Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix wikipedia , lookup
Tissue engineering wikipedia , lookup
Cell growth wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cell culture wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cellular differentiation wikipedia , lookup
Cytokinesis wikipedia , lookup
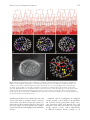
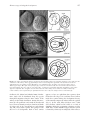
151 What a couple of dimensions can do for you: Comparative developmental studies using 4D microscopy—examples from tardigrade development Andreas Hejnol1,2 and Ralf Schnabel Technische Universität Braunschweig, Spielmannstrasse 7, 38106 Braunschweig, Germany Synopsis The development of an organism consists of processes occurring in space and time. To analyze this 4-dimensional development in embryogenesis, an appropriate method should be chosen. We present here a sophisticated method, 4D microscopy (3D time-lapse microscopy), initially developed to analyze the cell lineage of wild-type and mutant embryos of the nematode Caenorhabditis elegans. Our method records the entire development of an embryo and allows detailed analyses of events such as cleavage, cell migration, cell death (apoptosis), and cell differentiation during development. The 4D microscopy system has 3 main parts: a motorized microscope, trigger software, and a database that facilitates the analysis of recordings. Adopting the 4D microscopy technique for uses beyond the analysis of C. elegans makes it possible to discern the cell lineage of other small embryos. Our method fills a gap in the study of the development of diverse organisms that are impossible to observe with fluorescent labeling techniques using single blastomeres. The use of this technique to investigate the development of organisms such as tardigrades, acoelomorphs, rotifers, and gastrotrichs provides fresh insight into the evolution of developmental processes and the phylogenetic relationships between such taxa. Using tardigrade development as an example, we demonstrate that the use of 4D microscopy can reveal new characters and corroborate or disapprove old characters. We discuss the results in the light of recent phylogenetic hypotheses regarding the Arthropoda and their probable sister group, the Cycloneuralia, which together form the Ecdysozoa. Introduction Resolution of metazoan phylogenetic relationships is one of the most long standing and fascinating goals of biology, in which modern techniques from the fields of molecular biology, bioinformatics, paleontology, developmental biology, and morphology are integrated. Consistent technological advances, driven primarily by medical applications in model organisms, can also be fruitful when applied to the study of other organisms. Here we present a new form of microscopy, termed 4D microscopy (3D time-lapse microscopy), that was originally developed to investigate the molecular basis of the cell lineage of the nematode Caenorhabditis elegans (Schnabel and others 1997). Progress in analog and computational imaging technologies has made it possible to adapt this method to investigate other embryos. Questions raised by the publication of alternative phylogenetic hypotheses concerning the protostomian clades guided our choice of key organisms whose embryos are suitable for analysis using the 4D microscope system. We present here the phylogenetic implications of our study of the development of the eutardigrade Thulinia stephaniae (Hejnol and Schnabel 2005) in the light of recently published phylogenetic hypotheses. Not only do we either corroborate or reject prior assumptions concerning tardigrade development, but we also identify new characters (for example, cleavage pattern) that could be used for morphological data matrices in phylogenetic analysis. Our results contribute to the reconstruction of the arthropod and the ecdysozoan ground pattern. The 4D microscopy system The 4D microscopy system, first introduced as a multiple focal plane time-lapse recording system by Minden and colleagues (1989) and Hird and White (1993), was designed to analyze the development of living early embryos. The use of high-resolution enhanced Nomarski optics facilitates the detection of single cells of the embryo at different focus levels (Thomas and others 1996; Schnabel and others 1997). From the symposium “The New Microscopy: Toward a Phylogenetic Synthesis” presented at the annual meeting of the Society for Integrative and Comparative Biology, January 4–8, 2005, at San Diego, California. 