Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
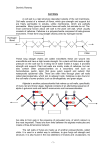
The Plant Journal (2008) 54, 559–568 doi: 10.1111/j.1365-313X.2008.03463.x HARNESSING PLANT BIOMASS FOR BIOFUELS AND BIOMATERIALS Cell-wall carbohydrates and their modification as a resource for biofuels Markus Pauly* and Kenneth Keegstra Department of Energy Plant Research Laboratory and Department of Biochemistry and Molecular Biology, Michigan State University, East Lansing, MI 48824, USA Received 1 December 2007; revised 5 February 2008; accepted 8 February 2008. * For correspondence (fax +1 517 353 9168; e-mail [email protected]). Summary Plant cell walls represent the most abundant renewable resource on this planet. Despite their great abundance, only 2% of this resource is currently used by humans. Hence, research into the feasibility of using plant cell walls in the production of cost-effective biofuels is desirable. The main bottleneck for using wall materials is the recalcitrance of walls to efficient degradation into fermentable sugars. Manipulation of the wall polysaccharide biosynthetic machinery or addition of wall structure-altering agents should make it possible to tailor wall composition and architecture to enhance sugar yields upon wall digestion for biofuel fermentation. Study of the biosynthetic machinery and its regulation is still in its infancy and represents a major scientific and technical research challenge. Of course, any change in wall structure to accommodate cost-efficient biofuel production may have detrimental effects on plant growth and development due to the diverse roles of walls in the life of a plant. However, the diversity and abundance of wall structures present in the plant kingdom gives hope that this challenge can be met. Keywords: cell walls, polysaccharide biosynthesis, hemicellulose, biofuel. Plant cell walls are the most abundant renewable resource on this planet It has been estimated that the net CO2 fixation by land plants per year is approximately 56 · 109 tonnes (Table 1) (Field et al., 1998) and that the worldwide biomass production by land plants is 170–200 · 109 tonnes (Lieth, 1975). Of this amount, 70% is estimated to represent plant cell walls (Duchesne and Larson, 1989; Poorter and Villar, 1997). Humans use these wall materials mainly in the form of wood for heat production (Table 2), and as a building material (timber), in the pulp and paper industry (Fenning and Gershenzon, 2002), and as raw material in the textile industry, e.g. cotton fibers (http://faostat.fao.org/site/567/ default.aspx; parameter settings: production quantity, cotton lint, world, 2006). Taken together, only 2% of the plant cell-wall-based biomass is currently utilized by humans (Table 2). It is thus not surprising that interest in using this resource as a material for biofuels has increased in recent years (Schubert, 2006). Important advantages of wall materials as feedstocks for biofuel production are their great ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd abundance and the fact that they do not serve as food for animals and humans as starch does, for example. All cell walls of higher plants contain cellulose, a homopolymer of b-1,4-linked glucose units, mainly in the form of crystalline microfibrils as well as in an amorphous form (Carpita and McCann, 2000). Walls also contain hemicelluloses (Figure 1), such as substituted glucans, xylans and/or mannans, and anionic components such as the galacturonic acid-containing pectic polysaccharides. Walls may also contain polyphenols such as lignins, and, to a minor extent, structural proteins. The prevalence of polysaccharides in the wall is particularly advantageous for plants, as they are generated directly from the products of photosynthesis without the utilization of large amounts of nitrogen or phosphorus, two macronutrients that frequently limit plant growth. Not all cell walls have the same polysaccharide composition When considering wall materials for the production of biofuels, one should be aware that walls from higher plants 559 560 Markus Pauly and Kenneth Keegstra Table 1 Annual production of plant cell walls Production Source Assimilated CO2 Land plants, net primary production Land, worldwide Land, worldwide Plant biomass Cell walls Tonnes 56 · 109 170–200 · 109 150–170 · 109 Table 2 Annual human utilization of plant cell walls Material Product Tonnes per year Wood Wood Energy Timber, pulp and paper 1.05 · 109 0.95 · 109 differ quite substantially in content, both qualitatively and quantitatively. While the cell is still elongating, a primary wall is formed. Primary walls contain cellulose and a hydrated (65% water) matrix consisting of hemicelluloses and pectins, with some structural proteins (Brett and Waldron, 1996). Based on their polysaccharide composition, primary cell walls are usually classified as type I or type II. Type I walls are present in dicots and non-commelinoid monocots; in addition to cellulose, they generally contain xyloglucan as the main hemicellulose and abundant amounts of pectic polysaccharides (Carpita and Gibeaut, 1993). In type II walls, the walls of the Poales, such as the grasses, arabinoxylan is the major hemicellulose. In addition, type II walls contain a higher percentage of cellulose and only negligible amounts of pectins and proteins (Carpita, 1996). Secondary walls, deposited once cell elongation ceases, are usually thicker than primary walls and may be deposited in a number of layers. Secondary walls contain cellulose and arabinoxylan and/or glucomannans as the major hemicellulose (Brett and Waldron, 1996). More importantly, in secondary walls, water is largely replaced by lignin, making them nearly impenetrable to solutes and enzymes. At the single-plant level, nearly all of the approximately 35 different cell types can be distinguished based on their varying wall structures, as observed by microscopy (Carpita and McCann, 2000), chemical composition analysis (Richmond and Somerville, 2001) and labeling of wall polymers with specific antibodies (Willats et al., 2000). Hence, it is not Figure 1. Examples of hemicellulose structures present in cell walls, including monosaccharides, and their linkage and ester constituents (modified from Somerville et al., 2004). Easily fermentable hexoses are distinguished by their square shape. Other sugars are shown in different shapes. ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 Cell-wall carbohydrates 561 Figure 2. Cell-wall polymer composition (cellulose, hemicellulose and lignin) and hemicellulose composition for a variety of plant materials that are currently under discussion for use as biofuel feedstocks (based on the data shown in Table 3 and references therein). surprising that plant-wall feedstocks that could be used for biofuel production can differ quite significantly in their composition, even though the materials derive mainly from secondary walls (Figure 2). In Table 3, some of the prominent plant materials that could be used for biofuel production are listed. Although the numbers cannot be compared with each other because different methods were used to establish them, some general features become evident. The most dominant polysaccharide in these walls is cellulose, making up 40.6–51.2% of the wall material. The next largest fraction comprises the hemicelluloses, representing 28.5–37.2% of the walls. Lignin occurs in these walls at a lower percentage (13.6–28.1%), with a more than twofold difference between walls of switchgrass, for example, and those of softwood. Viewed in greater detail, composition is seen to vary widely with regard to the hemicelluloses (Table 3, and references cited therein; for structures, see Figure 1). The grasses contain mainly arabinoxylan, but the degree of arabinosylation can vary greatly. Wheat straw contains the lowest degree of substitution, whereas sorghum xylans have an exceptionally high degree of substitution (Verbruggen et al., 1995). Hardwood (from angiosperm trees) also contains mainly xylan, but with a negligible degree of arabinosylation. The xylans here are mainly substituted with glucuronic acid or 4-O-methyl-glucuronic acid residues (Ebringerova and Heinze, 2000). In contrast, softwood (from conifers) contains mannans such as O-acetylated galactoglucomannans (Capek et al., 2002) as their main hemicellulose, although ample amounts of xylans are also present. Most importantly, the differences and ranges of wall components and their fine structures are also a result of the differences in tissues from which the feedstocks are derived (e.g. corn stover; Chundawat et al., 2007). Bottlenecks in utilizing cell-wall materials for biofuels Plant cell-wall materials can be converted in a number of ways. One way is the combustion and gasification of plant material. The resulting CO and hydrogen gas (also called syngas) can be converted to hydrocarbons of various lengths via a catalyzed chemical reaction (Fischer–Tropsch process). Hydrocracking of the large hydrocarbons can be used to produce diesel fuels (Tijmensen et al., 2002). Here, the main objective from a cell-wall perspective is simply an Table 3 Comparison of biomass feedstocks Feedstocka b Corn stover Wheat straw Rice straw Miscanthus Sorghum (whole sorghum pith and bark) Switchgrass (late cut) Sugar cane Hardwood (beech; Fagus sylvatica) Softwood (spruce; Picia abies) Cellulose Hemi cellulose Lignin Ash Protein Solubles Reference 39.4 34.9 41.6 41.9 15.0 33.1 22.5 31.5 26.6 12.3 14.9 21.3 12.5 13.3 5.8 ND 9.4 14.4a 3.2 0.4 3.7 ND ND ND ND 8.9 11.9 ND 15.0 66.5b Chundawat et al. (2007) Lynd et al. (1999) Jin and Chen (2007) Magid et al. (2004) Billa et al. (1997) 46.1 48.6 43.3 40.4 32.2 31.1 31.8 31.1 12.3 19.1 24.4 28.0 4.7 1.2 0.5 0.5 4.6 ND ND ND ND ND ND ND Lynd et al. (1999) Sanjuan et al. (2001) Fengel and Wegener (1989) Fengel and Wegener (1989) Values have been adjusted to a percentage basis (dry weight). Mainly silicate. b Mainly sucrose. ND, not determined in these studies. a ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 562 Markus Pauly and Kenneth Keegstra increase in the production of biomass (cell walls) per hectare, irrespective of its wall composition, although a low water and ash content is desirable (Tijmensen et al., 2002). A more sophisticated approach in biofuel production involves the degradation of wall materials to monosaccharides and subsequent fermentation to liquid fuels such as bioethanol (Schubert, 2006). However, plants have evolved wall structures to accommodate their needs in completing their lifecycle, not to suit mankind’s desire to exploit this resource for the production of biofuels. As a result, cell walls are naturally resistant to breakdown by mechanical and microbial forces, which are precisely the processes needed for the cost-effective and efficient production of monosaccharides. Hence, one major objective is to make walls more accessible to degradation (Himmel et al., 2007; Houghton et al., 2006). We could achieve this goal by increasing water solubility and hence access of enzymes to polysaccharides. One way to do this would be to add de novo synthesized, water-soluble polysaccharides to existing cells, leading to a greater abundance of these polysaccharides in the wall. One alternative would be a shift in the ratio of less soluble polysaccharides to soluble ones. This objective would require: (1) for cellulose, an increase in the abundance of amorphous glucan chains rather than crystalline microfibrils; (2) for hemicelluloses, addition of side chains to decrease hydrogen bonding with cellulose microfibrils; and (3) for lignins, a general reduction in their amount or amendment to a more easily degradable form (Akin, 2007), for example by introduction of specific monolignols (Boerjan et al., 2003; Chen and Dixon, 2007) and/or decrease of the existing lignin–hemicellulose linkages (Grabber et al., 2002). Another consideration is the fermentability of the wall degradation products, the resulting monosaccharides. Currently, the sugars most easily fermentable by yeasts are the hexoses, such as glucose and mannose, rather than the pentoses, although yeast and bacterial strains have been developed that can efficiently ferment pentoses (Chu and Lee, 2007). Hence, an increased production