Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Vaccination wikipedia , lookup
Anti-nuclear antibody wikipedia , lookup
DNA vaccination wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Molecular mimicry wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Immunocontraception wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Diagnosis of HIV/AIDS wikipedia , lookup
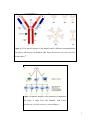
Application of Biophysical techniques in AIDS Vaccine research: Amit Anand MOAC, University of Warwick [email protected] 19th January 2005 ABSTRACT At the moment the number of people dying because of human immuno-deficiency virus (HIV) in a year is more than those dying due to natural calamities like floods, earth quake or volcano eruption. Virology researchers are not only collaborating internationally but they are also joining hands with other disciplines like physical sciences to solve the problem. Although the initial immune reaction against any new deadly virus will be insignificant but it will definitely be present. Some rare but potent antibodies of human origin which can neutralize HIV are been identified and their interactions are been visualized at atomic level. Examples are b12, 2G12, 2F5, 4E10 and Z13 (9). With the combination of multidisciplinary approach, biophysical techniques are opening new horizons to explore every possibility to solve the biggest challenge to the survival of human race on earth. This article discusses some of the successes researchers have achieved with the help of biophysical techniques. KEYWORDS: HIV Biophysical technique, X-ray crystallography, NMR, Mass Spectrometry 1 INTRODUCTION: The success of the polio vaccine is attributed to the work of a great scientist Jonas Salk (19141995). Salk launched the first polio vaccine in 1952 which was followed by oral live polio vaccine by Sabin in 1962 (1). In those days it was not possible to develop a vaccine strategically. Today by studying the structure of HIV-1 along with its neutralizing monoclonal antibodies, scientist are trying to study the possibility of making a successful vaccine. IMMUNE MECHANISM: Bacteria, virus, or any substance which is recognized as foreign invader by the body’s defense mechanism has an immune reaction against it. Foreign substances are called antigens and as a defense, body produces protein molecules called antibodies (immunoglobulin). These antibodies binds to specific sites of the antigen and thus makes the antigen non reactive or neutralized (2). These specific sites on antigens are called antigenic determinants or epitopes (3). GENERAL STRUCTURE AND CLASSIFICATION OF IMMUNOGLOBULIN: As is shown in the figure 1 below the basic structure of the antibody contains two identical light chains and two identical heavy chains linked together by disulfide bonds, making a shape of Y. Based on the structure of heavy chains immunoglobulin can be divided into five different classes: IgG, IgM, IgA, IgD, and IgE. 2 Figure 1(a) The general structure of an antibody and (b) different immunoglobulins. The figure is taken from Carl Branden, John Tooze-Introduction to protein structuresecond edition (4). Figure 2 Schematic diagram of the production of antibodies. The figure is taken from Carl Branden, John ToozeIntroduction to protein structure- second edition (4) 3 As shown in figure 2 antibodies are produce by B cells inside the human body. The important thing to note is that each different antibody is produced only by a single clone of B cells (2) (5). PRODUTION OF MONOCLONAL ANTIBODIES: By somatic-cell hybridization of B-cells and T-cells, hybridomas were produced (B and T cells are different type of immune cells). These hybridomas have some of the normal genes of B or T cells and immortal-growth properties of cancer cells. Here the function of B cells is to produce monoclonal