1 E-mail: [email protected] 2 Present address: Kewalo Marine Laboratory, University of Hawaii at Manoa, 41 Ahui Street, Honolulu, HI 96813, USA. Integrative and Comparative Biology, volume 46, number 2, pp. 151–161 doi:10.1093/icb/icj012 Advance Access publication February 16, 2006 Ó The Society for Integrative and Comparative Biology 2006. All rights reserved. For permissions, please email: journals.permissions@ oxfordjournals.org. 152 A. Hejnol and R. Schnabel Fig. 1 The 4D microscopy system. (A) Motorized microscope used for this study. The microscope is connected to a Hamamatsu analog camera. The pictures are digitized using a frame grabber in the connected computer. The software on the computer triggers the motor of the microscope and the collection of the single pictures. (B) Screenshot of the trigger software. In the right window the photograph collected by the camera is shown. All major modification buttons are visible on the screen and can be easily used during the recording. (C) Screenshot of the SIMI BioCell software for the analysis of the recordings. The left-hand window shows the cell lineage of the embryo; the central window shows the 3D reconstruction of the embryo. The right-hand window shows the picture of the corresponding level. Other small windows facilitate navigation of the lineage tree and show information about single cells. To handle the amount of data produced by a detailed recording of a complete embryogenesis and to facilitate the tracing of cell lineage, the software SIMI BioCell was developed (Schnabel and others 1997). Modifications to the system have allowed it to be successfully applied to other embryos, as demonstrated by investigations of Drosophila melanogaster (Urbach and others 2003), an isopod crustacean (Hejnol 2002; Dohle and others 2004), other nematode embryos (Dolinski and others 1998; Houthoofd and others 2003) and the tardigrade embryo presented here (Hejnol and Schnabel 2005). The system has 3 parts: a motorized microscope, control software for recording, and analysis software (Fig. 1). The microscope is a modified Zeiss compound microscope with a motorized light shutter and focus (Fig. 1A). Photographs are taken using a high-resolution analog camera, digitized with a frame grabber, and automatically saved onto the computer hard drive in a lossless wavelet compressed format. To ensure constant temperature during recording, the microscope is equipped with a thermostat set to a range of 10–37 C. The microscope software was programmed in Cþþ by A.-K. Schulz and R.S. and controls the whole capturing process from opening the light shutter, to capturing individual pictures step by step through the z-levels, to saving the z-stack onto the hard disk, to closing the light shutter (Fig. 1B). This process is repeated at defined time intervals during the entire development of the embryo. Parameters can be changed during the recording to correct for possible sinking of the embryo within the z-stack, to increase 153 4D microscopy of tardigrade development the number of optical sections when cells become smaller, and to increase time intervals when cell cycles are prolonged as development proceeds. The hardware and software are able to capture z-stacks of up to 70 levels every 40 s to ensure that the development of fast developing embryos with asynchronous cleavage patterns can be recorded. The tardigrade embryo was recorded with 45 optical sections every 45 s at a temperature of 24 C. To handle the large number of recorded digital pictures (for example, 1 recording of the tardigrade embryo consists of 140,000 individual images) we used the database SIMI BioCell, software specifically programmed for this type of cell lineage analysis (Fig. 1C). SIMI BioCell allows easy visualization of single pictures using the computer keyboard (up/down cursors to move through the z-stack; left/right cursors to move forward and backward in time). Cell nuclei are marked by clicking the mouse at appropriate time intervals, which saves their coordinates and the time to a database. By following and marking cells before and after cell divisions, a cell lineage tree is built in which each cell can be followed to its final cell fate. The software is able to represent the data as a cell lineage tree or as a 3D representation in which the position of every cell (nucleus) is shown as a sphere (Fig. 1C). The spheres can be colored to demonstrate cell clones and their localization. SIMI BioCell also includes several functions that facilitate the tracing of cells in the embryo. Because the resolution of Nomarski optics improves at high magnifications, 4D microscopy is ideal for recording small embryos (<60 mm) with hard eggshells, which are otherwise inaccessible via injection of blastomeres. A future goal is to combine 4D microscopy with fluorescent gene expression reporters and to visualize gene expression patterns over time at a single cell resolution. A 4D microscope system with 3 channels has been developed for this purpose. To exemplify the power of the system for comparative studies we discuss our results on tardigrade development in the light of the currently conflicting phylogenetic hypotheses. Tardigrade development: bringing light into a obscure field Tardigrades—also known as water bears—are tiny protostomes that live in marine and freshwater environments. Historically, their development has received little attention because the overwhelming interest has been in the developmental mechanisms in model organisms. With the rebirth, as “evo-devo,” of the field of comparative developmental biology and major rearrangements in the protostome phylogeny, some of the minor phyla are attracting renewed attention. One of these neglected taxa is the Tardigrada, which