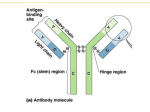
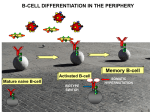
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
Molecular mimicry wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Adaptive immune system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
ª Society forby reserved. © The The Japanese Japanese Published Immunology. 2012. All rights Oxford University Press onreserved. behalf of For permissions, permissions, please please e-mail: [email protected] [email protected] For e-mail: The Japanese Society for Immunology 2009. International 1 of 6 7, International Immunology Immunology,2012; Vol. 22, 24, No. Immunology, Vol. No. 1, pp. pp. 403–408 7–12 doi:10.1093/intimm/dxs050 doi:10.1093/intimm/dxs050 doi:10.1093/intimm/dxp112 Advance Access Access publication publication 23 26 November March 20122009 Advance IL-21 androutes T follicular helper cells Multiple to B-cell memory Rosanne Spolski and Warren J. Leonard Kim L. Good-Jacobson1,2 and David M. Tarlinton1,2 Laboratory 1 of Molecular Immunology, National Heart, Lung, and Blood Institute, Building 10, Room 7B05, Bethesda, Immunology Division, 20892-1674, USA Walter and Eliza Hall Institute of Medical Research, Parkville, Victoria 3052, Australia Department of Medical Biology, University of Melbourne, Parkville, Victoria 3010, Australia MD 2 Received 23 January 2012, accepted 4 March 2012 Abstract Abstract Upon encounter with antigen, CD41 T cells differentiate into effector Th subsets with distinctive functions that are related to their unique cytokine profiles and anatomical locations. One of the most B-cell memory describes the populations of cells that provide long-term humoral immunity: long-lived important Th functions is to provide signals to developing B cells that induce specific and appropriate antibody-secreting plasma cells that reside mainly in the bone marrow and memory B cells. Interestingly, antibody responses. The major CD41 T cell subset that helps B cells is the T follicular helper (TFH) cell, the memory B-cell population is heterogenous, although the importance of this heterogeneity has been whose expression of the chemokine receptor CXCR5 [chemokine (C–X–C motif) receptor 5] serves to unclear. Recent studies have investigated the formation and function of memory in different settings. localize this cell to developing germinal centers (GCs) where it provides instructive signals leading to In particular, T-independent memory-like cells and T-dependent (TD) IgM memory B cells qualitatively Ig class switching and somatic mutation. TFH cells produce high levels of IL-21, a cytokine that is differ from canonical TD class-switched memory B cells; however, these studies suggest that IgM critical for GC formation and also for the generation of TFH cells. Although TFH cells have been found memory cells preserve the memory population over long periods of time. These subsets are evocative of to produce cytokines characteristic of other T subsets, they represent a distinct lineage whose the evolution of the humoral immune response,hwith memory-like cells appearing before acquisition of development is driven by the transcription factor B-cell CLL lymphoma-6 (BCL6). Consistent with germinal centers, suggesting that there are multiple pathways to producing B-cell memory. their critical role in the generation of antibody responses, dysregulated TFH function has been associated with the development of systemic autoimmunity. Here, we review the role of IL-21 in the Keywords: antibody, evolution, IgM memory, plasma cells, T-independent regulation of normal TFH development and function as well as in progression of autoimmune responses. Introduction Keywords: autoimmunity, BCL6, germinal center, Th subsets The immune system has one fundamental function—to recognize foreign invaders and reject them from the body, while ensuring that such defense mechanisms do not destroy the host. This function remains the center around which the immune system, at the cellular, biochemical, genetic and Introduction epigenetic levels, has evolved. The immune system has also, Specific CD4+ T cell effector responses can be delegated at various points in evolution, acquired the ability for heightto distinct subsets characterized as Th1, Th2, Th17 or reguened responses to pathogens if the host had been infected latory T cell (Treg) cells, with the development of each of prior—this is termed immune memory. As such, patients with these subsets being driven, respectively, by the master immunodeficiencies that affect the humoral system often have lineage-specific transcription factors T-bet [now also denoted an inability to form and/or maintain memory, resulting in as T-box 21 (TBX21)], GATA binding protein 3 (GATA-3), retiincreased susceptibility to infections (1, 2). Similarly, the ability noid-related orphan receptor ct (RORct) or forkhead box to induce B-cell (and T-cell) memory is the foundation for protein P3 (FOXP3) (1). T-cell-dependent antibody production protective immunity generated by vaccines. Yet, despite the is a critical component of the normal immune response, and clinical relevance of