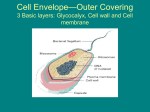
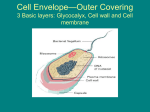
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
DNA vaccination wikipedia , lookup
Immune system wikipedia , lookup
Adaptive immune system wikipedia , lookup
Hygiene hypothesis wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Molecular mimicry wikipedia , lookup
Complement system wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Innate immune system wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
PEPTIDOGLYCAN RECOGNITION PROTEINS: MAJOR REGULATORS OF DROSOPHILA IMMUNITY PETER MELLROTH DEPARTMENT OF GENETICS, MICROBIOLOGY AND TOXICOLOGY STOCKHOLM UNIVERSITY 2005 Doctoral Thesis 2005 Department of Genetics, Microbiology and Toxicology, Stockholm University, 106 91 Stockholm, Sweden Abstract All eukaryotic organisms have an innate immune system characterized by germ-line encoded receptors and effector molecules, which mediate detection and clearance of microbes such as bacteria, fungi, and parasites. Vertebrate animals have, in addition to innate immune responses, evolved an adaptive immune system characterized by antibodies and T-cell receptors. Insects in general and the fruit fly in particular, have in recent years become an attractive model for the study of innate immunity. This is partly because it shares a great deal of similarity with the mammalian innate immune system. The versatility of Drosophila as a genetically tractable organism with a multitude of molecular-biology techniques available makes it an excellent tool for studying innate immunity. This thesis concerns the peptidoglycan recognition protein (PGRP) gene family in the fruit fly. The family consists of thirteen genes, of which a few have been reported to be part of the signaling pathways that regulates immune gene expression. PGRP proteins have affinity for peptidoglycan, an essential bacterial cell wall molecule. The main goal of this thesis has been to investigate which PGRP variants are required for immune pathway induction and which ligands are being recognized. Data presented show that the putative receptors have affinity for peptidoglycan, but not for lipopolysaccharide, or the fungal cell wall polymer beta-glucan. PGRP-SA, receptor of the Toll pathway, has a preference for peptidoglycan with a lysine in the crosslinking peptide. Furthermore PGRP-LCx, receptor of the Imd pathway, shows strong affinity for all types of peptidoglycan, both polymeric and monomeric. Activation of the Imd-pathway by polymeric, DAP-type, peptidoglycan is dependent on PGRP-LCx, whereas activation by monomeric peptidoglycan is dependent on both PGRP-LCx and PGRP-LCa. This result suggests that receptor dimerization is required for activation. Indeed, monomeric peptidoglycan can induce dimerization of the extracellular parts of PGRP-LCx and PGRP-LCa. However, PGRP-LCa does not have peptidoglycan affinity but is apparently only involved as an adapter molecule. These findings provide an explanation of the activation data for which a model is presented. In a search for novel PGRP receptors I found two PGRP proteins that instead displayed enzymatic activity towards peptidoglycan. They are of the N-actylmuramoyl Lalanine amidase type, which degrades peptidoglycan by splitting the crosslinking peptides from the glycan strands. The PGRP-SC1B-degraded Staphylococcus aureus peptidoglycan looses its immune elicitor capacity. This is in contrast to lysozyme-degraded peptidoglycan, which is still immune stimulating. Amino acid sequence homology showed as many as six of the thirteen Drosophila PGRPs to be potential enzymes. PGRP-SB1 is the other enzymatic PGRP described within this thesis. It has a more narrow activity spectrum as compared to SC1B implying that the fine-structure of peptidoglycan is important for the activity. In addition, PGRP-SB1 has antibacterial activity against Bacillus megaterium. In conclusion, receptor PGRP proteins binds bacterial peptidoglycan and triggers immune gene pathways and enzymatic PGRPs have the capacity to reduce the elicitor property of peptidoglycan. © 2005 Peter Mellroth ISBN 91-7155-105-0 (pp 1-58) Intellecta docusys AB, Stockholm 2005 The scientific theory I like best is that the rings of Saturn are composed entirely of lost airline luggage Mark Russell Table of contents Abstract .................................................................................................................................... II List of papers ........................................................................................................................ VII The need of an immune system ............................................................................................... 8 The insect immune system..................................................................................................... 10 Barrier epithelia............................................................................................................................................... 10 Blood ................................................................................................................................................................ 10 Humoral responses ...................................................................................................................................... 11 Cellular responses........................................................................................................................................ 13 Immune organs................................................................................................................................................. 14 Signaling pathways................................................................................................................. 14 The Toll pathway.............................................................................................................................................. 15 The Imd pathway .............................................................................................................................................. 16 Toll and Imd are the major immune pathways ................................................................................................. 17 Other pathways ................................................................................................................................................ 18 Pattern recognition................................................................................................................. 18 Microbial immune targets ..................................................................................................... 19 Teichoic acids................................................................................................................................................... 19 Lipopolysaccharide .......................................................................................................................................... 20 Fungal β–1,3-glucan ........................................................................................................................................ 20 Peptidoglycan................................................................................................................................................... 21 Peptidoglycan synthesis and turnover ......................................................................................................... 23 Peptidoglycan hydrolyzing enzymes ........................................................................................................... 27 Recognition molecules............................................................................................................ 28 C-type Lectins................................................................................................................................................... 28 Toll-like receptors ............................................................................................................................................ 29 Nod receptors ................................................................................................................................................... 29 Gram-negative binding proteins ...................................................................................................................... 30 Peptidoglycan recognition proteins ................................................................................................................. 30 PGRP in Drosophila: Expression pattern and protein topology predictions ............................................... 31 Functional studies of Drosophila PGRPs .................................................................................................... 32 PGRPs in other insects ................................................................................................................................ 33 Mammalian PGRPs ..................................................................................................................................... 34 PGRP structures........................................................................................................................................... 35 The present study ................................................................................................................... 38 Aim........................................................................................................................................... 38 A PGRP enzyme (Paper I) ................................................................................................................................ 38 Enzyme characterization (Paper I) ................................................................................................................... 39 A peptidoglycan scavenger (Paper I)................................................................................................................ 40 A bactericidal PGRP amidase (Paper II) ......................................................................................................... 41 LPS – to be, or not to be, an elicitor of the Drosophila immune system (Paper III and IV) ............................. 41 PGRP-LC – three of a kind (Paper III and IV)................................................................................................. 42 Polymeric peptidoglycan recognition (Paper III, IV, and V) ........................................................................... 43 Monomeric peptidoglycan recognition (Paper III, IV, and V) ......................................................................... 44 Concluding remarks............................................................................................................... 45 Acknowledgements................................................................................................................. 47 References ............................................................................................................................... 49 List of papers This thesis is based on the following publications: I Mellroth P., Karlsson J., Steiner H. (2003) A scavenger function for a Drosophila peptidoglycan recognition protein J. Biol. Chem. 278:7059-64 II Mellroth P., Steiner H. (2005) PGRP-SB1 – an N-acetylmuramoyl L-alanine amidase with antibacterial activity Manuscript III Werner T., Borge-Renberg K., Mellroth P., Steiner H., Hultmark D. (2003) Functional diversity of the Drosophila PGRP-LC gene cluster in the response to lipopolysaccharide and peptidoglycan J. Biol. Chem. 278:26319-22 IV Kaneko T., Goldman W.E., Mellroth P., Steiner H., Fukase K., Kusumoto S., Harley W., Fox A., Golenbock D., Silverman N. (2004) Monomeric and polymeric Gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway Immunity 20:637-49 V Mellroth P., Karlsson J., Håkansson J., Schultz N., Goldman W.E., Steiner H. (2005) Ligand-induced dimerization of Drosophila peptidoglycan recognition proteins in vitro Proc. Natl. Acad. Sci. USA 102:6455-60 The need of an immune system In the evolutionary race, all organisms have evolved means to defend the integrity of their genetic information contained in their genomes, which are under constant attack from viruses and other selfish genetic elements with a potential to modify or take control over the host’s DNA. Being sources of high energy molecules and nutrients, all eukaryotic organisms must also defend their cells and tissues from more faster propagating microorganisms. The enormous biodiversity among bacteria, however, set high demands on the host immune system in terms of detection and elimination. It must be specific enough to distinguish bacteria from the own cells, or the multitude of molecules produced by the own tissues. It must also be precise enough to kill the intruders without causing too much damage to the own tissues. The need of an efficient immune defense must certainly have been a cornerstone in the childhood of eukaryotic evolution and molecules with capacity to sense the presence of infectious microbes have been positively selected. A battery of molecules with capacity to eradicate all kinds of bacteria and fungi has also been contained in the evolutionary backpacks of the ancestors to the higher organisms that exist today. In common for the pre-existing eukaryotic life is the presence of an innate immune defense with a typical architecture. It is built on germ-line encoded sensors and effector molecules that recognizes and eliminates microbes that enter the host tissues. It is aiming at structural features of the microbial cell that has been seemingly unchanged during evolution. Even if distantly related organisms are using different molecules for detection of microbes the target epitopes are often the same. The recognition of a microbe is often linked to a signaling pathway that will direct the expression of genes that either encode effector molecules with capacity to kill the intruder, or encode cytokines and chemokines that engage other cell types in the microbial elimination process (Kimbrell and Beutler, 2001). It is striking that many factors of the innate immune system indeed are well conserved during evolution. A considerable number of genes encoding recognition molecules, as well as effector molecules, and components of signaling pathways, are conserved from organisms as divergent as humans and flies. These genes must thus have been present in the common ancestor to insects and mammals some 600 million years ago. In vertebrate animals, an adaptive immune system has evolved in addition to the innate immune defense. This adaptive immune system depends on B- and T-cells of lymphatic origin, which has the capacity to form antibodies and T-cell receptors through genetic rearrangement. In this elaborate process an 8 enormous potential of molecular recognition is accomplished allowing for detection of almost any possible microbial epitope. A major drawback of the adaptive immune system is that it can take up to three weeks until a full response has been established. However, once an intruder has been defeated, memory cells are formed which ensure a rapid response in case of repeated infections. The innate and adaptive systems works in parallel and the adaptive system partly depends on signals in form of cytokines and co-factors produced by cells of the innate system (Pasare and Medzhitov, 2004). A common theme for most organisms is that their immune responses are set in a resting-, or off-state, and is selectively activated whenever an infectious agent manages to pass an epithelium barrier. There are several reasons for this organization. One reason is that it is energetically costly to have a system that is always in the on-state and constantly producing immune effector molecules. Another reason is that most immune reactions are causing damages also to own tissues. In mammals, for example, an over-activated immune system can lead to a toxic-shock syndrome, often with fatal consequences. A third reason is that microbes can more easily evolve immune circumventing mechanisms if they are constantly being exposed to immune effector molecules. Such resting immune systems are dependent on efficient sensors of microbes that can selectively trigger the onset of the effector mechanisms. The microbial recognition event is thus of central importance. Bacteria are present everywhere in our environment. They have occupied all ecological niches and are specialists in extracting nutrients and energy from almost all imaginable sources. All high energetic biomasses like soil or dead animal corpses, lacking an active immune system, are soon overloaded with a multitude of decomposing bacteria. In our gut we have allied bacteria that help us to break down the food we ingest. They are adapted to this environment and also help us to exclude pathogens that first have to compete for space and nutrients with these commensal bacterial communities. However, these communities are so numerous that they by the tenfold exceed the number of cells in our own bodies and it is therefore essential to control these bacteria and limit their presence to the gut lumen. Also epidermal surfaces like our skin harbor complex bacterial societies that are mostly not harmful to us but have a potential to cause infection if they are getting access to a wound (Sansonetti, 2004). The physiological barriers that constitute the topological border dividing the organism’s interior from the exterior are guarded by the innate immune system. If bacteria manage to pass such a barrier, an elaborate alarm system is turned on triggering a multitude of responses aimed to track down and eliminate the intruders. The responses