Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Node of Ranvier wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
End-plate potential wikipedia , lookup
Electromyography wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Proprioception wikipedia , lookup
Circumventricular organs wikipedia , lookup
Synaptogenesis wikipedia , lookup
Fine Structural Analysis of Extraocular Muscle Spindles
of a Two-Year-Old Human Infant
Roland Blumer,' Julius-Robert Lukas,2 Martin Aigner,' Reginald Bittner,l
Isabella Baumgartner,2 and Robert Mayr1
PuRi'OSi:. To clarify whether structural peculiarities formerly described in extraocular muscle (EOM)
spindles of aged persons are already present in EOM spindles of a 2-year-old infant.
Mi-1 HODS. Distal halves of two EOMs obtained from a 2-year-old multiorgan donor were immersionfixed and prepared for electron microscopy. The fine structure of 10 muscle spindles and of 1 "false
spindle" was investigated.
Rrsui.TS. Extraocular muscle spindles of an infant 2 years of age had 2- to 4-layered outer capsules,
376 /Am (range, 217-606 jum) long and 97 ju.m (range, 55-140 jum) wide. In 10 EOM spindles, 4
to 16 intrafusal muscle fibers (mean. 7.9) were present. From a total of 79 intrafusal fibers, 43 (54%)
were nuclear chain fibers, and 8 of the 43 exhibited posttraumatic degenerative changes. Thirty-six
(46%) intrafusal fibers indistinguishable from extraiusal fibers were called anomalous fibers. No
nuclear bag libers were found. Each muscle spindle contained a variable number of chain fibers and
at least one anomalous fiber. Sensory nerve terminals were restricted to the 35 normal chain fibers
but were absent from damaged chain fibers and from anomalous fibers. One "false spindle" without
a periaxial space was composed of three anomalous fibers and one chain fiber, all of them devoid
of sensory terminals.
CONCLUSIONS. Most structural particularities of human EOM spindles described in aged persons are
already found in the infant. They cannot be interpreted as age-related changes, but rather they
represent specific features of human EOM spindles. (Invest Opbthalmol Vis Sci. 1999;4():55-64)
P
roprioception from human extraocular muscles (EOM)
is thought to play an important role in the development
of a normal binocular vision.1 Several reports attribute
increasing importance to proprioceptive input from EOMs
within the oculomotor system.2"'' In humans, muscle spindles
are accepted to be a regular constituent in EOMs, 5 ' 10 and their
characteristic distribution and high density in different human
EOMs are strong indications of the functional importance of
these structures."
Ultrastructural observations on human EOM spindles are
sparse. Other than a brief description of the fine structure of
the sensory endings, 1 ' only one detailed study of the ultrastructural features of human EOM spindles has been published. 7
This report was exclusively based on material of three aged
persons (58 years, 70 years, and 74 years). Transmission electron' and light microscopic observations Hy demonstrated that
aged human EOM spindles differ in their intrafusal muscle fiber
composition from skeletal muscle spindles and from EOM
spindles of various mammals.' 1 2 " 1 5 Nuclear bag fibers and
nuclear chain fibers are regular constituents of mammalian and
human skeletal muscle spindles and of EOM spindles of ungulates.7AZ-{5 in human EOM spindles, the intrafusal fiber com-
From the 'institute of Anatomy, and the ^Department of Ophthalmology and Opiometry, University of Vienna-General Hospital, Austria.
Submitted for publication December 19, 1997; revised July 10,
1998; accepted July 31, 1998.
Proprietary interest category: N.
Reprint requests: Roland Winner, Institute of Anatomy, Department 2, University of Vienna, 1090 Vienna, Austria.
position is different. Most human EOM spindles lack bag fibers,
which represent only 2% of EOM intrafusal fibers of aged
persons. Resides chain fibers, unmodified muscle fibers resembling extraiusal muscle fibers were found to be a regular
constituent of human EOM spindles, both representing approximately equal numbers of the total intrafusal fiber contingent of
aged persons.'" 10 Unmodified muscle fibers in aged human
EOM spindles were designated anomalous muscle fibers.7
In addition to a different intrafusal fiber composition,
human EOM spindles of old persons vary from skeletal muscle
spindles with respect to the periaxial space, which is either ill
defined or absent in human EOM spindles. 7 Another difference
from skeletal muscle spindles is that in aged human EOM
spindles the inner capsule never invested all intrafusal fibers.7
Several characteristics of human EOM spindles were considered to be age-related alterations.7
Up to now no study has reported on human EOM spindles
of younger individuals. The aim of this study was to compare
human EOM spindles of a 2-year-old infant with data from those
of aged persons 7 " to clarify whether the structural characteristics of old human EOM spindles are age-related alterations as
suggested by Kuskell,7 or whether they represent normal but
unique features of human EOM spindles.
MATERIALS AND METHODS
Distal halves of a left inferior rectus and a left superior rectus
were obtained from a 2-year-old multiorgan donor who died 72
hours after severe head trauma. This study was performed in
accordance with the Austrian federal transplantation law.
Investigative Ophthalmology & Visual Science, January 1999, Vol. -10, No. I
Copyright © Association for Research in Vision anil Ophthalmology
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
55
56
Blumer et al.
10VS, January 1999, Vol. 40, No. 1
45
40
35
30
25
20
15
10
I
5-7.5
•
7.6-10
10.1-12.5
12.6-15
15.1-17.5
17.6-20
20.1-22.5
22.6-25
25.1-27.5
FK.UKI: 1. Diameter distribution for intrafusal fibers (n = 79, black bars) in the: muscle spindle polar
regions and for cxirafusul fibers (» = 194. white bars) adjacent to muscle spindles. There is a considerable
overlap in diameters between intrafusal fibers in muscle spindle poles and neighboring extrafusal fibers.
