Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
SNARE (protein) wikipedia , lookup
Resting potential wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Patch clamp wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Electrophysiology wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
G protein-gated ion channel wikipedia , lookup
University of Iowa
Iowa Research Online
Theses and Dissertations
Spring 2015
Targeting membrane proteins to inner segments of
vertebrate photoreceptors
Yuan Pan
University of Iowa
Copyright 2015 Yuan Pan
This dissertation is available at Iowa Research Online: http://ir.uiowa.edu/etd/1720
Recommended Citation
Pan, Yuan. "Targeting membrane proteins to inner segments of vertebrate photoreceptors." PhD (Doctor of Philosophy) thesis,
University of Iowa, 2015.
http://ir.uiowa.edu/etd/1720.
Follow this and additional works at: http://ir.uiowa.edu/etd
Part of the Biochemistry Commons
TARGETING MEMBRANE PROTEINS TO INNER SEGMENTS OF VERTEBRATE
PHOTORECEPTORS
by
Yuan Pan
A thesis submitted in partial fulfillment
of the requirements for the Doctor of
Philosophy degree in Biochemistry
in the Graduate College of
The University of Iowa
May 2015
Thesis Supervisor: Assistant Professor Sheila A. Baker
Graduate College
The University of Iowa
Iowa City, Iowa
CERTIFICATE OF APPROVAL
______________________
PH.D. THESIS
_________________
This is to certify that the Ph.D. thesis of
Yuan Pan
has been approved by the Examining Committee for the
thesis requirement for the Doctor of Philosophy degree
in Biochemistry at the May 2015 graduation.
Thesis Supervisor:
__________________________
Sheila A. Baker, Thesis Supervisor
Thesis Committee:
__________________________
Daniel Weeks, Chair
__________________________
Brandon S. Davies
__________________________
Kris A. DeMali
__________________________
Peter Rubenstein
__________________________
Amy Lee
To my parents, for their love and support
ii
ACKNOWLEDGMENTS
I would like to express my deepest appreciation to my thesis advisor Dr. Sheila
Baker, who initiated the project and provided me the guidance on continuing the
research. You have been a great mentor for me and your advice on both conducting
scientific research and personal development are priceless to me. I appreciate your
patient instructions and inspiring ideas. Without your supervision and constant support
this dissertation would not have been possible.
I am very grateful to my thesis committee members – Dr. Daniel Weeks, Dr.
Peter Rubenstein, Dr. Kris DeMali, Dr. Brandon Davies, Dr. Amy Lee and Dr. Heather
Bartlett – for your brilliant suggestions and inspirational discussions. Additional thanks
to members of the Baker lab: Joseph Laird, David Yamaguchi, Vasily Kerov, Modestos
Modestou and Sarah Hengel. Thank you all for your kind help and I adore the friendship
we developed in the lab.
Special thanks to my family: my grandfather-in law, my grandparents, my motherin law, my mother, my father and my husband. Words cannot express how I value all of
you for the sacrifices and efforts you made on my behalf. Last but not least, I would also
like to thank all of my friends especially Suifang Mao and Xu Liu. I treasure the time we
spent together and support each other towards our goals in this journey.
iii
ABSTRACT
Photoreceptors are highly compartmentalized neurons in the retina, and they
function by detecting light and initiating signaling through the visual network. The
photoreceptor contains several compartments including the outer segment (OS) which
is a sensory cilium for detecting photons and the inner segment (IS) that carries out
important modulatory functions via its resident channels and transporters. Those
proteins are membrane proteins that function together to shape electrical properties of
the cell membrane during both rest and active states. Therefore it is essential to
maintain proper function of the membrane proteins in the IS. One important way to
regulate the function of a membrane protein is via controlling its trafficking to ensure a
proper amount of the protein in the proper cellular compartment. To date, little is known
about how IS membrane protein trafficking is controlled in photoreceptors. In this study,
our goal is to understand those mechanisms using cell biology and biochemistry
approaches. To achieve the goal, we investigated trafficking of two unrelated IS resident
proteins: the hyperpolarization-activated cyclic nucleotide-gated channel 1 (HCN1) that
mediates a feedback current in photoreceptors, and the sodium potassium ATPase
(NKA) which maintains the basic electrochemical property of the cell.