can be of several different types. Premade protein cascades can be triggered, specialized cells release the content of intracellular vesicles containing antibacterial factors, cells with a capacity to engulf bacteria are activated, and a synthesis of antimicrobial peptides can be 9 initiated. In mammals, small infections are often restricted to local and temporary inflammations but in insects a more systemic response is mounted. The bacterial detection event is critical to enable the innate immune system to mount a response. It is therefore astonishing that it relies only on some handful of receptors, but with a capacity to recognize almost all existing bacteria. The insect immune system Insects form the most successful (and sometimes irritating) animal group in terms of species diversity and abundance. To date about one million species are described, which is more than all other animal species combined, and they have occupied most terrestrial and aerial ecological niches on our planet. Part of their success is likely because they have evolved such an efficient immune defense. Many insects thrive in nutrient-rich environments such as soil or excrements with high contents of microbes and the need to efficiently recognize and battle bacteria is obvious. I will, as follows, briefly highlight the most important components of the fly immune system. Barrier epithelia The barrier epithelia serve as physiological shields against the environment and prevent microbes from entering interior tissues. The insect cuticle is a multilayered epithelium intertwined with protein matrixes and microfibers of chitin. It can be soft and flexible as in larvae or hard and sclerotized as in most adult insects. The digestive tract, tracheal tubules, and the genitals are also lined with epithelial tissues composed of cell layers and chitin/ protein matrixes, which secret factors like mucus and saliva. Cells of the midgut epithelium, for example, produces an extracellular matrix called the peritrophic membrane consisting of proteins, chitin, and proteoglycans and is working as a tough protective filter against microbes (Lehane, 1997). Certain epithelial tissues are also constitutively secreting antimicrobial peptides that aids in the protection (Ryu et al., 2004). Blood Bacteria that are getting access to the body cavities through wounds in the cuticle or by expressing virulence factors that enable passage through an epithelial barrier will reach the insect blood, named the hemolymph. The hemolymph is the main arena for the immune system and several different responses are involved in bacterial clearance. There are both cellular and humoral factors involved in the responses. 10 Insects have an open blood system with a heart that pumps the hemolymph to the body cavities surrounding the organs. The hemolymph consists of 10% hemocytes (blood cells) and 90% serum. The total cell concentration of the hemolymph is considerably lower than in human blood due to the lack of erythrocytes. In the absence of red blood cells, the appearance of the hemolymph ranges from clear to yellowish or greenish depending on insect species. Humoral responses The phenoloxidase system A melanization reaction can be triggered by the prophenol oxidase (PPO) cascade (Cerenius and Soderhall, 2004). Anyone who has injured a large insect has seen that the blood coming out is soon changing from clear to black. This blackening is due to the formation of the dark pigment melanin, which is the end product of the PPO cascade. The melanization reaction is considered to be an immune defense reaction even if the melanin itself is not especially harmful to microbes. However, some of the intermediates of the reaction are radicals and reactive oxygen species that might be harmful to bacteria. The reaction is catalyzed by the active phenoloxidase (PO) that catalyses oxidation of tyrosine to orthoquinones, which spontaneously polymerize to melanin. The enzymatically inactive PPO is cleaved into active PO in a process tightly controlled by serine proteases and their inhibitors. In many insects a pattern recognition receptor activates the cascade upon microbial infection (Ma and Kanost, 2000; Ochiai and Ashida, 1999; Yoshida et al., 1996; Yu et al., 1999) but in Drosophila no such receptor has been directly linked to PPO activation. However, it is shown that PPO activation is in part dependent on the Toll pathway (De Gregorio et al., 2002a; Ligoxygakis et al., 2002b) and that overexpression of PGRP-LE can induce melanization in larvae (Takehana et al., 2002). Melanin is deposited on parasites and becomes part of capsules and noodles formed around parasites by blood cells (see below). The prophenoloxidase cascade is present in insect and crustacean hemolymph but is not found in any deuterostome organism. Clotting The clotting system is another humoral system that can rapidly be activated if the cuticle is wounded (Bidla et al., 2005). Especially in larvae with its soft cuticle it is vital to stop the bleeding because the blood pressure makes them prone to leak. The molecular mechanism of how clot formation is triggered is not clear but it is evident that a soft clot is soon formed after wounding (Scherfer et al., 2004). This clot contains various proteins, including lectins but also ruptured blood cells. The soft clot will eventually be further cross-linked and 11 made hard by the melanization reaction. The clot will not only stop the blood from leaking out, it will also trap and hinder bacteria from entering the body cavities. Inducible antimicrobial peptides Even if the capacity of the insect to fight bacterial infections had been previously studied by scientists like Louis Pasteur and Elie Metchnikoff, the first biochemical analysis of anti-microbial factors in insect hemolymph was initiated by Hans G. Boman and colleagues in the late 1970s (Boman and Steiner, 1981). They discovered and characterized the first anti-microbial peptides and this became a starting point for a new field that was aiming to map the components of the insect immune system. The first anti-microbial peptide described was isolated from the Lepidoptera species Hyalophora cecropia and was named cecropin (Steiner et al., 1981). It was shown to be a cationic peptide of 37 amino acid residues having an unordered structure in solution. The net positive charge confers affinity for negatively charged bacterial surfaces. When cecropin binds to the bacterial membrane it folds into an ordered conformation consisting of two helices one of which being amphipathic (Steiner, 1982). Cecropin induces bacterial lysis because the peptide interferes with the integrity of bacterial membranes, presumably by forming leaky pores or patches. Cecropin has a broad spectrum of activity and potently kills both Gram-negative and Grampositive bacteria and is also active against fungi. In Drosophila, a total of seven different anti-microbial peptides plus isoforms are described (reviewed in (Imler and Bulet, 2005)). Attacin, a glycine-rich protein of 20 kDa is mainly active against Gram-negative bacteria and interferes with both bacterial membrane integrity as well as outer membrane protein gene expression (Carlsson et al., 1991). Drosocin and metchnikowin are proline-rich peptides with rather different antimicrobial spectra. Drosocin is a glycosylated peptide, which is active against Gram-negative bacteria whereas metchnikowin is active against Gram-positive bacteria and fungi (Bulet et al., 1993; Levashina et al., 1995). Diptericin is a 9 kDa peptide with a short proline-rich N-terminal domain and a glycine-rich Cterminal domain and is active against some Gram-negative bacteria (Wicker et al., 1990). Drosomycin and defensin are cysteine-rich peptides that form cyclic domains through disulfide bridge formation (Dimarcq et al., 1994; Fehlbaum et al., 1994). Drosomycin, an 8 kDa peptide containing four S-S bridges, has potent antifungal activity. In addition to these inducible peptides Drosophila also expresses seven lysozyme genes (Daffre et al., 1994). They are all of the classical lysozyme type cleaving the glycan strands of peptidoglycan by hydrolyzing the bond between N-acetyl-muramic acid (MurNAc) and N-acetylglucoseamine (GlcNAc) residues. 12 The bactericidal potency of the anti-microbial peptides was clearly illustrated in a study on Drosophila flies with deficient immune pathways. These flies are sensitive to bacterial infections but when manipulated to express a single anti-microbial peptide, an almost wild type phenotype was obtained (Tzou et al., 2002). Upon infection, a massive anti-microbial peptide expression is induced in the fatbody and the peptides are exported to the hemolymph. The expression is regulated by two signaling pathways, the Toll and the Imd pathway (reviewed in (Hultmark, 2003; Royet et al., 2005)). The Toll pathway mainly regulates the antifungal response, e.g. drosomycin expression, while the Imd pathway regulates the anti-bacterial peptide genes, e.g. diptericin and attacin. Even if the fatbody is the main site of expression, also blood cells are expressing antimicrobial peptides and contribute to the systemic humoral response (Johansson et al., 2005). In addition to the systemic response, barrier epithelial cells are capable of mounting a local inflammatory response involving antimicrobial peptide expression (Onfelt Tingvall et al., 2001; Tzou et al., 2000). In these tissues the immune gene induction is solely dependent on the Imd pathway. Some epithelial tissues also express anti-microbial peptides constitutively. The Drosophila lysozyme genes, for example, have a constitutive expression restricted to different parts of the gut (Daffre et al., 1994). This fact possibly reflects the lifestyle of the fly, which is often feeding on decaying fruits. Cellular responses Blood cells In Drosophila there are three classes of hemocytes described; plasmatocytes, lamellocytes, and crystal cells. The plasmatocytes is by far the most abundant cell type (Lanot et al., 2001). They are multi-functional cells that mostly resemble mammalian monocyte/ macrophage lineages. Being professional phagocytes, they are capable of efficiently engulf invading bacteria as well as own apoptotic cells. These are receptor-mediated responses and, at least partly, dependent on the expression of the PGRP-LC and croquemort receptors, respectively (Franc et al., 1996; Ramet et al., 2002b). Plasmatocytes are also involved in other immune-related responses such as synthesizing inducible antimicrobial peptides and producing important clotting factors. Recently they were also reported to be the primary producers of the endogenous peptide-cytokine spätzle (Irving et al., 2005). The crystal cells constitute about 5% of the blood cell population in larvae but are apparently absent in adult flies. These cells are characterized by having crystalline inclusion bodies of the prophenoloxidase enzyme required for the melanization reaction. Crystal cells are either circulating or sessile, sitting together with plasmatocytes in contact with the 13 cuticular epithelial layer (Lanot et al., 2001). If the epidermis gets wounded, the crystal cells will rupture and release their content to the surrounding tissues. This ensures a local melanization reaction at the wound-site and thus prevents the cytotoxic radicals produced to be formed throughout the hemocel. Lamellocytes are not present in healthy individuals but can form in lymph glands when larvae are under attack by parasites (Lanot et al., 2001). These cells are flat-shaped and are mainly involved in encapsulation of foreign objects that are too large to phagocytose. This specialized cell type has possibly evolved as a defense response against parasitic hymenoptera species that are specialized in laying their eggs in insect larvae. Lamellocytes moves towards and surround parasite eggs to form tough capsules together with melanin. Immune organs The fatbody is the major immune-related organ of insects. It is a functional equivalent of the mammalian liver and is the main site for synthesis of humoral factors taking part in the systemic immune response. Upon infection, a massive expression of immune genes is induced in the fatbody. Anti-microbial peptides are rapidly secreted to the hemolymph and can reach as high concentrations as 100 µM (Meister et al., 2000; Samakovlis et al., 1990). In the lymph glands, hematopoiesis (blood cell formation) takes place during larval stages, plasmatocytes proliferate, and lamellocytes differentiate under special conditions (Lanot et al., 2001). Signaling pathways The unraveling of how the immune-gene expression is controlled has been one of the main interests in the field during the last decennia. An important clue to how the gene-expression was regulated came when researchers examined the promoter regions for some anti-microbial peptide genes and found that they contained regulatory κB motifs (Engstrom et al., 1993; Sun and Faye, 1992). These motifs were known to be present in promoter regions in mammalian genes involved in inflammation responses controlled by nuclear factor κB (NFκB). This transcriptional activator is found in the cytoplasm bound to an inhibitory protein, IκB. When the signaling cascade is activated, the IκB protein is phosphorylated and degraded, leading to the unmasking of a nuclear import signal on NFκB. This promotes a translocation of NFκB from the cytoplasm to the nucleus where it binds to κB motifs and activates transcription. In Drosophila, three different Rel-homology proteins (homologous to NFκB) were found to interact with κB motifs and one of those, dorsal, had been shown to be an activator of the Toll-pathway, which defines the dorso-ventral polarity in Drosophila embryogenesis (Ip et al., 1991; Thisse et al., 1991). It was soon 14 evident that the same signaling pathway was utilized by the immune system in larval and adult stages but instead with another transactivator, named dorsalrelated immunity factor (Dif) (Ip et al., 1991; Ip et al., 1993; Lemaitre et al., 1996; Meng et al., 1999; Rosetto et al., 1995). Interestingly, most of the components in the Toll pathway are homologous to the interleukin-1 receptor pathway in mammals. These findings led to the important discovery of the Tolllike receptors (TLRs), which are now known to be cornerstones for microbial recognition by the mammalian innate immune system. The third Drosophila Rel-homology domain protein, Relish, was also shown to be implicated in the regulation of immune gene expression but is an activator of the other main signaling pathway, the immunity-deficiency pathway (Imd pathway). Today, many of the signal transducing components of the Toll- and Imd-pathways has been characterized (Fig.1). Figure 1. The Toll and Imd pathways. The Toll pathway The Toll signaling pathway and many of its components were characterized by Kathryn Andersson and colleagues in their studies of embryonic polarity establishment (reviewed in (Belvin and Anderson, 1996)). For immune gene 15 regulation, the pathway can be activated by receptor binding to bacterial peptidoglycan from many Gram-positive species, or by interactions with some species of fungi. For peptidoglycan activation the pathway requires peptidoglycan recognition protein (PGRP)-SA and Gram-negative binding protein (GNBP)-1, or PGRP-SD (Bischoff et al., 2004; Gobert et al., 2003; Michel et al., 2001; Pili-Floury et al., 2004). PGRP-SA is presumably acting together with Gram-negative binding protein-1 (GNBP1) in a recognition complex but PGRP-SD is seemingly independent of GNBP1. How the signal is transduced immediately after the recognition event is not clear. It is, however, likely that serine proteases are activated, which subsequently is leading to cleavage of the peptide-cytokine Spätzle to its active form. Spätzle then bind to the Toll-receptor in the cytoplasmatic membrane of fatbody cells. Binding of Spätzle to Toll induces receptor-dimerization and this in turn leads to the recruitment of the adaptor protein MyD88 to the cytoplamatic Toll-Interleukin-1 receptor (TIR) domains of Toll receptors (Sun et al., 2004). MyD88 interacts with the Toll receptors via its own TIR domain, and to the adaptor protein Tube and to Pelle via death domain interactions, and is thus initiating the formation of a signaling complex. Pelle is a serine-threonine kinase and upon activation it promotes phosphorylation of Cactus (an IκB homolog), which is bound to the transcription factor Dif (an NFκB homolog). Once phosphorylated, Cactus is targeted for degradation, and Dif is free to enter the nucleus and start transcription (Wu and Anderson, 1998). When the pathway is activated by fungi, the events that precedes Spätzle cleavage is following a different route than while activated by bacteria. The serine protease Persephone and the serine protease inhibitor Necrotic act upstream of Spätzle (Levashina et al., 1999; Ligoxygakis et al., 2002a) but the actual receptor(s) needed for pathway activation still remains to be found. Even if the Toll pathway is thoroughly studied there are still some question marks remaining. For example, it is not shown whether Pelle is actually phosphorylating Cactus or if there is another kinase(s) between Pelle and Cactus, and it is not clear if Dorsal is involved in immune gene activation, or if Dif is the only Toll-mediated transcription activator. The Imd pathway The Imd-pathway has many similarities with the mammalian Tumor-necrosis factor (TNF) receptor pathway. A major difference, however, is that the TNFreceptor itself is not present in the Imd pathway. Instead, membrane-spanning PGRP-LC receptors activate the pathway (Choe et al., 2002; Gottar et al., 2002; Ramet et al., 2002b) after binding to certain types of peptidoglycan (recognition/ activation specificities will be discussed later). These receptors have a PGRP domain outside the cytoplasmatic membrane, a single membrane-spanning helix, and a cytosolic ~30 kDa non-PGRP domain, which shares no significant 16 homology to any known protein. Downstream of PGRP-LC is the Imd protein and the corresponding gene gave name to the pathway when a null-mutant was found severely hampered in its antimicrobial peptide expression (Lemaitre et al., 1995). Recently, the Imd protein was shown to interact with the non-PGRP domain of PGRP-LC (Choe et al., 2005). The Imd protein has a C-terminal Death domain that interacts with the cytosolic protein FADD and the caspase Dredd. The N-terminal half of Imd interacts with the cytosolic non-PGRP domain of the PGRP–LC receptors and is required for pathway activation. The pathway has a branch-point downstream of the Imd protein and one branch activates FADD – Dredd while the other branch activates TAK1. TAK1 is a kinase and it is in turn constituting a new branch-point of which one branch leads to the activation of the IKK complex and the other to the activation of the Jun-N-terminal kinase (JNK) pathway controlling stress, and cellular immune responses (Silverman et al., 2003). The IKK complex is composed of the two subunits IKKβ (ird5) and IKKγ (kenny) (Lu et al., 2001; Rutschmann et al., 2000) and when activated it promotes phosphorylation of the NF-κB homolog Relish. Relish is a Rel-homology protein that has an N-terminal transcription activating domain and an internal IκB-like domain at the C-terminus. When phosphorylated, Relish is endoproteolytically cleaved (Stoven et al., 2000), presumably by the caspase Dredd after which the N-terminal domain translocates to the nucleus and initiates immune gene transcription. Toll and Imd are the major immune pathways How many signal transduction pathways are controlling the immune gene expression in Drosophila? Flies that are simultaneously mutated in the Toll-, and the Imd-pathway, are totally unresponsive in terms of anti-microbial peptide gene expression. Such flies were the subject of a gene microarray study, in which the immune response was compared to that of a wild-type fly and to flies defect in only one of the pathways (De Gregorio et al., 2002b). It was shown that the expression levels of nearly 300 genes were significantly affected (induced or repressed) by a septic injury in the wild-type fly and that 70% of these were affected in the mutant flies. Some genes are controlled by both pathways but most of the genes are dependent on only one of the pathways. Genes controlled by the Imd pathway includes most of the anti-microbial peptide genes but also many other immune related genes such as enzymes involved in the melanization process (Pale, Punch and Dhpr), prophenoloxidaseactivating enzyme (proPO-AE), and several PGRPs (-SD, -SB1, LB). Besides drosomycin, the Toll pathway controls expression of several serine proteases, serine protease inhibitors (serpins), and it also regulates the components of the Toll-pathway itself. However, it was also found that some immune related genes were not at all affected in the Toll/ Imd double mutants, suggesting that other pathways also regulate expression of at least some immune gene. As described 17 in the following section, at least two other immunity-related pathways are present. Other pathways In addition to the described NFκB signaling pathways there are at least two other signaling pathways involved in Drosophila immunity, the Janus kinase (JAK)/ signal transducers and activators of transcription (STAT) and the c-Jun N-terminal kinase (JNK) pathways. The immunological outcome of these two pathways is not as well-studied as that of the Toll and Imd pathways. In response to septic injury the JAK/ STAT pathway controls the production of at least two humoral factors synthesized in the fatbody, the thioester-contaning protein 1 (Tep1) and Turandot A (TotA) (Agaisse et al., 2003; Lagueux et al., 2000). Tep1 is a member of the complement C3/ α2-macroglobulin superfamily which has not been thoroughly investigated in Drosophila, but in Anopheles gambiae, the malaria vector, it has been shown to promote phagocytosis of Gram-negative bacteria (Levashina et al., 2001). The role of the Torandot genes is not known but their expression is upregulated in response to stress conditions (Ekengren and Hultmark, 2001). The JAK/ STAT pathway is also involved in blood cell proliferation and differentiation. The differentiation of hemocytes into lamellocytes during a parasitic wasp infection is clearly linked to this pathway (see (Agaisse and Perrimon, 2004) for a review). The JNK-pathway branches off from the Imd pathway and controls cytoskeletal, and proapoptotic genes, and is involved in wound healing (Boutros et al., 2002; Goberdhan and Wilson, 1998; Ramet et al., 2002a). In analogy with the Toll pathway both the JAK/ STAT and the JNK pathways are, in addition to the immunological involvement, also involved in the control of other processes during embryonic development, reviewed in (Luo and Dearolf, 2001; Noselli, 1998). Pattern recognition For many decennia immunologists were focused on antigen recognition by the adaptive immune system. In 1989, however, the so-called “pattern recognition concept” was formulated by Charles Janeway Jr (Janeway, 1989). He foresighted the existence of germ-line encoded pattern recognition receptors (PRR) that had capacity to detect molecular patterns present uniquely in microbial cells and consequently absent in the host. These “pathogen-associated molecular patterns”, or PAMPs, should be essential building blocks of the microbial cells and evolutionary conserved among large groups of microbes. This strategy enables a small number of germ-line encoded receptors to recognize an almost unlimited number of bacterial species. Eight year after publishing his theory, Janeway together with Ruslan Medzhitov, were involved in the discovery of the first pattern recognition receptor in mammalian host 18 defense, Toll-like receptor 4 (TLR4) (Medzhitov et al., 1997). This receptor was soon shown to have capacity to recognize lipopolysaccharide (Poltorak et al., 1998), a molecule present in the outer-membrane of Gram-negative bacterial cells. LPS fulfills the criteria for a good PAMP as it is an essential structural component of the outer membrane in Gram-negative bacterial cells. Moreover, LPS has for long been known to elicit conditions like inflammation and fever in mammals but the underlying mechanisms were still unknown. These initial findings opened many closed doors and a hunt for related pattern recognition receptors had started. Today, twelve different PRRs of two classes (TLRs and NODs) are described in mammalian host defense and they have the amazing capacity to recognize almost all different bacterial groups existing (reviewed in (Philpott and Girardin, 2004). After bacterial detection, a proinflammatory response is induced through NFκB activation, and the adaptive branch of the immune system is initiated through crosstalk with dendritic cells (Pasare and Medzhitov, 2004). The impact of the pattern recognition concept and discovery of the innate immune receptors is now appreciated as being one of the most important recent findings in the field of immunology. The term “pathogenassociated molecular patterns” should, however, not be associated with pathogens in particular but rather with microbes in general. Microbial immune targets A molecule present on a microbe must fullfill some basic criteria for being a good PAMP. It must be widespread among large groups of microorganisms and it must be fairly invariant during evolution. Typically, PAMPs are essential molecular building blocks of the microbial cell. In the following sections are examples of such immune target molecules. Teichoic acids Teichoic acid (Fig. 2) is an anionic polymer of glycerol-phosphate or ribitiolphosphate that is interwoven in the cell wall of Gram-positive bacteria and extends out and beyond the peptidoglycan layers. It can either be covalently connected to the peptidoglycan or have a lipid part that is anchored in the plasma membrane. Teichoic acids are functional analogs of lipopolysaccharide and confer a net negative charge to the bacterial cell. The degree of negative charge is modulated by the addition of protonated D-alanyl groups. Teichoic acids affect the traffic of ions, nutrients, proteins, and antibiotics across the cell wall (for an extensive review see (Neuhaus and Baddiley, 2003)). 19 Figure 2. Teichoic acid. Part of the polymeric molecule. Lipopolysaccharide The outer membrane of Gram-negative bacteria is a lipid bilayer, of which the inner leaflet is a phospholipid monolayer and the outer leaflet is a monolayer of lipopolysaccharide (LPS). The LPS molecule (Fig. 3) has a lipid part, lipid A. Connected to Lipid A is a core region consisting of various saccharides including 2-keto-3-deoxyoctulosonic acid (KDO), which is uniquely present among bacteria. Extending outside the core region is a polysaccharide O-chain, or O-antigen, which can be of variable composition and length. The composition of the Lipid A part and the core region is fairly invariant between bacterial species. It has for long been known that LPS induces inflammation and fever in humans. The microbial signature properties of LPS is strong since it is a ubiquitous and conserved molecule having unique features only found in bacteria. Figure 3. Structure of lipopolysaccharide, LPS. Fungal β–1,3-glucan Fungi form a diverse group of eukaryotic microorganisms, of which many species can cause infections in both invertebrates like Drosophila and in 20 vertebrates. A common fungal cell-wall molecule is β-1,3-glucan, which is a polymer of D-glucose in β(1,3) linkage with β(1,6) linked side chains of varying length and distribution. The structure is similar to that of chitin, which consists of D-glucose units coupled in β(1,4) linkage. Even if β-glucan is found in the cell walls of different plants (e.g. algae), it is not present in animals, which makes it a good target molecule for non-self recognition of fungal microorganisms (Brown and Gordon, 2003). Peptidoglycan Peptidoglycan is a polymeric macro-molecule present in the cell wall of almost all bacteria. It maintains the shape and gives tensile strength to the cell and it confers tolerance to changes in the osmotic pressure. Peptidoglycan is a network of unbranched glycan chains crosslinked with short peptides. Figure 4. General structure of peptidoglycan. Two tetrapeptides are linking glycan chains. A tetrapeptide is connected to the lactyl group of a MurNAc residue and to an adjacent tetrapeptide, either directly, or via an interpeptide bridge (shown as a dotted line). The glycan chains are composed of alternating residues of GlcNAc and MurNAc linked by β, 1→4 bonds (Fig. 4) and are typically 10 to 100 disaccharide units long (Harz et al., 1990). The peptide unit is composed of two 21 tetrapeptides (L-Ala – γ-D-Glu – DA – D-Ala) linked to the glycan chains via the amino group of L-Ala to the lactyl group of the MurNAc unit. The tetrapeptides, in turn, are linked to each other through the branching DA (diamino) group containing residue to the D-Ala residue. The DA residue is most commonly an L-Lys or a meso-diaminopimelic acid (DAP). Sometimes the two tetrapeptides are not directly linked but instead connected via an interpeptide bridge (Schleifer and Kandler, 1972). The general structure of peptidoglycan is much conserved among bacteria and the variation that is lies mainly in the tetrapeptide linkage region. The main differences are the identity of the third branching amino acid and the existence of an interpeptide bridge. However, the variation is rather small and large groups of bacteria have the same composition. Most common is the direct linked DAP-type peptidoglycan which is present in probably all Gram-negative species and in species belonging to Bacillaceae, Lactobacillaceae, Corynebacteriaceae, and Propionibacteriaceae. The second most common peptidoglycan type is those with an L-lysine residue in the third position of the tetrapeptide. Lys-type peptidoglycans are more heterogeneous because they often have an interpeptide bridge of variable but species-specific composition. For example, in Staphylococcus aureus the interpeptide bridge is composed of five glycine residues (Schleifer and Kandler, 1972). Besides these two major groups, DAP- and Lys-type peptidoglycan, other peptidoglycan variations include L-Orn, L,L-DAP, and hydroxyl-Lysine in the third tetrapeptide position. In rare cases the tetrapeptide linkage is instead at the D-Glu in second position. Peptidoglycan can be further modified. A common modification is amidation of carboxyl groups of meso-DAP or D-Glu and a less common is O-acetylation of muramic acid residues. The latter modification confers resistance to some peptidoglycan hydrolyzing enzymes (Bera et al., 2005; Schleifer and Kandler, 1972). The composition of the Gram-positive and Gram-negative cell wall is rather different. Gram-positive bacteria have a thick cell wall made up of 20 to 80 layers of peptidoglycan. In addition to peptidoglycan, the cell wall contains proteins and teichoic acids. In Gram-negative bacteria, the peptidoglycan is only one to three layers thick and is located in the periplasmatic space between the plasma membrane, and the outer membrane. The peptidoglycan layer is connected to the outer membrane by the covalently linked Brauns lipoprotein, which is imbedded in the inner leaflet of the outer membrane. The bacterial signature properties of peptidoglycan are indeed strong. MurNAc is a hexose, which is only present in bacteria. The presence of D-amino acid residues is also very rare among eukaryotes and the alternating motif of Dand L-amino acid residues in the peptidoglycan tetrapeptides is a feature unique to bacteria. The MurNAc – tetrapeptide motif can thus be considered as a bacterial bar-code and as such a perfect target for pattern recognition receptors. 22 Peptidoglycan synthesis and turnover When rod-shaped bacteria grow, the peptidoglycan sacculus is constantly rebuilt to enable elongation. In Gram-positive bacteria the cell wall biogenesis and turnover result in a shredding of peptidoglycan fragments to the environment. In most Gram-negative bacteria, on the other hand, an efficient peptidoglycan recycling pathway ensures that only 5% of the peptidoglycan fragments are released from the cell (Goodell and Schwarz, 1985; Greenway and Perkins, 1985; Park, 1995). There is an extensive peptidoglycan turnover during exponential growth whereby nearly half of the peptidoglycan is degraded and recycled to allow for the building of new glycan strands cross-linked with peptides. Peptidoglycan biosynthesis is an intricate process and the generally accepted model for Gram-negative bacteria invokes multimeric enzyme complexes comprising peptidoglycan synthesizing and degrading enzymes (Holtje, 1996b). Figure 5. Simplified view of the “3 for 1” model proposed for Gram-negative bacterial peptidoglycan biosynthesis. A multimeric enzyme complex digests one glycan strand and synthesizes three new strands in place. Dark-grey circles represent endopeptidases and lytic transglycosylases, which digest peptidoglycan and release the recycling fragments (the double-hexagone containing objects). The white ellipse and light grey circles represent glycosyltransferases and transpeptidases that incorporate new material transported from the cytoplasm. The figure is adapted from (Holtje, 1996a). 