Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Nucleic acid analogue wikipedia , lookup
Peptide synthesis wikipedia , lookup
Lipid signaling wikipedia , lookup
Zinc finger nuclease wikipedia , lookup
Metalloprotein wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Butyric acid wikipedia , lookup
Biosynthesis wikipedia , lookup
15-Hydroxyeicosatetraenoic acid wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Specialized pro-resolving mediators wikipedia , lookup
Amino acid synthesis wikipedia , lookup
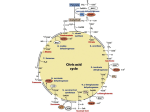
Journal of General Microbiology (I 977), gg, 43-48 Printed in Great Britain 43 Effect of Zinc on Tricarboxylic Acid Cycle Intermediates and Enzymes in Relation to Matoxin Biosynthesis By S . K. GUPTA, K. K. M A G G O N AND T. A. V E N K I T A S U B R A M A N I A N Depdrtment of Biochemistry, Vdllabhbhai Patel Chest Institute, University of Delhi, Delhi I 10007, India (Received 7 September I 976 ; revised I 8 October 1976) SUMMARY The stimulatory action of zinc on aflatoxin production by Aspergilluspdrdsiticus investigated by studying the levels of tricarboxylic acid (TCA) cycle intermediates and related enzymes in the fungal mycelium. During the stationary phase of growth, the levels of a-keto acids declined in zinc-sufficient cultures compared with those in zinc-deficient cultures. TCA cycle enzymes did not show any significant changes due to zinc availability. In zinc-deficient cultures, enzymes of the TCA cycle had maximum activity on the fourth day, after which their activity declined. In zinc-sufficient cultures, some enzymes showed maximum activity on the fourth day, others on the second day. N R R L ~ ~has ~ Obeen INTRODUCTION Several reports have described the stimulatory action of zinc on aflatoxin biosynthesis in Aspergillus PdrdSitiCW and A . $dVW (Mateles & Adye, 1965; Lee, Townsley & Walden, 1966; Gupta & Venkitasubramanian, 1975;Maggon, Gopal & Venkitasubramanian, 1g73), but the mechanism of this stimulatory action is not understood at present. The effect of zinc on the metabolism of A. pdrdsiticus has not been reported. It has been shown that soybean is a poor substrate for aflatoxin production because of its low content of zinc which.is bound to phytic acid (Gupta & Venkitasubramanian, 1975). Acetyl-CoA is a precursor for aflatoxin biosynthesis and also enters the tricarboxylic acid (TCA) cycle by condensing with oxaloacetate to form citrate. The study of TCA cycle intermediates and enzymes during zinc deficiency may reveal the mechanism of action of zinc in aflatoxin biosynthesis. We have therefore investigated the levels of TCA cycle enzymes and intermediates of A . parasiticus grown in zinc-deficient medium. METHODS Organism. Aspergilluspdrusiticus N R R L ~ ~ (toxigenic) ~ O obtained from Northern Regional Research Laboratory, Peoria, Illinois, U.S.A., was used throughout and was maintained as soil cultures (Hesseltine, Bradle & Benjamin, I 960). Medium dnd growth conditions. Sucrose/low salt (SLS) synthetic medium contained (per litre double-distilled water) :sucrose, 85 g; asparagine, 10 g; (NH,),SO,, 3.5 g; KH2P04, 10 g; MgSO,. 7H20,2 g; CaC1,.2H20,75 mg; ZnSO,. 7H20,10mg; MnCl,, 5 mg; ammonium molybdate, 2 mg; Na2B40,, 2 mg; and FeS0,.7H20, 2 mg. The medium was adjusted to pH 4.5 with HCl. