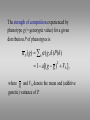Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Koinophilia wikipedia , lookup
Viral phylodynamics wikipedia , lookup
Genome (book) wikipedia , lookup
Public health genomics wikipedia , lookup
Designer baby wikipedia , lookup
Medical genetics wikipedia , lookup
History of genetic engineering wikipedia , lookup
Genetic testing wikipedia , lookup
Gene expression programming wikipedia , lookup
Dual inheritance theory wikipedia , lookup
Behavioural genetics wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Heritability of IQ wikipedia , lookup
Human genetic variation wikipedia , lookup
Genetic drift wikipedia , lookup
Quantitative trait locus wikipedia , lookup
Natural selection wikipedia , lookup
Microevolution wikipedia , lookup
Polymorphism (biology) wikipedia , lookup
Sasha Gimelfarb died on May 11, 2004 A Multilocus Analysis of Frequency-Dependent Selection on a Quantitative Trait Reinhard Bürger Department of Mathematics, University of Vienna Frequency-dependent selection has been invoked in the explanation of evolutionary phenomena such as • • • • Evolution of behavioral traits Maintenance of high levels of genetic variation Ecological character divergence Reproductive isolation and sympatric speciation Frequency-Dependent Selection Caused by Intraspecific Competition Intraspecific competition mediated by a quantitative trait under stabilizing selection: • • • • • • • • Bulmer (1974, 1980) Slatkin (1979, 1984) Christiansen & Loeschcke (1980), Loeschcke & Christiansen (1984) Clarke et al (1988), Mani et al (1990) Doebeli (1996), Dieckmann & Doebeli (1999) Matessi, Gimelfarb, & Gavrilets (2001) Bolnick & Doebeli (2003) Bürger (2002a,b), Bürger & Gimelfarb (2004) The General Model • A randomly mating, diploid population with discrete generations and equivalent sexes is considered. • Its size is large enough that random genetic drift can be ignored. • • Viability selection acts on a quantitative trait. Environmental effects are ignored (in particular, GxE interaction). Therefore, the genotypic value can be identified with the phenotype. Ecological Assumptions Fitness is determined by two components: • by stabilizing selection on a quantitative trait, S ( g ) 1 s( g ) 2 , and • by competition among individuals of similar trait value, ( g , h) 1 a( g h) . 2 The strength of competition experienced by phenotype g (= genotypic value) for a given distribution P of phenotypes is P ( g ) h ( g , h ) P ( h ) 1 a[( g g ) VA ], 2 where g and VA denote the mean and (additive genetic) variance of P. If stabilizing selection acts independently of competition, the fitness of an individual with phenotype g can be written as W ( g ) F ( N P ( g )) S ( g ), where F(N) describes population growth according to N´=NF(N). (F may be as in the discrete logistic, the Ricker, the Beverton-Holt, the Hassell, or the Maynard Smith model.) For weak selection (O( s 2 ) 0 , f = a/s constant), this fitness function is approximated by W ( g ) F ( N )(1 s( g ) 2 s ( N )[( g g ) 2 VA ]), where (N ) is a compound measure for the strength of frequency and density dependence relative to '( N ) stabilizing selection, i.e., ( N ) NF f . F(N ) Genetic Assumptions • The trait is determined by additive contributions from n diploid loci, i.e., there is neither dominance nor epistasis. • At each locus there are two alleles. The allelic effects at locus i are -gi/2 and gi/2. (This is general because the scaling constants can be absorbed by the position of the optimum and the strength of selection.) Genetical and Ecological Dynamics pi , pi´ : frequencies of gamete i in consecutive generations Wjk : fitness of zygote consisting of gametes j, k R(jk->i): probability that a jk-individual produces a gamete of type i through recombination W : mean fitness pi ' W 1 W jk p j pk R( jk i ) j ,k N ' NW Issues and problems to be addressed • What aspects of genetics and what aspects of ecology are relevant, and under what conditions? • When does FDS have important consequences for the genetic structure of a population? • • How does FDS affect the genetic structure? How much genetic variation is maintained by this kind of FDS? Numerical Results from a Statistical Approach (with A. Gimelfarb) Figure, poly, th=0 4.0 n = 2, = 0 n = 3, = 0 n = 4, = 0 3.5 Polymorphism 3.0 2.5 2.0 1.5 1.0 0.5 0.0 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 Strength of frequency dependence, f 2.2 2.4 Figure, poly, th=0, 0.75 4.0 n = 2, = 0 n = 3, = 0 n = 4, = 0 n = 2, = 0.25 n = 3, = 0.25 n = 4, = 0.25 3.5 Polymorphism 3.0 2.5 2.0 1.5 1.0 0.5 0.0 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 Strength of frequency dependence, f 2.2 2.4 4.0 Figure: poly n = 2, r = 0.5 n = 3, r = 0.5 n = 4, r = 0.5 n = 2, 0 < r < 0.01 n = 3, 0 < r < 0.01 n = 4, 0 < r < 0.01 3.5 Polymorphism 3.0 2.5 2.0 1.5 1.0 0.5 0.0 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 Strength of frequency dependence, f Selection: Quad. App.: Stabilizing Complicated Disruptive 1.8 The Weak-Selection or Linkage-Equilibrium Approximation • • • • The structure is the same as in Turelli and Barton 2004 (but vi ). The proofs of the results below use their results. The population is assumed to be in demographic equilibrium, i.e., N and η are treated as constants. All models of intraspecific competition and stabilizing selection I know have the above differential equation as their weak-selection approximation. ‘Arbitrary‘ population regulation, i.e., with a unique stable carrying capacity, is admitted. General Conclusions • The amount and properties of variation maintained depend in a nearly threshold-like way on , the strength of frequency and density dependence relative to stabilizing selection. • This critical value is independent of the number of loci and, apparently, also of the linkage map. Weak FDS • If more than two loci contribute to the trait, then weak frequency dependence ( < 1) can maintain significantly more genetic variance than pure stabilizing selection, but still not much. The more loci, the larger this effect. • FDS of such strength does not induce a qualitative change in the equilibrium structure relative to pure stabilizing selection. • Such FDS does not lead to disruptive selection, rather, stabilizing selection prevails. Strong FDS • Strong FDS ( > 1) causes a qualitative change in the genetic structure of a population by inducing highly polymorphic equilibria in positive linkage disequilibrium. • In parallel, such FDS induces strong disruptive selection, the fitness differences between phenotypes maintained in the population being much larger than under pure stabilizing selection. Disruptive Selection • Therefore, disruptive selection should be easy to detect empirically whenever FDS is strong enough to affect the equilibrium structure qualitatively. • Its strength (the curvature of the fitness function at equilibrium) is s( – 1). When Genetics Matters • • • The degree of polymorphism maintained by strong FDS depends on the number of loci and the distribution of their effects. Models based on popular symmetry assumptions, such as equal locus effects or symmetric selection, are often not representative (they maintain more polymorphism). Linkage becomes important only if tight. It produces clustering of the phenotypic distribution. Otherwise, the LE-approximation does a very good job. Outlook • Include assortative mating to study the conditions leading to divergence within a population (work in progress K. Schneider). • Determine the conditions under which sympatric speciation is induced by intraspecific competition.