have gained attention as a result of their intermediate position between Euarthropoda and groups of the former Nemathelminthes in the Ecdysozoa hypothesis (Gould 1995; Aguinaldo and others 1997; Kinchin 2000; Goldstein and Blaxter 2002). Hence, a reinvestigation of the development of tardigrades was advisable given the large gaps in our knowledge about their development. After an initial investigation by Kaufmann (1851), the majority of studies on tardigrade development were carried out in the early 20th century (von Erlanger 1895; von Wenck 1914; Marcus 1928, 1929). These studies describe cleavage, gastrulation, and germ layer formation of eutardigrade species and form the basis for the descriptions of tardigrade development in recent textbooks and of characteristics in current morphological data matrices. More recently, Eibye-Jacobsen (1997) added descriptions of the development of heterotardigrade embryos based on transmission electron microscopy (TEM) studies. These descriptions often conflict with the previous studies regarding the cleavage pattern and gastrulation, raising certain questions: Do tardigrade embryos show traits of spiral cleavage? Do they show a stereotypical cleavage pattern? When are the fates of the cells determined? Is the mesoderm really formed as outpouchings from the gut (enterocoely)? Eibye-Jacobsen (1997) cast doubt on the descriptions by Marcus (1928, 1929) and von Erlanger (1895) of the mesoderm being formed by enterocoely, and a careful comparison of the 2 publications by Marcus further confuses the discussion. Marcus (1928) stated that he could not detect cell borders after gastrulation in his histological slices although he included them in his drawings. One year later this statement is missing (Marcus 1929). R. M. Kristensen looked again at Marcus’ original sections, which are archived in the Smithsonian Institute, and confirmed that it is not possible to detect cell borders and furthermore that the sections are too thick (5–8 mm) to reconstruct enterocoely (personal communication from R. M. Kristensen; unreferenced). A reinvestigation of tardigrade development using modern techniques is necessary to resolve these concerns. The phylogenetic position of the tardigrades Tardigrades are generally associated with the Euarthropoda (chelicerates, myriapods, crustaceans, and insects) based on morphological data and 154 A. Hejnol and R. Schnabel Fig. 2 Embryonic cleavage patterns mapped onto alternative phylogenetic trees. (A) Evolution of cleavage patterns based on the Articulata hypothesis. Molluscs and annelids show spiral cleavage. Consequently, in the Arthropoda the spiral cleavage is plesiomorphic and was modified to a radial or syncytial cleavage pattern. (B) The alternative Ecdysozoa hypothesis, which postulates a sister relationship of Arthropoda and Cycloneuralia, predicts a radial, indeterminate cleavage in the ground pattern of this clade, since the Nematomorpha and Priapulida show a radial irregular cleavage pattern. Nematode development shows variable cleavage patterns. However, an outgroup comparison with the development of nematomorphs suggests that the stereotypical cell lineage, for example of C. elegans, is derived in the nematodes and that an indeterminate cleavage is thus part of the ground pattern of nematodes. This suggests that an indeterminate cleavage pattern is plesiomorphic for the Arthropoda and that the stereotypical cell lineages observed in some arthropods are a derived characteristic in the Arthropoda as in nematodes. Therefore, we propose that an indeterminate cleavage is part of the ground pattern in the Ecdysozoa. molecular data (Garey and others 1996, 1999; Giribet and others 1996; Garey 2001; Peterson and Eernisse 2001; Mallatt and Winchell 2002; Mallatt and others 2004). A 2005 molecular phylogenetic analysis (Philippe and others 2005) placed the tardigrades as a sister group to the nematodes. However, as there is no significant data to the contrary, we assume the position of the tardigrades as sister to arthropods (Fig. 2B). Thus, tardigrades share a common ancestor with the euarthropods, and we will discuss our results regarding the ground pattern of the Arthropoda. To specify the plesiomorphic characteristics, it is necessary to include the sister group of the arthropods in the discussion. As new phylogenies based on molecular data favor the Cycloneuralia (Priapulida, Loricifera, Kinorhyncha, Nematomorpha, and Nematoda) as the sister group (Aguinaldo and others 1997; Giribet and others 2000; Garey 2001; Giribet 2003; Philippe and others 2005), rather than the Annelida (Fig. 2), we will discuss some characteristics as possible plesiomorphies for the Arthropoda as well as the Ecdysozoa. [The morphological data matrices of these analyses that support the Ecdysozoa (Zrzavy and others 1998; Peterson and Eernisse 2001; Zrzavy 2001, 2003; Eernisse and Peterson 2004) have been criticized by Jenner and Scholtz (2005).] Results and discussion Axis determination and cleavage pattern We investigated the cell lineage of the eutardigrade T. stephaniae using the 4D microscopy system and a subset