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26 S. K. GUPTA AND OTHERS 44 A spore suspension in sterile double-distilled water was prepared from 5- to 6-day-old cultures of A . parasiticus grown on medium containing 2 :h (w/v) glucose, 2 yo(w/v) agar and I % (v/v) peptone. Flasks containing 100 ml SLS medium were inoculated with 3 x 106 to 3-4 x 1oSspores, and incubated without shaking at 26 5 I "C. After various periods of growth, mycelium was harvested either for determination of dry weight and aflatoxin content, or for the estimation of TCA cycle intermediates and enzymes. All operations were done below 4 "C. Extraction and estimation of qfiatoxins. Aflatoxins were extracted from the medium and mycelium with chloroform and separated by t.1.c. using the solvent system toluene/isoamyl alcohol/methanol (go :32 :3, by vol.) (Reddy, Viswanathan & Venkitasubramanian, 1970). Individual bands were eluted with methanol and estimated spectrophotometrically using the extinction coefficients reported by Nabney & Nesbitt (1965). Extraction and estimation of TCA cycle intermediates. Intermediates of the TCA cycle were extracted from mycelium as described by Williamson & Corkey (1973). Mycelium was quickly separated from the' medium by decanting and immediately frozen by pressing between plates of aluminium foil precooled in liquid nitrogen. Frozen mycelium was stored at -20 "C until used. The mycelium (500 mg) was ground with acid-washed sea sand in a precooled mortar (4 "C) containing 5 ml 8 % (w/v) perchloric acid in 40% (v/v) ethanol. The homogenate was centrifuged a t 25 ooo g for I o min, and the supernatant was decanted and stored at o to 4 "C. The residue was again extracted with 2 to 5 ml 6 % perchloric acid and centrifuged, and the supernatant was combined with the first extract. The combined extracts were brought to pH 6.0to 7.0 by slowly adding a solution of 3 M-K,CO, containing 0.5 M-triethanolamine or 0.5 M-KOH, and then centrifuged to remove precipitated KC10,. a-Keto acids, citric acid and malic acid were estimated in this supernatant. a-Keto acids, acetaldehyde, pyruvic acid, a-ketoglutaric acid and oxaloacetate, were estimated by preparing their phenylhydrazine derivatives and separating them by t.1.c. using the modified method of Friedemann & Haugen (I 943) : yellow phenylhydrazones were scraped from the plates and estimated quantitatively by the method of El Hawary & Thompson (1953). Citric acid was estimated by the method of Ettinger, Goldbaum & Smith (1952) and malic acid by the fluorometric method of Hummel (1949). Extraction and assay of TCA cycle enzymes. Mycelium was decanted from the medium after various periods of growth and washed with cold 0.05 M-Tris/HCl buffer, pH 7.3. Washed mycelium was ground with acid-washed sea sand in the same buffer in a precooled mortar and centrifuged at 15000 g for 10min. The supernatant was used as the source of enzymes. Malate dehydrogenase (EC. I . I . I .37) was assayed by the procedure of Yoshida (1973); malatedehydrogenase (decarboxylating; EC. I . I .I .40) and isocitrate dehydrogenase (NADP+,EC. I . I .I .42) accordingtoOchoa( I 955) ;succinate dehydrogenase (EC. I . 3 .99 . 1 ) by the method of Kalra, Krishna Murti & Brodie (1971); pyruvate dehydrogenase (EC. I . 2 . 4 . I .) and a-ketoglutarate dehydrogenase (EC. I . 2 . 4 . 2 ) as described by Stumpf, Zarudnaya & Green (1947). The activities reported are the means of six separate experiments. RESULTS AND DISCUSSION Preliminary observations indicated there was no fungal growth or aflatoxin production in the absence of zinc. To investigate the role of zinc in the metabolism of A . pdrdsiticus and the mechanism of its stimulatory action on aflatoxin production, two concentrations of zinc were used: 10 mg 1-1 which was optimum for aflatoxin production, and 0.25 mg 1-1 which was the minimum required for fungal growth. The growth of A . pdrdSitiCUS and Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26 45 TCA cycle in a$atoxin biosynthesis Table I.Eflect of zinc on growth and @atoxin synthesis by A . PardSitiCUS The results for mycelial dry wt and total aflatoxin are expressed as g (100 ml medium)-l and mg (IOO ml medium)-' respectively. Zinc-deficient SLS medium SLS medium h I f 7 Days of growth Mycelial dry wt Total aflatoxin Mycelial dry wt Total aflatoxin I -07 I '04 2-04 2 0.68 5-70 3'24 15-61 4 2.66 6.97 4-06 21-59 . 