23 The synthesizing enzymes include transferases and high-molecular weight penicillin-binding proteins whereas the degrading enzymes comprise endopeptidases and lytic transglycosylases. The model for peptidoglycan synthesis (Fig. 5) suggests that one glycan strand is being removed while three new strands are being synthesized in place (Holtje, 1996a; Koch, 1995). The penicillin-binding proteins/ transpeptidases and glycosyltransferases incorporate peptidoglycan precursor molecules at the sites which have been made available by the action of the degrading enzymes. The glycan chains typically have a MurNAc residue in 1,6 anhydro conformation at the reducing end (Vollmer and Holtje, 2004). This feature is characteristic for Gram-negative bacterial peptidoglycan and is a consequence of lytic transglycosylases cleaving the glycan stand between MurNAc and GlcNAc residues. The major product produced by the digestion/ synthesizing machinery is a GlcNAc-(1,6anhydro)MurNAc-tetrapeptide unit (Fig. 6)(Park, 2001). Figure 6. Structure of the major peptidoglycan recycling molecule of Gramnegative bacteria, the GlcNAc-(1,6-anhydro)MurNAc-tetrapeptide, also known as tracheal cytotoxin, TCT. 24 This molecule is recycled in a controlled process (Fig. 7) and its initial uptake from the periplasm and transport into the cytoplasm is facilitated by a membrane permease, in E. coli named AmpG (Cheng and Park, 2002). In the cytoplasm the sugar moieties are cleaved off from the stem peptide by AmpD, a bacterial Nacetylmuramoyl L-alanine amidase (Jacobs et al., 1995). The tetrapeptide is then further processed to a tripeptide, and then rebuilt to a UDP-MurNAcpentapeptide (Uehara and Park, 2004). Figure 7. Peptidoglycan recycling and synthesis in Gram-negative bacteria. During synthesis part of the pre-existing peptidoglycan is digested into TCT (GlcNAc-(anhydro)MurNAc-tetrapeptides), which constitute the major turnover product. This fragment is transported to the cytoplasm via the permease AmpG. In the cyotosol AmpD cleaves off the two sugar moieties and further processing yields an L-Ala – γ-D-Glu – mesoDAP tripeptide. The tripeptide is successively transformed into UDP-MurNAc-pentapeptide with aid of MurF. This precursor molecule is transported across the membrane and utilized for de novo peptidoglycan biosynthesis. Even though the turn-over products are efficiently taken back to the cytoplasm a small portion (~5%) of the recycling-product is typically lost to the environment during growth (Park, 1995). Interestingly, peptidoglycan fragments has for long been reported to elicit a range of immunity related effects in humans including adjuvanticity, cytotoxity, induction of arthritis, and modulation of sleep (reviewed in (Boneca, 2005)). A suggestion presented in this thesis is that such 25 peptidoglycan fragments are major regulators of insect immunity as well. Some bacteria release peptidoglycan fragments more actively, e.g. Neisseria gonorrhoeae and Bordatella pertussis secret vast amounts of the GlcNAc – (1,6anhydro)MurNAc – tetrapeptide (Cundell et al., 1994; Melly et al., 1984). For these bacteria the peptidoglycan fragment is a virulence factor that inflicts damage to cells of the fallopian tubes (Neisseria) or to ciliated epithelial cells in the respiratory tract (Bordetella). In the latter case the peptidoglycan fragment is more known as tracheal cytotoxin (TCT) (Fig. 6) and is the major causative agent for the symptoms of whooping cough. It is possible that there exist a selective interplay between the host innate defense and the peptidoglycan biosynthesis machinery of pathogenic bacteria. A fact that speaks in favor for this is that some pathogens are modifying their peptidoglycan making it resistant to host lysozyme. A pathogenic strain of Streptococcus pneumoniae, for example, has been shown to modify the GlcNAc units in the glycan back-bone by deacetylation. This modification confers resistance to lysozyme and was shown to depend on one single gene (pgdA) (Vollmer and Tomasz, 2002). In another example, AmdD mutants of Salmonella enterica serovar Typhimurium that are accumulating TCT in the cytosol, are less prone to invade macrophages and are better inducers of iNOS than wildtype bacteria. This suggests that the peptidoglycan recycling pathway is involved in virulence (Folkesson et al., 2005). 26 Figure 8. Peptidoglycan digesting enzymes. The general structure of Gramnegative bacterial peptidoglycan is shown and arrows indicate which bonds are cleaved by the different enzymes. Peptidoglycan hydrolyzing enzymes Below follows a brief overview of peptidoglycan degrading enzymes present in bacteria and some examples of viral and eukaryotic counterparts. β-N-acetylmuramidase: Hydrolyses the β(1,4) bond between MurNAc and GlcNAc in the glycan chains and is involved in bacterial autolysis (Shibata et al., 2005; Turner et al., 2004). This enzyme type to which hen egg white lysozyme belongs is widely spread among eukaryotes. In humans, it is present in high concentrations in sera and saliva and participates in immune defense reactions (Vanderwinkel et al., 1995). β-N-acetylglucosaminidase: Hydrolyses the β(1,4) bond between GlcNAc and MurNAc in the glycan chains. In E. coli NagZ is a cytosolic enzyme involved in peptidoglycan recycling (Cheng et al., 2000; Votsch and Templin, 2000). Lytic transglycosidase: Cleaves the same β-glycosidic bond as β-Nacetylmuramidase but in addition concomitantly forms a 1,6-anhydromuramoyl residue. Works in concert with a D,D-endopeptidase in the digestion/ synthesizing enzyme complex to produce the GlcNAc – (1,6-anhydro)MurNActetrapeptide recycling product. The enzyme is present in most bacteria except for Gram-positives that only rarely have the enzyme for use in the sporulation 27 process (Atrih and Foster, 2001). This enzyme type has to my knowledge not been reported from any eukaryote organism. N-acetylmuramoyl L-alanine amidase (NAMLAA): AmpD in E. coli is involved in the recycling-process and is present in the cytoplasm (Jacobs et al., 1995) where it cleaves the amide bond that unites the stempeptide and the glycan portion in the recycled molecule. Bacteriophage T7 lysozyme is another example that utilizes the enzyme to lyse host cells and facilitate phage escape from infected E. coli cells (Cheng et al., 1994). NAMLAA enzymes will also be the subject of papers I and II. D,D-carboxypeptidase: Engaged in the peptidoglycan synthesis process. Cleaves off the terminal D-Ala from the GlcNAc – MurNAc - pentapeptide precursor molecule to facilitate incorporation and transpeptidation to an adjacent tetrapeptide. L,D-carboxypeptidase: Cleaves the bond between the meso-DAP and the terminal D-Ala in the same tetrapeptide. Cytoplasmic LdcA is involved in peptidoglycan recycling (Templin et al., 1999). PGRP-SA in Drosophila is an example of a eukaryotic protein with this enzyme activity and it has possibly an immunomodulatory function (Chang et al., 2004). D,D-endopeptidase: Cleaves the bond between the mesoDAP of one tetrapeptide and the terminal (fourth) D-Ala of the adjacent tetrapeptide. Works in concert with lytic transglycosylases in the digestion/ synthesizing enzyme complex to produce the GlcNAc-(1,6-anhydro)MurNAc-tetrapeptide recycling product (Holtje, 1996b; Romeis and Holtje, 1994a; Romeis and Holtje, 1994b). Lysostaphin is an example of another type of endopeptidase that is active on Staphylococcal peptidoglycan and has specificity for the Gly – Gly bonds of the pentaglycine interpeptide bridge. Recognition molecules This section gives a brief description of some of the most important groups of innate immune receptors from both vertebrates and invertebrates including a more thoroughly description of the peptidoglycan recognition proteins. C-type Lectins Lectins can bind to sugar residues of different kinds. Specifically engaged in immune defense are the C-type lectins, which have affinity for sugars that are common among microbes but rare in the host. Mannan-binding lectin is a vertebrate serum protein with collagen like thread structures and with carbohydrate recognition domains at the ends. It binds to bacterial surface sugar residues such as mannose, fucose, N-acetylglucosamine (Ma et al., 2004). Ligand binding activates associated serine proteases which in turn initiates the recruitment of complement factors. Dectin-1 is another example of a C-type 28 lectin that recognizes fungi by binding to β-glucan. After binding it associates with TLR-2 to induce a NF-κB response (Brown et al., 2003; Gantner et al., 2003). An example of an invertebrate C-type lectin is the immunolectin-2 of the tobacco hornworm Manduca sexta, which specifically binds to LPS and initiates the prophenoloxidase cascade (Yu and Kanost, 2004). Toll-like receptors The Toll-like receptors (TLRs) has already been introduced in a previous section. For a more extensive review of TLRs including ligand specificity I refer to (Takeda and Akira, 2005). There are ten TLR homologues in the human genome data base and they all encode integral membrane proteins. The extracellular domain consists of leucine-rich repeats (LRR) and the cytoplasmic domain, called the Toll/ IL-1 receptor (TIR) domain, associates with adapter proteins of the NF-κB signaling pathway. The TLR can either be anchored in the plasma membrane or in the membrane of intracellular endosomes. The LRR domains interact with the bacterial associated ligands and each TLR displays its own unique affinity. Additional co-factors are in some cases also required for recognition. The LPS receptor TLR-4 for example, is dependent on the adaptor proteins CD14 and MD-2 to be fully active (da Silva Correia et al., 2001). In some cases the recognition event requires receptor heterodimerization. TLR-2 associates with both TLR-1 and TLR-6 to detect lipopeptides (Takeda et al., 2002). TLR-3, -7, -8, and -9 are associated with endosomes and recognizes bacterial or viral nucleic acids (Alexopoulou et al., 2001; Heil et al., 2004; Hemmi et al., 2000). TLR-2 was initially thought to be a peptidoglycan receptor but it is now believed to recognize lipoteichoic acids and lipoproteins (Hirschfeld et al., 1999; Hirschfeld et al., 2000; Schwandner et al., 1999). Not only are the TLRs mediating an innate immune response but they are also cross-talking with the adaptive branch of the immune system. Especially dendritic cells are engaged in this cross-talk since they process/ present antigen and express TLR-mediated co-stimulatory molecules (Takeda and Akira, 2005). Nod receptors The nucleotide-binding oligomerisation domain (Nod) proteins are cytosolic peptidoglycan receptors of the vertebrate innate immune system. They consists of a C-terminal LRR domain, a central nucleotide binding domain, and one (Nod1) or two (Nod2) caspase-activating and recruitment domains at the Nterminus. Both Nod1 and Nod2 recognize peptidoglycan but they differ in the specific epitope requirement. Nod1 binds the γ-D-Glu-meso-DAP dipeptide whereas Nod2 instead recognizes a MurNAc-L-Ala-γ-D-Glu subunit. These specificities define Nod2 to be a more general receptor and Nod1 to be restricted to sense bacteria having DAP-type peptidoglycan. Upon ligand binding Nod 29 receptors, just like TLRs, triggers an NFκB response. Interestingly, mutations in Nod2 have been implicated Crohn’s disease, a chronic inflammatory disease in the intestine, and in the Blau syndrome, which is an inflammatory disease affecting joints and eyes (see (Philpott and Girardin, 2004; Royet and Reichhart, 2003) for reviews on Nods). Gram-negative binding proteins The Gram-negative binding protein (GNBP)/ β-glucan recognition protein family was originally discovered in lepidopteran species where protein members were isolated and characterized biochemically (Ma and Kanost, 2000; Ochiai and Ashida, 1988). The proteins contain an N-terminal glucan binding domain and a C-terminal domain that shares homology to bacterial glucanases. However, the catalytic amino acid residues required for enzymatic activity are not retained (Zhang et al., 2003). Members of this protein family have been reported to bind LPS in Bombyx mori and β-1,3-glucan M. sexta. The Drosophila genome comprises three GNBP members and one of these, GNBP1, is part of the Toll pathway (Gobert et al., 2003; Kim et al., 2000; Pili-Floury et al., 2004). Somewhat surprising is that even though this protein has affinity for both LPS and β-1,3-glucan it is required for the induction with Gram-positive bacteria, which do not contain these molecules. However, it is not clear if the protein takes part in the recognition event or if it is mediating the signal from the PGRP-SA receptor. The expression of a GNBP protein has been found specifically uppregulated in the gut region of A. gamiae in response to a Plasmodium falciparum infection (Tahar et al., 2002). Peptidoglycan recognition proteins PGRP is the most studied pattern recognition receptor in insects. The following sections surveys the PGRP research of past and present. The PGRP-story of our laboratory starts in 1995 when Håkan Steiner and Daiwu Kang, a Chinese PhD student, decided to investigate how insects defend themselves against virus infections. They planned to set up a differential display screen with the aim to identify the genes that were up- and downregulated by viral infections. For training purposes a screen was initiated using bacteria instead of viruses and larvae from the Lepidopteran species Trichoplusia ni were infected. Differentially expressed were identified using data-base mining and homology searches (Kang et al., 1996). One of the cloned genes had similarities a protein from the silk moth B. mori. A few N-terminal residues of had been sequenced of this protein and it was named peptidoglycan recognition protein (PGRP) (Yoshida et al., 1986). This protein could bind 30 peptidoglycan and was required for prophenoloxidase cascade activation. A similar protein could be a good candidate also for a receptor controlling antimicrobial peptide genes. The interest in PGRP increased when the laboratory cloned homologous genes from mouse and man. The T. ni PGRP and a mouse homolog was expressed and purified and both proteins did show peptidoglycan affinity (Kang et al., 1998). Since mice are devoid of the PPO cascade, the function of at least that particular PGRP variant could not be the same as in B. mori. It was found that PGRP is anciently related to bacteriophage T7 lysozyme, a peptidoglycan degrading enzyme. However, no such activity could be recorded for the mouse or the T. ni PGRP. The lack of enzymatic activity pointed instead towards a possible receptor function for these proteins. Daiwu Kang also cloned a PGRP homolog from Drosophila melanogaster, later named PGRP-SA. The flanking sequence placed the gene adjacent to an RNA polymerase in the genome. Collaboration was initiated with Dan Hultmark’s group in Umeå to make a PGRP mutant fly. However, it was soon evident that the Drosophila genome contains several PGRP genes and the risk was therefore obvious that a knock-out of a single PGRP would lack a phenotype. Instead, a general characterization of all the Drosophila PGRPs was performed including tissue distribution and possible inducibility by bacterial fragments (Werner et al., 2000). PGRP in Drosophila: Expression pattern and protein topology predictions When the Drosophila genome was completely sequenced in 2000, it was found to harbor a total of 13 PGRP genes (Khush and Lemaitre, 2000). They have been grouped into two groups according to the length of their transcripts, the short (S), and the long (L) forms (Werner et al., 2000). There are seven short forms which all encode proteins that contain the conserved PGRP domain flanked by an N-terminal signal peptides that target these proteins for cellular export. The short forms include PGRP-SA, -SB1, -SB2, -SC1A, -SC1B, -SC2, and -SD. The majority of the short PGRPs are expressed in the fatbody and the expression is often up-regulated upon bacterial infection. The N-terminal signal sequence and the fatbody expression strongly suggest that these proteins are secreted into the hemolymph. Some short forms (PGRP-SC) are also expressed in the gut region. An export to the gut lumen is likely and the proteins are possibly taking part in bacterial decomposition. Another possibility is that the proteins take part in the defense of the barrier epithelium lining the gut. The longer forms, which typically contain another domain in addition to the PGRP part, include PGRP-LA, -LB, -LC, -LD, -LE, and -LF. The additional non-PGRP domains differ in amino acid length and they neither share homology to each other nor to any other protein. The amino acid sequences of PGRP-LA, -LC, and -LF contain putative membrane spanning regions and intracellular non-PGRP domains, in addition to the extracellular PGRP domain. 31 Membrane localizations for these forms are therefore likely and have been experimentally shown for PGRP-LC (Paper IV, V). PGRP-LF is unique since it is the only Drosophila PGRP that contains two PGRP domains, located in tandem outside the putative transmembrane domain. Topology predictions for the other long forms are more uncertain. The cDNA for PGRP-LB encodes a short protein more similar to the short (S) forms. However, it does not have any obvious signal peptide sequence and was therefore initially predicted to be an intracellular protein (Werner et al., 2000). The protein sequences of PGRP-LD, and –LE, both contains a non-PGRP part connected to the amino-terminal of the PGRP domain, and also for these proteins are no signal peptide present. However, an investigation shows that when PGRP-LE is overexpressed, the recombinant protein is anyhow secreted to the hemolymph (Takehana et al., 2002). Functional studies of Drosophila PGRPs The capacity of PGRP to bind the bacterial cell wall polymer peptidoglycan points by itself toward functions involved in immune reactions. Also the bacteria-inducible expression patterns of most PGRPs and the localized expression in immune related tissues strongly corroborates an involvement in the immune defense (Werner et al., 2000). The first Drosophila PGRP to be assigned a function was PGRP-SA. It was found that the protein was required for Toll pathway activation (Michel et al., 2001). Epistatic gene knockout experiments placed PGRP-SA upstream of Toll and showed that it is required for down-stream processing of the peptide cytokine Spätzle, presumably by activating a series of serine proteases. PGRPSA is thus a soluble receptor involved in recognition of certain types of Grampositive bacteria such as Micrococcus luteus and Streptococcus faecalis and knock-out flies are more susceptible to infections by these bacteria. PGRP-SA has later been suggested to be engaged in an extracellular complex-formation with GNBP-1 (Gobert et al., 2003; Pili-Floury et al., 2004). Moreover, PGRPSA have been found to possess L,D-carboxypeptidase enzymatic activity (Chang et al., 2004). This might have an immunomodulatory function important for Toll-pathway signaling but its particular role needs to be further investigated. PGRP-SD is another short form that has also been reported to be an extracellular factor of the Toll pathway (Bischoff et al., 2004), in parallel to PGRP-SA and GNBP-1. The PGRP-SD mutant is hampered in the response to infections of S. aureus and Streptococcus pyogenes and shows partial redundancy with PGRP-SA/ GNBP1. PGRP-SC1B, and –SB1, are characterized in Paper I and II, respectively, and will be discussed later in this thesis. No functional study has yet been done for the short forms PGRP-SB2, SC1A, and SC2, but they are further discussed in Paper I and II. 32 PGRP-LC was the first of the long membrane-bound forms to be functionally characterized. It was shown to be a receptor of the Imd-pathway upstream of the Imd protein and was absolutely required for pathway activation. This finding was reported in three independent studies in April 2002 (Choe et al., 2002; Gottar et al., 2002; Ramet et al., 2002b). The PGRP-LC knockout flies were highly susceptible to Gram-negative bacterial infections. One of the studies (Ramet et al., 2002b), showed that RNAi knock-down of PGRP-LC expression impaired a Drosophila Scheider-2 cell line in the phagocytic response against E. coli cells. The PGRP-LC gene locus is somewhat complex and three different gene products can be formed by alternative splicing. The explicit role of the different spliceforms as Imd-receptors, and biochemical data on peptidoglycan affinities is the subject of Paper III, IV, and V. PGRP-LE has also been reported to function as a receptor for the Imd-pathway and genetic epistasis experiments place the protein, somewhat remarkable, both upstream and in parallel with PGRP-LC (Takehana et al., 2004). PGRP-LE binds DAP-type peptidoglycan and its overexpression in Drosophila larvae leads to melanin formation which implies a possible role as a receptor also for the PPO cascade (Takehana et al., 2002). PGRP-LB was the first PGRP for which the crystal structure was solved. It was also shown to possess N-acetylmuramoyl L-alanine amidase activity and is thereby a peptidoglycan degrading enzyme (Kim et al., 2003). No other functional study of PGRP-LB has been carried out. The possible involvement of PGRP-LA and PGRP-LD in Imdpathway activation or phagocytosis of bacteria has been investigated using RNA-interference (RNAi) to knock-down their expression. However, no clear phenotypes could be detected (Ramet et al., 2002b). RNAi against PGRP-LF gives a stronger Imd-mediated expression of antibacterial peptide genes upon bacterial challenge (Paper III and Carina Persson, personal communication). This suggests that the protein has an inhibitory effect to the Imd-pathway. PGRPs in other insects Drosophila melanogaster is by far the most studied insect. The understanding of its immune system is un-matched in any other insect species. Even though PGRP genes and proteins have been characterized in other insect species, it is not known if they play as central roles as they do in Drosophila. The malaria vector Anopheles gambiae that belongs to the same insect order as Drosophila (Diptera) harbors seven PGRP genes, of which many isoforms are remarkably similar to their Drosophila counterparts. A striking example is the PGRP-LC locus in which the organization of the exons producing three different spliceforms is fully conserved (Christophides et al., 2002). 