Methods for securing human tissue were humane, included
proper consent and approval, and complied with the tenets of
the Declaration of Helsinki.
Muscles were cut longitudinally into four strips and cut
transversely with a length of 6 mm and a diameter of 3 mm.
The tissue was immersion-fixed in 2% parafornialdehyde and
2.5% glutaraldehyde in 0.1 M cacodylate buffer at pll 7.4 for 1
day. After rinsing in the same buffer, specimens were postfixed
in buffered 1% osmium tetroxide, dehydrated in graded solutions of ethanol, and embedded in Epon.
Semi-thin cross sections (1 jam) were retained at intervals
of 20 /urn, stained with toluidine blue, and examined under the
light microscope (LM) for the presence of muscle spindles.
When muscle spindles were identified in the LM, every semithin section was mounted on a slide. Ultrathin sections were
cut at appropriate intervals, mounted on dioxane formvarcoated copper grids, immersed in an aqueous solution of 2%
uranyl acetate followed by a solution of 0.4% lead citrate in 0.1
M sodium hydroxide, and examined under a transmission electron microscope (TEM, model EM9; Zeiss, Oberkochen, Germany).
Ten human EOM spindles (seven spindles of the left inferior rectus and three spindles of the left superior reel us) and
one false spindle of the left inferior rectus were investigated.
All muscle spindles and the false spindle were analyzed
throughout their capsule length, and in some muscle spindles
the investigations were continued up to 100 jam into the
extracapsular regions. The lengths of the muscle spindle capsules were calculated by counting both their serial semi-thin
sections (1 jam) and their ultrathin sections (100 nm). One
tissue block containing muscle spindles was reorientated and
cut longitudinally to assess the degree of" muscle fiber contraction due to immersion fixation.
RESULTS
Identification of human FOM spindles in cross sections was
only possible if the intrafusal fibers were surrounded by a
distinct capsule. Polar regions outside the capsules could not
be recognized because the extrafusal fibers did not differ from
extracapsular intrafusal fibers, neither in diameter (extrafusal
fibers, 14.6 ± 3.3 ju.ni; intrafusal fibers, 12.5 ± 4.2 jam) nor in
structure (Fig. 1). As was noted in a former study, all muscle
spindles investigated were located in the transition zone between the orbital and global layers of the F.OMs."
Outer Capsule
The capsule lengths varied from 93 ju.m to 6()6 jam with a mean
of 376 jam. The largest diameters of all 10 muscle spindles
varied between 55 jam and 140 ju,m with a mean of 97 jam
(Table 1).
The intrafusal fibers of the human I-OM spindles were
encircled by a capsule that consisted of concentric layers of
flattened perineurial cells. The perineurial cells were covered
with a basal lamina on both sides, and the spaces between
adjacent perineurial cell layers contained collagenous fibers.
Blood vessels were occasionally observed between neighboring perineurial cell layers. In the polar regions of the muscle
spindles, the capsules consisted of one to two cell layers
closely encircling the intrafusal fibers. Toward the equatorial
regions, the diameter of the spindles and the thickness of their
capsules gradually increased, the latter to two to four layers.
In two spindles, all intrafusal fibers in the equatorial region were concentrated in the center of the muscle spindle
cross sections, and a well-developed periaxial space separated
the intrafusal fibers from the spindle capsule (Fig. 2). In four
muscle spindles the intrafusal libers were uniformly distributed
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
Infant Extraocular Muscle Spindles
10VS, January 1999, Vol. 40, No. 1
TABLE
57
1. Morphological Features of 10 Human EOM Spindles of a 2-Year-Old Infant
Intrafusal Fibers per
Muscle Spindle
Nuclear Chain
Fibers
4
3
6
6
2 + 2*
1 + 2*
i + r
6
7
7
8
8
11
16
79 (7.9)
Anomalous
Fibers
1
%
4-
55
l
7.
120
90
95
110
140
140
36 (3.6)
967 (97)
3759 (376)
3
i
4 + 1*
3 + 2*
I
»
7
35 + 8*
(3.5 + 0.8*)
Capsule
Length (jmm)
220
606
420
280
550
93
508
217
350
515
2
4
9
Equatorial
Diameter (/u,m)
65
77
75
Periaxial
Space
+
+
+
+
+
+
+
+
+
Values in parentheses are mean values.
* Damaged chain fibers without sensory terminals.
on cross sections of the equatorial region, and the periaxial
space was less developed. In three muscle spindles a variable
number of intrafusal fibers were located in the center within
the equatorial region, whereas numerous intrafusal fibers exclusively of the anomalous type had an eccentric position (Fig,
3). A periaxial space was visible only around the intrafusal
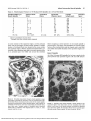
FIGURE 2. Semi-thin cross section through a muscle spindle, its equatorial region containing six chain fibers, and one anomalous fiber (AF),
All intrafusal fibers concentrated in the center of the muscle spindle.
Concentric periaxial space around intrafusal fibers. Inner capsule 0)
consisting of two layers: outer layer surrounding all intrafusal fibers,
inner layer only a variable number of intrafusal fibers, associated
muscle fiber (*) between outer capsule layers. N, nerve; C, outer
capsule. Scale bar, 10 jam.
fibers occupying a central position. In one muscle spindle a
periaxial space was absent, and througliout the capsule length
between the intrafusal fibers and the capsule only a small cleft
was visible that hardly increased in the equatorial region (Table 1).