of developmental stages of the heterotardigrade Echiniscoides sigismundii. Cleavage is total and equal in both embryos. The cleavage program of T. stephaniae is irregular, and early blastomeres show an unexpected potential for regulation when early cells are ablated (Hejnol and Schnabel 2005). It was not possible to identify corresponding blastomeres between embryos owing to their equal size, the lack of morphological characteristics, and the variable spindle directions (Fig. 3D–F). The timing of cell divisions also differed among individual embryos (Fig. 3A–C). The embryo appears to lack a predetermined egg axis, and the localization of the polar body does not correspond to any of the future dorsal/ventral or anterior/posterior axes (Hejnol and Schnabel 2005). The observed absence of predetermined cytoplasm is supported by cell ablation experiments in which the embryo was able to form all main axes after 1 cell of the 2-cell and 2 cells of the 4-cell stage were ablated (Hejnol and Schnabel 2005). Related euarthropods show a gradient of cleavage types ranging from total to syncytial cleavage, and 4D microscopy of tardigrade development 155 Fig. 3 Cleavage and gastrulation in the eutardigrade T. stephaniae. (A–C) Cell lineages and cell pattern of 3 different embryos of the tardigrade T. stephaniae (126 cell stage). The lineage trees show different cell division timings in all 3 embryos from early on. (D–F) Arrangements of the descendants of 2 corresponding blastomeres (determined by the future egg axes) differ in each embryo (dark blue versus pink). The 2 bright yellow cells show the 2 primordial germ cells. (G) Gastrulation of a tardigrade embryo: ventral view, anterior is left. The blastopore is located at the future position of the mouth. (H) Arrangement of the germ layer precursors before cell migration at the site of gastrulation shown with the 3D reconstruction of SIMI BioCell. The germ cells are the first to migrate into the embryo, followed by the endodermal and mesodermal precursors. including intermediate forms, which begin with total cleavage and switch to syncytial cleavage (Anderson 1973; Scholtz 1997). Both cleavage type extremes are found scattered in the arthropod lineages, and related taxa show opposite cleavage patterns (for example, isopods with syncytial and amphipods with holoblastic cleavage). Compared with the euarthropods, the tardigrade cleavage program is most similar to the holoblastic and irregular cleaving pycnogonids (Dogiel 1913), some crustaceans (Fuchs 1914; Benesch 1969), and myriapods (Tiegs 1940, 1947; Dohle 1964). Syncytial cleavage appears to have evolved independently in different arthropod lineages, perhaps as an 156 adaptation to a terrestrial life cycle. As an outgroup comparison would help to determine the cleavage type of the stem species of the Arthropoda, tardigrades may reflect the ancestral type of cleavage in the arthropod clade. In light of the Ecdysozoa hypothesis, irregular and indeterminate tardigrade cleavage would be a plesiomorphic condition (Fig. 2B). The clades that are regarded as the new adelphotaxon of the Arthropoda, either the whole Cycloneuralia or some members, show holoblastic cleavage. In addition, their cleavage pattern appears to be irregular based on the study of certain nematodes (Malakhov and Spiridonov 1984; Voronov and Panchin 1998; Voronov 1999; Schierenberg 2005) and priapulids (Zhinkin 1949; Zhinkin and Korsakova 1953). As annelids are the sister group to Arthropoda according to the Articulata hypothesis (Fig. 2A), the cleavage type in the arthropods should be derived from a spiral cleaving ancestor (Scholtz 1997; Nielsen 2001). No spiral cleaving arthropod embryo has been properly described, and Anderson’s wideranging description (1969) of the cirripede cleavage as spiral is based on dubious homologization of single cells and badly needs reinvestigation using modern methods (Zilch 1979). Under the Ecdysozoa hypothesis, holoblastic irregular cleavage appears to be the plesiomorphic condition for arthropods as well as for all of the Ecdysozoa (Fig. 2B). Which molecular mechanisms regulate cell fate and axis determination in the tardigrade embryo remains enigmatic, as no known mechanism from protostome model organisms can explain this ancestral type of development. Gastrulation Gastrulation in T. stephaniae starts with the immigration of cells at the position of the future oral opening (Hejnol and Schnabel 2005). The embryo does not form a clear blastocoel. Simultaneously, some cells migrate through the blastopore and other cells sink temporarily into the embryo at a nearby area that later in development gives rise to the proctodaeum of the adult (Fig. 3G). In T. stephaniae, the first evidence of the anterior/posterior and dorsal/ventral axes of the embryo is recognizable at gastrulation with the blastopore being anterior and ventral (Hejnol and Schnabel 2005). In all embryos recorded, the germ cells were the first to migrate, followed by the endoderm precursors and mesoderm. During migration these cells are indistinguishable, with only future cell fate in terms of their position