B-cell memory, the mechanisms underlyTh2 cells were originally believed to be the predominant ing formation and function of memory B cells remain elusive. source of B cell help because of their production of IL-4, a Various questions have been posed about B-cell memory. cytokine known to be involved in B cell proliferation as well What signals and genetic networks are required for its as Ig class switching (1). formation? How do memory B cells respond more rapidly Subsequently, IL-21 was identified as a T -derived, type I, and robustly during a secondary response?h What maintains four-a-helical bundle cytokine that was critical for plasma cell the population for long periods of time and, on the flipside, generation as well as isotype switching (2) and normal Ig what prevents exhaustion of the population? Although many production (3), consistent with IL-21 being a Th2-specific cymolecules have been identified to be important for the tokine (4); however, other data indicated that IL-21 had Th1formation, maintenance and regulation of germinal center like properties as well (5). (GC) responses (reviewed in ref. 3), thus impacting on the More recently it became clear that the CD4+ T cells involved quality and quantity of B-cell memory, these fundamental in germinal center (GC) formation and function—denoted questions remain unsolved. T follicular helper cells (TFH cells)—were distinct from any In the past decade, studies on B-cell memory in non-disease of these previously identified subsets. These TFH cells states have had three foci: gene expression studies, utilization of gene-deficient animals to study the roles of genes expressed during humoral responses and the formation of memory subsets. In particular, recent studies have provided a more in-depth characterization of T-independent (TI) formation of B-cell memory (4–6) and the function of murine IgM memory B cells (7, 8). The identification of multiple routes expressed high levels of the chemokine receptor CXCR5 to memory B-cell formation has led to the idea that memory [chemokine (C–X–C motif) receptor 5], allowing them to home can be formed in ‘layers’, reminiscent of how evolution of the to and be retained by the lymphoid follicle, where contact with immune system has been described (9). In this review, we exantigen-primed B cells led to B cell proliferation, isotype amine recent data demonstrating that memory B cells are switching and somatic mutation of the Ig repertoire (6, 7). formed in response to either TI or T-dependent (TD) stimuli. Gene microarray analysis revealed that follicle-localized CXCR5+ Th cells had a very distinctive transcriptional profile that distinguished these cells Th1 or Th2 cells, with highMemory B-cell formation in from TD responses level IL-21 and B-cell CLL lymphoma-6 (BCL6) messenger During a primary TD humoral response, antigen-stimulated RNAs (mRNAs) (5), both of which are now considered hallB cells interact with activated T cells and other accessory marks of TFH cells (6). cells, resulting in the formation of foci of short-lived plasma IL-21 is a type I cytokine that signals via a specific recells (PCs), GC-independent ‘early’ memory cells or a GC ceptor protein, IL-21R (8, 9), and the common cytokine reaction, which can last a number of weeks before subsiding receptor c chain, c , which is shared by the receptors for (reviewed in ref. 9).c Within the GC, B cells undergo rounds IL-2, IL-4, IL-7, IL-9, IL-15 and IL-21 (10); cc is mutated in of division and affinity maturation, in which antibody affinity humans with X-linked SCID (11). IL-21 signals in part via increases due to somatic hypermutation (SHM). Class switch STAT3 (signal transducer and activator of transcription 3) recombination (CSR) from IgM to IgG, IgA or IgE can occur, (12), with actions on a wide range of lineages, including although these processes can also occur outside a GC. T cells, B cells, NK cells and dendritic cells (10). Specifically, Eventually, high-affinity mutants are+ selected and IL-21 can promote the expansion of CD8 T cells, is critical differentiate into memory B cells or long-lived PCs (LLPCs). for normal Ig production by B cells, can inhibit dendritic cell During a secondary response, memory B cells differentiate function and, interestingly, can be pro-apoptotic for B cells rapidly into high-affinity plasmablasts, thus clearing the and NK cells (10). Whereas IL-21 was first identified as infection more quickly than the primary response. REVIEW Correspondence to: W. J. Leonard; E-mail: [email protected] Correspondence to: K. L. Good-Jacobson, Immunology Division, Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade Parkville, Received 25 September 2009, accepted 30 October 2009 Victoria 3052, Australia; E-mail: [email protected] 2404 B-cell memory B-cell memory Memory B-cell subsets The memory B-cell population in higher vertebrates is heterogenous (3). Subsets in both humans and mice can be clearly identified by different Ig isotypes (e.g. IgM or classswitched isotypes) (7, 8, 10–12), and IgM memory B cells and IgG memory B cells have been shown to be intrinsically different (13). However, there