Inner Capsule
All of the 10 human EOM spindles had an inner capsule in the
para-equatorial and equatorial regions. LM and TEM observa-
FIGURE 3. Semi-thin cross section through a muscle spindle in the
equatorial region. Four chain fibers and one anomalous fiber (AF)
placed in the center of the muscle spindle surrounded by periaxial
space and inner capsule. Six anomalous fibers with an eccentric position and outside of inner capsule. N, nerve. Scale bar, 10 fim.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
58
IOVS, January 1999, Vol. 40, No. 1
Blumer et al
tions revealed that in one muscle spindle this inner capsule
consisted of two cell layers: the outer layer surrounded all
intrafusal fibers and the inner one invested a different number
of intrafusal fibers (Tig. 2). Nine muscle spindles exhibited a
one-layered inner capsule ensheathing a variable number of
intrafusal fibers. Anomalous muscle fibers located adjacent to
the spindle capsule were not enveloped by inner capsule cells
(Fig. 3).
The cells of the inner capsule consisted of a prominent
cell body that appeared to be rounded or irregularly shaped. A
variable number of thin cytoplasmic cell processes emanated
from the cell body to encircle individual intrafusal fibers. Adjacent cell processes had cell-to-cell contacts at numerous
points. TEM demonstrated that the cells of the inner capsule
lacked a basal lamina investment.
Intrafusal Fibers
Ten EOM spindles of the infant contained between 4 and 16
intrafusal fibers, with a mean of 7.9 intrafusal fibers. A total of
79 intrafusal fibers were observed, 43 nuclear chain fibers
(54%) and 36 anomalous fibers (46%). In addition to nuclear
chainfibers,anomalous fibers were a regular constituent of the
infant spindles. Each muscle spindle contained at least one
anomalous fiber (Table 1).
Nuclear Chain Fibers
Thirty-five of 43 nuclear chain fibers exhibited a normal morphology. In the polar regions of the muscle spindles most of
these typical nuclear chain fibers had subsarcolemmal myonuclei, and their diameters were similar to those of the anomalous
fibers. A small number of nuclear chainfibershad single central
myonuclei in their polar regions and also in parts of their
extracapsular regions. In the para-equatorial regions the myonuclei of all nuclear chain fibers were found in a central
position, separated from each other by interspaces filled with
undifferentiated electron translucent sarcoplasm. In the equatorial regions these interspaces gradually diminished in such a
way that adjacent myonuclei of the chain were touching each
Other. In the equatorial regions the mean diameter of the
nuclear chain fibers was significantly smaller than that of the
spindle poles (equator: 8.5 ± 2.2 ju,m [mean ± SD], pole:
10.8 ± 2.8 /im, Student's f-test, P < 0.001, n = 35), and the
central myonuclei were only surrounded by a thin layer of
sarcoplasm (Figs. 4, 5). Four nuclear chain fibers exhibited
short areas in their equatorial regions in which two myonuclei
were lying side by side (Fig. 6). Length measurements of
sarcomeres in longitudinally cut extrafusal fibers revealed that
numerous muscle fibers exhibited contracted sarcomeres
(length, 1.6 furi). In one nuclear chain fiber the sarcomeres
were contracted to an extent that no I-bands were visible (Fig.
7). The contraction of the muscle fibers might have been
caused by the fixation procedure. Thereby in nuclear chain
fibers, myonuclei that were originally arranged in a chain came
to lie side by side.
Ultrastructural images revealed that throughout their capsule length the sarcomeres of the nuclear chainfibersconsisted
of thick Z-lines and small I-bands. Within the H-bands, a distinct
M-line was present (Fig. 7). All nuclear chain fibers were
endowed with sensory terminals. Three muscle fibers received
a single motor terminal within the capsules of their muscle
spindles.
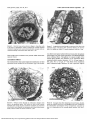
FIGURE 4. Ultrathin cross section through the equatorial region of a
muscle spindle composed of six chain fibers and one anomalous fiber.
Strikingly reduced diameters of chain fibers are seen in the equatorial
region; central myonuclei are surrounded by a thin layer of sarcoplasm,
with all chain fibers receiving sensory terminals (ST). I, inner capsule;
AF, anomalous fiber; N, nerve. Scale bar, 5 jitm.
Eight of 43 nuclear chainfibers(Table 1) exhibited degenerative changes that may have been induced by the severe head
trauma of the donor. In these damaged nuclear chain fibers
clustered myofilaments, areas of myofibrillolysis, swelling of
the mitochondria, and dilatation of the sarcoplasmic reticulum
were present; in addition, karyorrhexis of central myonuclei
and pyknotic central myonuclei were evident (Fig. 8). In these
nuclear chain fibers, three to nine single central myonuclei
were present on cross sections throughout their capsule
length. They never formed a continuous chain but were always
separated by variable intervals of sarcoplasm. A small number
of extrafusal muscle fibers exhibited comparable ultrastructural changes. All damaged nuclear chain fibers were devoid of
sensory terminals (Table 1, Fig. 8). On two of these muscle
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
JOVS, January 1999, Vol. 40, No. I
FIGURK 5. Detail of cross section shown in Figure 4, chain fiber with
a thin layer of sarcoplasm. Sensor}' terminal (ST) contains electron
translucent axoplasm and numerous mitochondria, synaptic cleft (arrowhead) lacking a basal lamina (BL). Scale bar, 1 |U,m.
fibers single motor terminals were present within their ultracapsular course.