antibodies and T cells secrete various growth factors (5) (6). Figure 3 normal and cancerous hybridoma cells. The figure is taken from http://www.whfreeman.com/immunology/CH02/hy bridomas.htm (6) AIDS (ACQUIRED IMMUNODEFICIENCY SYNDROME): AIDS is cause by the destruction of CD4+ lymphocytes by the human immunodeficiency viruses (HIV-1, HIV-2, and the related simian immunodeficiency viruses SIV) in their respective host. STRUCTURE: HIV is a retro virus of class lenti-virus. RNA is the main genetic material. They also contain an enzyme reverse transcriptase which upon entering into the host cells converts the RNA into DNA. The 4 RNA genetic material is surrounded by a capsid protein (p24). The outer layer consists of matrix protein (P17) surrounded on the out by lipid bi-layer. The outer layer also consists of glycoprotein gp120 and gp41 which helps in attaching the virus to the host cells (4, 7). Figure 4 diagram of a HIV. Figure taken from “The EMBO J. Vol.18 No.5 pp.1124–1136, 1999” (8) 5 Figure 5 Schematic diagram showing the entry mechanism of HIV. Figure taken from S.J. Flint, L.W. Krug, V.R. Racaniello, A.M. Skalka- Principles of Virology- 2000 edition (7). MECHANISM OF VIRUS ENTERING INTO CELLS: The main receptor for HIV attachment on the host cell is CD4 which is present on T-cells and macrophages. Along with the receptor, the virus also needs co-receptors or chemokine. The interaction of the virus glyco-protein with chemokine triggers the fusion of virus and cellular membranes and thus the virus gains entry into the cytoplasm. HIV-1 virus is further classified on basis of the type of co-receptors it binds. -chemokine (Cxcr4) is present on T-cell hence the HIV is called Tcell-line-tropic strain of HIV-1 (X4) where as -chemokine (Ccr5) is present on macrophages so the name given is macrophage-tropic strain of HIV-1 (R5) (4)(7). HIV VACCINE: The reason why there is a lot of problem in making a vaccine for AIDS is that HIV is a very tricky virus and somehow avoids antibody binding. One of the tricks is that it has sugar molecules which 6 make the immune system believe that the virus is not foreign invader. Moreover the site with which it binds to CD4 is hidden behind loops of proteins and carbohydrates, so it is very difficult to neutralize it. On the other hand there is so much variation among different types of HIV that the antibody binding to one will not bind to another (9). The current trend in HIV vaccine research is by reverse immunology (which means designing a vaccine or an immunoglobulin by studying antibodies that have been induced to that antigen (10)). General Epitopes of the HIV-1 envelope glycoprotein along with the human monoclonal antibodies are listed in table 1. Some of these examples will be discussed briefly. In one research (11) scientist solved the X-ray crystallographic structure of HIV gp120 in complex with CD4 receptor and a neutralizing human antibody at 2.5 Angstrom resolution Table 1 shows HIV epitopes and Human antibodies released against them. Data taken from Nature march 2004 200-210 (10). Epitopes Human monoclonal Antibodies Cluster I of gp41 Clone 3, 246-D Transmembrane-proximal region of gp41 2F5, 4E10, Z13 CD-4 binding domain of gp120 Ig G1b12, 559/64D, 15e CD-4 induced epitope of gp120 17b,48D 1-2 mannose residues of gp120 2G12 V2 loop of gp120 697-D V3 loop of gp120 447/52-D, 19b,2182 7 Methods for getting proteins: The gp120 used in the experiment was from HXBc2 stain of HIV-1. They were produced from Drosophila Schneider 2 lines under the control of an inducible metallothionein. Chinese hamster ovarian cells were used to produce the two domain CD4 (D1D2, residues1-182). For monoclonal antibody 17b production, B-cell clone were used. B-cells were immortalized by Epstein-Barr virus, isolated from an HIV-1 infected individual and were fused with murine