33 As already mentioned, the first PGRP that was characterized was purified from hemolymph of the silk moth B. mori by Ashida’s group (Yoshida et al., 1996; Yoshida et al., 1986). They found that this 19 kDa protein binds to peptidoglycan and is required for peptidoglycan-induced activation of the prophenoloxidase cascade. The first PGRP cDNA was cloned from the cabbage looper T. ni and was performed in our laboratory (Kang et al., 1998). The gene showed high sequence similarity to bacteriophage T7 lysozyme and the protein has peptidoglycan affinity but no enzymatic activity. The functional role for the T. ni PGRP is still not fully understood. A PGRP was isolated from the hemolymph of the large beetle Holotrichia diomphalia and was found to bind both peptidoglycan and the fungal cell wall component β-1,3-glucan (Lee et al., 2004). Interestingly, it was shown to be required for pro-phenoloxidase activation by β-1,3-glucan but not by peptidoglycan. Mammalian PGRPs It was realized at an early stage of PGRP research that PGRP homologues were also present in mammalian genomes (Kang et al., 1998). Now when the human genome has been fully sequenced it is evident that it contains a total of four PGRP homologues. There is one short (S) form, one long (L) form, and two of intermediate (I) size (Dziarski, 2004). The PGRP-S is most prominently expressed in the bone marrow and in neutrophils. In murine neutrophils, PGRP-S is contained within intracellular vesicles and has been shown to inhibit growth of Gram-positive bacteria (Liu et al., 2000). A recent study (Cho et al., 2005) shows that also the human PGRP-S is localized in neutrophil granules and accordingly inhibits growth of both Gram-positive bacteria (S. aureus) and Gram-negative bacteria (E. coli). In addition, this protein shows synergistic antibacterial activity with lysozyme. A Russian research group has found PGRP-S expressed in murine tumors and suggested a cytokine function for the protein (Kiselev et al., 1998) and later they reported that it can form a cytotoxic complex together with heat-shock protein 70 (Sashchenko et al., 2004). The PGRP-Iα and PGRP-Iβ are putative membrane bound proteins with a rather short intracellular part. Both these proteins have a localized expression in the esophagus epithelium (Liu et al., 2001) of which relatively little is known in terms of innate immune recognition and defense. No function has yet been suggested for these proteins. However, the crystal structure of PGRP-Iα has been solved and will be discussed in a section below. PGRP-L contains a relatively large non-PGRP domain (~40 kDa) with two hydrophobic stretches that weakly indicates putative membrane spanning domains. It was therefore suggested to be a membrane bound protein 34 (Dziarski, 2004) and as such considered a strong immune receptor candidate. However, it is pre-dominantly expressed in the liver and has later been found to be an enzyme with peptidoglycan degrading activity (Gelius et al., 2003; Wang et al., 2003). It was also realized that PGRP-L is identical to the Nacetylmuramoyl L-alanine amidase, or NAMLAA, that had been purified from human sera and biochemically characterized in 1980s and -90s (Hoijer et al., 1998; Hoijer et al., 1997; Hoijer et al., 1996; Mollner and Braun, 1984; Valinger et al., 1982). These findings was recently reconfirmed (Zhang et al., 2005). A recent study showed that in addition to the tissues showing major PGRP expression, the expression of all four PGRPs could be markedly induced in an NF-κB dependent manner in oral epithelial cells when challenged with synthetic TLR- or Nod-ligands (Uehara et al., 2005). This tissue is a major site for bacterial entrance to our bodies and it typically harbor bacterial communities. However, not much is known of the interplay between these bacteria and the innate immune system. A bovine PGRP ortholog was isolated from neutrophil and eosinophil granules and was reported to possess antibacterial activity against Gram-positive and Gram-negative bacteria and surprisingly also against yeast (Tydell et al., 2002). Since yeast does not contain peptidoglycan, this protein was instead named bovine oligosaccharide-binding protein, bOBP. A camel PGRP has been isolated from milk and was suggested to function as an antibacterial factor (Kappeler et al., 2004). A rat PGRP has been found up-regulated when animals have been deprived from sleep (Rehman et al., 2001). Sleep-depravation can also be induced by peptidoglycan fragments (Johannsen et al., 1990). A functional role for the rat PGRP as a modulator of peptidoglycan-induced sleep deprivation was suggested (Rehman et al., 2001). PGRP structures The crystal structures of the Drosophila PGRP-LB and PGRP-SA and the human PGRP-IαC and PGRP-S have been solved (Chang et al., 2004; Guan et al., 2004a; Guan et al., 2004b; Guan et al., 2005a; Guan et al., 2005b; Kim et al., 2003). They all have a similar fold as phage T7 lysozyme (Cheng et al., 1994), which was expected considering the considerable amino acid sequence similarity. The general structure contains three α-helices and a central β-sheet, composed of four parallel strands and one anti-parallel strand (Fig. 9). However, PGRP-SA has some additional minor helices and β-strands (Chang et al., 2004). They all share a similar peptidoglycan-binding groove with the beta-sheet as a floor and with walls flanked by one of the helices and five of the loop-regions. The enzymatic PGRP-LB has, just like phage T7 lysozyme, a zinc ion bound to one cysteine and two histidine residues in the catalytic centre. The 35 metal co-factor is not included in the other PGRP structures. Common to all PGRP structures, but not present in T7 lysozyme, is an extra loop-region located on the back of the protein relative to the binding cleft. It has been suggested that this part could be important for protein – protein interactions and thus might facilitate PGRP dimerization or complex-formation such as that suggested for PGRP-SA and GNBP-1. Adapted from Kim et al. Nature Immunology (2003) 4,787-793 Figure 9. Figure show ribbon diagrams of PGRP-LB (left) and bacteriophage T7 lysozyme (right) structures. The zinc ion is symbolized by a ball, disulphide bonds shown in “ball and stick”, and PGRP specific segment in yellow. Also in contrast to T7 lysozyme, all PGRP structures contain disulphide bridges, of which at least one is seemingly conserved in all PGRPs except in Drosophila PGRP-LE (Guan et al., 2005b). This feature speaks against the proposed cytoplasmatic localization of some PGRPs lacking an N-terminal signal-peptide, because the reducing cytosolic environment will most likely disrupt disulphide bonds. 36 Protein D.m.PGRP-SA D.m.PGRP-SB1 D.m.PGRP-SB2 D.m.PGRP-SC1A D.m.PGRP-SC1B D.m.PGRP-SC2 D.m.PGRP-SD D.m.PGRP-LA D.m.PGRP-LB D.m.PGRP-LCa D.m.PGRP-LCx D.m.PGRP-LCy D.m.PGRP-LD D.m.PGRP-LE D.m.PGRP-LF B.m.PGRP H.d.PGRP C.d.PGRP B.t.PGRP, bOBP R.n.PGRP M.m.PGRP-S M.m.PGRP-L H.s.PGRP-S H.s.PGRP-Iα H.s.PGRP-Iβ H.s.PGRP-L Function Receptor of Toll pathway, L,D-carboxypeptidase activity. N-acetylmuramoyl L-alanine amidase (NAMLAA), antibacterial activity. Putative NAMLAA, mainly expressed in prepupal stage. Putative NAMLAA. NAMLAA, peptidoglycan scavenger Putative NAMLAA. Extracellular factor of the Toll pathway, putative receptor. Function unknown, putative membrane protein. NAMLAA. Putative adapter-protein to PGRP-LCx required for Imd-activation with monomeric peptidoglycan. Main receptor of Imd-pathway. Function unknown, membrane bound. Function unknown. Receptor of Imd-pathway and possibly involved in PPO activation. Function unclear, negatively affects Imd-pathway signaling, membrane bound, protein contains two PGRP-domains. Receptor for PPO cascade activation Ref (Chang et al., 2004; Michel et al., 2001) II I I I, (Gelius et al., 2003) I (Bischoff et al., 2004) (Werner et al., 2000) (Kim et al., 2003) III,IV,V III,IV,V III,IV (Werner et al., 2000) (Takehana et al., 2002; Takehana et al., 2004) III, Carina Persson (personal comm.) (Yoshida et al., 1996; Yoshida et al., 1986) Receptor for PPO cascade activation, binds β- (Lee et al., 2004) glucan and peptidoglycan Present in milk (Kappeler et al., 2004) Antibacterial against Gram-positive and Gram- (Tydell et al., 2002) negative bacteria and fungi Suggest involvement in sleep deprivation (Rehman et al., 2001) responses Bacteriostatic. Expressed in neutrophil granula (Liu et al., 2000) NAMLAA (Gelius et al., 2003) Bacteriostatic. Expressed in neutrophil granula (Cho et al., 2005) Function unknown. Expressed in esophagus (Liu et al., 2001) Function unknown. Expressed in esophagus (Liu et al., 2001) NAMLAA. Mainly expressed in liver. Present in (Wang et al., 2003) sera. Table 1. Overview of the functional properties of PGRPs. D.m. Drosophila melanogaster (Fruit fly), B.m. Bombyx mori (Silk moth), H.d. Holotrichia diomphalia (Korean Large beetle), C.d. Camelus dromedaries (Camel), B.t. Bos Taurus (Cow), R.n. Rattus norvegicus (Rat), M.m. Mus musculus (Mouse), H.s. Homo sapiens (Human). 37 The present study Aim The general aim of this thesis has been to investigate how the innate immune system recognizes bacteria to initiate immune gene expression using Drosophila as a model system. In particular, the involvement of peptidoglycan recognition proteins, PGRPs, as immune receptors was the subject of the investigation. However, the goal was extended during the course of the study to include the characterization of enzymatic PGRPs. A PGRP enzyme (Paper I) The first project was to express and purify recombinant Drosophila PGRP proteins and investigate their possible involvement as immune receptors or modulators of immune gene expression. At this time, many of the intracellular components of the Toll and Imd pathways were well-characterized but a receptor triggering the pathways had yet not been found. The Toll receptor itself was clearly not a bacterial receptor in striking contrast to the mammalian Tolllike receptors (TLRs), which had been shown to bind a repertoire of different microbial ligands. We imagined that the soluble Drosophila PGRPs could either function as opsonins that bind to bacteria and stimulate phagocytosis, or else that they could function as co-receptors to a membrane receptor, e.g. Toll. A similar function had been reported for the mammalian CD14 and LBP that binds LPS and work as co-receptors for TLR 4 (Guha and Mackman, 2001). One of our strategies was to assay immune gene induction in the Drosophila S2 cell line after challenge with bacterial products and to assay the expression of antibacterial peptide genes after addition of PGRP to the cell culture medium. Initial studies by Gang Liu had shown that pretreatment with PGRP-SA had an inhibitory effect on cecropin expression instead of the elevated expression that was expected if the protein had been a receptor (the result is fully explainable in light of later findings). To compare the inhibitory effect of PGRP-SA with another soluble PGRP, a cell line expressing PGRPSC1B was established. When I finally had expressed and purified enough protein to perform comparative studies with PGRP-SA, a paper appeared (Michel et al., 2001) showing PGRP-SA to be a soluble receptor of the Toll pathway. It was surprising that a knock-out of a soluble PGRP displayed such a clear phenotype because the Drosophila genome comprises six very similar 38 short putatively exported PGRP proteins. The knock-out mutant placed PGRPSA upstream of Spätzle. I felt confident that PGRP-SC1B also was a receptor that perhaps recognized some other type of peptidoglycan and I consequently compared the affinities of PGRP-SA and SC1B towards different peptidoglycan types. Insoluble peptidoglycan was incubated with protein and surprisingly, the reaction tubes with PGRP-SC1B was found to have a much smaller (or no) peptidoglycan pellet as compared to those with PGRP-SA present. We then recollected the relationship of PGRP to phage T7 lysozyme and its enzymatic nature. The realization that PGRP-SC1B possesses enzymatic activity readily explains the diminished peptidoglycan pellets. This finding was also the starting point of a new track of PGRP research. Enzyme characterization (Paper I) Insoluble peptidoglycan isolated from bacteria is a complex polymeric macromolecule of undefined size. Using peptidoglycan as an enzyme substrate limits the possibilities to define enzyme kinetic constants that typically require a defined substrate that is converted to a defined product possible to measure. To obtain a rough estimate of the enzymatic properties of PGRP-SC1B we analyzed the rate by which a turbid solution of peptidoglycan turns clear. We found that PGRP-SC1B indeed is an efficient enzyme with an activity in the same range as hen egg white lysozyme. The highest activity was recorded against S. aureus peptidoglycan and we found that the presence of teichoic acids had an inhibitory effect. PGRP-SC1B has a higher pH optimum than lysozyme and if the two enzymes are compared at pH 8.0 instead of pH 7.2, SC1B actually shows a higher activity against S. aureus peptidoglycan than lysozyme (unpublished data). The crystal structure of T7 lysozyme shows a mixed α/β conformation with a prominent cleft with a zinc ion bound to three amino acid ligands (Cheng et al., 1994). A mutational analysis pointed out two additional residues as being required for the amidase activity. Aligning the active-site residues of T7 lysozyme and PGRPs made evident that SC1B retains all five residues. Interestingly, in total six Drosophila PGRPs have these residues conserved and this is also true for the long mammalian PGRPs (PGRP-L). Amidase activity has later been shown also for Drosophila PGRP-LB and for the mouse and human PGRP-L (Gelius et al., 2003; Kim et al., 2003; Wang et al., 2003). Interesting to note was that none of the receptor PGRPs has all the five residues retained. A critical cysteine residue, being one of the zinc ligands, is replaced by a serine residue in the peptidoglycan binding receptor-PGRPs i.e. SA, SD, LE, LCx, and B. mori PGRP. We decided to examine what role this cysteine had for SC1B activity. By mutagenesis I did exchange it with a serine or an alanine residue. As expected, the cysteine was absolutely required for the enzymatic activity but the mutants had not lost their peptidoglycan binding capabilities. To more 39 specifically determine the enzymatic specificity of PGRP-SC1B, we undertook an analysis of the cleavage products by means of HPLC fractionation and determination of molecular masses by Maldi-TOF mass spectrometry. The mass spectrum shows a number of peaks but all correlate to products produced by the action of an N-acetylmuramoyl L-alanine amidase cleavage and it was firmly established that PGRP-SC1B has the same activity as T7 lysozyme. As described in Paper II we have also cloned and characterized PGRP-SB1, which is one of the amidase candidates postulated in Paper I. We experimentally demonstrate that it is an enzyme with N-acetylmuramoyl Lalanine amidase activity and that it has a preference for DAP-type peptidoglycan. This stands in contrast to PGRP-SC1B that has a broader activity spectrum and are highly active against peptidoglycan of