Anomalous Fibers
All intrafusal fibers that lacked equatorial modification of their
myonuclei were ascribed to the anomalous type. The anoma-
Infant Extraocular Muscle Spindles
59
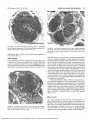
FIGURE 7. Longitudinal section through a contracted chain fiber in its
equatorial region. Sarcomeres have prominent M-lines and thick Zlines. No I-bands are visible. ST, sensory terminals, Scale bar, 1 fxm.
lous fibers had typical subsarcolemmal myonuclei throughout
their lengths, and, thus, their appearance was indistinguishable
from extrafusaJ fibers. Throughout the capsule length of the
human EOM spindles, the diameter of the anomalous fibers
remained rather constant (equator: 14.7 ± 4.0 ju,m [mean ±
SD], pole: 14.3 ± 4.3 jam, n = 36). Most of the anomalous
fibers exhibited larger diameters in their equatorial regions
M
FIGURE 6. Oblique section through an artifactually changed chain
fiber in its equatorial region. Two nuclei are side by side, sensory
terminals (ST) contain electron translucent axoplasm and numerous
mitochondria, one obliquely cut sarcomere with a clear M-line (arrowhead) is shown within the H-zone. Scale bar, 5 Mm-
FIGURE 8. Damaged chain fiber displaying structural lesions. Myofilaments are clustered and areas of myofibrillolysis are seen. Swelling of
the mitochondria and dilatation of the sarcoplasmatic reticulum are
seen, as is karyorrhexis of the centrally placed nucleus. Scale bar, 5
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
60
Blumer et al.
IOVS, January 1999, Vol. 40, No. 1
were called associated muscle fibers. Associated muscle fibers
were observed in two human EOM spindles (Fig. 2). As associated muscle fibers had subsarcolemmal myonuclei throughout their length they were indistinguishable from anomalous
fibers and extrafusal fibers. Neither sensory nor motor terminals were found on associated muscle fibers within their
course between outer capsule layers.
Myosatellite Cells
All chain fibers outside their areas of sensory innervation, all
damaged chainfibers,anomalous fibers, and associated muscle
fibers throughout their lengths had typical satellite cells like
extrafusal fibers. Satellite cells were located on the intrafusal
fiber surface and covered by the basal lamina of the muscle
fiber as in skeletal muscle. They had typical oval shapes and
moderately indented the surface of their intrafusal fibers and
associated muscle fibers. They possessed a large nucleus that
was surrounded by a thin layer of cytoplasm.
Sensory Terminals
9. Chainfiberis shown immediately beforefiberbreakage,
myofibrillar material ts dispersed, sarcoplasm contains swollen mitochondria characterized by degeneration of cristae (arrowhead). ST,
sensory terminal. Scale bar, 5 ixm.
FIGURE
than normal chain fibers. In seven muscle spindles the anomalous fibers remained within their capsule space throughout
the whole capsule lengths. In three muscle spindles anomalous
fibers were penetrating at least the inner layer of the outer
capsule at variable points: In one muscle spindle, three anomalous fibers coming from outside entered the muscle spindle
and, after running a short distance within the capsule space,
these anomalous fibers exited the muscle spindle. In another
muscle spindle serial sections demonstrated that two anomalous fibers were running in the equatorial zone for a distance of
120 jum between adjacent capsule layers. In a third muscle
spindle, three anomalous fibers successively penetrated the
muscle spindle capsule in the polar region.
TEM images revealed that the anomalous fibers contained
numerous mitochondria with occasional subsarcolemmal accumulations. Sensory terminals were never found on anomalous
fibers. Single motor terminals were observed on six anomalous
fibers within the capsular regions.
Breakage, Interruption, and Intrafusal Fiber
Splitting
Four nuclear chain fibers with areas of sensory innervation in
three muscle spindles abruptly ended at variable points within
the encapsulated regions. TEM pictures demonstrated that
shortly before breakage, these muscle fibers contained swollen
mitochondria, and their myofibrillar material appeared dispersed (Fig. 9)- One nuclear chain fiber with sensory terminals
was interrupted for a distance of 35 ju,m. Another nuclear chain
fiber with sensory terminals split after having passed the equatorial region.
Associated Muscle Fibers
Musclefibersrunning for variable distances between neighboring outer capsule layers without entering the capsule space
Sensory terminals were present on all normal chainfibers,and
they enwrapped the individual fibers, forming irregular coils.
Their areas of sensory innervation varied between 30 /xm and
360 jLtm. Sensory terminals were located not only in the equatorial regions, but in most chainfibersthey also covered at least
one side of the para-equatorial regions. In one muscle spindle
the sensory terminals on two chainfibersextended on one side
into the extracapsular region of the muscle spindle. TEM observations indicated that the sensory terminals often deeply
indented the surfaces of the chain fibers. Rarely, the sensory
terminals extended from the surface into the center of the
intrafusal fibers (Fig. 4). Sensory terminals contained numerous
mitochondria that were either distributed uniformly or were
concentrated near the synaptic cleft. Their axoplasm was electron translucent. The intrafusal fiber basal lamina covered the
outer surfaces of the sensory terminals but was absent within
the synaptic clefts. The synaptic clefts measured 25 nm, In a
small number of terminals, there were junctional complexes
that were shaped like discs between opposed plasma membranes of sensory terminals and intrafusal fibers (Figs. 5, 6, 7).
On four chain fibers in three muscle spindles sensory
nerve endings exhibited a dense axoplasm full of mitochondria
with swelling of the matrix and reduction of their cristae,
which may be interpreted as Wallerian degeneration. Three
chain fibers were exclusively endowed with such degenerated
sensory endings. On one chain fiber normal and degenerated
sensory terminals were observed. Degenerated sensory terminals covered fiber lengths between 30 jam and 130 fxm (Fig.