and differentiation allowing us to reconstruct their identity during this stage. The 2 germ cells migrate into the gonad and the mesodermal cells form the somites A. Hejnol and R. Schnabel and later the muscles of the limbs. The arrangement of the cells prior to migration is shown in Figure 3H, in which 2 germ cells are surrounded by the endoderm and mesoderm precursors. We were unable to determine whether there is a stereotypical pattern in the arrangement of the mesodermal and endodermal precursors owing to the low number of embryos in which the cell lineage was traced (n ¼ 3). Which mechanism induces the fate of the germ cells or the fate of the germ layer precursors remains unknown, but the arrangement of these cells at the blastopore is reminiscent of some total cleaving crustaceans (Kühn 1913; Fuchs 1914; Benesch 1969; Gerberding and others 2002; Gerberding and Patel 2004) and thus suggests the presence of a common mechanism. Owing to the variation in gastrulation across the arthropods (Weygoldt 1979) it is difficult to determine whether gastrulation in tardigrades reflects an ancestral type for the arthropods. A comparison with the putative sister group of arthropods, the Cycloneuralia, is rather uninformative because of the lack of thorough investigations of the gastrulation of loriciferans, kinorhynchs, and priapulids. The descriptions of gastrulation in nematodes demonstrate the variety present in this taxon (Sulston and others 1983; Voronov and Panchin 1998; Schierenberg 2005) and the description of a nematomorph gastrulation (Inoue 1958) is completely incongruent with other cycloneuralians, in that the mouth is formed at a site other than the blastopore. Mesoderm development The development of the third germ layer of tardigrades has been controversial from the beginning of tardigrade investigation. Von Erlanger (1895) stated that the mesoderm is formed by outpouching of the gut. His illustrations clearly show coelomic hollows and a gut lumen in the developing embryo. We did not observe these lumens in living embryos of either the heterotardigrade E. sigismundii (Fig. 4A) or the eutardigrade T. stephaniae (Fig. 4B and C). Enterocoelic formation was corroborated by Marcus (1928, 1929). More recently, Eibye-Jacobson (1997) expressed doubts concerning the formation of the mesoderm by enterocoely in trardigrades. Tracing back the cell lineage of single cells of the mesodermal somites of T. stephaniae shows that their origin is not the endodermal gut; rather they develop from mesodermal precursors that migrate through the blastopore into the embryo (Hejnol and Schnabel 2005). These mesoderm precursor cells form mesodermal cords ventrolateral to the gut (Fig. 4B). The precursor cells do not form a clear growth zone nor 4D microscopy of tardigrade development 157 Fig. 4 Later stages of tardigrade embryos showing the mesodermal somites. The schematic drawings on the right show the orientation of the embryo in the egg. (A) Embryos of the heterotardigrade E. sigismundii. The black arrows show the epithelial border surrounding somites. (B) Embryo of T. stephaniae before elongation. The spheres indicate cells of different germ layers. One part of the pharynx (ph) consists of endodermal cells. The mesodermal cells form lateral mesodermal bands. The inner cells are surrounded by a 1-cell layered ectodermal epithelium. (C) Embryo of the eutardigrade T. stephaniae after elongation: view from the left side of the embryo. Four somites are visible, indicated by roman numbers; each is separated by an epithelium from the other cells. The embryo forms a ventral fold (vf). teloblastic cells (Hejnol and Schnabel 2005). Furthermore, single cells are distributed along the egg axis throughout migration along the inner surface of the embryo to their final destinations. During this movement, the cells proliferate and extend the mesodermal layer. The mesodermal germ layer forms lateral bands on either side of the entodermal gut and pharynx (Fig. 4B). These bands separate into groups of cells after 22 h of development. Each of these somites appears to have an epithelium that separates them from other cells (Fig. 4). We could not detect a hollow inside these somites during the development because of the naturally compressed embryo inside the eggshell (Fig. 4). In her work, Eibye-Jacobsen (1997) could detect hollows, which become visible as a result of shrinking during the preparation technique used for TEM. These results combined suggest that the coelom is formed by schizocoely in tardigrades. This reflects a 158 A. Hejnol and R. Schnabel Table 1 Morphological characters changed in the data matrix of Eernisse and Peterson (2004) No. Character Modification 17 Position of polar bodies Added characteristic state 4 (irregular position) 24 One axis specified during oogenesis “?” to “0” (absent) 28 Stereotypical cleavage pattern “?” to “0” 29 Spiral cleavage “?” to “0” 32 Gastrulation “?” to “1” (present) 33 Blastopore associated with larval/adult mouth “?” to “1” 35 Endomesodermal muscle cells “?” to “1” 36 Endomesoderm derived from gut “?” to “0” 37 Ectomesenchyme “?” to “0” 38 4d