B-cell. Strategy for crystallization: The biggest hurdle for the researchers was that gp120 had extensive glycosylation and conformational heterogeneity. Hence the sample was deglycosylated and complexes were formed with various ligands. Moreover they also made truncations at termini and variable loops in various combinations. So 90% of carbohydrate was removed where as 80% of the non variable protein was retained. Theoretically they believed that the probability of crystal formation is greatly increased by such reduction of surface heterogeneity. This way they obtained crystals for ternary complex composed of truncated gp120, N-terminal two domains (DID2) of CD4 and a Fab from the human neutralizing monoclonal antibody 17b (see figure 6 below). A combination of techniques was used to solve the ternary structure via molecular replacement, isomorphous replacement and density modification. The two heavy atom compounds used were K3IrCl6 and K2OsCl6. Figure 6 shows the overall X-ray crystallographic structure of cd4, gp120 and fab 17b. Gp120 is in red, cd4 in yellow and fab 17b in light blue (light chain) and puple. 8 Figure is taken from Nature (1998) 393, 648–659 (11). In a recent study (12), a similar interaction between gp120 and a broadly neutralizing human antibody 2G12 was seen using X-ray crystallography (see figure 7 below). This antibody 2G12 was isolated from patients infected with HIV. It is very special in the sense that it is the only immunoglobulin isolated which successfully binds to carbohydrate part of glycoprotein. This carbohydrate is the one which shields potential antigenic epitopes. Figure 7 shows the x-ray crystallography structure of gp120 and 2G12. Figure taken from SCIENCE VOL 300 27 JUNE 2003 (12) Another good research regarding structure and function of monoclonal antibodies can be seen where scientist were trying to see the interaction of P24 (HIV capsid protein) with Fab13B5 antibody (see figure 8 shown below) (13). The importance of p24 is seen during virus assembly, maturation, and disassembly. Inside the mature virions lipid bilayer, p24 forms characteristically conical shaped shell surrounding the RNAnucleoprotein complex. Antibody-antigen interactions are very important as their detection serves as 9 diagnostic for HIV infection. Moreover the antibodies presence in larger amount is correlated with delayed progression to AIDS. Only 243 residues (called RH24) of p24 were considered for the research and they were derived from HXB2 strain of HIV-1. X-ray crystallography structure of the antigen antibody complex was determined in the previous research at 3 angstrom. In this research they only developed the x-ray structure of the antibody at 1.8 angstrom resolution. The most fascinating part of the research was that structural changes were observed between Fab bound to p24 and the free Fab as well as p24 bound to Fab and free p24. Figure 8 shows the interaction between P24 and the antibody part Vh and Ch. Figure taken from Structure, Vol. 8, 1069–1077, October, 2000(13) In effort for AIDS vaccine development researchers were trying to trace the binding site of a neutralizing monoclonal antibody 2F5 to HIV glycoprotein gp41 (14). The biophysical technique they use was Matrix-assisted laser desorption ionization mass spectrometry (MALDI-MS) in combination with 10 proteolytic protection assays. 