In order to study trafficking of HCN1, we first investigated the dependence of
HCN1 trafficking in photoreceptors on TRIP8b, an accessory subunit that influences
trafficking of HCN1 in hippocampal neurons. By studying TRIP8b knockout mice we
found that TRIP8b is dispensable for HCN1 trafficking in photoreceptors but required for
maintaining the maximal expression level of HCN1. Since we revealed that HCN1
trafficking can be regulated in a cell-type specific manner, we subsequently focused on
iv
the amino acid sequence of HCN1 to identify novel trafficking signals that function in
photoreceptors. By examining localization of a series of HCN1 mutants in transgenic
Xenopus photoreceptors, we discovered a di-arginine ER retention motif and a leucinebased ER export motif. These two sequence motifs must function together to maintain
equilibrium of HCN1 level between the endomembrane system and the cell surface. The
study of HCN1 uncovered a mechanism for the photoreceptor to control membrane
protein trafficking via the early secretory pathways.
To reveal additional trafficking machineries in photoreceptors, we investigated
trafficking of NKA. We first tested for an interaction with ankyrin, an adaptor protein that
regulates NKA trafficking in epithelial cells, and found these proteins do not co-localize
in photoreceptors. We then aimed to identify novel trafficking signals by studying the
trafficking behavior of two NKA isozymes: NKAα3 and NKAα4. When expressed in
transgenic Xenopus photoreceptors, these two proteins localize to the IS and the OS
respectively. By studying localization of multiple chimeras and truncation mutants, we
found that the distinct localization pattern is due to a VxP OS/ciliary targeting motif
present in NKAα4. Since NKAα4 is naturally expressed in the ciliary compartment of the
sperm, our finding in the photoreceptor suggests a mechanism for NKAα4 trafficking in
its native environment. Overall, our studies of HCN1 and NKA together provide new
insights into controlling membrane protein trafficking in photoreceptors and help
establish the basics for future therapeutic intervention targeting trafficking pathways that
are linked to about one third of proteins reported in retinal diseases.
v
PUBLIC ABSTRACT
Photoreceptors are neurons responsible for detecting light in the eyes. The inner
segment (IS) is a specialized structure within a photoreceptor and contains several
resident membrane proteins that function to control electrical properties of the cell
membrane. The specialized localization of those proteins is controlled by protein
transport mechanisms and is critical for functions of the proteins. Our goal is to
understand how IS membrane proteins are transported in photoreceptors using cell
biology and biochemistry approaches. To achieve this goal, we investigated trafficking
of two unrelated IS proteins: the hyperpolarization-activated cyclic nucleotide-gated
channel 1 (HCN1) that is required for fast light responses, and the sodium potassium
ATPase (NKA) that shuttles ions across the cell membrane.
To study how HCN1 is transported, we first investigated the role of TRIP8b, a
regulator of HCN1 transport in hippocampal neurons. Using genetically modified mice
we found that TRIP8b is not required for proper HCN1 transport in photoreceptors.
Using genetically modified frogs we subsequently identified two amino acid sequences
within HCN1 itself that are required for transporting HCN1 from the inside of the cell to
the cell surface. To study how NKA is transported, we studied transport of two NKA
proteins that are transported to different places in the photoreceptor. Using genetically
modified frogs we found that this different transport phenomenon is due to a ciliary
transport sequence. Our findings provide new ideas in protein targeting and the basics
for future therapeutic tools targeting protein transport mechanisms.