both Lys-type e.g. S. aureus peptidoglycan, and of DAP-type e.g. E. coli peptidoglycan. The best substrates for PGRP-SB1 are peptidoglycan from B. megaterium and E. coli. A peptidoglycan scavenger (Paper I) The T7 lysozyme mediates lysis of the E. coli cell to facilitate the release of newly synthesized phage particles from the infected bacterium. To examine if PGRP-SC1B is bacteriolytic we extensively assayed antibacterial activity against a range of different bacteria but with negative results. What could the physiological role of a peptidoglycan degrading enzyme be if it was not antibacterial? As peptidoglycan is a highly immune-stimulatory macromolecule, a possible function could be to modify this property. We speculated that the degradation products could either give a stronger immune response than the intact cell wall because the ligands would more widely spread to cells and tissues, or else that the degradation products would be less stimulatory. To test this we challenged Drosophila cells with the SC1B degradation products and it was clear that the immune induction was much reduced as compared to intact peptidoglycan or lysozyme-degraded peptidoglycan. This observation led us to suggest a scavenger function for the protein. Peptidoglycan fragments will most likely spread systemically when bacterial lysis is caused by antibacterial peptides or other factors during an infection. A peptidoglycan scavenger function would be beneficial for the host in order to restrict the duration of the immune response and thereby limit the damages caused to own tissues. To bring levels of immune stimulatory molecules back to zero is likely a prerequisite for the fly to be able to respond to a second infection. The scavenging effect of PGRP-SC1B is readily explained because it cleaves the same motif in the peptidoglycan molecule that is being used by the receptor-PGRPs. It must be stressed, however, that this is an indicative observation in vitro and a possible PGRP scavenger function still remains to be shown in vivo. The likelihood of PGRP-SC1B being a scavenger can be further discussed as the expression pattern of PGRP-SC1B suggests that the main expression is restricted to the gut 40 region. However, there is also a weak expression from the fatbody (Werner et al., 2000). Possibly, SC1B is exported to the gut and is taking part in bacterial degradation as a digestive enzyme. This would be in analogy with the six Drosophila lysozymes that are expressed in the gut (Daffre et al., 1994) and might play a role in the degradation of the bacteria present in the decomposing fruits on which larvae feed. A bactericidal PGRP amidase (Paper II) As discussed above, PGRP-SC1B did not show any antibacterial activity and therefore it was surprising to find that PGRP-SB1 is highly bactericidal against B. megaterium. However, this is the only bacterial species PGRP-SB1 showed activity against, despite an exhaustive search for antibacterial activity against other bacteria including several Bacillus species. At present we do not know the feature(s) of the B. megaterium cell wall that makes the peptidoglycan accessible for PGRP-SB1 degradation. The removal of teichoic acids enhances the activity of PGRP-SC1B in vitro (Paper I). A possible explanation to the PGRP-SB1 bactericidal activity could be that the distribution or composition of teichoic acids being different in B. megaterium compared to other bacilli. LPS – to be, or not to be, an elicitor of the Drosophila immune system (Paper III and IV) Bacterial lipopolysaccharide, LPS, has for long been known to induce inflammation in humans. It was the first bacterial pattern molecule to be linked to a specific receptor (TLR4) of the human innate immune system (Poltorak et al., 1998; Rock et al., 1998). Also in studies of the insect immune system, LPS has been extensively used to elicit immune gene expression. An LPS receptor was therefore expected to be found in control of the Drosophila immunity pathways. The reports that describe PGRP-LC as the Imd-pathway receptor clearly show that the knock-out flies have a hampered response to LPS and live Gram-negative bacteria in addition to peptidoglycan (Choe et al., 2002; Gottar et al., 2002; Ramet et al., 2002b). However, PGRP proteins had been shown to bind peptidoglycan and not LPS. Even if peptidoglycan is a constituent of the Gram-negative bacterial cell-wall, it is hidden underneath the LPScontaining outer membrane. It is not easy to envisage how a receptor would be able to penetrate the lipid-bilayer to get access to the peptidoglycan. The observation that the LPS response is blocked in the PGRP-LC mutant fly indicates a broader PGRP-LC ligand specificity. However, an alternative explanation could be the presence of peptidoglycan impurities in the LPS preparation. The purity issue has been central in the study of mammalian Toll41 like receptors and their ligands. TLR2, for example, was initially believed to be an LPS receptor until extensive re-purification of LPS abolished the TLR2 mediated response (Hirschfeld et al., 2000). However, our discovery of a peptidoglycan scavenger, PGRP-SC1B, as described in Paper I, presented a way to decontaminate LPS from peptidoglycan impurities. In the two studies presented in Papers III and IV, PGRP-SC1B-treated LPS has been used to challenge Drosophila cells. In both studies this LPS is eliciting an immune response, in Paper IV at a concentration as low as 10 ng/ ml. The conclusion from Paper III is therefore that LPS really is an elicitor of the Imd-pathway. This conclusion, however, was drawn under the assumption that the peptidoglycan fragments produced by PGRP-SC1B lack activity. The study in Paper IV takes the impurity issue a step further and analyses the LPS preparation after it has been fractionated by gelfiltration. Indeed, the material that induces Drosophila cells from the LPS-solution that had been treated with a peptidoglycan digesting enzyme has a lower molecular weight than un-treated LPS. This is not the case for the LPS-responsive HEK293/TLR4 cells, which respond to material of the same molecular weight in the treated and un-treated LPS. Paper IV clearly shows that Drosophila S2 cells, having a functional Imd-pathway, are unresponsive to LPS. The first study suggesting that LPS is not an inducer of the Imdpathway was from Bruno Lemaitre’s laboratory (Leulier et al., 2003). They showed instead that the Toll and the Imd-pathway discriminately can be activated by peptidoglycan of Lys-type and DAP-type, respectively. An LPSinjection to the flies gave the same response as an injection of sterile water. However, the response from the sterile control injection was high and could possibly have masked a weak LPS response. PGRP-LC – three of a kind (Paper III and IV) The PGRP-LC gene-cluster encodes membrane-bound proteins generated by alternative splicing. A common intracellular non-PGRP domain and a transmembrane domain are spliced together with any of three different PGRP domains LCa, LCx, or LCy. Two additional PGRP domains was thought to be part of the PGRP-LC gene-cluster but as shown in paper III these are transcribed as a single transcript including both domains and therefore defines a unique gene, PGRP-LF. Gene knock-out experiments have placed PGRP-LC as a crucial component of the Imd-pathway up-stream of the Imd-protein (Choe et al., 2002; Gottar et al., 2002; Ramet et al., 2002b). In one of these studies a chromosomal mutation in the non-PGRP domain, thus affecting all three spliceforms, totally blocks the Imd-pathway (Gottar et al., 2002). A similar phenotype was reported in one of the other studies characterizing a chromosomal mutation affecting the PGRP-LCx domain (Choe et al., 2002). 42 In studies presented in Paper III and IV a similar approach was employed to determine the relative importance of the three different splice forms. In a cell culture system, RNAi was directed toward the different spliceforms to knock-down their expression, one at a time. Different immune elicitors were then added and the effect on immune gene induction was assayed. The results show that regardless of immune elicitor, RNAi directed against the PGRP-LCx domain or against the common non-PGRP domain totally block the Imd-pathway. This supports the notion that PGRP-LCx is a necessary spliceform and a major receptor of the Imd-pathway. RNAi directed against PGRPLCy, on the other hand, has no effect at all on Imd-pathway activation by the stimuli used. The effect of RNAi directed against the PGRP-LCa spliceform is intricate. In Paper III we report that LPS treated with PGRP-SC1B (but not untreated LPS) or living Gram-negative bacterial cells is dependent on PGRPLCa. The results in Paper IV directly show that LCa is required for induction with monomeric peptidoglycan. Also shown is that the PGRP-SC1B treatment does not fully remove the stimulants from LPS but reduces the immunostimulatory capacity 100 times. Is it possible that only Lys-type peptidoglycan can be fully inactivated by PGRP-SC1B? A more recent paper from our laboratory (Gelius et al., 2003) shows that even if PGRP-SC1B is highly active against E. coli peptidoglycan it can not cleave TCT. Thus, PGRP-SC1B can degrade most of the E. coli peptidoglycan but leave monomeric TCT-like molecules. As shown by Stenbak and colleagues (Stenbak et al., 2004), a peptidoglycan monomer with a muramic acid in anhydo conformation in combination with a DAP residue is required for full Imd-pathway activation. It is thus possible that the immune eliciting factors giving an LCa-dependent response in Paper III in fact are TCT-like molecules. Taking this into account provides an explanation why similar experimental results were interpreted differently in Paper III and IV. Polymeric peptidoglycan recognition (Paper III, IV, and V) It has for long been realized that the Toll pathway mainly responds towards fungi and certain Gram-positive bacteria such as M. luteus and that the Imd pathway is activated by Gram-negative bacteria and some Gram-positives such as Bacillus species. It was recently suggested that the receptors of the two pathways could discriminate between peptidoglycans having an L-lysine or a meso-DAP residue in the branching position of the tetrapeptide (Leulier et al., 2002). As no biochemical data were available, we decided to examine the relative binding affinities of the PGRP receptors and to correlate these with the known elicitor specificities of the two pathways. As shown in Paper V PGRPSA, the soluble receptor of the Toll-pathway indeed has affinity for peptidoglycan of Lys-type and not for peptidoglycan from B. megaterium, being 43 of DAP-type. Somewhat surprising however, we found that PGRP-SA has good affinity to DAP-type peptidoglycans from other species (E. coli and L. plantarum). In future experiments, it would be interesting to examine how well these peptidoglycans activate the Toll pathway. The peptidoglycan binding data for PGRP-LCx, the receptor of the Imd-pathway, are more intriguing. PGRP-LCx shows a remarkable high affinity for all types of peptidoglycan, which stands in contrast to the DAP-type peptidoglycan preference that has been reported for the Imd-pathway activation. A possible explanation of the strong activation seen with DAP-type peptidoglycan could be the anhydro-MurNAc present at the reducing ends of the glycan strands in Gram-negative bacterial peptidoglycan. The anhydo bond not only confers a different appearance to the MurNAc residue but also fixes the amino-hexose in a locked conformation in contrast to the open conformation of unreduced MurNAc that is oscillating between α- and β-conformation. It is thus possible that even if PGRP-LCx can bind to any polymeric peptidoglycan, the presence of an anhydro-MurNAc is required for a strong induction of the Imdpathway. This idea is supported by in a recent study (Stenbak et al., 2004). They found that the anhydro-MurNAc in combination with a DAP residue indeed is required for a full Imd response. Another recent study also supports this idea that the conformation of the MurNAc is instrumental for Toll and Imd activation (Filipe et al., 2005). These investigators show that the Toll pathway, in contrast to the Imd pathway, is positively activated by peptidoglycan fragments in which the MurNAc has an unreduced flexible conformation. In the model, presented in Paper V, we postulate that an Imdactivating PGRP-LCx homodimer will form when one of the dimer partners binds to a peptidoglycan motif that includes an 1,6-anhydro MurNAc residue and the other binds to an adjacent common peptidoglycan motif. Monomeric peptidoglycan recognition (Paper III, IV, and V) The analysis of the relative requirements of the different PGRP-LC spliceforms for activation of the Imd-pathway, presented in Paper III and IV, suggests that both the PGRP-LCa and LCx form are needed when the Imd-pathway is activated by monomeric peptidoglycan, or by impure LPS. These results suggest that the activating unit is a heterodimer complex formed by PGRP-LCa and LCx. In our biochemical analysis of the recombinant PGRP-LC domains, presented in Paper V, we found unexpectedly that PGRP-LCa does not bind to monomeric peptidoglycan and that instead PGRP-LCx has affinity for this elicitor. As activation data could be explained by receptor dimerization, an assay was set up that would enable testing if the proteins possibly get affinity for each other in the presence of the proper ligand. In this assay one of the recombinant proteins contains a histidine-tag and the other protein interaction partner lacks such tag. The two protein forms are incubated in the presence or absence of a 44 ligand and the incubation mixture is passed through a solid-phase histidinebinding matrix. If the proteins have affinity for each other then they are both trapped in the matrix, otherwise only the tagged form stays bound. In a second step, the proteins that are bound to the matrix are eluted. Using this method, we found that PGRP-LCa and -LCx indeed have affinity for each other in the presence of monomeric peptidoglycan (TCT). The interaction is specific for these PGRP forms since dimerization is not induced by any other combination of PGRP-LCa, LCx and SA. Our model for Imd activation is mainly based on the results from paper III, IV, and V. It is proposed that the Imd-pathway can be activated by PGRP-LCx binding to monomeric peptidoglycan, which causes a minor conformational change that attracts PGRP-LCa. The two proteins will thus constitute an activating dimer-complex with LCa serving as an adaptor or transducer and not as a receptor. Again, I want to stress that this model of Imdpathway activation is based on experiments in vitro using cell lines and recombinant proteins. Note: during the preparation of this thesis an article was published describing the crystal structure of PGRP-LCa (Chang et al., 2005). The structure shows that LCa has a distorted binding groove. Binding data presented are in full consonance with, but do not add anything to, the findings we had already published in paper V. Concluding remarks The work in this thesis has been focused on the recognition of bacteria by innate immune receptors which initiate the triggering of the pathways controlling immune gene expression. Most of the experiments have been performed in vitro using biochemical protein interaction approaches and molecular biology techniques to derive and manipulate cell lines. The main goal has been to elucidate the biological role of members of the PGRP gene family. Many questions have been answered and many new questions have been formulated. It is still an open question how Gram-negative bacteria are being recognized as we showed that LPS is not an inducer of the Imd-pathway. If the peptidoglycan sacculus is being recognized then PGRP proteins must in some way penetrate the LPS-containing outer membrane. Alternatively, peptidoglycan monomers which are lost to the environment by growing Gram-negative bacteria could possible offer an explanation to how these bacteria are being recognized. The existence of the dedicated splice variant PGRP-LCa indicates that recognition of peptidoglycan subunits is instrumental for bacterial recognition. We have demonstrated that the heterodimerization of PGRP-LCx and -LCa can be induced by monomeric peptidoglycan and it is likely that homodimerisation of PGRP-LCx receptors will be a consequence of polymeric peptidoglycan binding. However, these dimerization events remain to be shown also in vivo. 45 The elucidation of the physiological roles played by the peptidoglycan degrading enzymes with N-acetylmuramoyl L-alanine amidase activity will have to await the establishment of knock-out mutants. One can, however, foresee that they play important roles by modulating the property of the major stimulant of the Drosophila immune system, namely peptidoglycan. 