Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Polyclonal B cell response wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Innate immune system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Hygiene hypothesis wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
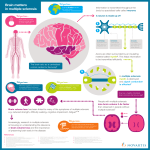
Management of multiple sclerosis wikipedia , lookup
Multiple sclerosis signs and symptoms wikipedia , lookup
Pathophysiology of multiple sclerosis wikipedia , lookup
Sjögren syndrome wikipedia , lookup