are also less distinct differences, such as the levels of expression of different surface proteins (e.g. co-stimulatory factors) (14, 15) that may reflect the different experiences of each cell during the primary response (3). In these latter studies, memory B cells can be categorized into different subgroups based on cell surface markers, and this could be correlated to precursor frequency and mutation rate (but not to class switch) (15). Other groups had previously identified heterogeneity within the memory B-cell population (14–19) in both humans and mice. For example, Cooper et al. have identified a tissue-specific population of memory B cells in humans, identified by the expression of FCRL4 (17, 18). Finally, atypical memory B-cell populations found in HIV and malaria patients have high expression of inhibitory receptors, the presence of which is correlated to defective function of the humoral system in these patients (20, 21). Taken together, these studies suggest that the memory population may be subdivided based on origin and function. Interestingly, many gene-deficient mice and immunodeficient patients that lack productive GCs have cells that resemble memory cells, but the population appears to have an incomplete phenotype, such as few mutations, inadequate affinity maturation or no class-switched isotypes (reviewed in ref. 1, 3). These studies have revealed many genes that are important for different phases of the GC response—i.e. formation, magnitude or resolution—affecting output of the GC and thus the quality and quantity of the memory population but not whether the memory population exists. As a result, the definition of a memory B cell has become contentious. Some researchers have made a priori assumptions based on observations of the cells that have been found to persist in mice and man; i.e. the precursor cell of a canonical memory B cell is a GC cell, and therefore, its Ig must be class switched and mutated. Thus, the main sources of contention are whether IgM and/or low-affinity early memory B cells, which can be found in the absence of a GC [or paucity of GCs; (1, 3, 22–24)], can be defined as memory cells. Similarly, TI B cells have not traditionally been considered as canonical memory B cells. We will focus this review on the recent in vivo research into what might be regarded as non-canonical memory formation. TI memory TI antigens can be TI type I (TI-I; for soluble antigens) or TI-II (where antigen is cell-bound). Recently, various groups have described LLPCs and memory-like cells formed during TI responses in mice. First, LLPCs can be generated to TI-II antigens, which secrete protective antibody from both the secondary lymphoid organs (25) and the bone marrow (4, 5). These LLPCs can either be IgM-secreting or be IgGsecreting; however, in at least one study, TI LLPCs secreted lower amounts of antibody compared with LLPCs generated during a TD response (4). Unlike ‘conventional’ B cells (B2 cells), B1 cells were thought to self-renew in the periphery, to be TI, to express IgM but not to form memory cells; B1-a and B1-b subsets have been described but it is not clear exactly how mouse B1-a/B1-b subsets relate to the human equivalents. Alugupalli et al. (26) demonstrated that mouse B1-b cells give rise to an IgM-expressing B-cell population that persists, controls Borrelia hermsii infection and can confer protection on transfer, suggesting that these are in fact memory cells. Obukhanych and Nussenzweig used (4-hydroxy-3-nitrophenyl)-acetyl (NP)-Ficoll, a TI-II antigen, to generate cells that persisted for many months post-immunization (6). When these cells were adoptively transferred into naive hosts, they rapidly divided, although it is unclear whether they generated a burst of plasmablasts. Interestingly, both these groups confirmed older studies demonstrating that serum Ig, specifically class-switched Ig, inhibits the proliferation of TI memory cells (27, 28). It remains unclear whether the B1-b memory population is continually proliferating or whether, once generated, these cells have an intrinsic survival advantage. There were some differences between these studies. For example, in response to NP-Ficoll, the memory population contains both IgG+ and IgG cells (6), whereas in the study by Alugupalli et al., the B1-b memory population was only IgM+ (26). Furthermore, it is unclear if the precursor cells that respond to NP-Ficoll are of B1 or B2 origin or both, or even whether aborted GCs were generated (29). In sum, both these studies (6, 26) demonstrate that immunity formed during a TI response can be conferred when transferred into naive hosts and that these ‘recall’ responses are inhibited by serum Ig. However, it appears that this is a quantitative rather than qualitative attribute; thus, it is as yet unexplored whether these cells have an intrinsic advantage—as happens with TD memory in vivo and IgM and IgG memory in humans (1, 13, 30, 31)—to respond more rapidly and robustly compared with naive B cells. Thus, it is clear that at least TI-II antigens can form B-cell memory, although it is different to TD memory in regards to its