2F5 is also among those antibodies which are isolated from HIV-1 infected individuals. It can neutralize most primary HIV-1 isolates in vitro. Therefore it is of significant interest to vaccine designers. Monoclonal antibody 2F5 was obtained from the AIDS Research and reference reagent program. The antihuman Fc-specific immunoglobulin secondary antibody was bought from Sigma chemical company. The gp140 protein was expressed from the HIV-1 JR-FL env gene. This gene contains the expression of gp120 linked to gp41 with disulfide linkage. The designated name of the protein is sos gp140. Under physiological conditions SOS gp140 was bound to 2F5 monoclonal antibody. Gp140 residues which were not protected by the antibody were cleaved using series of proteolytic enzymes (as shown in figure 9 below). Figure 9 shows a self explanatory diagrammatic presentation of the experiment. Figure taken from JOURNAL OF VIROLOGY, Nov. 2001, p. 10906–10911 Vol. 75, No. 22 (14). 11 The 16 amino acid long sequence shown in the diagram is found to be the main epitope on gp41. The research paper also mentions that, this sequence is significantly longer then the ELDKWA core epitope previously determined for 2F5 by peptide ELISA. This shows that MALDI-MS is a better method than ELISA. A typical mass spectrum of the experiment is shown below in figure 10. Figure 10 shows a mass spectrum detecting the16 amino acid chain residue. Figure taken from JOURNAL OF VIROLOGY, Nov. 2001, p. 10906–10911 Vol. 75, No. 22 (14). Another ground breaking research was reported by Susan Zolla-Pazner and co-scientist (8c). They identified a neutralizing antibody known as 447-52D which binds to V3 loop of the glycoprotein gp120. The structure of V3 was found to be similar to hairpins present in chemokine co-receptors (CCR5 and CXCR4) for HIV. All the structures were solved using NMR. 12 HIV needs to bind to chemokine co-receptors along with the main receptor CD4 in order to enter the host cell. This indicates that mechanism stimulating antibodies similar to 447-52D might help in developing a good vaccine against HIV. Figure 11. The first figure on the top is the backbone superimposition of 29 lowest-energy structure of V3 and the second figure is the ribbon diagram of the energy minimized average structure. Figure taken from Structure, Vol. 11, 225–236, February, 2003 (15). VACCINE DESIGN STRATEGIES: Among the most successful way of vaccine design is the Salk’s polio vaccine method. It should contain killed virus surface proteins eg in case of HIV, gp120, gp41 or p24. Next important one is Sabin’s polio vaccine method using live vector viruses engineered to carry genes encoding HIV proteins. A good example which is under clinical trial is where combination of elements such as pure gp120 and canary pox vector are used (combination of both Salk and Sabin method). In some trials even naked DNA is also been tried which code for one or more HIV genes. Live harmless bacteria engineered to carry genes encoding HIV protein are also under trial. Some more examples are Pseudovirions (non- 13 replicating HIV-like particles) and Replicons (non-HIV viruses engineered to carry genes encoding HIV proteins but they do not completely replicate) (16). THE DISCOURAGEMENT: Unlike Salk’s whole killed poliovirus HIV’s destroyed viruses couldn’t start immune reaction because the structure was disrupted. A biotech company Genentech tried the clinical trial with only gp 120 but it was only Lab HIV specific and the HIV mutates very fast. Even Sabin’s live attenuated strains of polio vaccine method also could not serve as a model because of the fear of active HIV getting in the human body of volunteers (17). CONCLUSION: HIV is a tricky virus and it is creating a lot of problem. To solve the problem a multidisciplinary research approach is a must. Identification of successful antibodies like 2G12 and 2F5 and their atomic level structure determination to study the interaction is a big success in keeping the hope up. There is no doubt that physical techniques like X-ray, NMR and spectroscopy are bringing us closer to understand our limitation in making an AIDS vaccine but still there is a long wait before a success story will be made. REFERENCES: (1) http://www.pbs.org/wgbh/aso/databank/entries/bmsalk.html (2) Carl Branden, John Tooze-Introduction