Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
DNA vaccination wikipedia , lookup
Innate immune system wikipedia , lookup
Human leukocyte antigen wikipedia , lookup
Duffy antigen system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Gluten immunochemistry wikipedia , lookup
Adaptive immune system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Monoclonal antibody wikipedia , lookup
Antimicrobial peptides wikipedia , lookup
Major histocompatibility complex wikipedia , lookup
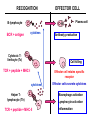
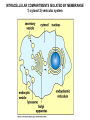
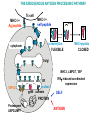
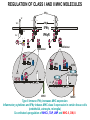
ANTIGEN PRESENTATION Lymphocyte subsets CLP T CELLS B CELLS Common lymphoid precursor T Th Activate B cells and macrophages T HELPER CELLS CTL Kill virusinfected cells CYTOTOXIC T LYMPHOCYTES B PC Produce antibodies PLASMA CELLS RECOGNITION EFFECTOR CELL Plasma cell B-lymphocyte BCR + antigen cytokines Cytotoxic Tlimfocyte (Tc) Antibody production Cell killing TCR + peptide + MHC-I Effector cell retains specific receptor cytokines Helper Tlymphocyte (Th) TCR + peptide + MHC-II Effector cells secrete cytokines Macrophage activation Lymphocyte activation Inflammation RECOGNITION OF EXOGENOUS AND ENDOGENOUS ANTIGENES BY T-LYMPHOCYTES Tc Th Exogenous Ag Endogenous Ag Peptides of endogenous proteins (virus, tumor) bind to class I MHC molecules Peptides of exogenous proteins (toxin, bacteria, allergen) bind to class II MHC molecules The number of different T cell antigen receptors is estimated to be 1,000,000,000,000,000 (1015 - 17) How can 6 invariant molecules have the capacity to bind to 1,000,000,000,000,000 different peptides? A flexible binding site? A binding site that is flexible enough to bind any peptide? At the cell surface, such a binding site would be unable to • allow a high enough binding affinity to form a trimolecular complex with the T cell antigen receptor • prevent exchange of the peptide with others in the extracellular milieu A flexible binding site? A binding site that is flexible at an early, intracellular stage of maturation formed by folding the MHC molecules around the peptide. Venus fly trap Floppy Compact Allows a single type of MHC molecule to • bind many different peptides • bind peptides with high affinity • form stable complexes at the cell surface • Export only molecules that have captured a peptide to the cell surface MHC molecules • Adopt a flexible “floppy” conformation until a peptide binds • Fold around the peptide to increase stability of the complex •The captured peptides contribute to the stabilization of the complex • Use a small number of anchor residues to tether the peptide - this allows different sequences between anchors and different lengths of peptides WHERE PEPTIDE BINDING OCCURS? INTRACELLULAR COMPARTMENTS ISOLATED BY MEMBRANSE 1) cytosol 2) vesicular system THE ENDOGENOUS ANTIGEN PROCESSING PATHWAY Tc-cell MHC-I + Ag peptide MHC-I + self peptide α-chain+β2m cytoplasm citopla zma FLEXIBLE MHC+peptide CLOSED Golgi MHC-I, LMP2/7, TAP ER TAP1/2 α-chain gp96 calnexin IFN induced coordinated expression SELF PROTEIN Proteasome LMP2/LMP7 ANTIGEN CYTOSOL-DERIVED PEPTIDES ARE PRESENTED BY MHC-I FOR T-CELLS Degradation of endogenous proteins in (immune) proteasomes TAP: Transporter associated with antigen processing Transporters associated with antigen processing (TAP1 & 2) Hydrophobic transmembrane domain Lumen of ER Peptide ER membrane Cytosol Peptide Peptide Peptide antigens from proteasome ATP-binding cassette (ABC) domain Transporter has preference for longer than 8 amino acid peptides with hydrophobic C termini. PEPTIDE-MHC INTERACTION IS SUPPORTED BY MULTIPLE PROTEINS IN THE ENDOPLASMIC RETICULUM TRIMMING OF PEPTIDES FOR OPTIMAL SIZE BY BY ERAP THE EXOGENOUS ANTIGEN PROCESSING PATHWAY Th-cell MHC-II + Ag peptide MHC-II + self peptide CLIP DMA/B Ii+αβ MCII CLIP DMA/B C IIV Golgi li li MHCII ER FLEXIBLE CLOSED INVARIANT CHAIN (Ii) 