longevity, secretion and phenotype. TD IgM memory B cells Memory B cells that retain IgM on their surface have been shown to exist in both mice (32) and man (1, 10–12). The identity of these cells in humans is contentious, with some groups identifying them as memory based on their functional and molecular similarity to IgG memory and others suggesting that they are ‘circulating’ marginal zone (MZ) B cells (33) (splenic MZ B cells are generally mature, IgMexpressing and non-recirculating cells). Two recent studies have investigated the origin, function and persistence of murine IgM memory cells during a TD response (7, 8). First, Weill et al. utilized a mouse encoding a reporter for activation-induced cytidine deaminase expression to track cells that have been activated during an immunization with sheep red blood cells (7) (a classical TD antigen). Taking this a step further, Jenkins et al. immunized B-cell memory4053 B-cell memory mice with PE (a fluorescent TD antigen) and thus were able to track antigen-specific cells throughout the response (8). Taken together, both groups (7, 8) confirm that IgM memory can be formed, and there are qualitative differences between IgM and IgG1 memory. The two main findings were, first, that IgM memory persists longer than IgG1 memory and, second, that class-switched memory B cells differentiate rapidly into plasmablasts during a secondary response, whereas IgM memory B cells had a slower turnover rate and were less likely to participate in a secondary response in the presence of serum Ig (8). These IgM memory B cells were less mutated than IgG memory B cells, paralleling the IgM+CD27+ memory B cells found in healthy people. Weill et al. proposed the concept of ‘layers’ of memory (7), describing the existence of IgM memory and IgG memory, as well as their proliferating precursors within chronic GCs. Thus, it appears that IgM memory B cells can be generated in both TI and TD responses and in the presence of different adjuvants. Although TI and TD IgM memory cells may qualitatively differ and/or have different precursors, they have overarching characteristics regardless of the response. The population can persist for months in vivo in mice, they divide and confer protection when transferred into naive recipients and are inhibited by serum Ig. Hence, it was concluded that IgM memory B cells are important for persistence of the population, whereas IgG memory B cells are the frontline responders. It is still unclear whether these murine populations are equivalent to IgM+CD27+ memory B cells found in humans, although Obukhanych and Nussenzweig suggest that the inhibition by IgG is an important regulatory mechanism in humans, highlighted by the fact that immunodeficient patients that do not generate IgG display hyperIgM syndrome (6). Can subsets inform memory biology? It could be proposed that the presence of memory subsets answers some long-standing questions about memory B-cell biology. Arguably, the three most important questions regarding memory biology are (Q1) how is the population maintained over long periods of time, sometimes for the life of the host; (Q2) what are the mechanisms underlying rapid and robust secondary responses and (Q3) how are memory B cells formed? We will discuss these recent papers within the context of the first two questions. The data presented in papers by both Weill and Jenkins (7, 8) suggest that IgM memory B cells may maintain the memory population (Q1). Both papers demonstrate that IgM memory B cells persist for longer periods of time than IgG memory B cells—in fact, the same number of IgM memory cells is maintained after an initial decrease during the resolution of the primary response (8), whereas IgG memory B cells diminish in number over time. It is important to note that all these studies are in mice, and therefore, it is difficult to directly compare how long these cells may be maintained in humans. Despite this, the data in these papers suggest that it may be IgM memory B cells that maintain the memory population. On the other hand, these data do not explain how a qualitatively better response is generated, beyond the known abilities of canonical IgG memory B cells [Q2; (34)] as murine IgM memory B cells do not have enhanced response kinetics over naive B cells in vivo (8). It is important to note that, in contrast, human IgM memory B cells have an intrinsic advantage over IgM+ naive B cells with respect to proliferation, co-stimulation, time taken to first division and survival (13, 30, 31); therefore, it is likely that there are modifications in addition to expression of IgG rather than IgM that account for enhanced secondary responses. In mice, however, IgM memory B cells appear to have only two main advantages over IgM+ naive B cells: an increase in number over naive B cells and a higher-affinity B-cell receptor. Would this be enough to induce rapid and robust secondary responses after the eventual loss of IgG memory B cells? On the flipside, the presence of IgM memory B cells may be why the memory B-cell population is not exhausted during a secondary response—although they are poor