10).
Motor Terminals
On two chain fibers single motor terminals were found in the
immediate vicinity of the sensory regions. TEM images revealed
that they were positioned superficially without causing a depression in the surface of the intrafusal fibers. Their subsynaptic membranes were smooth.
Single neuromuscular junctions were observed on two
chain fibers and on six anomalous fibers at variable points
within the spindle capsules. On three chain fibers single motor
terminals were located in their extracapsular regions. All these
motor terminals had numerous synaptic knobs. These knobs
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
Infant Extraocular Muscle Spindles
IOVS, January 1999, Vol. 40, No. 1
FIGURE 10. Chain fiber with densely appearing sensory terminals (ST)
clue to Wallerian degeneration. The mitochondria show swelling of the
matrix and reduction of the cristae. Scale bar, 5 p-m.
6l
FIGURE 12. Cross section through a false muscle spindle containing
one muscle fiber with a central nucleus and three anomalous fibers.
One of them contains a motor terminal (*) with synaptic knobs. Scale
bar, 5 Jim.
indented the fibers' surfaces and the subsynaptic membranes
were folded (Fig- H).
False Spindle
One encapsulated structure contained four muscle fibers but
lacked sensory terminals throughout the capsule length (330
jam). It is extremely unlikely that sensory endings have been
missed, because they always covered a substantial length of the
muscle fiber, and because ultrathin sections were taken at very
short intervals (5-7 jutm). This structure without sensory investment was therefore classified as a false spindle. The capsule
consisted of three cell layers and was always in close contact to
the muscle fibers throughout its length. A periaxial space was
absent, and its maximum width was 35 /xm. The capsule cells
were covered on both sides with basal laminae. Individual
muscle fibers were encircled by processes that extended from
the innermost layer of the capsule. Reconstruction of serial
sections of the encapsulated region revealed that there were
three muscle fibers of the anomalous type with only subsarcolemmal myonuclei. One muscle fiber exhibited a row of five
central myonuclei with variable amounts of sarcoplasm between them. Thus, it is questionable whether this fiber should
be classified as a chain fiber.
Motor terminals were found within the encapsulated region of this false spindle on two anomalous fibers. One anomalous fiber was multiply innervated with three neuromuscular
contacts, whereas the other anomalous fiber had a single motor
terminal. Apart from their larger content of densely packed
vesicles, motor terminals in the false spindle did not differ in
their morphology from extracapsular motor endings on chain
fibers and those on anomalous fibers in infant EOM spindles
(Figs. 12, 13).
DISCUSSION
FIGURE 11. Anomalous fiber motor terminal (*) with five synaptic
knobs indenting the muscle fiber surface and exhibiting short synaptic
folds. C, outer capsule. Scale bar, 5 /wm.
This study describes for the first time the ultrastructure of
human EOM spindles at an age at which the development of
binocular vision and interocular alignment were not yet complete. Most morphologic characteristics of human EOM spindles as described by Ruskell7 and Lukas et al.8 in aged persons
were also present at the age of 2 years. In particular, EOM
spindles of the 2-year-old infant and those of old individuals7"1*
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
62
Blumer et al
IOVS, January 1999, Vol. 40, No. 1
•/»-•*& .V*:xt
.ft-
•-
••**»**?•'
FIGURE 13. Detail of cross section shown in Figure 12. Two knobs of
motor terminal, one containing numerous mitochondria and region
with densely packed vesicles, synaptic cleft containing basal lamina
(arrowhead), and subsynaptic membrane with short folds. Scale bar, 1
had almost the same unique intrafusal fiber composition. Besides typical nuclear chain fibers, anomalous fibers lacking
equatorial nucleation were a regular constituent of human
EOM spindles. All spindles of the infant and most spindles of
the aged persons7"9 were devoid of nuclear bag fibers. In
contrast to that, mammalian skeletal muscle spindles and EOM
spindles of ungulates were reported to contain exclusively bag
fibers and chainfibers,with the regular presence of at least one
bag fiber per spindle.712"15 In infant EOM spindles the total
number of chain fibers (54%) was higher than that of the
anomalous fibers (.46%). In the aged,7'8 the number of bag
fibers was only 2% of the total; anomalous fibers were 50% to
73% in one study7 and 43% in another8 (Table 2). Another
characteristic of EOMs described by Ruskell7 and confirmed by
Lukas et al.8'9 was the occurrence of false spindles, encapsulated structures containing muscle fibers without any sensory
innervation. One false spindle was identified and analyzed in
the infant.
Other features distinguishing human EOM spindles from
mammalian muscle spindles are the limited or absent perdaxial
space7 and the fact that intrafusal fibers and adjacent extrafusal
fibers exhibit similar diameters. Although measurements (Table 1) revealed that the mean equatorial diameter and the mean
TABLE
number of intrafusal fibers per spindle were similar in the
infant and in aged persons, the periaxial space seemed to be
more pronounced in infant spindles. The absolute diameter of
the periaxial space remained constant with increasing age,
whereas intrafusal fibers in the infant are smaller (12.5 p.m)
compared with the aged (153 |xm),7 The periaxial space protects the sensory nerve terminals of the intrafusal fibers from
nonspecific stimuli like contraction of neighboring extrafusal
fibers.16 Thus, because of ..a relatively larger periaxial space,
infant muscle spindles appear to be better protected from
mechanical interference.