of hexose-containing polymers such as cellulose, glucomannans and to some extent xyloglucan is more desirable than an increase in arabinoxylans, for example (see hexose and pentose annotation in Figure 1). Depending on the fermentation- or catalyst-based chemical process (Huber et al., 2005) used to produce fuels, monosaccharide fermentation-inhibiting components, such as aliphatic acids (e.g. acetic acid) or phenolic compounds (Larsson et al., 1999) are present to varying degrees in the degraded biomass. One goal should be to reduce the abundance of such compounds to a minimum. All of the above-mentioned changes could be accomplished by either manipulation of the plant biosynthetic pathways for the respective polymers and/or post-deposition metabolism alterations in planta. Manipulation of the biosynthetic pathways The natural variability in wall compositional quantity and quality (Figure 2) suggests that there is an opportunity for altering the abundance of specific wall components without compromising the life cycle of a plant. Such a feat could be accomplished by manipulation of the biosynthesis of specific wall polysaccharides. The two most abundant polysaccharides of plant cell walls, cellulose and hemicellulose, are synthesized in different compartments by significantly different processes. Cellulose, generally the most abundant component in secondary cell walls (see Figure 1), is synthesized at the plasma membrane by a complex machinery that we are just beginning to understand (Somerville, 2006). Whereas the glucosyl residues come from UDP-glucose molecules that are present in the cytosol, the cellulose microfibrils are deposited into the extracellular wall matrix at a location adjacent to the plasma membrane (Somerville, 2006). On the other hand, the hemicellulosic polysaccharides present are synthesized in the Golgi and packaged into secretory vesicles before delivery to the cell surface and incorporation into the wall matrix. The assembly events that combine these components into the composite that exists in the wall matrix are still poorly understood. The CesA proteins are thought to be the catalytic subunits of the cellulose synthase complexes (Somerville, 2006). These proteins are encoded by a family of CesA genes that are found throughout the plant kingdom (Hazen et al., 2002; Richmond and Somerville, 2000). Genetic studies have led to the conclusion that three CesA genes are needed for cellulose biosynthesis in primary cell walls (Persson et al., 2007b), and another set of three CesA genes is required for cellulose synthesis in secondary cell walls (Somerville, 2006). The three different CesA proteins are thought to cluster into a higher-order structure, which forms the rosette structure observed in the plasma membrane (Somerville, 2006). The rosettes containing multiple CesA proteins are thought to move in the plasma membrane in a direction that is defined by cortical microtubules, thereby producing cellulose microfibrils outside the plasma membrane. These microfibrils are deposited in a pattern that reflects the orientation of the cortical microtubules present on the cytosolic side of the plasma membrane (Paredez et al., 2006). Despite this emerging outline of how cellulose is deposited, many important issues remain unresolved. Genetic experiments provide evidence that additional proteins are involved in cellulose deposition (Lane et al., 2001; Pagant et al., 2002). One of these proteins has been shown to be a membrane-bound endoglucanase/cellulase and is thought to act as an editing/repairing protein during cellulose biosynthesis (Mølhoj et al., 2002). However, the precise roles of this and the other proteins are still not clear. ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 Cell-wall carbohydrates 563 Whether the encoded proteins are part of the rosette structure or whether they have other roles in cellulose deposition remain to be determined. As more is learned about the details of cellulose biosynthesis, it may be possible to alter these processes in ways that would render the walls more easily digestible during processing to biofuels. For example, if one understood the details of how the glucan chains come together to form crystalline cellulose, it might be possible to modify this process such that cellulose microfibrils would have larger amorphous regions. The biosynthesis of hemicellulosic polysaccharides in the Golgi apparatus differs significantly from cellulose biosynthesis. In the case of the mannans, the backbone is synthesized by CslA proteins (Liepman et al., 2007) that have been identified in a number of species (Dhugga et al., 2004; Liepman et al., 2005, 2007; Suzuki et al., 2006). Each plant species for which the complete genome is available has a small family of CslA genes that are part of the CesA superfamily. The CslA proteins produced in heterologous systems not only have the ability to synthesize mannan when GDP-mannose is present, they also have the ability to synthesize glucomannan when a mixture of GDP-mannose and GDP-glucose is present (Liepman et al., 2005, 2007; Suzuki et al., 2006). Thus, the same protein is able to incorporate both sugars into the backbone in vitro, and it is likely that the same proteins produce both mannans and glucomannans in vivo. The degree of galactosylation of the mannan backbone has implications for mannan solubility. Mannans with a low degree of galactosyl substitution have limited solubility in water, whereas polymers with a high degree of substitution have important properties as emulsifiers (Reid et al., 1988). The galactosyltransferase enzymes that add side chains to the mannan and glucomannan backbones have been identified and characterized (Edwards et al., 1999, 2002). The levels of these enzymes appear to control the degree of substitution of the backbone, with the side chains being added in patterns that are described by hidden Markov models (Edwards et al., 2004). Altering the degree of sidechain substitutions will be vital in engineering more soluble mannans. The mannans are attractive candidates for enhancing wall composition with the aim of creating improved