46 Acknowledgements Hmm, have you already been reading this far? Noo, I hardly believe you… Maybe you didn’t know that those who skip pages in this book will be reborn as a laboratory fly in their next lives? First, I want to express my gratitude for my supervisor professor Håkan Steiner who brought my attention to this interesting field of pattern recognition. I appreciate the warm and open atmosphere in your lab, the free hands you gave me, and the immense support I had whenever I needed it. Your generosity in sharing your knowledge (and humor) has been invaluable. -I will never forget the mantra ‘PGRP - besser als Toll’… Next, I want to thank the other inhabitants of our research group from past and present; Gang for introducing me to the lab during my exam-work time, Annika for ‘lending’ me numerous acrylamide gels, Eva for fruitful discussions concerning amidases and for a never-ending flood of Swedish/ German jokes, Jenny, the best collaborator ever, who will never learn to say ‘kex’, Carina for your unpredictable comments on all aspects of life - I will be reminded of you whenever I’ll eat any Grapefruit, Sandra for your warm attitude and for dragging me around in the forest every week, and MacGyvor for fixing the machines and tools that I occasionally broke. The other seniors at the microbiology department, Leif, for introducing me to “who is who in the bacterial community”, Monica, for recurrent updates on male javelin throwers… Margareta for being supportive in teaching issues, IngaBritt, IB, for chats and laughs in the kitchen, cross-word solving, massage, and hjortron-berries, Helena, Anita, Olga, Eva, and Annette, for lunch-time chats and for keeping the institution going, Björn for preventing my computer from crashing, Inger Holmgren for extensive support at the teaching lab and for all the cakes and cookies. My comrades at the translation lab: Ernesto, Victor, Qing (how could you possibly write your thesis that fast? I’m impressed), Sergey, Richard and Mirja for many interesting discussions Georgina and Lily, an advice for your own best, -keep yourselves updated on male javelin throwers (especially those from the Czech Republic)… The people that have left the department during the years: Peter H for one or two games of chess, badminton and billiard, Oliver take care of Jenny and Leon in Copenhagen, Martin for your anti-optimism, Magnus, Magda and Josefin our parties have been missing you, Ann for all the rules and for all the cakes, Robert S, for exiting sub-cloning discussions. The staff at the rest of our department, the Geneticists and Toxicologists, especially the athletes attending the Innebandy fights. 47 The other innate immunity researchers at the university from Ylva’s, Uli’s, Ingrid’s groups, you are truly a resource of knowledge! Reseach collaborators: Dan Hultmark and your group in Umeå for nice collaboration, it is always inspiring to discuss with you. Niklas Schultz for sharing your knowledge and for running the confocal, Janet Håkansson for being an outstanding exam-worker, Neal Silverman, Takashi Kaneko, and William Goldman for collaboration, TCT, and research discussions. People outside the university… Thomas and Richard for bowling and for crazy parties in the ‘good old days’, Mathias and family for billiard games and dinners, Eddy, Helen, Emma, Viktor, and Lovisa for numerous dinners and for helping me reinvent the relativity-theory during late night-hours, Niklas alias Gegge for chips and electricity know-how. My sister Gunlis, and family, Olle, Fred and Annika for extensive hospitality and support during one of the toughest periods of my life. My brother Roger, and family, Anna-Karin, Emmie and Lise for care, and for introducing me to ornithology, which was the main reason I started this education in the first place (-how did I end up with this fly research? I still cannot give a proper answer…). My parents Margit and Harry, thanks for your support, it is always so nice to come to your place and to be taken care of… Finally, my family, -Therese, for love, support, Grådask, and for making this work possible -Fabian and Johannes, you will definitely be the next Zlatan, but times two! -min dotter Stina, du är mitt käraste skogssmultron ♥ Tack för att du låter UllaBerta och Doris låna mig en arm att sova på ibland! 6 Bryt upp, bryt upp! Den nya dagen gryr. Oändligt är vårt stora äventyr. 48 References Agaisse, H., and Perrimon, N. (2004). The roles of JAK/STAT signaling in Drosophila immune responses. Immunol Rev 198, 72-82. Agaisse, H., Petersen, U. M., Boutros, M., Mathey-Prevot, B., and Perrimon, N. (2003). Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev Cell 5, 441-450. Alexopoulou, L., Holt, A. C., Medzhitov, R., and Flavell, R. A. (2001). Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 413, 732-738. Atrih, A., and Foster, S. J. (2001). In vivo roles of the germination-specific lytic enzymes of Bacillus subtilis 168. Microbiology 147, 2925-2932. Belvin, M. P., and Anderson, K. V. (1996). A conserved signaling pathway: the Drosophila toll-dorsal pathway. Annu Rev Cell Dev Biol 12, 393-416. Bera, A., Herbert, S., Jakob, A., Vollmer, W., and Gotz, F. (2005). Why are pathogenic staphylococci so lysozyme resistant? The peptidoglycan O-acetyltransferase OatA is the major determinant for lysozyme resistance of Staphylococcus aureus. Mol Microbiol 55, 778-787. Bidla, G., Lindgren, M., Theopold, U., and Dushay, M. S. (2005). Hemolymph coagulation and phenoloxidase in Drosophila larvae. Dev Comp Immunol 29, 669-679. Bischoff, V., Vignal, C., Boneca, I. G., Michel, T., Hoffmann, J. A., and Royet, J. (2004). Function of the drosophila pattern-recognition receptor PGRP-SD in the detection of Gram-positive bacteria. Nat Immunol 5, 1175-1180. Boman, H. G., and Steiner, H. (1981). Humoral immunity in Cecropia pupae. Curr Top Microbiol Immunol 94-95, 75-91. Boneca, I. G. (2005). The role of peptidoglycan in pathogenesis. Curr Opin Microbiol 8, 4653. Boutros, M., Agaisse, H., and Perrimon, N. (2002). Sequential activation of signaling pathways during innate immune responses in Drosophila. Dev Cell 3, 711-722. Brown, G. D., and Gordon, S. (2003). Fungal beta-glucans and mammalian immunity. Immunity 19, 311-315. Brown, G. D., Herre, J., Williams, D. L., Willment, J. A., Marshall, A. S., and Gordon, S. (2003). Dectin-1 mediates the biological effects of beta-glucans. J Exp Med 197, 11191124. Bulet, P., Dimarcq, J. L., Hetru, C., Lagueux, M., Charlet, M., Hegy, G., Van Dorsselaer, A., and Hoffmann, J. A. (1993). A novel inducible antibacterial peptide of Drosophila carries an O-glycosylated substitution. J Biol Chem 268, 14893-14897. Carlsson, A., Engstrom, P., Palva, E. T., and Bennich, H. (1991). Attacin, an antibacterial protein from Hyalophora cecropia, inhibits synthesis of outer membrane proteins in Escherichia coli by interfering with omp gene transcription. Infect Immun 59, 3040-3045. Cerenius, L., and Soderhall, K. (2004). The prophenoloxidase-activating system in invertebrates. Immunol Rev 198, 116-126. Chang, C. I., Ihara, K., Chelliah, Y., Mengin-Lecreulx, D., Wakatsuki, S., and Deisenhofer, J. (2005). Structure of the ectodomain of Drosophila peptidoglycan-recognition protein LCa suggests a molecular mechanism for pattern recognition. Proc Natl Acad Sci U S A 102, 10279-10284. 49 Chang, C. I., Pili-Floury, S., Herve, M., Parquet, C., Chelliah, Y., Lemaitre, B., MenginLecreulx, D., and Deisenhofer, J. (2004). A Drosophila pattern recognition receptor contains a peptidoglycan docking groove and unusual L,D-carboxypeptidase activity. PLoS Biol 2, E277. Cheng, Q., Li, H., Merdek, K., and Park, J. T. (2000). Molecular characterization of the betaN-acetylglucosaminidase of Escherichia coli and its role in cell wall recycling. J Bacteriol 182, 4836-4840. Cheng, Q., and Park, J. T. (2002). Substrate specificity of the AmpG permease required for recycling of cell wall anhydro-muropeptides. J Bacteriol 184, 6434-6436. Cheng, X., Zhang, X., Pflugrath, J. W., and Studier, F. W. (1994). The structure of bacteriophage T7 lysozyme, a zinc amidase and an inhibitor of T7 RNA polymerase. Proc Natl Acad Sci U S A 91, 4034-4038. Cho, J. H., Fraser, I. P., Fukase, K., Kusumoto, S., Fujimoto, Y., Stahl, G. L., and Ezekowitz, R. A. (2005). Human peptidoglycan recognition protein-S is an effector of neutrophilmediated innate immunity. Blood. Choe, K. M., Lee, H., and Anderson, K. V. (2005). Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc Natl Acad Sci U S A 102, 1122-1126. Choe, K. M., Werner, T., Stoven, S., Hultmark, D., and Anderson, K. V. (2002). Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 296, 359-362. Christophides, G. K., Zdobnov, E., Barillas-Mury, C., Birney, E., Blandin, S., Blass, C., Brey, P. T., Collins, F. H., Danielli, A., Dimopoulos, G., et al. (2002). Immunity-related genes and gene families in Anopheles gambiae. Science 298, 159-165. Cundell, D. R., Kanthakumar, K., Taylor, G. W., Goldman, W. E., Flak, T., Cole, P. J., and Wilson, R. (1994). Effect of tracheal cytotoxin from Bordetella pertussis on human neutrophil function in vitro. Infect Immun 62, 639-643. da Silva Correia, J., Soldau, K., Christen, U., Tobias, P. S., and Ulevitch, R. J. (2001). Lipopolysaccharide is in close proximity to each of the proteins in its membrane receptor complex. transfer from CD14 to TLR4 and MD-2. J Biol Chem 276, 21129-21135. Daffre, S., Kylsten, P., Samakovlis, C., and Hultmark, D. (1994). The lysozyme locus in Drosophila melanogaster: an expanded gene family adapted for expression in the digestive tract. Mol Gen Genet 242, 152-162. De Gregorio, E., Han, S. J., Lee, W. J., Baek, M. J., Osaki, T., Kawabata, S., Lee, B. L., Iwanaga, S., Lemaitre, B., and Brey, P. T. (2002a). An immune-responsive Serpin regulates the melanization cascade in Drosophila. Dev Cell 3, 581-592. De Gregorio, E., Spellman, P. T., Tzou, P., Rubin, G. M., and Lemaitre, B. (2002b). The Toll and Imd pathways are the major regulators of the immune response in Drosophila. Embo J 21, 2568-2579. Dimarcq, J. L., Hoffmann, D., Meister, M., Bulet, P., Lanot, R., Reichhart, J. M., and Hoffmann, J. A. (1994). Characterization and transcriptional profiles of a Drosophila gene encoding an insect defensin. A study in insect immunity. Eur J Biochem 221, 201209. Dziarski, R. (2004). Peptidoglycan recognition proteins (PGRPs). Mol Immunol 40, 877-886. Ekengren, S., and Hultmark, D. (2001). A family of Turandot-related genes in the humoral stress response of Drosophila. Biochem Biophys Res Commun 284, 998-1003. Engstrom, Y., Kadalayil, L., Sun, S. C., Samakovlis, C., Hultmark, D., and Faye, I. (1993). kappa B-like motifs regulate the induction of immune genes in Drosophila. J Mol Biol 232, 327-333. 50 Fehlbaum, P., Bulet, P., Michaut, L., Lagueux, M., Broekaert, W. F., Hetru, C., and Hoffmann, J. A. (1994). Insect immunity. Septic injury of Drosophila induces the synthesis of a potent antifungal peptide with sequence homology to plant antifungal peptides. J Biol Chem 269, 33159-33163. Filipe, S. R., Tomasz, A., and Ligoxygakis, P. (2005). Requirements of peptidoglycan structure that allow detection by the Drosophila Toll pathway. EMBO Rep 6, 327-333. Folkesson, A., Eriksson, S., Andersson, M., Park, J. T., and Normark, S. (2005). Components of the peptidoglycan-recycling pathway modulate invasion and intracellular survival of Salmonella enterica serovar Typhimurium. Cell Microbiol 7, 147-155. Franc, N. C., Dimarcq, J. L., Lagueux, M., Hoffmann, J., and Ezekowitz, R. A. (1996). Croquemort, a novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity 4, 431-443. Gantner, B. N., Simmons, R. M., Canavera, S. J., Akira, S., and Underhill, D. M. (2003). Collaborative induction of inflammatory responses by dectin-1 and Toll-like receptor 2. J Exp Med 197, 1107-1117. Gelius, E., Persson, C., Karlsson, J., and Steiner, H. (2003). A mammalian peptidoglycan recognition protein with N-acetylmuramoyl-L-alanine amidase activity. Biochem Biophys Res Commun 306, 988-994. Goberdhan, D. C., and Wilson, C. (1998). JNK, cytoskeletal regulator and stress response kinase? A Drosophila perspective. Bioessays 20, 1009-1019. Gobert, V., Gottar, M., Matskevich, A. A., Rutschmann, S., Royet, J., Belvin, M., Hoffmann, J. A., and Ferrandon, D. (2003). Dual activation of the Drosophila toll pathway by two pattern recognition receptors. Science 302, 2126-2130. Goodell, E. W., and Schwarz, U. (1985). Release of cell wall peptides into culture medium by exponentially growing Escherichia coli. J Bacteriol 162, 391-397. Gottar, M., Gobert, V., Michel, T., Belvin, M., Duyk, G., Hoffmann, J. A., Ferrandon, D., and Royet, J. (2002). The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 416, 640-644. Greenway, D. L., and Perkins, H. R. (1985). Turnover of the cell wall peptidoglycan during growth of Neisseria gonorrhoeae and Escherichia coli. Relative stability of newly synthesized material. J Gen Microbiol 131, 253-263. Guan, R., Malchiodi, E. L., Wang, Q., Schuck, P., and Mariuzza, R. A. (2004a). Crystal structure of the C-terminal peptidoglycan-binding domain of human peptidoglycan recognition protein Ialpha. J Biol Chem 279, 31873-31882. Guan, R., Roychowdhury, A., Ember, B., Kumar, S., Boons, G. J., and Mariuzza, R. A. (2004b). Structural basis for peptidoglycan binding by peptidoglycan recognition proteins. Proc Natl Acad Sci U S A 101, 17168-17173. Guan, R., Roychowdury, A., Ember, B., Kumar, S., Boons, G. J., and Mariuzza, R. A. (2005a). Crystal structure of a peptidoglycan recognition protein (PGRP) in complex with a muramyl tripeptide from Gram-positive bacteria. J Endotoxin Res 11, 41-46. Guan, R., Wang, Q., Sundberg, E. J., and Mariuzza, R. A. (2005b). Crystal structure of human peptidoglycan recognition protein S (PGRP-S) at 1.70 A resolution. J Mol Biol 347, 683691. Guha, M., and Mackman, N. (2001). LPS induction of gene expression in human monocytes. Cell Signal 13, 85-94. Harz, H., Burgdorf, K., and Holtje, J. V. (1990). Isolation and separation of the glycan strands from murein of Escherichia coli by reversed-phase high-performance liquid chromatography. Anal Biochem 190, 120-128. 51 Heil, F., Hemmi, H., Hochrein, H., Ampenberger, F., Kirschning, C., Akira, S., Lipford, G., Wagner, H., and Bauer, S. (2004). Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 303, 1526-1529. Hemmi, H., Takeuchi, O., Kawai, T., Kaisho, T., Sato, S., Sanjo, H., Matsumoto, M., Hoshino, K., Wagner, H., Takeda, K., and Akira, S. (2000). A Toll-like receptor recognizes bacterial DNA. Nature 408, 740-745. Hirschfeld, M., Kirschning, C. J., Schwandner, R., Wesche, H., Weis, J. H., Wooten, R. M., and Weis, J. J. (1999). Cutting edge: inflammatory signaling by Borrelia burgdorferi lipoproteins is mediated by toll-like receptor 2. J Immunol 163, 2382-2386. Hirschfeld, M., Ma, Y., Weis, J. H., Vogel, S. N., and Weis, J. J. (2000). Cutting edge: repurification of lipopolysaccharide eliminates signaling through both human and murine toll-like receptor 2. J Immunol 165, 618-622. Hoijer, M. A., de Groot, R., van Lieshout, L., Jacobs, B. C., Melief, M. J., and Hazenberg, M. P. (1998). Differences in N-acetylmuramyl-L-alanine amidase and lysozyme in serum and cerebrospinal fluid of patients with bacterial meningitis. J Infect Dis 177, 102-106. Hoijer, M. A., Melief, M. J., Calafat, J., Roos, D., van den Beemd, R. W., van Dongen, J. J., and Hazenberg, M. P. (1997). Expression and intracellular localization of the human Nacetylmuramyl-L-alanine amidase, a bacterial cell wall-degrading enzyme. Blood 90, 1246-1254. Hoijer, M. A., Melief, M. J., Keck, W., and Hazenberg, M. P. (1996). Purification and characterization of N-acetylmuramyl-L-alanine amidase from human plasma using monoclonal antibodies. Biochim Biophys Acta 1289, 57-64. Holtje, J. V. (1996a). A hypothetical holoenzyme involved in the replication of the murein sacculus of Escherichia coli. Microbiology 142 ( Pt 8), 1911-1918. Holtje, J. V. (1996b). Molecular interplay of murein synthases and murein hydrolases in Escherichia coli. Microb Drug Resist 2, 99-103. Hultmark, D. (2003). Drosophila immunity: paths and patterns. Curr Opin Immunol 15, 1219. Imler, J. L., and Bulet, P. (2005). Antimicrobial peptides in Drosophila: structures,activities and gene regulation. Chem Immunol Allergy 86, 1-21. Ip, Y. T., Kraut, R., Levine, M., and Rushlow, C. A. (1991). The dorsal morphogen is a sequence-specific DNA-binding protein that interacts with a long-range repression element in Drosophila. Cell 64, 439-446. Ip, Y. T., Reach, M., Engstrom, Y., Kadalayil, L., Cai, H., Gonzalez-Crespo, S., Tatei, K., and Levine, M. (1993). Dif, a dorsal-related gene that mediates an immune response in Drosophila. Cell 75, 753-763. Irving, P., Ubeda, J. M., Doucet, D., Troxler, L., Lagueux, M., Zachary, D., Hoffmann, J. A., Hetru, C., and Meister, M. (2005). New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol 7, 335-350. Jacobs, C., Joris, B., Jamin, M., Klarsov, K., Van Beeumen, J., Mengin-Lecreulx, D., van Heijenoort, J., Park, J. T., Normark, S., and Frere, J. M. (1995). AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramylL-alanine amidase. Mol Microbiol 15, 553-559. Janeway, C. A., Jr. (1989). Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol 54 Pt 1, 1-13. Johannsen, L., Toth, L. A., Rosenthal, R. S., Opp, M. R., Obal, F., Jr., Cady, A. B., and Krueger, J. M. (1990). Somnogenic, pyrogenic, and hematologic effects of bacterial peptidoglycan. Am J Physiol 258, R182-186. Johansson, K. C., Metzendorf, C., and Soderhall, K. (2005). Microarray analysis of immune challenged Drosophila hemocytes. Exp Cell Res 305, 145-155. 52 Kang, D., Liu, G., Gunne, H., and Steiner, H. (1996). PCR differential display of immune gene expression in Trichoplusia ni. Insect Biochem Mol Biol 26, 177-184. Kang, D., Liu, G., Lundstrom, A., Gelius, E., and Steiner, H. (1998). A peptidoglycan recognition protein in innate immunity conserved from insects to humans. Proc Natl Acad Sci U S A 95, 10078-10082. Kappeler, S. R., Heuberger, C., Farah, Z., and Puhan, Z. (2004). Expression of the peptidoglycan recognition protein, PGRP, in the lactating mammary gland. J Dairy Sci 87, 2660-2668. Khush, R. S., and Lemaitre, B. (2000). Genes that fight infection: what the Drosophila genome says about animal immunity. Trends Genet 16, 442-449. Kim, M. S., Byun, M., and Oh, B. H. (2003). Crystal structure of peptidoglycan recognition protein LB from Drosophila melanogaster. Nat Immunol 4, 787-793. Kim, Y. S., Ryu, J. H., Han, S. J., Choi, K. H., Nam, K. B., Jang, I. H., Lemaitre, B., Brey, P. T., and Lee, W. J. (2000). Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J Biol Chem 275, 32721-32727. Kimbrell, D. A., and Beutler, B. (2001). The evolution and genetics of innate immunity. Nat Rev Genet 2, 256-267. Kiselev, S. L., Kustikova, O. S., Korobko, E. V., Prokhortchouk, E. B., Kabishev, A. A., Lukanidin, E. M., and Georgiev, G. P. (1998). Molecular cloning and characterization of the mouse tag7 gene encoding a novel cytokine. J Biol Chem 273, 18633-18639. Koch, A. L. (1995). Bacterial growth and form, Chapman & Hall, New York, N.Y). Lagueux, M., Perrodou, E., Levashina, E. A., Capovilla, M., and Hoffmann, J. A. (2000). Constitutive expression of a complement-like protein in toll and JAK gain-of-function mutants of Drosophila. Proc Natl Acad Sci U S A 97, 11427-11432. Lanot, R., Zachary, D., Holder, F., and Meister, M. (2001). Postembryonic hematopoiesis in Drosophila. Dev Biol 230, 243-257. Lee, M. H., Osaki, T., Lee, J. Y., Baek, M. J., Zhang, R., Park, J. W., Kawabata, S., Soderhall, K., and Lee, B. L. (2004). Peptidoglycan recognition proteins involved in 1,3-beta-Dglucan-dependent prophenoloxidase activation system of insect. J Biol Chem 279, 32183227. Lehane, M. J. (1997). Peritrophic matrix structure and function. Annu Rev Entomol 42, 525550. Lemaitre, B., Kromer-Metzger, E., Michaut, L., Nicolas, E., Meister, M., Georgel, P., Reichhart, J. M., and Hoffmann, J. A. (1995). A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense. Proc Natl Acad Sci U S A 92, 9465-9469. Lemaitre, B., Nicolas, E., Michaut, L., Reichhart, J. M., and Hoffmann, J. A. (1996). The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86, 973-983. Leulier, F., Parquet, C., Pili-Floury, S., Ryu, J. H., Caroff, M., Lee, W. J., Mengin-Lecreulx, D., and Lemaitre, B. (2003). The Drosophila immune system detects bacteria through specific peptidoglycan recognition. Nat Immunol 4, 478-484. Leulier, F., Vidal, S., Saigo, K., Ueda, R., and Lemaitre, B. (2002). Inducible expression of double-stranded RNA reveals a role for dFADD in the regulation of the antibacterial response in Drosophila adults. Curr Biol 12, 996-1000. Levashina, E. A., Langley, E., Green, C., Gubb, D., Ashburner, M., Hoffmann, J. A., and Reichhart, J. M. (1999). Constitutive activation of toll-mediated antifungal defense in serpin-deficient Drosophila. Science 285, 1917-1919. 53 Levashina, E. A., Moita, L. F., Blandin, S., Vriend, G., Lagueux, M., and Kafatos, F. C. (2001). Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae. Cell 104, 709-718. Levashina, E. A., Ohresser, S., Bulet, P., Reichhart, J. M., Hetru, C., and Hoffmann, J. A. (1995). Metchnikowin, a novel immune-inducible proline-rich peptide from Drosophila with antibacterial and antifungal properties. Eur J Biochem 233, 694-700. Ligoxygakis, P., Pelte, N., Hoffmann, J. A., and Reichhart, J. M. (2002a). Activation of Drosophila Toll during fungal infection by a blood serine protease. Science 297, 114-116. Ligoxygakis, P., Pelte, N., Ji, C., Leclerc, V., Duvic, B., Belvin, M., Jiang, H., Hoffmann, J. A., and Reichhart, J. M. (2002b). A serpin mutant links Toll activation to melanization in the host defence of Drosophila. Embo J 21, 6330-6337. Liu, C., Gelius, E., Liu, G., Steiner, H., and Dziarski, R. (2000). Mammalian peptidoglycan recognition protein binds peptidoglycan with high affinity, is expressed in neutrophils, and inhibits bacterial growth. J Biol Chem 275, 24490-24499. Liu, C., Xu, Z., Gupta, D., and Dziarski, R. (2001). Peptidoglycan recognition proteins: a novel family of four human innate immunity pattern recognition molecules. J Biol Chem 276, 34686-34694. Lu, Y., Wu, L. P., and Anderson, K. V. (2001). The antibacterial arm of the drosophila innate immune response requires an IkappaB kinase. Genes Dev 15, 104-110. Luo, H., and Dearolf, C. R. (2001). The JAK/STAT pathway and Drosophila development. Bioessays 23, 1138-1147. Ma, C., and Kanost, M. R. (2000). A beta1,3-glucan recognition protein from an insect, Manduca sexta, agglutinates microorganisms and activates the phenoloxidase cascade. J Biol Chem 275, 7505-7514. Ma, Y. G., Cho, M. Y., Zhao, M., Park, J. W., Matsushita, M., Fujita, T., and Lee, B. L. (2004). Human mannose-binding lectin and L-ficolin function as specific pattern recognition proteins in the lectin activation pathway of complement. J Biol Chem 279, 25307-25312. Medzhitov, R., Preston-Hurlburt, P., and Janeway, C. A., Jr. (1997). A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 388, 394397. Meister, M., Hetru, C., and Hoffmann, J. A. (2000). The antimicrobial host defense of Drosophila. Curr Top Microbiol Immunol 248, 17-36. Melly, M. A., McGee, Z. A., and Rosenthal, R. S. (1984). Ability of monomeric peptidoglycan fragments from Neisseria gonorrhoeae to damage human fallopian-tube mucosa. J Infect Dis 149, 378-386. Meng, X., Khanuja, B. S., and Ip, Y. T. (1999). Toll receptor-mediated Drosophila immune response requires Dif, an NF-kappaB factor. Genes Dev 13, 792-797. Michel, T., Reichhart, J. M., Hoffmann, J. A., and Royet, J. (2001). Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature 414, 756-759. Mollner, S., and Braun, V. (1984). Murein hydrolase (N-acetyl-muramyl-L-alanine amidase) in human serum. Arch Microbiol 140, 171-177. Neuhaus, F. C., and Baddiley, J. (2003). A continuum of anionic charge: structures and functions of D-alanyl-teichoic acids in gram-positive bacteria. Microbiol Mol Biol Rev 67, 686-723. Noselli, S. (1998). JNK signaling and morphogenesis in Drosophila. Trends Genet 14, 33-38. Ochiai, M., and Ashida, M. (1988). Purification of a beta-1,3-glucan recognition protein in the prophenoloxidase activating system from hemolymph of the silkworm, Bombyx mori. J Biol Chem 263, 12056-12062. 54 Ochiai, M., and Ashida, M. (1999). A pattern recognition protein for peptidoglycan. Cloning the cDNA and the gene of the silkworm, Bombyx mori. J Biol Chem 274, 11854-11858. Onfelt Tingvall, T., Roos, E., and Engstrom, Y. (2001). The imd gene is required for local Cecropin expression in Drosophila barrier epithelia. EMBO Rep 2, 239-243. Park, J. T. (1995). Why does Escherichia coli recycle its cell wall peptides? Mol Microbiol 17, 421-426. Park, J. T. (2001). Identification of a dedicated recycling pathway for anhydro-Nacetylmuramic acid and N-acetylglucosamine derived from Escherichia coli cell wall murein. J Bacteriol 183, 3842-3847. Pasare, C., and Medzhitov, R. (2004). Toll-like receptors: linking innate and adaptive immunity. Microbes Infect 6, 1382-1387. Philpott, D. J., and Girardin, S. E. (2004). The role of Toll-like receptors and Nod proteins in bacterial infection. Mol Immunol 41, 1099-1108. Pili-Floury, S., Leulier, F., Takahashi, K., Saigo, K., Samain, E., Ueda, R., and Lemaitre, B. (2004). In vivo RNA interference analysis reveals an unexpected role for GNBP1 in the defense against Gram-positive bacterial infection in Drosophila adults. J Biol Chem 279, 12848-12853. Poltorak, A., He, X., Smirnova, I., Liu, M. Y., Van Huffel, C., Du, X., Birdwell, D., Alejos, E., Silva, M., Galanos, C., et al. (1998). Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282, 2085-2088. Ramet, M., Lanot, R., Zachary, D., and Manfruelli, P. (2002a). JNK signaling pathway is required for efficient wound healing in Drosophila. Dev Biol 241, 145-156. Ramet, M., Manfruelli, P., Pearson, A., Mathey-Prevot, B., and Ezekowitz, R. A. (2002b). Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature 416, 644-648. Rehman, A., Taishi, P., Fang, J., Majde, J. A., and Krueger, J. M. (2001). The cloning of a rat peptidoglycan recognition protein (PGRP) and its induction in brain by sleep deprivation. Cytokine 13, 8-17. Rock, F. L., Hardiman, G., Timans, J. C., Kastelein, R. A., and Bazan, J. F. (1998). A family of human receptors structurally related to Drosophila Toll. Proc Natl Acad Sci U S A 95, 588-593. Romeis, T., and Holtje, J. V. (1994a). Penicillin-binding protein 7/8 of Escherichia coli is a DD-endopeptidase. Eur J Biochem 224, 597-604. Romeis, T., and Holtje, J. V. (1994b). Specific interaction of penicillin-binding proteins 3 and 7/8 with soluble lytic transglycosylase in Escherichia coli. J Biol Chem 269, 2160321607. Rosetto, M., Engstrom, Y., Baldari, C. T., Telford, J. L., and Hultmark, D. (1995). Signals from the IL-1 receptor homolog, Toll, can activate an immune response in a Drosophila hemocyte cell line. Biochem Biophys Res Commun 209, 111-116. Royet, J., and Reichhart, J. M. (2003). Detection of peptidoglycans by NOD proteins. Trends Cell Biol 13, 610-614. Royet, J., Reichhart, J. M., and Hoffmann, J. A. (2005). Sensing and signaling during infection in Drosophila. Curr Opin Immunol 17, 11-17. Rutschmann, S., Jung, A. C., Zhou, R., Silverman, N., Hoffmann, J. A., and Ferrandon, D. (2000). Role of Drosophila IKK gamma in a toll-independent antibacterial immune response. Nat Immunol 1, 342-347. Ryu, J. H., Nam, K. B., Oh, C. T., Nam, H. J., Kim, S. H., Yoon, J. H., Seong, J. K., Yoo, M. A., Jang, I. H., Brey, P. T., and Lee, W. J. (2004). The homeobox gene Caudal regulates constitutive local expression of antimicrobial peptide genes in Drosophila epithelia. Mol Cell Biol 24, 172-185. 55 Samakovlis, C., Kimbrell, D. A., Kylsten, P., Engstrom, A., and Hultmark, D. (1990). The immune response in Drosophila: pattern of cecropin expression and biological activity. Embo J 9, 2969-2976. Sansonetti, P. J. (2004). War and peace at mucosal surfaces. Nat Rev Immunol 4, 953-964. Sashchenko, L. P., Dukhanina, E. A., Yashin, D. V., Shatalov, Y. V., Romanova, E. A., Korobko, E. V., Demin, A. V., Lukyanova, T. I., Kabanova, O. D., Khaidukov, S. V., et al. (2004). Peptidoglycan recognition protein tag7 forms a cytotoxic complex with heat shock protein 70 in solution and in lymphocytes. J Biol Chem 279, 2117-2124. Scherfer, C., Karlsson, C., Loseva, O., Bidla, G., Goto, A., Havemann, J., Dushay, M. S., and Theopold, U. (2004). Isolation and characterization of hemolymph clotting factors in Drosophila melanogaster by a pullout method. Curr Biol 14, 625-629. Schleifer, K. H., and Kandler, O. (1972). Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol Rev 36, 407-477. Schwandner, R., Dziarski, R., Wesche, H., Rothe, M., and Kirschning, C. J. (1999). Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by toll-like receptor 2. J Biol Chem 274, 17406-17409. Shibata, Y., Kawada, M., Nakano, Y., Toyoshima, K., and Yamashita, Y. (2005). Identification and characterization of an autolysin-encoding gene of Streptococcus mutans. Infect Immun 73, 3512-3520. Silverman, N., Zhou, R., Erlich, R. L., Hunter, M., Bernstein, E., Schneider, D., and Maniatis, T. (2003). Immune activation of NF-kappaB and JNK requires Drosophila TAK1. J Biol Chem 278, 48928-48934. Steiner, H. (1982). Secondary structure of the cecropins: antibacterial peptides from the moth Hyalophora cecropia. FEBS Lett 137, 283-287. Steiner, H., Hultmark, D., Engstrom, A., Bennich, H., and Boman, H. G. (1981). Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 292, 246-248. Stenbak, C. R., Ryu, J. H., Leulier, F., Pili-Floury, S., Parquet, C., Herve, M., Chaput, C., Boneca, I. G., Lee, W. J., Lemaitre, B., and Mengin-Lecreulx, D. (2004). Peptidoglycan molecular requirements allowing detection by the Drosophila immune deficiency pathway. J Immunol 173, 7339-7348. Stoven, S., Ando, I., Kadalayil, L., Engstrom, Y., and Hultmark, D. (2000). Activation of the Drosophila NF-kappaB factor Relish by rapid endoproteolytic cleavage. EMBO Rep 1, 347-352. Sun, H., Towb, P., Chiem, D. N., Foster, B. A., and Wasserman, S. A. (2004). Regulated assembly of the Toll signaling complex drives Drosophila dorsoventral patterning. Embo J 23, 100-110. Sun, S. C., and Faye, I. (1992). Cecropia immunoresponsive factor, an insect immunoresponsive factor with DNA-binding properties similar to nuclear-factor kappa B. Eur J Biochem 204, 885-892. Tahar, R., Boudin, C., Thiery, I., and Bourgouin, C. (2002). Immune response of Anopheles gambiae to the early sporogonic stages of the human malaria parasite Plasmodium falciparum. Embo J 21, 6673-6680. Takeda, K., and Akira, S. (2005). Toll-like receptors in innate immunity. Int Immunol 17, 114. Takeda, K., Takeuchi, O., and Akira, S. (2002). Recognition of lipopeptides by Toll-like receptors. J Endotoxin Res 8, 459-463. Takehana, A., Katsuyama, T., Yano, T., Oshima, Y., Takada, H., Aigaki, T., and Kurata, S. (2002). Overexpression of a pattern-recognition receptor, peptidoglycan-recognition 56 protein-LE, activates imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc Natl Acad Sci U S A 99, 13705-13710. Takehana, A., Yano, T., Mita, S., Kotani, A., Oshima, Y., and Kurata, S. (2004). Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. Embo J 23, 4690-4700. Templin, M. F., Ursinus, A., and Holtje, J. V. (1999). A defect in cell wall recycling triggers autolysis during the stationary growth phase of Escherichia coli. Embo J 18, 4108-4117. Thisse, C., Perrin-Schmitt, F., Stoetzel, C., and Thisse, B. (1991). Sequence-specific transactivation of the Drosophila twist gene by the dorsal gene product. Cell 65, 11911201. Turner, M. S., Hafner, L. M., Walsh, T., and Giffard, P. M. (2004). Identification, characterisation and specificity of a cell wall lytic enzyme from Lactobacillus fermentum BR11. FEMS Microbiol Lett 238, 9-15. Tydell, C. C., Yount, N., Tran, D., Yuan, J., and Selsted, M. E. (2002). Isolation, characterization, and antimicrobial properties of bovine oligosaccharide-binding protein. A microbicidal granule protein of eosinophils and neutrophils. J Biol Chem 277, 1965819664. Tzou, P., Ohresser, S., Ferrandon, D., Capovilla, M., Reichhart, J. M., Lemaitre, B., Hoffmann, J. A., and Imler, J. L. (2000). Tissue-specific inducible expression of antimicrobial peptide genes in Drosophila surface epithelia. Immunity 13, 737-748. Tzou, P., Reichhart, J. M., and Lemaitre, B. (2002). Constitutive expression of a single antimicrobial peptide can restore wild-type resistance to infection in immunodeficient Drosophila mutants. Proc Natl Acad Sci U S A 99, 2152-2157. Uehara, A., Sugawara, Y., Kurata, S., Fujimoto, Y., Fukase, K., Kusumoto, S., Satta, Y., Sasano, T., Sugawara, S., and Takada, H. (2005). Chemically synthesized pathogenassociated molecular patterns increase the expression of peptidoglycan recognition proteins via toll-like receptors, NOD1 and NOD2 in human oral epithelial cells. Cell Microbiol 7, 675-686. Uehara, T., and Park, J. T. (2004). The N-acetyl-D-glucosamine kinase of Escherichia coli and its role in murein recycling. J Bacteriol 186, 7273-7279. Valinger, Z., Ladesic, B., and Tomasic, J. (1982). Partial purification and characterization of N-acetylmuramyl-L-alanine amidase from human and mouse serum. Biochim Biophys Acta 701, 63-71. Vanderwinkel, E., de Pauw, P., Philipp, D., Ten Have, J. P., and Bainter, K. (1995). The human and mammalian N-acetylmuramyl-L-alanine amidase: distribution, action on different bacterial peptidoglycans, and comparison with the human lysozyme activities. Biochem Mol Med 54, 26-32. Wang, Z. M., Li, X., Cocklin, R. R., Wang, M., Fukase, K., Inamura, S., Kusumoto, S., Gupta, D., and Dziarski, R. (2003). Human peptidoglycan recognition protein-L is an Nacetylmuramoyl-L-alanine amidase. J Biol Chem 278, 49044-49052. Werner, T., Liu, G., Kang, D., Ekengren, S., Steiner, H., and Hultmark, D. (2000). A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc Natl Acad Sci U S A 97, 13772-13777. Wicker, C., Reichhart, J. M., Hoffmann, D., Hultmark, D., Samakovlis, C., and Hoffmann, J. A. (1990). Insect immunity. Characterization of a Drosophila cDNA encoding a novel member of the diptericin family of immune peptides. J Biol Chem 265, 22493-22498. Vollmer, W., and Holtje, J. V. (2004). The architecture of the murein (peptidoglycan) in gram-negative bacteria: vertical scaffold or horizontal layer(s)? J Bacteriol 186, 59785987. 57 Vollmer, W., and Tomasz, A. (2002). Peptidoglycan N-acetylglucosamine deacetylase, a putative virulence factor in Streptococcus pneumoniae. Infect Immun 70, 7176-7178. Votsch, W., and Templin, M. F. (2000). Characterization of a beta -N-acetylglucosaminidase of Escherichia coli and elucidation of its role in muropeptide recycling and beta lactamase induction. J Biol Chem 275, 39032-39038. Wu, L. P., and Anderson, K. V. (1998). Regulated nuclear import of Rel proteins in the Drosophila immune response. Nature 392, 93-97. Yoshida, H., Kinoshita, K., and Ashida, M. (1996). Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J Biol Chem 271, 13854-13860. Yoshida, H., Ochiai, M., and Ashida, M. (1986). Beta-1,3-glucan receptor and peptidoglycan receptor are present as separate entities within insect prophenoloxidase activating system. Biochem Biophys Res Commun 141, 1177-1184. Yu, X. Q., Gan, H., and Kanost, M. R. (1999). Immulectin, an inducible C-type lectin from an insect, Manduca sexta, stimulates activation of plasma prophenol oxidase. Insect Biochem Mol Biol 29, 585-597. Yu, X. Q., and Kanost, M. R. (2004). Immulectin-2, a pattern recognition receptor that stimulates hemocyte encapsulation and melanization in the tobacco hornworm, Manduca sexta. Dev Comp Immunol 28, 891-900. Zhang, R., Cho, H. Y., Kim, H. S., Ma, Y. G., Osaki, T., Kawabata, S., Soderhall, K., and Lee, B. L. (2003). Characterization and properties of a 1,3-beta-D-glucan pattern recognition protein of Tenebrio molitor larvae that is specifically degraded by serine protease during prophenoloxidase activation. J Biol Chem 278, 42072-42079. Zhang, Y., van der Fits, L., Voerman, J. S., Melief, M. J., Laman, J. D., Wang, M., Wang, H., Li, X., Walls, C. D., Gupta, D., and Dziarski, R. (2005). Identification of serum Nacetylmuramoyl-l-alanine amidase as liver peptidoglycan recognition protein 2. Biochim Biophys Acta. 58