6 3'36 6-05 4'74 27'94 8 4'45 h \ Table 2. Eflect of zinc on the content of TCA cycle intermediates in A . parasiticus All results are expressed as pmol (g dry wt mycelium)-'. Days of growth Acetaldehyde Zinc-deficient SLS medium 2 3'63 64'59 4 6 31-60 8 17-19 SLS meQium 2 4 6 8 57'67 29'55 24'05 19-55 Pyruvic acid 3'76 5'38 2-57 1-81 4'17 3-05 2-51 1-61 a-KetoOxaloacetate glutaric acid 2.69 4'34 2.86 I -26 I '92 3-41 2-39 1-19 Citric acid Malic acid 0.65 0.57 0.17 0.42 0.39 0.96 5-89 3'55 6.39 6.3I 1-01 0.05 0.82 0'12 6-02 3'36 4'67 0.68 1'22 0.87 0.53 0.25 0.62 3-12 aflatoxin production were both inhibited by zinc deficiency, but aflatoxin synthesis more so than growth (Table I). This agrees with the reports of previous workers (Lee et al., 1966; Maggon et d.,1973). The effect of zinc on pyruvic acid and TCA cycle intermediates is shown in Table 2. In zinc-deficient medium, the levels of a-keto acids reached a maximum on the fourth day of growth. In zinc-sufficient medium, the levels of acetaldehyde, pyruvate and a-ketoglutarate were maximal on day 2, during the late-exponential phase of growth (Detroy & Hesseltine, 1g70),and oxaloacetate was maximal on day 4. Thus, the zinc-sufficient cultures had higher levels of these intermediates during growth and lower levels in the stationary phase than did the zinc-deficient cultures. The only exceptions were oxaloacetate and citrate which showed lower levels in zinc-sufficient medium. In the stationary phase, A. pdrdsiticus had lower levels of a-keto acids in zinc-sufficient medium. The build up of TCA cycle intermediates in zinc-sufficient cultures at the end of the exponential growth phase may play a role in triggering the onset of aflatoxin synthesis. Bu'Lock (I 9-75), Weinberg (I 971)and Demain (I 972)have postulated that the accumulation of primary precursors at the end of exponential growth is necessary for the initiation of secondary metabolism. The results in Table 2 seem to provide evidence for this. The decrease in TCA cycle intermediates in zinc-sufficient cultures during the stationary phase (days 4 to 8) indicates the diversion and possible utilization of these intermediates into secondary biosynthesis, particularly that of aflatoxin. During the stationary phase in zincdeficient cultures, TCA cycle intermediates accumulated and there was reduced aflatoxin formation. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26 46 S. K. G U P T A A N D OTHERS Table 3. Eflect of zinc on the speciJic activities of TCA cycle enzymes of A. parasiticus Specific activities [units (mg protein)-'] A f Days of growth Pyruvate dehydrogenase Isocitrate dehydrogenase a-Ketoglutarate dehydrogenase Succinate dehydrogenase Malate dehydrogenase \ Malate dehydrogenase (decarboxylating) Zinc-deficient SLS medium 2 4 6 8 ' SLS medium 2 4 6 8 I -80 7'14 2.3 I I -70 4'74 I *g8 3-04 I -96 3'25 I -65 2-48 3'56 2-80 2-97 I '00 2.00 3-14 1-55 2.14 3.20 3-70 1.38 I -03 1 '55 2-32 3'97 3-80 5.00 1.52 2-57 I -05 1.38 63-30 100~00 66-60 2 I -30 242'50 67'44 57'94 47'04 9-60 7.10 5'70 5'1 5 8.60 5-60 6-00 I 0.50 In zinc-deficient cultures these precursors are not used to form secondary metabolites at the same rate as in zinc-sufficient cultures. There have been many reports of the accumulation of citric and malic acids during zinc deficiency (Foster, 1949; Trumpy & Mills, 1963; Shu & Johnson, 1948; Martin, 1955; Sanchez-Marroquin, Carreno & Ledezma, 1970; Lockwood, 1975). Foster (1949) suggested that zinc plays a part in the utilization of carbohydrate by fungi because, under conditions of partial zinc deficiency, less carbohydrate is completely oxidized and a greater proportion is diverted to form organic acids. Foster & Waksman (1939) concluded that in RJzizopus nigricans zinc has a specific role in carbohydrate metabolism : they reported accumulation of fumaric