In the investigation by Ruskell,7 five nuclear bag fibers
were described. Only one of the bag fibers had an accumulation of myonuclei in its "bag" region, but four of them had two
myonuclei abreast over a short length of each fiber. The classification of intrafusal fibers with two nuclei abreast as bag
fibers needs attention. In skeletal muscle spindles a bag fiber is
defined as an intrafusal fiber with an accumulation of myonuclei in its "bag" region.17 In the tenuissimus of the cat, Adal18
qualified intrafusal fibers with two nuclei side by side as nuclear chain fibers. In most skeletal muscle spindles the bag
fibers are of a larger diameter throughout their length than
chain fibers.17 Furthermore, in skeletal muscle spindles, bag
fibers and chain fibers differ in the M-band structure of the
equatorial sarcomeres. Bag fibers contain two faint M-lines
within the H-band, whereas chain fibers show one single welldefined M-line.19'20 In the present study four intrafusal fibers
were found with two myonuclei abreast. However, apart from
that, these four fibers were indistinguishable in their morphology from chainfibers.We concluded that intrafusal fibers with
only two nuclei abreast are most likely chain fibers that were
artifactually changed during the fixation procedure.
The localized pathologic changes observed in eight chain
fibers, and scattered extrafusal fibers can be explained by the
severe head trauma that caused the death of the infant 72 hours
after injury.21 Damages of the myelin sheaths of single axons
and electron dense axoplasm and altered mitochondria observed in four sensory endings are considered to be signs of
Wallerian degeneration, probably caused by a rupture of the
axolemma (Lassmann H, personal communication, March
1997). The majority of muscle fibers, axons, and nerve terminals have not been visibly damaged by this trauma. We believe
that, in general, the severe head trauma did not affect the
reliability of the results presented in this study.
In humans22 in contrast to other species (e.g., rats),23'2''
the development of skeletal muscle spindles is completed
before birth. So far, data on the development of human EOM
spindles are not available in the literature. In the present study
we did not observe any features of continuing spindle development such as an incomplete capsule, an ongoing fusion of
myoblasts, or close apposition of intrafusal fibers. However,
2. Intrafusal Fiber Composition in Human EOM Spindles of Persons of Different Ages
Author
Ruskell7
Ruskell7
Lukas et al.8
This study
Age of the
Individuals (y)
% of Nuclear
Chain Fibers
% of Anomalous
Fibers
70,74
58
67, 72, 83
25
48
55
54
73
50
43
46
2
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
% of Nuclear
Bag Fibers
IOVS, January 1999, Vol. 40, No. I
postnatal maturation of extrafusal libers with respect to their
fiber thickness and their miiochondrial pattern occurs in cat23
and rat26 IZOMs. Maturation of extrafusal libers continues up to
the adult age of these animals. In monkeys, the singly innervated muscle fibers of the orbital layer attain their definitive
size and mitochondrial content within the first 6 months.27 By
comparing the mean intrafusal liber diameter in the EOMs of
the infant (12.5 /xm) with that in the EOMs of aged persons
(15.3 jum), an increase in the intrafusal fiber thickness is noted.
The circumstances of a high incidence of anomalous fibers
in aged human EOMs, such as frequent lipofuscin deposits, loss
of liber length, and fiber fragmentation of regular intrafusal
fibers, stimulated suggestions by Ruskell' that anomalous fibers
are arguably incursions of extrafusal fibers replacing degenerated intrafusal fibers and that false spindles may represent the
culmination of spindle degeneration, with all nuclear bag and
nuclear chain libers destroyed and replaced by extrafusal fibers. In this study in the infant, anomalous fibers were almost
equal in number to chain libers (Table 2). Loss of fiber length
was observed in four chain libers, fragmentation in one chain
fiber, and splitting in another chain fiber. However, lipofuscin
was not present at the age of 2 years. The false spindle examined was composed of three anomalous libers and one chain
liber, with all four lacking any sensory innervation. Apart from
the presence of lipofuscin most aberrant features of human
EOM spindles cannot be interpreted as age-related changes. It
has been suggested that the special muscle spindle morphology could be an expression of phylogenetic redundancy.'
However, it seems very unlikely that evolutionary redundant
receptors would be seen so frequently. As previously reported,H muscle spindles were at least as frequent in human HO Ms
as in skeletal muscles, with known high muscle spindle density
when compared gram for gram.
Experimental studies in skeletal muscles gave evidence
that dcvascularization28 and application of myotoxic substances29 could induce muscle spindle degeneration with subsequent regeneration. It is important to note that in all these
studies regeneration of intrafusal fibers was found to originate
from myosatellite cells. Necrotic sarcoplasm of the degenerated intrafusal libers is removed by phagocytes. The myosatellite cells survive, and during regeneration they segregate
myoblasts, which fuse to myotubes. Analogous to the transformation of undifferentiated intrafusal fibers into bag fibers and
chain fibers by primary sensory nerve terminals in normal
dcvelopmenl,22l2VS(> innervaiion with sensory terminals was
reported to induce maturation to bag libers and chain fibers
with typical equatorial nucleation in spindle regeneration even
in adult rats.2* In human EOM spindles of the infant, myosatellile cells were regularly present in intrafusal fibers except in
their equatorial regions, which are endowed with sensor)'
terminals. These facts do not support the hypothesis of replacement of degenerated intrafusal fibers in human EOMs by
incursion of extrafusal fibers.'