to protein structure- second edition. (3) http://cancerweb.ncl.ac.uk/cgi-bin/omd?query=epitopes&action=Search+OMD (4) David M. Knipe, Peter M. Howley- Fundamental Virology-Fourth Edition. (5) http://www.whfreeman.com/immunology/CH05/kuby05.htm 14 (6) http://www.whfreeman.com/immunology/CH02/hybridomas.htm (please try Google or cut and paste in web browser) (7) S.J. Flint, L.W. Krug, V.R. Racaniello, A.M. Skalka- Principles of Virology- 2000 edition. (8) Carmen Berthet-Colominas, Ste´ phanie Monaco, Armelle Novelli1, Genevie`ve Sibaı¨2, Franc¸ois Mallet1 and Stephen Cusack3--Head-to-tail dimers and interdomain flexibility revealed by the crystal structure of HIV-1 capsid protein (p24) complexed with a monoclonal antibody Fab-- The EMBO Journal Vol.18 No.5 pp.1124–1136, 1999 http://www.nature.com/cgi-taf/DynaPage.taf?file=/emboj/journal/v18/n5/full/7591546a.html&filetype=pdf (9) Review Michael Hortens The Quest for Neutralizing Antibodies to HIV A I D S VA C C I N E A D V O C A C Y C O A L I T I O N • WWW. AVAC.ORG http://avac.org.phtemp.com/pdf/reports/AbResearch.pdf (10) Review Susan Zolla-Pazner- IDENTIFYING EPITOPES OF HIV-1 THAT INDUCE PROTECTIVE ANTIBODIES- Nature march 2004 200-210. http://www.nature.com/nri/journal/v4/n3/pdf/nri1307.pdf (11) Article Hendrickson, Kwong, P.D., Wyatt, R., Robinson, J., Sweet, R.W., Sodroski, J., and W.A. (1998)-- Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody --Nature 393, 648–659. http://www.nature.com/cgitaf/DynaPage.taf?file=/nature/journal/v393/n6686/full/393648a0_fs.html&content_filetype=pdf (12) Daniel A. Calarese, Christopher N. Scanlan, Michael B. Zwick, Songpon Deechongkit, Yusuke Mimura, Renate Kunert, Ping Zhu, Mark R. Wormald, Robyn L. Stanfield, Kenneth H. Roux, Jeffery W. Kelly, Pauline M. Rudd, Raymond A. Dwek, Hermann Katinger, Dennis R. Burton, 15 Ian A. Wilson,-Antibody Domain Exchange Is an Immunological Solution to Carbohydrate Cluster Recognition SCIENCE VOL 300 27 JUNE 2003 http://medicine.ucsd.edu/molpath/PDF/Wilson_CelBiol.pdf (13) Ste´phanie Monaco-Malbet, k Carmen Berthet-Colominas, Armelle Novelli, Nicole Battaı, Nadia Piga, Vale´ rie Cheynet,† Franc¸ ois Mallet, and Stephen Cusack-- Mutual Conformational Adaptations in Antigen and Antibody upon Complex Formation between an Fab and HIV-1 Capsid Protein p24 Structure, Vol. 8, 1069–1077, October, 2 http://www.sciencedirect.com/science?_ob=MImg&_imagekey=B6VSR-41M94YH-7G&_cdi=6269&_user=585204&_orig=search&_coverDate=10%2F31%2F2000&_qd=1&_sk=999919989&view=c& wchp=dGLbVtz-zSkzS&md5=36f4c2d713c65410a03bef4d8552d7bc&ie=/sdarticle.pdf (14) CAROL E. PARKER,1* LEESA J. DETERDING,1* CHRISTINE HAGER-BRAUN,1 JAMES M. BINLEY,2 NORBERT SCHU¨ LKE,3 HERMANN KATINGER,4 JOHN P. MOORE,2 AND KENNETH B. TOMER1 Fine Definition of the Epitope on the gp41 Glycoprotein of Human Immunodeficiency Virus Type 1 for the Neutralizing Monoclonal Antibody 2F5-JOURNAL OF VIROLOGY, Nov. 2001, p. 10906–10911 Vol. 75, No. 22 http://jvi.asm.org/cgi/reprint/75/22/10906 (15) Michal Sharon,1 Naama Kessler,1 Rina Levy,1 Susan Zolla-Pazner,2 Matthias Go¨ rlach,3 and Jacob Anglister--Alternative Conformations of HIV-1 V3 Loops Mimic _ Hairpins in Chemokines, Suggesting a Mechanism for Coreceptor Selectivity --Structure, Vol. 11, 225–236, February, 2003 http://www.sciencedirect.com/science?_ob=MImg&_imagekey=B6VSR-47WC25R-GF&_cdi=6269&_user=585204&_orig=browse&_coverDate=02%2F28%2F2003&_sk=999889997&view=c&wchp =dGLbVzz-zSkzV&md5=d3bbc4b5da7f2b46e84c700b1203fa63&ie=/sdarticle.pdf 16 (16) http://www.niaid.nih.gov/daids/vaccine/slidehivvacupdate/slide15.htm (17) http://pubs.acs.org/cen/topstory/7935/7935sci1.html 17



























![Anti-PCB antibody [3H2AD9] ab110314 Product datasheet 3 Images Overview](http://s1.studyres.com/store/data/000076345_1-acbfa58e194757c519d151062b812354-150x150.png)