responders in the presence of serum Ig, in the absence of it, they can generate secondary GCs and replenish the IgG memory compartment. Although these studies do not reveal the molecular mechanisms underlying the formation of memory B cells, they raise an important question: are there multiple independent pathways to becoming B-cell memory (Fig. 1)? Or are the different ‘types’ of memory (TI, IgM memory and early memory) in GC-deficient conditions intrinsically related? As Weill et al. suggest, have we formed layers of memory over time? Evolution of memory B cells Protection, defined as the ability to produce a heightened response to a previously seen infection, has been identified in lower vertebrates, such as bony fish and amphibians [tables summarizing data for secondary responses in vertebrates can be found in (35–37)]. For example, repeated immunizations of trout have demonstrated that successive responses induce heightened antibody levels compared with the primary response. It has even been shown that invertebrates, such as the bumblebee, are able to clear secondary infections more rapidly and with specificity (38). Therefore, it is likely that a memory-like cells exists in lower vertebrates and several invertebrates. The protective antibody responses observed in lower vertebrates do not show an increase in affinity compared with the primary response. Unlike higher vertebrates that can regulate body temperature (homeotherms), poikilotherms do not have the ability to form GCs (36). Yet, lower vertebrates have transcription factors that have homology to those that regulate B-cell differentiation in higher vertebrates, such as Bcl-6 and Blimp-1 (39–42). Although the series of evolutionary events that occurred in order to form GCs are unclear, it appears to at least correlate with enhanced T-cell responses (and thereby T-cell help) (43, 44) that occurred upon the increase and regulation of body temperature (36) and the existence of secondary lymphoid organs with complex organization and the presence of follicular dendritic cells (reviewed in ref. 37). Evidence of affinity maturation and selection appears with GCs in higher vertebrates (36, 37) explaining why secondary antibody responses in lower vertebrates are not of higher affinity than the primary response. 4406 B-cell memory B-cell memory Fig. 1. Layers of memory. (A) Evolution of memory B-cell attributes. In some invertebrates and lower vertebrates, heightened secondary responses have been detected, suggesting immune memory. Ig class switching and SHM have been detected in Xenopus. These traits were found in animals that do not have GCs, which appeared relatively late in the evolutionary tree. With the addition of secondary lymphoid organs and GCs, came affinity maturation and selection, leading to the canonical TD memory seen in humans and mice today. (B) Different pathways to memory. In addition to TD memory [e.g. canonical GC-derived class-switched memory cells derived from follicular (FO) B cells], there are various subsets of memory that have been described in both humans and mice. Shown are memory cells and their possible precursors, demonstrating the complexity of determining lineage relationships. A different presentation of memory B subsets (C) demonstrates that different types of memory have a layering of traits associated with the function of canonical TD memory, as seen with the evolution of memory B cells in invertebrates and vertebrates over time (A). Humans with immunodeficient diseases that affect GC development, as well as lower vertebrates, similarly do not form fully functional humoral responses (as discussed in detail in ref. 45). Various GC-deficient or GC-defective mice, such as those that lack Bcl-6, ICOS or IL-21 (22–24), are still able to produce early memory-like cells, as can B1-b cells in mice. Interestingly, B cells in lower vertebrates can undergo SHM in the absence of a GC. The amphibian Xenopus, for example, contains B cells with somatic mutations (46); however, because there is a lack of evidence for affinity maturation in secondary responses in this species, it appears that these somatic mutants are not expanded and selected into the pool of persisting cells. Similarly, patients with mutations in Cd40L (47) or Sh2d1a (48) that have a paucity of GCs still have B cells that undergo SHM, generating mutated IgM+ cells with a memory phenotype. B cells in these patients can class-switch in the absence of a GC, as can those in lower vertebrates such as Xenopus (which switch to IgY) (46). Therefore, the cellular machinery for SHM and CSR, normally associated with affinity maturation and selection, appears to exist in B cells in both lower vertebrates and patients that lack effective GC responses. Taken together, investigation of primitive immunity demonstrates that B cells in lower vertebrates are able to somatically mutate, differentiate into antibody -producing cells and produce heightened protective responses, indicating that a memory-like population exists in these animals. Are memory B cells an unintended consequence of infection in lower vertebrates? Memory B cells may have been