biofuel crops for several reasons. First, the genes and proteins needed for mannan biosynthesis have been identified. Second, the genes needed for mannan biosynthesis appear to be present in all land plants, although their expression levels are such that few mannans are present in the walls of most angiosperms. However, mannans accumulate to high levels in the seeds of many plants, where they serve as storage carbohydrates (Meier and Reid, 1982). During germination, seedlings have the ability to rapidly degrade the mannans and use the resulting sugars as a source of carbon for early seedling development. Because the released sugars are all hexoses, they can easily speed up the central metabolism of the developing seedling. Given these circumstances, it may be possible to enhance mannan levels in vegetative tissues such that the polymers could be easily degraded after harvest to yield hexoses, which could be converted to biofuels more efficiently than the pentoses released from the more abundant xylans. Although xyloglucan is found mainly in the primary cell walls of many plants (Hayashi, 1989), its biosynthesis is relevant to our general understanding of hemicellulose biosynthesis and therefore will be briefly summarized here. All the glycosyltransferases involved in synthesis of its side chains have been tentatively identified (see reviews by Scheible and Pauly, 2004; Lerouxel et al., 2006). However, it remains unclear how the enzymes achieve the structural side-chain diversity found in this polymer. Cocuron et al. (2007) recently presented evidence that the glucan synthase required for making the backbone of xyloglucan is encoded by a CslC gene. When CslC genes were expressed in Pichia pastoris cells, the cells accumulated significant quantities of oligosaccharides containing b1,4-linked glucosyl residues. When one of the xyloglucan xylosyltransferase genes, which is responsible for substituting the glucan backbone with xylosyl residues (Faik et al., 2002), was co-expressed with the CslC gene, the cells produced large quantities of unsubstituted b-1,4-glucan (Cocuron et al., 2007). These observations provide evidence that these xylosyltransferase and glucan synthase enzymes interact to form a complex that has an impact on the nature of the resulting product, even though one of them does not exhibit any activity. Further work is needed to confirm this interesting hypothesis, as protein complexes involved in hemicellulose biosynthesis have yet to be discovered. Given that xylans are the most abundant hemicellulose present in the secondary walls of plants being considered for use in biofuel production (see Figure 1), it is unfortunate that we know so little about their biosynthesis. Recently, several groups have begun to make progress in this difficult area. One of the most interesting observations comes from the work of Peña et al. (2007), who examined the xylan polysaccharides present in two mutant lines of Arabidopsis that have irregular xylem phenotypes. First, these authors rediscovered an older, but little noticed, observation that xylan polysaccharides often have an unusual oligosaccharide at the reducing end. This oligosaccharide contains the glycosyl sequence 4-b-D-Xyl-(1,4)-b-D-Xyl-(1,3)-a-L-Rha-(1,2)-a-D-GalA(1,4)-D-Xyl. Because it is at the reducing end of the polysaccharide, it is possible that this oligosaccharide serves as the primer for chain initiation, if chain elongation occurs from the reducing end toward the non-reducing end, as is commonly hypothesized. Both of the mutants have reduced levels of xylan in the secondary walls of xylem elements, leading to the irregular xylem phenotype. ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 564 Markus Pauly and Kenneth Keegstra One of the mutants, irx9, has increased levels of the unusual oligosaccharide, but the chains containing it are shorter than in wild-type plants, suggesting that the IRX9 gene is involved in elongating the xylan chains. On the other hand, the other mutant, irx8, has little of the unusual oligosaccharide and lower quantities of xylan (Persson et al., 2007a); the xylan that is present is longer and more heterodisperse in size. These observations suggest that IRX8 may be involved in synthesizing the unusual oligosaccharide, which may serve as a primer in wild-type plants. These observations highlight the complexity of xylan biosynthesis, but offer some hope that these new observations can lead to an improved understanding of this important cell-wall polymer. As illustrated above, much is to be learned about the biosynthetic machinery of polysaccharides. We are just beginning to understand the carbon flux into the specific wall polysaccharides (Sharples and Fry, 2007) and regulation of polysaccharide biosynthesis. Post-deposition wall changes Another way to make wall structures more enzyme-accessible is to add ‘loosening agents’ through transgenic approaches. Such agents include the expansins, plant proteins that have been shown to induce the extensibility of plant tissues under stress (Cosgrove, 2000). The precise mechanism of expansin action is unknown, but it is thought that they act at the interface of hemicellulose polymers and cellulose microfibrils (Cosgrove, 2000). Proteins with a similar mode of action are the fungal swollenins, proteins that consist of a cellulose-binding domain and an expansinlike domain. They are thought to disrupt cellulose microfibrils without hydrolytic activity, i.e. the release of reducing sugars (Saloheimo et al., 2002). Adding expansins to wall materials can double the yield of sugars released by fungal cellulases (Cosgrove, 2001a,b), and it is expected that swollenin might have a similar effect. Another class of proteins that could be used to make the wall more accessible is glycanases, in particular endoglucanases. Expression of a poplar endoglucanase in Arabidopsis leads to increased cell elongation and subsequent plant growth (Park et al., 2003). A similar effect was found when a fungal xyloglucanase (a xyloglucan-specific endoglucanase; Pauly et al., 1999) was expressed in poplar (Park et al., 2004). This is not surprising, as