endomesoderm “?” to “0” 39 Mesodermal bands from 4d “?” to “0” 40 Lateral coelom derived from mesodermal bands “0” to “1” (also corrected for arthropods and onychophorans) 42 Teloblastic growth “?” to “0” 43 Somatoblast “?” to “0” Tetrahedral 4-cell stage “?” to “1” 157 well-known phenomenom in arthropods, including the onychophorans (Anderson 1973; Dohle 1979). Later in development, these somites disappear and some of the cells migrate into the limb buds to form the muscles. The other cells could not be traced further, but they may form the “mixocoel,” which is common to all euarthropod taxa and the onychophorans (Weygoldt 1986; Nielsen 2001; Scholtz 2002; Mayer and others 2004). The establishment of mesodermal bands that separate into segmental, paired somites to later form coelomic cavities by schizocoely is a mechanism shared between annelids and arthropods (Weygoldt 1963; Anderson 1973; Dohle 1979) and has been used to unite annelids and arthropods in defense of the Articulata hypothesis (Scholtz 2002, 2003). In nematodes and nematomorphs, mesodermal bands are also present (Inoue 1958; Schierenberg 2005), but they do not form segmental somites. Segmentation and posterior growth zone Tardigrades have 4 segmental leg pairs and a brain consisting of 3 parts (Dewel and Dewel 1996), although we could not corroborate the presence of a tripartite brain in our study. Our study of the development of the eutardigrade T. stephaniae shows no sequential development of the 4 trunk segments, including the somites and limb buds (Fig. 4A and C). Teloblasts in the ectoderm or mesoderm are not present in the embryo and a posterior growth zone is also absent (Hejnol and Schnabel 2005). A common mechanism to elongate the body during development in annelids and arthropods is the sequential addition of segments from a posterior growth zone. The absence of a growth zone in tardigrades may be interpreted as a derived state resulting from the small size of the embryo and the low number of segments. Some authors regard a teloblastic growth zone as an apomorphy supporting a close relationship between the Annelida and Arthropoda (Anderson 1973; Ax 1999; Nielsen 2003). However, recent evidence rejects teloblasts in the ground pattern of the Arthropoda (Dohle and Scholtz 1988; Scholtz 2002). The description of the presence of ectoteloblasts in annelids (Anderson 1973) is only true for the clitellates (Dohle 1999). Our cell lineage studies for the polychaete Trilobodrilus axi show that the cells that form the ectoteloblasts in clitellates do not form ectoteloblasts in polychaetes and the “mesoteloblast” derived from the 4d cell does not divide in a teloblast fashion (unpublished data, A.H.). If the Ecdysozoa hypothesis is correct and no member of the Cycloneuralia displays an embryological posterior growth zone, then it has evolved in the arthropods convergently. Further investigations of priapulid, kinorhynch, and loriciferan development are needed before an understanding of the evolution of segmentation in the protostomes will be complete. Phylogenetic analysis Our investigations of the development of the eutardigrade T. stephaniae yielded new insights and useful characters for the reconstruction of phylogenetic 4D microscopy of tardigrade development trees. To determine the extent to which the new findings corroborate or weaken the position of the Tardigrada, we modified the morphological data matrix of Eernisse and Peterson (2004) and recalculated the Bootstrap and Bremer support with Paup* (Swofford 2002) of a parsimony analysis. Of 166 morphological characters, 15 were changed in the matrix according to Table 1, and both trees were recalculated with 2000 replicants. The topology of the relationship of Tardigrada to Onychophora and Arthropoda was not changed, but the Bootstrap values and Bremer indices differed. The node separating Tardigrada from Onychophora/Arthropoda is supported in both trees with a Bremer index of 4, and the Bootstrap value varies marginally between the original tree (87) and the modified one (86). Interestingly, the sister group relationship between onychophorans and arthropods receives a Bootstrap value of 83 and Bremer index of 3 in the original tree and is less supported after introducing our modifications (Bootstrap 69, Bremer index 2). Although support for a sister group relationship of the tardigrades to the Onychophora/ Arthropoda remains relatively high in both trees, the introduction of our findings to the data matrix of Eernisse and Peterson (2004) could not help to clarify the phylogenetic relationships between these taxa. Acknowledgments We thank Reinhardt M. Kristensen for species determination and his help and support in collecting E. sigismundii. The field collection trip for E. sigismundii to Denmark for A.H. was financed by the COBICE program of the EU. We thank Amy Maxmen, Kevin Pang, and 3 anonymous referees for critically reading and improving the manuscript and Kevin Peterson for sending us the nexus file. References Aguinaldo AM, Turbeville JM, Linford LS, Rivera MC, Garey JR, Raff RA, Lake JA. 1997. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387:489–93. Anderson DT. 1969. On the embryology of the cirripede crustaceans Tetraclita rosea (Krauss), Tetraclita purpurascens (Wood), Chthalamus antennatus (Darwin) and Chamaesipho columna (Spengler) and some considerations of crustacean phylogenetic relationships. Philos Trans R Soc B 256:183–235. Anderson DT. 1973. Embryology and phylogeny in annelids and arthropods. Oxford: Pergamon Press. Ax P. 1999. Das System der Metazoa II. Stuttgart: Gustav Fischer. Benesch R. 1969. Zur Ontogenie und Morphologie von Artemia salina L. Zool Jb Anat 86:307–458. Dewel RA, Dewel WC. 1996. The brain of Echiniscus viridissimus Peterfi 1956 (Heterotardigrada): a key to understanding the 159 phylogenetic position of tardigrades and the evolution of the arthropod head. Zool J Linn Soc 116:35–49. Dogiel V. 1913. Embryologische Studien an Pantopoden. Z Wiss Zool 107:575–741, plates 17–22. Dohle W. 1964. Die Embryonalentwicklung von Glomeris marginata (Villers) im Vergleich zur Entwicklung anderer Diplopoden. Zool Jb Anat 81:241–310. Dohle W. 1979. Vergleichende Entwicklungsgeschichte des Mesoderms bei Articulaten. Fortschr Zool Syst Evolutionsforsch 1:120–40. Dohle W. 1999. The ancestral cleavage pattern of the clitellates and its phylogentic deviations. Hydrobiologia 402:267–83. Dohle W, Gerberding M, Hejnol A, Scholtz G. 2004. Cell lineage, segment differentiation and gene expression in crustaceans. In: Scholtz G, editor. Evolutionary developmental biology of Crustacea. Lisse: A.A. Balkema. p 95–133. Dohle W, Scholtz G. 1988. Clonal analysis of the crustacean segment: the discordance between genealogical and segmental borders. Development 104 (Suppl):147–60. Dolinski C, Borgonie G, Schnabel R, Baldwin JG. 1998. Buccal capsule development as a consideration for phylogenetic analysis of Rhabditida (Nemata). Dev Genes Evol 208:495–503. Eernisse D, Peterson K. 2004. The history of animals. In: Cracraft J, Donoghue M, editors. Assembling the tree of life. New York: Oxford University Press. p 197–208. Eibye-Jacobsen J. 1997. New observations on the embryology of the Tardigrada. Zool Anz 235:201–16. Fuchs F. 1914. Die Keimblätterentwicklung von Cyclops viridis Jurine. Zool Jb Anat 38:103–56. Garey JR. 2001. Ecdysozoa: the relationship between Cyloneuralia and Panarthropoda. Zool Anz 240:321–30. Garey JR, Krotec M, Nelson DR, Brooks J. 1996. Molecular analysis supports a tardigrade-arthropod association. Invertebr Biol 115:115–21. Garey JR, Nelson DR, Mackey LY, Li J. 1999. Tardigrade phylogeny: congruency of morphological and molecular evidence. Zool Anz 238:205–10. Gerberding M, Browne WE, Patel NH. 2002. Cell lineage analysis of the amphipod crustacean Parhyale hawaiensis reveals an early restriction of cell fates. Development 129:5789–801. Gerberding M, Patel NH. 2004. Gastrulation in crustaceans. In: Stern C, editor. Gastrulation. Cold Spring Harbor, NY: CSHL Press. p 79–89. Giribet G. 2003. Molecules, development and fossils in the study of metazoan evolution; Articulata versus Ecdysozoa revisited. Zoology 106:303–26. Giribet G, Carranza S, Baguna J, Riutort M, Ribera C. 1996. First molecular evidence for the existence of a Tardigrada Arthropoda clade. Mol Biol Evol 13:76–84. Giribet G, Distel DL, Polz M, Sterrer W, Wheeler WC. 2000. Triploblastic relationships with emphasis on the acoelomates and the position of Gnathostomulida, Cycliophora, Plathelminthes, and Chaetognatha: a combined approach of 18S rDNA sequences and morphology. Syst Biol 49:539–62. 160 Goldstein B, Blaxter M. 2002. Tardigrades. Curr Biol 12:R475. Gould SJ. 1995. Of tongue worms, velvet worms and water bears. Nat Hist 95:6–15. Hejnol A. 2002. Der postnaupliale Keimstreif von Porcellio scaber und Orchestia cavimana (Crustacea, Peracarida): Zellinie, Genexpression und Beginn der Morphogenese [thesis]. Berlin: HU Berlin. Hejnol A, Schnabel R. 2005. The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. Development 132:1349–61. Hird SN, White JG. 1993. Cortical and cytoplasmic flow polarity in early embryonic cells of Caenorhabditis elegans. J Cell Biol 121:1343–55. Houthoofd W, Jacobsen K, Mertens C, Vangestel S, Coomans A, Borgonie G. 2003. Embryonic cell lineage of the marine nematode Pellioditis marina. Dev Biol 258:57–69. Inoue I. 1958. Studies on the life history of Chordodes japonensis, a species of Gordiacea. I, The development and structure of the larva. Jap J Zool 12:203–18. Jenner RA, Scholtz G. 2005. Playing another round of metazoan phylogenetics: historical epistemology, sensitivity analysis, and the position of Arthropoda within the Metazoa on the basis of morphology. In: Koenemann S, Jenner RA, editors. Crustacea and arthropod relationships. Lisse: A.A. Balkema. p 355–86. Kaufmann J. 1851. Ueber die Entwicklung und systematische Stellung der Tardigraden. Z Wiss Zool 3:220–32. Kinchin IM. 2000. On the significance of a “minor” phylum (the Tardigrada) in the context of a constructivist view of knowledge. Perspect Biol Med 43:243–51. Kühn A. 1913. Die Sondierung der Keimesbezirke in der Entwicklung der Sommereier von Polyphemus. Zool Jb Anat 35: 243–340 Malakhov VV, Spiridonov