On the other hand, reconstructions of 10 muscle spindles
of the 2-year-old infant revealed that muscle libers indistinguishable from adjacent extrafusal fibers were found to be
interposed for variable distances between the layers of the
outer spindle capsule (= associated muscle fibers) and as
so-called "anomalous" fibers in an intrafusal position. In contrast to typical intrafusal fibers, most anomalous fibers were not
ensheathed by an inner capsule. In the infant, two muscle
spindles contained one associated muscle liber each, but all 10
Infant Extraocular Muscle Spindles
63
muscle spindles had anomalous libers (1-7 per spindle) together with nuclear chain libers as regular constituents. Eight
of 36 anomalous fibers did not run the whole length within the
capsule space, but rather penetrated one muscle spindle capsule in the para-equatorial region and left the capsule space in
another muscle spindle to run between outer capsule layers
before reentering at an equatorial level. Furthermore, anomalous fibers entered one muscle spindle from outside and, after
running within the capsule space, left the muscle spindle
again. These findings together with abrupt ending and fragmentation of intrafusal fibers with typical equatorial nucleation
are in conformity with the situation in the aged adult.7 In
contrast to aged human EOM spindles, where 8 of 120 anomalous fibers were generously supplied with sensory endings,7
all 36 anomalous fibers of the infant were lacking sensory
terminals. However, three anomalous fibers endowed with
sensory endings were observed in two human EOM spindles of
a 17-year-old boy (authors' unpublished observation, June
1997).
Anomalous libers supplied with sensor)1 terminals were a
major argument for RuskelTs7 hypothesis of spindle reorganization imposed by degeneration. Although they were not observed at 2 years of age, the question remains as to whether
anomalous libers are an expression of a regular turnover of
intrafusal libers or whether they exert a special function of
their own. In a similar rein, evidence for the continuing plasticity of EOM libers during postnatal life was given by mvosin
immunohistochemistry. In EOMs of adult rats^1 *~ and in EOMs
of humansv* numerous morphologically mature extrafusal fibers of the orbital layer coexpress both fast and neonatal, or
neonatal and embryonic, mvosin heavy chain (MHC). In adult
mammalian skeletal muscles, expression of embryonic and
neonatal MHC is confined to the intrafusal fibers.•vl~i(1 Because
many extrafusal fibers in human EOMs imitate a MHC expression only found during development and in intrafusal fibers in
other mammalian skeletal muscle, one might speculate that
this indicates a continuing capacity for differentiation. Uy incorporation of such extrafusal fibers into human EOM spindles,
sensor)' contacts on anomalous fibers (erstwhile extrafusal
fibers) might induce their modification into chain libers in a
manner similar to that in skeletal muscle spindle development. 2 ^ 2 " 50 The caveat here remains, that EOM fibers expressing embryonic and neonatal MHC are otherwise completely
mature muscle libers.
Because of the unique morphology of human EOM spindles, previous reports doubted their capacity to provide useful
proprioceptive information.7 In contrast, the following observations clearly indicate their functional importance. First, a
high and constant number and a characteristic distribution of
spindles within each of the six different EOMs were demonstrated." Second, the morphology of terminals on chain libers
was similar to that of terminals in skeletal muscle spindles.
Third, the characteristic morphology of EOM spindles is also
present at the age of 2 years and must not be interpreted as
age-related degeneration. More likely, their unique morphology represents special functional properties. The lack of nuclear bag fibers in human EOM spindles might indicate that
human EOM spindles have predominantly a static function and
that they monitor the degree of muscle length changes rather
than the contraction velocity.8 Finally, widespread afferent
input from EOMs to the central nervous system suggests propriocepior activity whereby muscle spindles and myotencli-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017
64
Blumcr ct al.
nous cylinders-*' are the only known sources in EOMs. In
particular, afferenfs from EOMs were described to terminate in
the trigeminal ganglion, superior colliculus, the lateral geniculate body, the pulvinar of the thalamus, the tegmentum, the
gigantocellular nucleus, the vestibular nuclei, the nucleus prepositus hypoglossi and the cerebellum, llrodmann areas 17 and
18, the Clare Bishop area, and the frontal cortex.1 In all these
structures involved in vision, oculomotor control, or both,
EOM afferents interact with afferents from the vestibular apparatus. Also, in humans, proprioceptive input from EOMs to the
brain could play an important role in the development of
binocular vision as was demonstrated in cats.1 However, the
exact role of EOM proprioceptors in oculomotor control is still
far from clear. Further research will be needed to elucidate this
complex interaction.
Acknoivledgments
The authors thank Hans Lassmann for helpful discussion and Christianc
Krivanck. Marietta I.ipowcc. and Fakhcr Hen Mustapha for their valuable technical aid.
References
1. Huisseret P. Influence of cxtraocular muscle proprioception on
vision. I'hysiol Rev. 1995:75:323-338.
2. Fiorcntini A. Hcrardi N. Mallei I.. Role of extraocular proprioception in the orienting behavior of cats, lixp Brain Res. 1982;48:
113-120.
3. Gaulliicr GM, Nommay D. Vcrehcr J-L The role of ocular muscle
proprioccption in visual localization of targets. Science. 1990:249:
58-61.
4. Graves A. Trotter Y, Fregnac Y. Role of ocular muscle proprioccption in (he development of depth perception in cats. ./ Neuropbysiol. 1987:58:816-831.
5. Cooper S, Daniel PM. Muscle spindles in human extrinsic eye
muscles, lira in. 1949:72:1-2.
6. Merrillees NCR. Sundcrland S. Hayhovv \V. Neuromuscular spindles in the extraocular muscles in man. Anat Ret: 1950:108:23-30.
7. Ruskell GI.. The line structure of human extraocular muscle spindles and their potential propriocepiive capacity. J Anat. 1989:167:
199-214.
8. Lukas JR, Aigner M, Hlumer R, Hein/I H, Mayr R. Number and
distribution of neuromuscular spindles in human extraocular muscles. Invest 0/)btbal»iol Vis Sci. 1994:35:4317-4327.