this enzyme is the only protein other than expansin that is known to induce wall extension (Yuan et al., 2001). One effect in the transgenic poplar material is an observed increase in cellulose, which leads to material with higher hexose content. A side-effect of the enhanced growth of these transgenic plants is an increase in the photosynthetic canopy, potentially allowing more biomass to accumulate. Other agents that work on the hemicellulose–cellulose network are xyloglucan transglycosylases/hydrolases. This class of enzymes is thought to be involved in either incorporating newly synthesized hemicelluloses and/or remodeling existing hemicelluloses present in the wall by loosening/re-ligating xyloglucan (Fry et al., 1992; Nishitani, 1997). It has been demonstrated that xyloglucan transglycosylases/hydrolases are active in cell elongation and act at the cellulose/xyloglucan interface (Vissenberg et al., 2000, 2005). Manipulating the levels of this agent thus has the potential to loosen cell-wall structures. Other examples of glycanases that have been expressed in plants are pectin-degrading enzymes. Expression of a galactanase in potato tubers led to significant wall alteration (i.e. reduction of galactans), but had no effect on plant or tuber development (Sørensen et al., 2000). Interestingly, the tubers exhibited a marked change in physical tissue properties (Ulvskov et al., 2005). In particular, the water-binding capacity was changed, indicating that removing pectin side chains would probably render such wall material less degradable. Agents that work on the hemicellulose–lignin interface, i.e. that break the covalent bonds between the polymers, can lead to more easily degradable wall materials. For example, expression of phenolic esterases improves the release of fermentable sugar (Akin, 2007). Modification of plant cell walls will challenge their biological function Any strategy to improve wall materials in planta as feedstocks for biofuels needs to take into account the possibility of functional ‘failure’ of the cell wall, which could be detrimental to plant growth, leading to a concomitant reduction in wall biomass and ultimately threatening the very survival of the plant. Cell walls are important for structural integrity of the cell and indeed the whole plant. Through the evolutionary introduction of polyphenol incorporation into the wall, land plants were able to increase their sunlight-harvesting capacity by increasing the plant canopy, not only in terms of width, but also in terms of height, enabling the plant to compete with other plants for sunlight. A number of examples have demonstrated that altering walls can lead to structural concessions such as dwarfism (Desprez et al., 2007) and even to lethality (Goubet et al., 2003). Cell walls determine the shape of the cell, so altering them can lead to changes in morphology, such as an irregular xylem (Turner et al., 2007), that may be disadvantageous to water transport. Walls, in particular the pectinaceous middle lamellae, ensure attachment of the cells. A number of mutants with altered pectic polysaccharides have been shown to have reduced cell adhesion (Bouton et al., 2002; Iwai et al., 2002; Krupkova et al., 2007), probably ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 Cell-wall carbohydrates 565 limiting the plant’s ability to withstand certain mechanical stresses (such as wind). On the other hand, wall materials from such plants might be more accessible to walldegrading enzymes, making processing of these plants more rapid, easier, and thus more cost-effective. Another function of the wall is keeping plant pathogens such as bacteria and fungi away from the nutritious cytosolic content of the cells. In addition to a simple mechanical line of defense, walls contain signaling molecules that allow the plant cell to recognize a pathogen attack and to respond with various lines of defense (Cote and Hahn, 1994; Vorwerk et al., 2004). Alterations in wall composition and architecture thus also introduce the possibility of increased susceptibility to pathogens, or endogenous release of wall-derived oligosaccharides might lead to disease symptoms. For example, expression of a fungal arabinanase in potato tubers led to a severely stressed plant morphology (Skjot et al., 2002), presumably through the release of apoplastic arabinan-oligosaccharides. This morphology was overcome when the enzyme was targeted to the Golgi apparatus instead of the apoplast. Targeting and retaining the arabinanase in the Golgi led to plants and tubers with unaltered appearance but with a significant decrease in pectic arabinans. In a few cases where inhibition of polysaccharide biosynthesis through genetic engineering of the glycan synthases was achieved, these alterations did not result in diminishment of pathogen resistance (Jacobs et al., 2003), and in some cases even increased resistance (HernandezBlanco et al., 2007). The plant cell wall is a dynamic entity that undergoes delicate metabolic changes during cell elongation and differentiation. Throughout the elongation process, the cell must balance loosening the wall with maintaining turgor pressure and cohesiveness of the wall structure. It is thought that metabolism of the hemicelluloses interlacing cellulose microfibrils with wall-loosening enzymes such as endoglucanases, xyloglucantransglycosylases and expansins allows slippage of cellulose microfibrils and thus controlled cell elongation (Cosgrove, 2001a,b). As cells elongate, new cell-wall material is deposited (Refregier et al., 2004), probably leading to strengthening of the wall. Consequently, changing the abundance or structure of wall polymers may stiffen the wall to the extent that the cell cannot enlarge effectively, or may lead to mechanical failure and hence bursting of the cell during the elongation process. It has become clear that the plant cell has a hitherto unknown mechanism for monitoring wall integrity and compensating for change (Humphrey et al., 2007; Pilling and Höfte, 2003). Candidates for such a monitoring activity are plasma membrane-localized, wall-associated kinases (Wagner and