acid during zinc deficiency. Romano, Bright & Scott (1967) observed that addition of zinc increased cell synthesis with a more efficient utilization of glucose and a corresponding decrease in fumaric acid yields. Chester & Rolinson (195I) reported that zinc-deficient cultures of A . niger produced large quantities of citric acid. The results of this study are supported by the findings of Bryzgalova & Orlova ( I975) who concluded that conditions which favour the synthesis of secondary metabolites decrease the accumulation of organic acids and vice versa. The activities of the enzymes of the TCA cycle of A . parasiticus in zinc-deficient media reached a maximum on the fourth day of growth but declined as the cultures aged (Table 3). In zinc-sufficient medium the activities of pyruvate dehydrogenase, isocitrate dehydrogenase and a-ketoglutarate dehydrogenase reached a maximum on the fourth day of growth; those of succinate dehydrogenase, malate dehydrogenase and malate dehydrogenase (decarboxylating) remained low throughout. The activities of other enzymes were high in zincsufficient cultures during the exponential growth phase, but variable during the stationary phase. Bu'Lock et ul. (1965) observed a marked decrease in the activity of the TCA cycle at the end of the exponential growth phase in Penicillium urticae. A decline in TCA cycle enzyme activity can lead to accumulation of acetyl-CoA, glycolytic intermediates and citrate (Bu'Lock, 1965, 1975; Demain, 1968). Similar results have been obtained in this study (Table 2). The activation of acetyl-CoA carboxylase by citrate and a-glycerophosphate would result in the formation of more malonyl-CoA (Resmussen & Klein, 1960). Increased formation of malonyl-CoA would lead to enhanced aflatoxin formation as observed in zincsufficient cultures. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26 TCA cycle in aflatoxin biosynthesis 47 Part of this work was supported by a PL-480 grant no. FG-111-438.S. K. G. is grateful to Dr D. N. Gupta, University College of Medical Science, New Delhi, for his keen interest and constant encouragement. K. K. M. is grateful to the Council of Scientific and Industrial Research, New Delhi, for placement in the Scientists Pool. REFERENCES BRYZGALOVA, T. E. & ORLOVA, N. V. (1975). Organic acid production by an active strain of Actinomyces rimosus and an inactive mutant in oxytetracycline biosynthesis. Antibiotiki 20, I 1-1 5. BU’LOCK,J. D. (1965). Aspects of secondary metabolism in fungi. In Biogenesis of Antibiotic Substances, pp. 61-72. Edited by Z. Vanek and Z. Hostalek. Prague: Publication House(CzechAcademyof Sciences). BU’LOCK, J. D. (1975). Secondary metabolism in fungi and its relationships to growth and development. In The Filamentous Fungi, vol. I, Industrial Mycology, pp. 33-58. Edited by J. E. Smith and D. R. Berry. London: Edward Arnold. BU’LOCK, J. D., HAMILTO~, D., HULME, M. A., POWELL, A. J., SMALLEY, H. M., SHEPHARD, D. & SMITH, G. N. (1965). Metabolic developments and secondary biosynthesis in Penicillium urticae. Canadian Journal of Microbiology 11, 765-778. CHESTER, C. G. C. & ROLINSON, G. N. (1951). Zinc in the metabolism of a strain of Aspergillus niger. Journal of General Microbiology 5, 553-565. DEMAIN,A. L. (1968). Regulatory mechanisms and the industrial production of microbial metabolites. Lloydia 31, 395-418. DEMAIN, A. L. (1972). Cellular and environmental factors affecting the synthesis and excretion of metabolites. Journal of Applied Chemical Biotechnology 22, 345-372. DETROY, R. W. & HESSELTINE, C. W. (1970). Secondary biosynthesis of aflatoxin B, in Aspergillusparasiticus. Canadian Journal of Microbiology 16,959-963. ELHAWARY, M. F. S. &THOMPSON, R. H. S. (1953).Separation and estimation of blood keto acids by paper chromatography. Biochemical Journal 53, 340-347, ETTINGER,R. H., GOLDBAUM, L. R. & SMITH,L. H. (1952). A