an intermediate state between naive B cells and PCs, i.e. activated B cells that have undergone at least one round of division but do not get signals to differentiate into a PC; instead, they were able to escape death signals and revert back to a ‘naive-like’ state. If so, at some point during evolution, the cells within this population acquired the ability to persist. Evidence for an early induction of a memory state post-activation has been observed in T cells: CD4+ T cells that have been rested 2 days post-activation were observed to have almost identical transcriptional profiles to those that had been rested for 2 months (49). Differentiation into PCs induces a large change in transcriptional networks. In contrast, naive and memory B cells have very similar transcriptional profiles. While no ‘master regulator’ of memory B-cell differentiation has been found, B-cell memory4075 B-cell memory the differences in transcription consist of a number of changes in cell cycle regulators, as well as ligands and receptors that allow interaction with T cells (13, 30, 50, 51). Similarly, upon activation, B cells integrate signals delivered via surface receptors, leading to a genetic re-programing that allows cell cycle and differentiation. Taken together, we hypothesize that an activated B cell has the ability to differentiate into a memory B cell after only a few rounds of division, and without the large changes in transcriptional networks seen in GCs and PCs. GC responses were an addition to the lineage relationship of naive and memory B cells that further honed memory B-cell qualities. Once GCs were part of the microenvironment, these cells were then able to go into a GC ‘state’ and undergo affinity maturation and selection. Memory B-cell evolution can therefore be split into two stages, separated into the ability to produce a cell that can persist to provide protection and the acquisition of qualities that further enhance the efficiency of secondary responses. Evolution of mechanisms that regulate the humoral response was necessary for stable GC function and have in turn shaped the characteristics of the memory B-cell population. Within the GC, regulatory mechanisms provided by Fas–Fas-L, PD-1–PD-L and limiting the amount of T-cell help are vital in shaping the quality and quantity of the memory population (22, 52–55). This is evident in mice lacking molecules important for a fully functional GC—rarely does the ability to form a memory cell become completely disrupted; however, the quality and quantity of the memory population are different to that present in a control animal (reviewed in ref. 3). The characteristics attributed to memory that are attained within the GC, such as selection of high-affinity mutants, are layered over other the characteristics required for persistence. Heterogeneity within the immune system may therefore reflect the importance of layering of attributes of the memory population, both during the immune response and over time. Moving forward There are of course questions that remain unanswered. Are human IgM memory B cells counterparts of B1 or B2 memory in mice? Why is there a difference in longevity and responses of IgM and IgG memory B cells? Is this difference governed by intrinsic differences, and if so, are they at the genetic or epigenetic level? In particular, research into histone modifications that shutdown cell cycle to allow differentiation, or alternatively to allow quick transcription of genes during a secondary response, may reveal the answers to the questions that still consume the memory field. Conclusions Adaptation is required for survival—this is the main tenet of evolution. Thus, it would be advantageous for a naive B cell to be structured in such a way that it is able to differentiate into PCs, GCs or memory B cells, with extrinsic factors and epigenetic modifications influencing the fate of the activated B cell. When the humoral adaptive system is deconstructed into its components—i.e. the ability to respond to and clear foreign antigen, the ability to produce antibody, the ability for a population of antigen-specific cells to persist and the ability of these cells to respond to and clear the immunizing antigen more rapidly during secondary responses—it becomes clear that during evolution, the qualities and subsets of memory B cells routinely studied in mice and humans have been layered over the more primitive structure that originally existed. Genetic and epigenetic changes that allowed persistence of the memory population, and thus induced protection, were beneficial when selective pressures were applied. Therefore, evolution has resulted in the continual selection of traits that enhance the main function of the immune system—to protect the host. Funding This work is supported by National Health and Medical Research Council (NHMRC) Australia program grant to D.M.T. (356202) and project grant to K.L.G-J (1022458). K.L.G-J and DMT are supported by an NHMRC Biomedical Research Fellowship and an NHMRC Research Fellowship, respectively. Acknowledgements We thank Jennifer Walker for critical reading of this review. Note added in proof: While in proof, an article was published in Journal of Experimental Medicine (Taylor et al., 2012 Journal of Experimental Medicine, DOI: 10.1084/jem.20111696) demonstrating that IgM and class-switched memory B cells can be generated through a GC-independent pathway. References 1 Tangye, S. G. and Good, K. L. 2007. Human IgM+CD27+ B cells: memory B cells or ‘‘memory’’ B cells? J. Immunol. 