerol, in 10mM of Krebs bicarbonate medium. Nail 118.5mM NaHCO3 28.9mM KCl 4.7mM KH2PO4 1.2mM CACl2·6H2O 2.5mM MgSO4·7H2O 1.2mM Flasks containing 5 ml of precursor solution in Krebs bicarbonate medium, and control flasks containing only 5ml Krebs are preincubated in a shaking water bath at 37º while gassing with 95% oxygen and 5% CO2 gas mixture. The kidney from the starved rat is placed in a Perspex holder and sliced by drawing a razor blade through the holder. The slices are then washed, and bottled in the flasks containing substrates. The process is repeated for kidney slices from the fed rat. After the stoppers have been placed, the flasks are gassed with the gas mixture continuously during incubation. The importance of oxygen is to allow cellular respiration, oxidative phosphorylation and ATP synthesis; the importance of CO2 is to stabilize the equilibrium of the bicarbonate buffer and supply CO2 for carboxylase reactions (e.g. Pyruvate carboxylase). The slices are incubated at 37º for 30 minutes with shaking. At the end of the incubation period, the slices are removed, blotted gently and weighed. Thin slices and efficient shaking are important features of this experiment because it’s important to provide efficient access of substrates to metabolizing tissue. Tissue weights: substrate propionate lactate alanine acetate control control fed (g) starved (g) 0.083 0.1 0.088 0.1 0.093 0.12 0.099 0.08 0.1 0.092 0.093 0.1 The concentration of glucose in the residual incubation medium must now be estimated. Calibration samples will also be required. Glucose oxydase reagent (Perid) and standard glucose solutions are used to determine a graph (0.05mM, 0.1mM, 0.15mM). The glucose reagent is preferable to chemical methods for general detection of reducing sugars since it is enzyme based, therefore specific for glucose, even if other reducing sugars are present. 5 ml glucose reagent is added to 1 ml from each incubation flask. 5 ml glucose reagent is added to 1 ml of each standard glucose solution and one reagent blank (1 ml water) total – 16 test tubes. The tubes are shaken to mix the contents and are placed in a water bath at 37º for 15 minutes. Mixing is important for assays to achieve homogeneous solutions. The glucose analysis is based on the following reactions: Hydrogen peroxide is formed when glucose is oxidized to gluconolactone and the hydrogen peroxide in turn oxidizes the chomagen ABTS (diammunium 2,2’-azino-bis(ethylbenzothiazoline-6-sulphonate)) in the presence of peroxydase to produce a green color. Next color absorbance will be measured (610 nm) and glucose content of the samples will be calculated. glucose concentration amount of glucose Absorbance in standard solution in 1 ml (610 nm) 0 0 0.04 0.05 0.05 0.16 0.1 0.1 0.28 0.15 0.15 0.41 Equation (regression) Calculate the rate of glucose formation as μmol/g tissue/hour for the incubated slices from the starved rats. You will need to: Use the standard plot Calculate the micromols of glucose in each of the 5 ml flasks Allow for incubation time (30 minutes) Allow for tissue weight. fed propionate lactate alanine acetate control control g absorbance μmol/g tissue/hour 0.083 0.09 2.55 0.088 0.117 3.65 0.093 0.153 5.03 0.099 0.124 3.53 0.1 0.078 1.63 0.093 0.076 1.66 starved g absorbance μmol/g tissue/hour propionate 0.1 0.17 5.36 lactate 0.1 0.27 9.43 alanine 0.12 0.4 12.26 acetate 0.08 0.25 10.77 control 0.092 0.087 2.22 control 0.1 0.092 2.25 The diagrams indicate a range of values obtained from 10 rats. How do your result compare with these? How would you extend this investigation? An interpretation of experimental results must be based upon a realistic estimation of the potential errors, for example: The kidney samples may dry out if cutting and selection is too slow. Technique in carrying out procedures The kidney slices will vary in thickness and composition. We have corrected for variations in weight, should we have corrected for surface area or variations in proportions of cortex? How could we do this? The rate of gluconeogenesis is controlled by a number of key enzymes which catalyze the ‘essentially irreversible’ reactions. It is at these stages that the catabolic pathway of glucolysis and the biosynthetic pathway of gluconeogensis differ. The activity of the enzymes controlling these reactions should be higher in the rats starved for 24 hours. Do your results agree with this? –yes, gluconeogenesis is more evident in the starved rats. propionate lactate fed 2.55 3.65 starved 5.36 9.43 alanine acetate control control 5.03 3.53 1.63 1.66 12.26 10.77 2.22 2.25 The gluconeogensis is produced only from certain precursors – for example, acetate does not serve as precursor since it has similar gluconeogenic rate as the control group, while other precursors have a much higher gluconeogenic rate. In ruminant animals it is more likely that the gluconeogenesis will be more efficient with propionate, while in carnivores gluconeogenesis will be more efficient gluconeogenic amino acids.