vi
TABLE OF CONTENTS
LIST OF TABLES ........................................................................................................... xii
LIST OF FIGURES ........................................................................................................ xiii
LIST OF ABBREVIATIONS AND SYMBOLS ................................................................ xiv
CHAPTER I - INTRODUCTION ...................................................................................... 1
Functional Compartmentalization of Vertebrate Photoreceptors .................................. 1
Membrane Protein Trafficking ...................................................................................... 3
Regulation of Membrane Protein Trafficking in Photoreceptors ................................... 6
Function and Structure of HCN channels ..................................................................... 8
HCN1 Trafficking and TRIP8b.................................................................................... 10
Function and Structure of NKA .................................................................................. 12
NKA Trafficking and Ankyrin ...................................................................................... 14
Focus of the Thesis .................................................................................................... 16
CHAPTER II - ROLE OF TRIP8B IN REGULATING HCN1 TRAFFICKING IN
PHOTORECEPTORS ................................................................................................... 25
Abstract ...................................................................................................................... 25
Introduction ................................................................................................................ 26
Materials and Methods ............................................................................................... 28
Animals ................................................................................................................... 28
Immunohistochemistry ............................................................................................ 28
vii
Laser capture micro-dissection ............................................................................... 29
RT-PCR .................................................................................................................. 29
Fractionation of cytosolic and membrane proteins.................................................. 30
Immunoprecipitation ............................................................................................... 30
Biotinylation assay .................................................................................................. 31
Western blotting ...................................................................................................... 32
Electroretinography................................................................................................. 32
Results ....................................................................................................................... 33
Multiple TRIP8b isoforms are present in the retina ................................................. 33
Interaction of HCN1 and TRIP8b in the retina ........................................................ 35
HCN1 surface expression in the absence of TRIP8b.............................................. 37
Discussion.................................................................................................................. 38
CHAPTER III - IDENTIFICATION OF A DI-ARGININE ER RETENTION SIGNAL IN
THE C-TERMINUS OF HCN1 ....................................................................................... 48
Abstract ...................................................................................................................... 48
Introduction ................................................................................................................ 48
Materials and Methods ............................................................................................... 51
Animals ................................................................................................................... 51
Molecular cloning .................................................................................................... 52
Immunostaining ...................................................................................................... 52
viii
Cell culture .............................................................................................................. 53
Biotinylation assays ................................................................................................ 53
Results ....................................................................................................................... 54
The C-terminus of HCN1 redirects an integral membrane reporter to the ER in
Xenopus photoreceptors ......................................................................................... 54
The ER targeting signal is located within the intrinsically disordered region ........... 55
The ER retention signal regulates the trafficking of HCN1 to the cell surface......... 57
Discussion.................................................................................................................. 59
CHAPTER IV - AN N-TERMINAL ER EXPORT SIGNAL FACILITATES THE PLASMA
MEMBRANE TARGETING OF HCN1 CHANNELS IN PHOTORECEPTORS .............. 69
Abstract ...................................................................................................................... 69
Introduction ................................................................................................................ 70
Materials and Methods ............................................................................................... 71
Velocity sedimentation ............................................................................................ 71
Results ....................................................................................................................... 71
The N-terminus is required for HCN1 to target the ISPM........................................ 71
Identification of an ER export motif in HCN1 .......................................................... 73
The ER export motif overrides the function of the ER retention motif ..................... 76
Discussion.................................................................................................................. 77