1. Chaperone – conformation 2. Inhibition of peptide binding 3. Transport/retention DMA/DMB 1. Support the peptide receptive conformation 2. Exchange of CLIP for exogenous peptides THE INVARIANT CHAIN PROTECTS THE MHC CLASS II BINDING SITE UNTIL REACHING THE APPROPRIATE COMPARTMENT DMA/DMB INVARIANT CHAIN LÁNC (Ii) 1. Chaperon – Conformation 2. Inhibition of the peptide binding site 3. Transport and retention 1. Stabilization of peptide accessable conformation 2. Exchange of CLIP to peptides derived from exogenous proteins GENERATION OF MHC – I EPITOPES GENERATION OF MHC – II EPITOPES HLA-DR1/HLA-DR4 Viral protein B35 A2 C42 B27 HLA-DQ2/HLA-DQ7 HLA-A,B,C binding Overlapping peptides The Tc response is focused to few epitopes The Th response is directed to overlapping epitopes ENSURE RECOGNITION OF MANY PATHOGENIC PROTEIN ENSURE RECOGNITION OF ALL PROTEINS TARGETS OF EPSTEIN-BARR VIRUS-SPECIFIC Tc (CTL) RESPONSES LATENT ANTIGENS + LMP2 ± (?) EBNA5 ± EBNA2 ++ ++ ++ EBNA3 EBNA4 EBNA6 - + EBNA1 LMP1 Nhet C W W W W W W H F QU PO M S L E ZR K B G D W WW W W Y LYTIC ANTIGENS BHRF1 ++ BMLF1 BZLF1 BMRF1 ++ ++ A Nhet TXV I BARF0 ++ • A poliklonális CTL válasz elsősorban a litikus antigének és az EBNA3,4,6 nukleáris fehérjék ellen irányul • Erősen fókuszált egy adott MHC - peptid kombinációra • Az endogén EBNA1 nem processzálódik és így nem ismerhető fel Bound peptide Generation of peptide MHC class I MHC class II source self or foreign proteins self or foreign proteins size 8-10 amino acids 13-21 amino acids heterogenity limited overlapping set of peptides natural cytoplasmic and nuclear proteins ~70% MHC derived, membrane and extracellular proteins site cytoplasm vesicles, endo/lysosomes enzyme proteasome LMP-2, LMP-7 regulatory subunits vesicular acidic proteases cathepsins transport TAP - size and C-terminal dependent cytoplasm ER no no Ii - target, retention ER vesicular system special compartment site ER vesicles, CIIV chaperons calnexin, toposin Ii - CLIP, DMA/B stable complexes reflecting the endogenous environment of the cell few instable empty molecules stable complexes reflecting the exogenous/endogenous environment of the cell few re-circulating molecules complexed with CLIP MHC transport MHC - peptide interaction MHC - peptide complexes In the cell surface REGULATION OF CLASS I AND II MHC MOLECULES IFNg TNF a a b JAK1 TRADD TRAF JAK2 p65 p55 IKK P IFNγ JAK1 IFNγR P–Y Ik B b IFNgR TNFRI JAK2 P–Y P Y–P P– Y Y–P P– Y p65 p55 STAT-dimer STAT-dimer Y–P P– Y NFkB A IRF-1 Y–P P– Y Y–P P– Y IRF-1 Co-a ktivátor Y–P p58 P– Y ISRE CIITA B W/S CAAT TATA RFX X2b p NF-Y X1 MHCI X2 Y CCAAT TATA MHCII Type II immune IFNγ increases MHC expression Inflammatory cytokines and IFNγ induces MHC class II expression in certain tissue cells (endothelial, astrocyte, microglia) Co-ordinated upregulation of MHC-I, TAP, LMP and MHC-II, DM, Ii B-cell T-cell Appearance of antigen Soluble, particles Any cell surface molecule Cells carrying self MHC-peptide complexes Nature of the antigén Natíve proteins, carbohydrate, lipids, metals, any structure Processed protein fragments = peptides Ligand Conformational determinant Ssequential determinant MHC-peptide complex Antigen recognizing receptor on the cell surface Variable BCR ligand (antigen) – spcific bivalent Variable TCR MHC + peptide pecific monovalent Soluble antigen recognizing receptor antibody - Collaboration of other cells - Antigen processing and presenting cells APC – interaction of two cells Antigen processing, presentation - Intracellular enzymatic degradation, peptide or MHC transportation Result of full activation Production of effector molecule antibody = soluble BCR Activation of new genes Activation molecules, production of lymphokines, TCR on the cell surface Possibililties of cell activation FULL plasma cell, antibody PARTIAL funcional anergy APOPTOSIS FULL Various lymphokines PARTIAL functional anergy certain lymphokines APOPTOSIS CD19, CD21, CD4, CD8, CD28/CTLA4, CD2, CD38 Co-receptors