Kohorn, 2001), which that have been shown to bind to the pectin matrix in the apoplast (Kohorn et al., 2006). Recently, another plasma membrane-localized receptor has been identified that may also act as such a wall sensor (Hematy et al., 2007). Manipulating putative sensing mechanisms has the potential to overcome unexpected wall structural changes, even though they might be beneficial, such as decreasing lignin content but increasing the relative content of cellulose (Hu et al., 1999). Concluding remarks Owing to the abundance of cell-wall material generated by plants, cell walls could play a prominent role in our quest for the reduced utilization of carbon dioxide-emitting fossil fuels. The scientific and technical challenges inherent in realizing this goal are enormous. The production of walls with tailored polysaccharide composition and structures is still in its infancy due to our lack of knowledge of polysaccharide biosynthesis and its regulation. Despite the above-mentioned difficulty of making the wall polysaccharides more degradable, the current recalcitrance of wall materials brings with it the advantage that harvested wall materials, unlike grains and fruits, can be stored relatively easily for extended periods without loss of yield prior to factory processing. Also, identification of specific bioenergy crop species with high biomass yields grown in various climatic regions has just begun, as have breeding programs for the increased production of biomass. References Akin, D.E. (2007) Grass lignocellulose – strategies to overcome recalcitrance. Appl. Biochem. Biotechnol. 137, 3–15. Billa, E., Koullas, D.P., Monties, B. and Koukios, E.G. (1997) Structure and composition of sweet sorghum stalk components. Indust. Crops Prod. 6, 297–302. Boerjan, W., Ralph, J. and Baucher, M. (2003) Lignin biosynthesis. Annu. Rev. Plant Biol. 54, 519–546. Bouton, S., Leboeuf, E., Mouille, G., Leydecker, M.T., Talbotec, J., Granier, F., Lahaye, M., Höfte, H. and Truong, H.N. (2002) Quasimodo1 encodes a putative membrane-bound glycosyltransferase required for normal pectin synthesis and cell adhesion in Arabidopsis. Plant Cell, 14, 2577–2590. Brett, C. and Waldron, K. (1996) Physiology and Biochemistry of Plant Cell Walls. Topics in Plant Functional Biology. London: Chapman and Hall. Capek, P., Alfoldi, J. and Liskova, D. (2002) An acetylated galactoglucomannan from Picea abies L. Karst. Carbohydr. Res. 337, 1033–1037. Carpita, N.C. (1996) Structure and biogenesis of the cell walls of grasses. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 445– 476. Carpita, N.C. and Gibeaut, D.M. (1993) Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant J. 3, 1–30. Carpita, N. and McCann, M. (2000) The cell wall. In Biochemistry and Molecular Biology of Plants (Buchanan, B.B., Gruissem, W. and ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 566 Markus Pauly and Kenneth Keegstra Jones, R.L., eds). Rockville, MD: American Society of Plant Physiologists, pp. 52–108. Chen, F. and Dixon, R.A. (2007) Lignin modification improves fermentable sugar yields for biofuel production. Nat. Biotechnol. 25, 759–761. Chu, B.C.H. and Lee, H. (2007) Genetic improvement of Saccharomyces cerevisiae for xylose fermentation. Biotechnol. Adv. 25, 425–441. Chundawat, S.P.S., Venkatesh, B. and Dale, B.E. (2007) Effect of particle size based separation of milled corn stover on AFEX pretreatment and enzymatic digestibility. Biotechnol. Bioeng. 96, 219–231. Cocuron, J.C., Lerouxel, O., Drakakaki, G., Alonso, A.P., Liepman, A.H., Keegstra, K., Raikhel, N.V. and Wilkerson, C.G. (2007) A gene from the cellulose synthase-like C family encodes a b–1,4 glucan synthase. Proc. Natl Acad. Sci. USA, 104, 8550–8555. Cosgrove, D.J. (2000) Loosening of plant cell walls by expansins. Nature 407, 321–326. Cosgrove, D.J. (2001a) Wall structure and wall loosening. A look backwards and forwards. Plant Physiol. 125, 131–134. Cosgrove, D.J. (2001b) Enhancement of Accessibility of Cellulose by Expansins. US Patent No. 6326470. Cote, F. and Hahn, M.G. (1994) Oligosaccharins: structures and signal-transduction. Plant Mol. Biol. 26, 1379–1411. Desprez, T., Juraniec, M., Crowell, E.F., Jouy, H., Pochylova, Z., Parcy, F., Höfte, H., Gonneau, M. and Vernhettes, S. (2007) Organization of cellulose synthase complexes involved in primary cell wall synthesis in Arabidopsis thaliana. Proc. Natl Acad. Sci. USA, 104, 15572–15577. Dhugga, K.S., Barreiro, R., Whitten, B. et al. (2004) Guar seed b-mannan synthase is a member of the cellulose synthase super gene family. Science, 303, 363–366. Duchesne, L.C. and Larson, D.W. (1989) Cellulose and the evolution of plant life. Bioscience, 39, 238–241. Ebringerova, A. and Heinze, T. (2000) Xylan and xylan derivatives – biopolymers with valuable properties. 1 – Naturally occurring xylans: structures, isolation procedures and properties. Macromol. Rapid Commun. 21, 542–556. Edwards, M.E., Dickson, C.A., Chengappa, S., Sidebottom, C., Gidley, M.J. and Reid, J.S.G. (1999) Molecular characterization of a membrane-bound galactosyltransferase of plant cell wall matrix polysaccharide biosynthesis. Plant J. 19, 691–697. Edwards, M.E., Marshall, E., Gidley, M.J. and Reid, J.S.G. (2002) Transfer specificity of detergent-solubilized fenugreek galactomannan galactosyltransferase. Plant Physiol. 129, 1391–1397. Edwards, M.E., Choo, T.S., Dickson, C.A., Scott, C., Gidley, M.J. and Reid, J.S.G. (2004) The seeds of Lotus japonicus lines transformed with sense, antisense, and sense/antisense galactomannan galactosyltransferase constructs have structurally altered galactomannans in their endosperm cell walls. Plant Physiol. 134, 1153–1162. Faik, A., Price, N.J., Raikhel, N.V. and Keegstra, K. (2002) An Arabidopsis gene encoding an a-xylosyltransferase involved in xyloglucan biosynthesis. Proc. Natl Acad. Sci. USA, 99, 7797–7802. Fengel, D. and Wegener, G. (1989) Wood-Chemistry, Ultrastructure, Reactions. Berlin: Walter de Gryter. Fenning, T.M. and Gershenzon, J. (2002) Where will the wood come from? Plantation forests and the role of biotechnology. Trends Biotechnol. 