simplified photometric method for the determination of citric acid in biological fluids. Journal sf Biological Chemistry 199,53 1-536. FOSTER, J. W. (1949). Chemical Activities of Fungi. New York: Academic Press. FOSTER, J. W. & WAKSMAN, S. A. (1939). The specific effect of zinc and other heavy metals on the growth and nutrition of Rhizopus. Journal of Bacteriology 37,599-617. FRIEDEMANN, T. E. & HAUGEN, G. E. (1943). Pyruvic acid. 11. The determination of keto acids in blood and urine. Journal of Biological Chemistry 147,415-442. GUPTA,S. K. & VENKITASUBRAMANIAN, T. A. (1975). Production of aflatoxin on soybeans. Applied Microbiology 29, 834-836. HESSELTINE, C. W., BRADLE, B. J. & BENJAMIN, C. R. (1960). Further investigations on the preservation of molds. Mycologia 52, 762-774. HUMMEL, J. P. (1949). Fluorometric determination of malic acid. Journal of Biological Chemistry 180, I 225-1 228. KALRA,V. K., KRISHNAMURTI,C. R. & ERODIE, A. F. (1971). Resolution and reconstitution of the succinoxidase pathway of Mycobacterium phlei. Archives of Biochemistry and Biophysics 147,734-743. LEE,E. G. H., TOWNSLEY, P. M. & WALDEN, C. C. (1966). Effect of bivalent metals on the production of a t o x i n in submerged cultures. Journal of Food Science 31,432-436. LOCKWOOD, L. B. (1975). Organic acid production. In The Filamentous Fungi, vol. I , Industrial Mycology, pp. 140-157. Edited by J. E. Smith and D. R. Berry. London: Edward Arnold. MAGGON, K. K., GOPAL,S. & VENKITASUBRAMANIAN, T. A. (1973). Effect of trace metals on aflatoxin production by Aspergillus &vus. Biochemische Physiologie der PJIanzen 164, 523-530. MARTIN, S. M. (1955). Effect of ferrocyanide on growth and citric acid production by Aspergillus nker. Canadian Journal of Microbiology I, 644-652. MATELES, R. I. & ADYE,J. C. (1965). Production of aflatoxin in submerged cultures. Applied Microbiology 13, 208-21 I . NABNEY, J. & NESBITT, B. F. (1965). A spectrophotometric method for determining the aflatoxins. Analyst 90,155--J60. OCHOA, S . (1955). I. Isocitrate dehydrogenase system (TPN) from pig heart. 11. Malic dehydrogenase from pig heart. Methods in Enzymology I, 699-704 and 735-739. REDDY,T. V., VISWANATHAN, L. & VENKITASUBRAMANIAN, T. A. (1970). Thin-layer chromatography of aflatoxins. Analytical Biochemistry 38, 568-57 I . RESMUSSEN, R. K. & KLEIN,H. P. (1968). Activation of fatty acid synthesis in cell free extracts of Saccharomyces cerevisiae. Journal of Bacteriology 95, I 57-161. M I C 99 4 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26 48 S. K. GUPTA AND OTHERS ROMANO, A. H., BRIGHT,M. M. & SCOTT,W. E. (1967). Mechanism of fumaric acid accumulation in Rhizopus nigricans. Journal of Bacteriology 93, 600-604. N. (1970). Effect of trace elements on citric acid ferSANCHEZ-MARROQUIN, A., CARRENO, R. & LEDEZMA, mentation by Aspergillus niger. Applied Microbiology 20, 888-892. SHU,P. &JOHNSON, M. L. (1948). The interdependence of medium constitutents in citric acid production by submerged fermentation. Journal of Bacteriology 50, 577-585. STUMPF, P. K., ZARUDNAYA, K. & GREEN,D. E. (1947). Pyruvic acid and a-ketoglutaric oxidase of animal tissue. Journal of Biological Chemistry 167,817-825. TRUMPY, B. M. & MILLS, N. F. (1963). Nutritional requirements of an Aspergillus niger mutant for citric acid production. Journal of General Microbiology 30, 38 1-393. WEINBERG, E. D. (1971). Secondary metabolism. Raison d'Ctre. Perspectives in Biology and Medicine 14, 565-577. WILLIAMSON, J. R. & CORKEY, B. E. (1973). Assays of intermediates of the citric acid cycle and related compounds by fluorometric enzyme methods. Methods in Enzymology 13,434-443. A. (1973). L-Malate dehydrogenase from B. subtilis. Methods in Enzymology 13, 141-145. YOSHIDA, Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 06 May 2017 04:26:26