179:13. 2 Rezaei, N., Hedayat, M., Aghamohammadi, A. and Nichols, K. E. 2011. Primary immunodeficiency diseases associated with increased susceptibility to viral infections and malignancies. J. Allergy Clin. Immunol. 127:1329 e2; quiz 1342. 3 Good-Jacobson, K. L. and Shlomchik, M. J. 2010. Plasticity and heterogeneity in the generation of memory B cells and longlived plasma cells: the influence of germinal center interactions and dynamics. J. Immunol. 185:3117. 4 Taillardet, M., Haffar, G., Mondiere, P. et al. 2009. The thymusindependent immunity conferred by a pneumococcal polysaccharide is mediated by long-lived plasma cells. Blood 114:4432. 5 Foote, J. B., Mahmoud, T. I., Vale, A. M. and Kearney, J. F. 2012. Long-term maintenance of polysaccharide-specific antibodies by IgM-secreting cells. J. Immunol. 188:57. 6 Obukhanych, T. V. and Nussenzweig, M. C. 2006. T-independent type II immune responses generate memory B cells. J. Exp. Med. 203:305. 7 Dogan, I., Bertocci, B., Vilmont, V. et al. 2009. Multiple layers of B cell memory with different effector functions. Nat. Immunol. 10:1292. 8 Pape, K. A., Taylor, J. J., Maul, R. W., Gearhart, P. J. and Jenkins, M. K. 2011. Different B cell populations mediate early and late memory during an endogenous immune response. Science 331:1203. 9 Herzenberg, L. A. 1989. Toward a layered immune system. Cell 59:953. 10 Tangye, S. G., Liu, Y. J., Aversa, G., Phillips, J. H. and de Vries, J. E. 1998. Identification of functional human splenic memory B cells by expression of CD148 and CD27. J. Exp. Med. 188:1691. 11 Klein, U., Kuppers, R. and Rajewsky, K. 1997. Evidence for a large compartment of IgM-expressing memory B cells in humans. Blood 89:1288. 12 Klein, U., Rajewsky, K. and Kuppers, R. 1998. Human immunoglobulin (Ig)M+IgD+ peripheral blood B cells expressing the CD27 cell surface antigen carry somatically mutated variable region 6408 B-cell memory B-cell memory 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 genes: cD27 as a general marker for somatically mutated (memory) B cells. J. Exp. Med. 188:1679. Good, K. L., Avery, D. T. and Tangye, S. G. 2009. Resting human memory B cells are intrinsically programmed for enhanced survival and responsiveness to diverse stimuli compared to naive B cells. J. Immunol. 182:890. Anderson, S. M., Tomayko, M. M., Ahuja, A., Haberman, A. M. and Shlomchik, M. J. 2007. New markers for murine memory B cells that define mutated and unmutated subsets. J. Exp. Med. 204:2103. Tomayko, M. M., Steinel, N. C., Anderson, S. M. and Shlomchik, M. J. 2010. Cutting edge: hierarchy of maturity of murine memory B cell subsets. J. Immunol. 185:7146. Bar-Or, A., Oliveira, E. M., Anderson, D. E. et al. 2001. Immunological memory: contribution of memory B cells expressing costimulatory molecules in the resting state. J. Immunol. 167:5669. Ehrhardt, G. R., Hijikata, A., Kitamura, H., Ohara, O., Wang, J. Y. and Cooper, M. D. 2008. Discriminating gene expression profiles of memory B cell subpopulations. J. Exp. Med. 205:1807. Ehrhardt, G. R., Hsu, J. T., Gartland, L. et al. 2005. Expression of the immunoregulatory molecule FcRH4 defines a distinctive tissue-based population of memory B cells. J. Exp. Med. 202:783. Fecteau, J. F., Cote, G. and Neron, S. 2006. A New memory CD27IgG+ B cell population in peripheral blood expressing VH genes with low frequency of somatic mutation. J. Immunol. 177:3728. Moir, S., Ho, J., Malaspina, A. et al. 2008. Evidence for HIVassociated B cell exhaustion in a dysfunctional memory B cell compartment in HIV-infected viremic individuals. J. Exp. Med. 205:1797. Weiss, G. E., Crompton, P. D., Li, S. et al. 2009. Atypical memory B cells are greatly expanded in individuals living in a malariaendemic area. J. Immunol. 183:2176. Zotos, D., Coquet, J. M., Zhang, Y. et al. 2010. IL-21 regulates germinal center B cell differentiation and proliferation through a B cell-intrinsic mechanism. J. Exp. Med. 207:365. Inamine, A., Takahashi, Y., Baba, N. et al. 2005. Two waves of memory B-cell generation in the primary immune response. Int. Immunol. 17:581. Toyama, H., Okada, S., Hatano, M. et al. 2002. Memory B cells without somatic hypermutation are generated from Bcl6-deficient B cells. Immunity 17:329. Hsu, M. C., Toellner, K. M., Vinuesa, C. G. and Maclennan, I. C. 2006. B cell clones that sustain long-term plasmablast growth in T-independent extrafollicular antibody responses. Proc. Natl. Acad. Sci. USA 103:5905. Alugupalli, K. R., Leong, J. M., Woodland, R. T., Muramatsu, M., Honjo, T. and Gerstein, R. M. 2004. B1b lymphocytes confer T cellindependent long-lasting immunity. Immunity 21:379. Brodeur, P. H. and Wortis, H. H. 1980. Regulation of thymusindependent responses: unresponsiveness to a second challenge of TNP-Ficoll is mediated by hapten-specific antibodies. J. Immunol. 