20, 291–296. Field, C.B., Behrenfeld, M.J., Randerson, J.T. and Falkowski, P. (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 281, 237–240. Fry, S.C., Smith, R.C., Renwick, K.F., Martin, D.J., Hodge, S.K. and Matthews, K.J. (1992) Xyloglucan endotransglycosylase, a new wall-loosening enzyme activity from plants. Biochem. J. 282, 821–828. Goubet, F., Misrahi, A., Park, S.K., Zhang, Z.N., Twell, D. and Dupree, P. (2003) AtCSLA7, a cellulose synthase-like putative glycosyltransferase, is important for pollen tube growth and embryogenesis in Arabidopsis. Plant Physiol. 131, 547–557. Grabber, J.H., Ralph, J. and Hatfield, R.D. (2002) Model studies of ferulate-coniferyl alcohol cross-product formation in primary maize walls: implications for lignification in grasses. J. Agric. Food Chem. 50, 6008–6016. Hayashi, T. (1989) Xyloglucans in the primary cell wall. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 139–168. Hazen, S.P., Scott-Craig, J.S. and Walton, J.D. (2002) Cellulose synthase-like genes of rice. Plant Physiol. 128, 336–340. Hematy, K., Sado, P.E., Van Tuinen, A., Rochange, S., Desnos, T., Balzergue, S., Pelletier, S., Renou, J.P. and Höfte, H. (2007) A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr. Biol. 17, 922–931. Hernandez-Blanco, C., Feng, D.X., Hu, J. et al. (2007) Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance. Plant Cell, 19, 890–903. Himmel, M.E., Ding, S.Y., Johnson, D.K., Adney, W.S., Nimlos, M.R., Brady, J.W. and Foust, T.D. (2007) Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science, 315, 804–807. Houghton, J., Weatherwax, S. and Ferrell, J. (2006) Breaking the Biological Barriers to Cellulosic Ethanol: A Joint Research Agenda. Washington: US Department of Energy Office of Science and Office of Energy Efficiency and Renewable Energy, http:// www.doegenomestolife.org/biofuels. Hu, W.J., Harding, S.A., Lung, J., Popko, J.L., Ralph, J., Stokke, D.D., Tsai, C.J. and Chiang, V.L. (1999) Repression of lignin biosynthesis promotes cellulose accumulation and growth in transgenic trees. Nat. Biotechnol. 17, 808–812. Huber, G.W., Chheda, J.N., Barrett, C.J. and Dumesic, J.A. (2005) Production of liquid alkanes by aqueous-phase processing of biomass-derived carbohydrates. Science, 308, 1446–1450. Humphrey, T.V., Bonetta, D.T. and Goring, D.R. (2007) Sentinels at the wall: cell wall receptors and sensors. New Phytol. 176, 7–21. Iwai, H., Masaoka, N., Ishii, T. and Satoh, S. (2002) A pectin glucuronyltransferase gene is essential for intercellular attachment in the plant meristem. Proc. Natl Acad. Sci. USA, 99, 16319– 16324. Jacobs, A.K., Lipka, V., Burton, R.A., Panstruga, R., Strizhov, N., Schulze-Lefert, P. and Fincher, G.B. (2003) An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell, 15, 2503–2513. Jin, S.Y. and Chen, H.Z. (2007) Near-infrared analysis of the chemical composition of rice straw. Indust. Crops Prod. 26, 207–211. Kohorn, B.D., Kobayashi, M., Johansen, S., Riese, J., Huang, L.F., Koch, K., Fu, S., Dotson, A. and Byers, N. (2006) An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 46, 307–316. Krupkova, E., Immerzeel, P., Pauly, M. and Schmulling, T. (2007) The tumorous shoot development2 gene of Arabidopsis encoding a putative methyltransferase is required for cell adhesion and co-ordinated plant development. Plant J. 50, 735–750. Lane, D.R., Wiedemeier, A., Peng, L.C. et al. (2001) Temperaturesensitive alleles of RSW2 link the KORRIGAN endo-1,4-beta-glucanase to cellulose synthesis and cytokinesis in Arabidopsis. Plant Physiol. 126, 278–288. ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 Cell-wall carbohydrates 567 Larsson, S., Reimann, A., Nilvebrant, N.O. and Jonsson, L.J. (1999) Comparison of different methods for the detoxification of lignocellulose hydrolyzates of spruce. Appl. Biochem. Biotechnol. 77, 91–103. Lerouxel, O., Cavalier, D.M., Liepman, A.H. and Keegstra, K. (2006) Biosynthesis of plant cell wall polysaccharides – a complex process. Curr. Opin. Plant Biol. 9, 621–630. Liepman, A.H., Wilkerson, C.G. and Keegstra, K. (2005) Expression of cellulose synthase-like (Csl) genes in insect cells reveals that CslA family members encode mannan synthases. Proc. Natl Acad. Sci. USA, 102, 2221–2226. Liepman, A.H., Nairn, C.J., Willats, W.G.T., Sørensen, I., Roberts, A.W. and Keegstra, K. (2007) Functional genomic analysis supports conservation of function among cellulose synthase-like A gene family members and suggests diverse roles of mannans in plants. Plant Physiol. 143, 1881–1893. Lieth, H. (1975) Primary production of the major vegetation units of the world. In Primary Production of the Biosphere (Lieth, H. and Whittaker, R.H., eds). Berlin: Springer-Verlag, pp. 203–215. Lynd, L.R., Wyman, C.E. and Gerngross, T.U. (1999) Biocommodity engineering. Biotechnol. Prog. 15, 777–793. Magid, J., Luxhoi, J. and Lyshede, O.B. (2004) Decomposition of plant residues at low temperatures separates turnover of nitrogen and energy rich tissue components in time. Plant Soil, 258, 351– 365. Meier, H. and Reid, J.S.G. (1982) Reserve polysaccharides other than starch in higher plants. In Encyclopedia of Plant Physiology, Vol. 13A (Loewus, F.A. and Tanner, W., eds). Berlin: Springer, pp. 418–471. Mølhoj, M., Pagant, S.R. and Hofte, H. (2002) Towards understanding the role of membrane-bound endo-beta–1,4-glucanases in cellulose biosynthesis. Plant Cell Physiol. 43, 1399–1406. Nishitani, K. (1997) The role of endoxyloglucan transferase in the organization of plant cell walls. Int. Rev. Cytol. 173, 157–206. Pagant, S., Bichet, A., Sugimoto, K., Lerouxel, O., Desprez, T., McCann, M., Lerouge, P., Vernhettes, S. and Höfte, H. (2002) KOBITO1 encodes a novel plasma membrane protein necessary for normal synthesis of cellulose during cell expansion in Arabidopsis. Plant Cell, 14, 2001–2013. Paredez, A.R., Somerville, C.R. and Ehrhardt, D.W. (2006) Visualization of cellulose synthase demonstrates functional association with microtubules. Science, 312, 1491–1495. Park, Y.W., Tominaga, R., Sugiyama, J., Furuta, Y., Tanimoto, E., Samejima, M., Sakai, F. and Hayashi, T. (2003) Enhancement of growth by expression of poplar cellulase in Arabidopsis thaliana. Plant J. 33, 1099–1106. Park, Y.W., Baba, K., Furuta, Y., Iida, I., Sameshima, K., Arai, M. and Hayashi, T. (2004) Enhancement of growth and cellulose accumulation by overexpression of xyloglucanase in poplar. FEBS Lett. 564, 183–187. Pauly, M., Andersen, L.N., Kauppinen, S., Kofod, L.V., York, W.S., Albersheim, P. and Darvill, A. (1999) A xyloglucan-specific endobeta–1,4-glucanase from Aspergillus aculeatus: expression cloning in yeast, purification and characterization of the recombinant enzyme. Glycobiology, 9, 93–100. Peña, M.J., Zhong, R., Zhou, G.