9. Lukas JR. Blumcr R, Aigner M. Denk M. Haumganner I, Mayr R.
Proprioception from human extraocular muscles: on the morphology of their neuromuscular spindles. Klin Monatsbl Augenheilkd.
1997:211:183-187.
10. Lukas JR, Hlumer K, Aigner M. Denk M, Mayr R. Effects of eye
muscle proprioceptive activation: morphological particularities of
human extraocular muscle spindles. Graefes Arch Clin lixp Opbibalnwl. 1998:236:238 -239.
I I. Mukuno K, Nomura T. Fine structure of the muscle spindle in the
human extraocular muscles. Ada Soc Opbtbalmol Jap. 1969;73:
21 19-2127.
12. Maier A, De Santis M, lildred F. The occurrence of muscle spindles
in extraocular muscles of various vertebrates../ Morpbol. 1974:
143:397-408.
13- Harker D\V. The structure and innervarion of sheep superior
rectus and Icvaior palpebrae extraocular muscles. II: muscle spin-
dles. Invest Opbtbalmol Vis Sci. 1972:1 1:970-979.
IOVS, January 1999, Vol. 40, No. 1
14. Kubota M. Ultrastructural observations on muscle spindles in extraocular muscles of pig. Anat Anz. 1988; 165:205-228.
15- Hanker HO, Girvin JP. The ulirasiructural features of the mammalian muscle spindle./ iXenropatbol lixp Nenrol. 1971 ;30:155-195.
16. Hr/e/inski DK. Untersuchungen zur Histochemie der Muskelspindeln. II. Mitteilung. Zur Topochemie und Funktion des Spindclraums und der Spindelkapsel. Ada Histocbemica. 1961; 12:277288.
17. Matthews PHC. The structure of the receptors. In: II Davson, ADM
Greenfield, R Whittam, GS Hrindley. eds. Mammalian Muscle
Receptors and Their Central Actions. London: Fdward Arnold Ltd:
1972:24-26.
18. Adal M. The fine srructure of the sensory region of cat muscle
spindles../ Ultrastrud Res. 1969:26:332-354.
19. Hanks RW. Harker D\V. Stacey MJ. A study of mammalian intrafusal
muscle libers using a combined histochemical and ultrastructural
technique../ Anat. 1977; 123:783-796.
20. Ovalle \VK. Fine structure of rat intrafusal muscle fibers: the
equatorial region../ Cell Biol. 1972:52:382-396.
21. Schroder JM. Pathologic der Muskulatur. New York: Springer:
1982:441-452.
22. Howden RUM. Muscle spindles in the human foetus. Ada Biol
Szeged. 1963:9:35-59.
23- Landon DM. The fine structure of the equatorial regions of developing muscle spindles in the rat../ Neurocytol. 1972; 1:189-210.
24. Milburn A. The early development of muscle spindles in rat. / Cell
Sci. 1973; 12:175-195.
25. Hanson.). Lennerstrand G. Nichols KG. The postnatal development
of the inferior oblique muscle of cat. III: fiber size and histochemical properties. Ada Pbysiol Stand. 1980:108:61-71.
26. Nag AC. Cheng M. Differentiation of fiber types in an exlraocular
muscle of the rat../ limbryol lixp Morpbol. 1982;71:171-191.
27. Porter.|D, Haker RS. Prenatal morphogenesis of primate extraocular muscle: neuromuscular junction formation and fiber type differentiation. Invest Opbtbalmol Vis Sci. 1992:33:657-670.
28. Divvan H, Milburn A. The effect of temporary ischacmia on rat
muscle spindles../ limbryol lixp Morpbol. 1986:92:223-254.
29. Milburn A. The effect of the local anaestethic bupivacaine on the
muscle spindle of rat../ Neurocytol. 1976:5:425-446.
30. Zelena J. Morphogeneric inlluence of innervation on the ontogenetic development of muscle spindles../ limbryol lixp Morpbol.
1957:5:283-292.
31. HruecknerJK. Itkis O. Porter JD. Spatial and temporal patterns of
myosin heavy chain expression in developing rat cxtraocular muscle. ./ Muscle Res Cell Motil. 1996; 17:297-312.
32. Wieczorek Dl\ Periasamy M. Huller-Hrovvne GS, Whalen RG.
N'adal-Ginard H. Co-expression of multiple myosin heavy chain
genes, in addition to a tissue-specific one. in extraocular musculature. ./ Cell Biol. 1985; 101:618- 629.
33- Hitiner RH. Lukas JR. Turhani D. Mayr R. Human extraocular
muscles: predominant and persistent expression of embryonic and
neonatal myosin heavy chain in the orbital layer. Verb Anat Ges.
I995;177(suppl):43.
34. Soukop T. Pedrosa-Domellof l:, Thornell LI:. Lxpression of myosin
heavy chain isoforms and myogenesis of intrafusal fibers in rat
muscle spindles. Microsc Res Tecbnol. 1995;3O:39O-4O7.
35. l-rikson PC). Huiler-Hrowne GS. Thornell LF. Imnnmohistochcmical characterization of human masseter muscle spindles. Muscle
Nerve. 1994; 17:31 -4 1.
36. Pedrosa-Domellof F. Gohlsch II, Thornell I.E. Pette D. Llectrophoretically defined myosin heavy chain patterns of single human
muscle spindles. I'liBS Lett. 1993:335:239-242.
37. Steinbach M.|. Proprioceptive knowledge of eye position. Vision
Res. 1987:27:1737-1744.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933210/ on 06/17/2017