125:1499. Hosokawa, T. 1979. Studies on B-cell memory. II. T-cell independent antigen can induce B-cell memory. Immunology 38:291. de Vinuesa, C. G., Cook, M. C., Ball, J. et al. 2000. Germinal centers without T cells. J. Exp. Med. 191:485. Good, K. L. and Tangye, S. G. 2007. Decreased expression of Kruppel-like factors in memory B cells induces the rapid response typical of secondary antibody responses. Proc. Natl. Acad. Sci. USA 104:13420. Good, K. L., Bryant, V. L. and Tangye, S. G. 2006. Kinetics of human B cell behavior and amplification of proliferative responses following stimulation with IL-21. J. Immunol. 177:5236. Dell, C. L., Lu, Y. X. and Claflin, J. L. 1989. Molecular analysis of clonal stability and longevity in B cell memory. J. Immunol. 143:3364. Weller, S., Braun, M. C., Tan, B. K. et al. 2004. Human blood IgM ‘‘memory’’ B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood 104:3647. Martin, S. W. and Goodnow, C. C. 2002. Burst-enhancing role of the IgG membrane tail as a molecular determinant of memory [comment]. Nat. Immunol. 3:182. 35 Du Pasquier, L. 1993. Phylogeny of B-cell development. Curr. Opin. Immunol. 5:185. 36 Nahm, M. H., Kroese, F. G. and Hoffmann, J. W. 1992. The evolution of immune memory and germinal centers. Immunol. Today 13:438. 37 Hofmann, J., Greter, M., Du Pasquier, L. and Becher, B. 2010. B-cells need a proper house, whereas T-cells are happy in a cave: the dependence of lymphocytes on secondary lymphoid tissues during evolution. Trends Immunol. 31:144. 38 Sadd, B. M. and Schmid-Hempel, P. 2006. Insect immunity shows specificity in protection upon secondary pathogen exposure. Curr. Biol. 16:1206. 39 Ohtani, M. and Miyadai, T. 2011. Functional analysis of fish BCL-6 and Blimp-1 in vitro: transcriptional repressors for B-cell terminal differentiation in fugu (Takifugu rubripes). Mol. Immunol. 48:818. 40 Ohtani, M., Miyadai, T. and Hiroishi, S. 2006. Identification of genes encoding critical factors regulating B-cell terminal differentiation in torafugu (Takifugu rubripes). Comp. Biochem. Physiol. Part D Genomics Proteomics 1:109. 41 Ohtani, M., Miyadai, T. and Hiroishi, S. 2006. Molecular cloning of the BCL-6 gene, a transcriptional repressor for B-cell differentiation, in torafugu (Takifugu rubripes). Mol. Immunol. 43:1047. 42 Ohtani, M., Miyadai, T. and Hiroishi, S. 2006. B-lymphocyteinduced maturation protein-1 (Blimp-1) gene of torafugu (Takifugu rubripes). Fish Shellfish Immunol. 20:409. 43 Avtalion, R. R., Wojdani, A., Malik, Z., Shahrabani, R. and Duczyminer, M. 1973. Influence of environmental temperature on the immune response in fish. Curr. Top. Microbiol. Immunol. 61:1. 44 Rijkers, G. T., Frederix-Wolters, E. M. and van Muiswinkel, W. B. 1980. The immune system of cyprinid fish. Kinetics and temperature dependence of antibody-producing cells in carp (Cyprinus carpio). Immunology 41:91. 45 Cooper, M. D. 2002. Exploring lymphocyte differentiation pathways. Immunol. Rev. 185:175. 46 Wilson, M., Hsu, E., Marcuz, A., Courtet, M., Du Pasquier, L. and Steinberg, C. 1992. What limits affinity maturation of antibodies in Xenopus—the rate of somatic mutation or the ability to select mutants? EMBO J. 11:4337. 47 Facchetti, F., Appiani, C., Salvi, L., Levy, J. and Notarangelo, L. D. 1995. Immunohistologic analysis of ineffective CD40-CD40 ligand interaction in lymphoid tissues from patients with X-linked immunodeficiency with hyper-IgM. Abortive germinal center cell reaction and severe depletion of follicular dendritic cells. J. Immunol. 154:6624. 48 Ma, C. S., Hare, N. J., Nichols, K. E. et al. 2005. Impaired humoral immunity in X-linked lymphoproliferative disease is associated with defective IL-10 production by CD4+ T cells. J. Clin. Invest. 115:1049. 49 McKinstry, K. K., Golech, S., Lee, W. H., Huston, G., Weng, N. P. and Swain, S. L. 2007. Rapid default transition of CD4 T cell effectors to functional memory cells. J. Exp. Med. 204:2199. 50 Tomayko, M. M., Anderson, S. M., Brayton, C. E. et al. 2008. Systematic comparison of gene expression between murine memory and naive B cells demonstrates that memory B cells have unique signaling capabilities. J. Immunol. 181:27. 51 Klein, U., Tu, Y., Stolovitzky, G. A. et al. 2003. Transcriptional analysis of the B cell germinal center reaction. Proc. Natl. Acad. Sci. USA 100:2639. 52 Takahashi, Y., Ohta, H. and Takemori, T. 2001. Fas is required for clonal selection in germinal centers and the subsequent establishment of the memory B cell repertoire. Immunity 14:181. 53 Good-Jacobson, K. L., Szumilas, C. G., Chen, L., Sharpe, A. H., Tomayko, M. M. and Shlomchik, M. J. 2010. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nat. Immunol. 11:535. 54 Linterman, M. A., Beaton, L., Yu, D. et al. 2010. IL-21 acts directly on B cells to regulate Bcl-6 expression and germinal center responses. J. Exp. Med. 207:353. 55 Victora, G. D., Schwickert, T. A., Fooksman, D. R. et al. 2010. Germinal center dynamics revealed by multiphoton microscopy with a photoactivatable fluorescent reporter. Cell 143:592.