-K., Richardson, E.A., O’Neill, M.A., Darvill, A.G., York, W.S. and Ye, Z.-H. (2007) Arabidopsis irregular xylem8 and irregular xylem9: implications for the complexity of glucuronoxylan biosynthesis. Plant Cell, 19, 549–563. Persson, S., Caffall, K.H., Freshour, G., Hilley, M.T., Bauer, S., Poindexter, P., Hahn, M.G., Mohnen, D. and Somerville, C. (2007a) The Arabidopsis irregular xylem8 mutant is deficient in glucuronoxylan and homogalacturonan, which are essential for secondary cell wall integrity. Plant Cell, 19, 237–255. Persson, S., Paredez, A., Carroll, A., Palsdottir, H., Doblin, M., Poindexter, P., Khitrov, N., Auer, M. and Somerville, C.R. (2007b) Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl Acad. Sci. USA, 104, 15566–15571. Pilling, E. and Höfte, H. (2003) Feedback from the wall. Curr. Opin. Plant Biol. 6, 611–616. Poorter, H. and Villar, R. (1997) The fate of acquired carbon in plants: chemical composition and construction costs. In Plant Resource Allocation (Bazzaz, F.A. and Grace, J., eds). San Diego, CA: Academic Press, pp. 39–72. Refregier, G., Pelletier, S., Jaillard, D. and Höfte, H. (2004) Interaction between wall deposition and cell elongation in dark-grown hypocotyl cells in Arabidopsis. Plant Physiol. 135, 959–968. Reid, J.S.G., Edwards, M. and Dea, I.C.M. (1988) Enzymatic modification of natural seed gums. In Gums and Stabilizers for the Food Industry (Phillips, G.O., Wedlock, D.J. and Williams, P.A., eds). Washington DC: IRL Press, pp. 391–398. Richmond, T.A. and Somerville, C.R. (2000) The cellulose synthase superfamily. Plant Physiol. 124, 495–498. Richmond, T.A. and Somerville, C.R. (2001) Integrative approaches to determining Csl function. Plant Mol. Biol. 47, 131–143. Saloheimo, M., Paloheimo, M., Hakola, S., Pere, J., Swanson, B., Nyyssonen, E., Bhatia, A., Ward, M. and Penttila, M. (2002) Swollenin, a Trichoderma reesei protein with sequence similarity to the plant expansins, exhibits disruption activity on cellulosic materials. Eur. J. Biochem. 269, 4202–4211. Sanjuan, R., Anzaldo, J., Vargas, J., Turrado, J. and Patt, R. (2001) Morphological and chemical composition of pith and fibers from Mexican sugarcane bagasse. Holz als Roh- und Werkstoff. 59, 447–450. Scheible, W.-R. and Pauly, M. (2004) Glycosyltransferases and cell wall biosynthesis: novel players and insights. Curr. Opin. Plant Biol. 7, 285–295. Schubert, C. (2006) Can biofuels finally take center stage? Nat. Biotechnol. 24, 777–784. Sharples, S.C. and Fry, S.C. (2007) Radioisotope ratios discriminate between competing pathways of cell wall polysaccharide and RNA biosynthesis in living plant cells. Plant J. 52, 252–262. Skjot, M., Pauly, M., Bush, M.S., Borkhardt, B., McCann, M.C. and Ulvskov, P. (2002) Direct interference with rhamnogalacturonan I biosynthesis in Golgi vesicles. Plant Physiol. 129, 95–102. Somerville, C. (2006) Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol. 22, 53–78. Somerville, C., Bauer, S., Brininstool, G. et al. (2004) Toward a systems approach to understanding plant-cell walls. Science, 306, 2206–2211. Sørensen, S.O., Pauly, M., Bush, M., Skjot, M., McCann, M.C., Borkhardt, B. and Ulvskov, P. (2000) Pectin engineering: modification of potato pectin by in vivo expression of an endo-1,4–betaD-galactanase. Proc. Natl Acad. Sci. USA, 97, 7639–7644. Suzuki, S., Li, L.G., Sun, Y.H. and Chiang, V.L. (2006) The cellulose synthase gene superfamily and biochemical functions of xylemspecific cellulose synthase-like genes in Populus trichocarpa. Plant Physiol. 142, 1233–1245. Tijmensen, M.J.A., Faaij, A.P.C., Hamelinck, C.N. and van Hardeveld, M.R.M. (2002) Exploration of the possibilities for production of Fischer Tropsch liquids and power via biomass gasification. Biomass Bioenerg. 23, 129–152. Turner, S., Gallois, P. and Brown, D. (2007) Tracheary element differentiation. Annu. Rev. Plant Biol. 58, 407–433. Ulvskov, P., Wium, H., Bruce, D., Jorgensen, B., Qvist, K.B., Skjot, M., Hepworth, D., Borkhardt, B. and Sorensen, S.O. (2005) Biophysical consequences of remodeling the neutral side chains ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568 568 Markus Pauly and Kenneth Keegstra of rhamnogalacturonan I in tubers of transgenic potatoes. Planta, 220, 609–620. Verbruggen, M.A., Beldman, G. and Voragen, A.G.J. (1995) The selective extraction of glucuronoarabinoxylans from sorghum endosperm cell-walls using barium and potassium hydroxide solutions. J. Cereal Sci. 21, 271–282. Vissenberg, K., Martinez-Vilchez, I.M., Verbelen, J.P., Miller, J.G. and Fry, S.C. (2000) In vivo colocalization of xyloglucan endotransglycosylase activity and its donor substrate in the elongation zone of Arabidopsis roots. Plant Cell, 12, 1229–1237. Vissenberg, K., Fry, S.C., Pauly, M., Höfte, H. and Verbelen, J.P. (2005) XTH acts at the microfibril–matrix interface during cell elongation. J. Exp. Bot. 56, 673–683. Vorwerk, S., Somerville, S. and Somerville, C. (2004) The role of plant cell wall polysaccharide composition in disease resistance. Trends Plant Sci. 9, 203–209. Wagner, T.A. and Kohorn, B.D. (2001) Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell, 13, 303–318. Willats, W.G.T., Steele-King, C.G., McCartney, L., Orfila, C., Marcus, S.E. and Knox, J.P. (2000) Making and using antibody probes to study plant cell walls. Plant Physiol. Biochem. 38, 27–36. Yuan, S., Wu, Y.J. and Cosgrove, D.J. (2001) A fungal endoglucanase with plant cell wall extension activity. Plant Physiol. 127, 324– 333. ª 2008 The Authors Journal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 559–568