SE. 1984. The embryogenesis of Gordius sp. from Turkmenia, with special reference to the position of the Nematomorpha in the animal kingdom. Zool Zh 63:1285–97. Mallatt JM, Garey JR, Shultz JW. 2004. Ecdysozoan phylogeny and Bayesian inference: First use of nearly complete 28S and 18S rRNA gene sequences to classify the arthropods and their kin. Mol Phylogenet Evol 31:178–91. Mallatt JM, Winchell CJ. 2002. Testing the new animal phylogeny: first use of combined large-subunit and small-subunit rRNA gene sequences to classify the protostomes. Mol Biol Evol 19:289–301. Marcus E. 1928. Zur Embryologie der Tardigraden. Verh Deutsch Zool Ges 32:134–46. Marcus E. 1929. Zur Embryologie der Tardigraden. Zool Jb Anat 50:334–79. Mayer G, Ruhberg H, Bartolomaeus T. 2004. When an epithelium ceases to exist: an ultrastructural study on the fate of the embryonic coelom in Epiperipatus biolleyi (Onychophora, Peripatidae). Acta Zool 85:163–70. A. Hejnol and R. Schnabel Minden JS, Agard DA, Sedat JW, Alberts BM. 1989. Direct cell lineage analysis in Drosophila melanogaster by time-lapse, three-dimensional optical microscopy of living embryos. J Cell Biol 109:505–16. Nielsen C. 2001. Animal evolution. New York: Oxford University Press. Nielsen C. 2003. Proposing a solution of the ArticulataEcdysozoa controversy. Zool Scr 32:475–83. Peterson KJ, Eernisse DJ. 2001. Animal phylogeny and the ancestry of bilaterians: inferences from morphology and 18S rDNA gene sequences. Evol Dev 3:170–205. Philippe H, Lartillot N, Brinkmann H. 2005. Multigene analyses of bilaterian animals corroborate the monophyly of Ecdysozoa, Lophotrochozoa and Protostomia. Mol Biol Evol 22:1246–53. Schierenberg E. 2005. Unusual cleavage and gastrulation in a freshwater nematode: developmental and phylogenetic implications. Dev Genes Evol 215:103–8. Schnabel R, Hutter H, Moerman D, Schnabel H. 1997. Assessing normal embryogenesis in Caenorhabditis elegans using a 4D microscope: variability of development and regional specification. Dev Biol 184:234–65. Scholtz G. 1997. Cleavage, germ band formation and head segmentation: the ground pattern of the Euarthropoda. In: Fortey RA, Thomas RH, editors. Arthropod relationships. London: Chapman & Hall. p 317–332. Scholtz G. 2002. The Articulata hypothesis—or what is a segment? Org Divers Evol 2:197–215. Scholtz G. 2003. Is the taxon Articulata obsolete? Arguments in favour of a close relationship between annelids and arthropods. In: Legakis A, Sfenthourakis S, Polymeni R, ThessalouLegaki M, editors. The new panorama of animal evolution. 18th International Congress of Zoology Sofia: Pensoft. p 489–501. Sulston JE, Schierenberg E, White JG, Thomson JN. 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev Biol 100:64–119. Swofford D. 2002. Paup? phylogenetic analysis using parsimony (and other methods), ver. 4.0b10 for Unix. Sunderland, MA: Sinauer Associates. Thomas C, DeVries P, Hardin J, White J. 1996. Fourdimensional imaging: computer visualization of 3D movements in living specimens. Science 273:603–7. Tiegs OW. 1940. The embryology and affinities of the Symphyla, based on a study of Hanseniella agilis. Q J Micros Sci 82:1–225. Tiegs OW. 1947. The development and affinities of the Pauropoda, based on a study of Pauropus sylvaticus. Q J Micros Sci 88:165–267, 275–336. Urbach R, Schnabel R, Technau GM. 2003. The pattern of neuroblast formation, mitotic domains and proneural gene expression during early brain development in Drosophila. Development 130:3589–606. von Erlanger R. 1895. Beiträge zur Morphologie der Tardigraden. I, Zur Embryologie eines Tardigraden: Macrobiotus macronyx Dujardin. Morph Jb 22:491–513. 4D microscopy of tardigrade development von Wenck W. 1914. Entwicklungsgeschichtliche Untersuchungen an Tardigraden (Macrobiotus lacustris Duj.). Zool Jb Anat 37:465–514. Voronov DA. 1999. The embryonic development of Pontonema vulgare (Enoplida: Onycholaimidea) with a discussion of nematode phylogeny. Russ J Nematol 7:105–14. 161 Zhinkin L. 1949. Early stages in the development of Priapulus caudatus [in Russian]. Dokl Akad Nauk SSSR 65:409–12. Zhinkin L, Korsakova G. 1953. Early stages in the development of Halicryptus spinulosus [in Russian]. Dokl Akad Nauk SSSR 88:571–3. Voronov DA, Panchin YV. 1998. Cell lineage in marine nematode Enoplus brevis. Development 125:143–50. Weygoldt P. 1963. Grundorganisation und Primitiventwicklung bei Articulaten. Zool Anz 171:363–76. Weygoldt P. 1979. Gastrulation in the Arthropoda? Fortschr Zool Syst Evolutionsforsch 1:73–81. Zilch R. 1979. Cell lineage in arthropods? Fortschr Zool Syst Evolutionsforsch 1:19–41. Zrzavy J. 2001. Myzostomida are not annelids: molecular and morphological support for a clade of animals with anterior sperm flagella. Cladistics 17:170–98. Zrzavy J. 2003. Gastrotricha and metazoan phylogeny. Zool Scr 32:61–81. Weygoldt P. 1986. Arthropod interrelationships: the phylogenetic-systematic approach. Z Zool Syst Evolutionsforsch 24:19–35. Zrzavy J, Mihulka S, Kepka P, Bezdek A, Tietz D. 1998. Phylogeny of the Metazoa based on morphological and 18S ribosomal DNA evidence. Cladistics 14:249–85.