ix
CHAPTER V – A VxP CILIARY TARGETING MOTIF IS REQUIRED FOR THE
DIFFERENTIAL TRAFFICKING OF NKAα3 AND NKAα4 ............................................ 89
Abstract ...................................................................................................................... 89
Introduction ................................................................................................................ 90
Materials and Methods ............................................................................................... 92
Molecular cloning .................................................................................................... 92
Animals ................................................................................................................... 92
Immunohistochemistry ............................................................................................ 93
Subcellular fractionation ......................................................................................... 93
Results ....................................................................................................................... 94
Distinct compartmentalization of ankyrins and NKAα in photoreceptors................. 94
The N-terminus of NKAα contains information for their differential trafficking......... 95
A VxP OS/ciliary targeting motif is required for NKAα4 to target the OS ................ 97
Discussion.................................................................................................................. 98
CHAPTER VI – DISCUSSION AND FUTURE DIRECTIONS ..................................... 108
Summary of the Thesis ............................................................................................ 108
Regulation of HCN1 by TRIP8b in Photoreceptors .................................................. 109
Other Candidates for Regulating HCN1 Trafficking in Photoreceptors .................... 111
Trafficking of HCN1 via the Early Secretory Pathways ............................................ 112
Potential Roles of Post-translational Modifications in HCN1 Trafficking .................. 115
x
Role of Ankyrin in the Retinal Localization of NKA................................................... 117
Trafficking of NKAα to the ISPM of Photoreceptors ................................................. 118
Conclusions ............................................................................................................. 120
REFERENCES ............................................................................................................ 126
xi
LIST OF TABLES
Table 1-1. Roles of TRIP8b isoforms in regulating HCN1 trafficking ............................. 20
Table 3-1. A summary of the design and targeting behaviors of all constructs
expressed in transgenic Xenopus photoreceptors ....................................... 63
Table 4-1. A summary of the design and targeting behaviors of all constructs
expressed in transgenic Xenopus photoreceptors ....................................... 82
Table 5-1. A summary of NKA constructs expressed in transgenic Xenopus
photoreceptors ........................................................................................... 102
Table 6-1. A summary of interactions between di-basic ER retention motifs and
COPI or 14-3-3 proteins ............................................................................. 122
xii
LIST OF FIGURES
Figure 1-1. Compartmentalization and function of vertebrate photoreceptors ............... 21
Figure 1-2. Major membrane protein trafficking pathways in the cell............................. 22
Figure 1-3. Structure and retinal localization of HCN1 .................................................. 23
Figure 1-4. Structure and retinal localization of NKAα................................................... 24
Figure 2-1. TRIP8b co-localizes with HCN1 in the mouse retina................................... 41
Figure 2-2. Multiple TRIP8b splice variants are expressed in retina ............................. 42
Figure 2-3. Multiple TRIP8b splice variants are expressed in photoreceptors............... 43
Figure 2-4. HCN1 is required to fully recruit TRIP8b to the membrane ......................... 44
Figure 2-5. HCN1 protein levels are reduced in the absence of TRIP8b ....................... 45
Figure 2-6. Flicker frequency responses are normal in the absence of TRIP8b ............ 46
Figure 2-7. The surface expression of HCN1 is maintained in TRIP8b-/- mice .............. 47
Figure 3-1. The C-terminus of HCN1 directs localization to the ER .............................. 64
Figure 3-2. The C-linker and CNBD are not required for ER localization ...................... 65
Figure 3-3. Identification of a di-arginine ER retention signal ........................................ 66
Figure 3-4. The di-arginine motif influences plasma membrane localization of HCN1 .. 67
Figure 3-5. Mutating the di-arginine motif enhances surface expression ...................... 68
Figure 4-1. The N-terminus drives a membrane-associated reporter to the IS.............. 83
Figure 4-2. The N-terminus is required for HCN1 to target the ISPM ............................ 84
Figure 4-3. Identification of a VxxxSL ER export motif in HCN1.................................... 85
Figure 4-4. A bulky hydrophobic residue is key for the ER export signal to function ..... 86
Figure 4-5. Disrupting the ER export motif does not affect the assembly of HCN1 ....... 87
Figure 4-6. The ER export signal counters the action of the ER retention signal .......... 88
Figure 5-1. The interaction between ankyrins and NKA in the bovine retina ............... 103
Figure 5-2. The N-terminus of NKAα3 and/or NKAα4 determines their distinct
localization ................................................................................................ 104
Figure 5-3. The first 14 amino acids of NKAα3 and/or α4 contain the targeting
information ................................................................................................ 105
Figure 5-4. The first 14 amino acids of NKAα4 are required for its OS localization..... 106
Figure 5-5. Identification of a VxP OS targeting motif in NKAα4 ................................. 107
Figure 6-1. A model for photoreceptor trafficking of HCN1 and NKA .......................... 123
Figure 6-2. A proposed model for HCN1 trafficking via early secretory pathways ....... 124
Figure 6-3. Regions of NKAα3 with putative ISPM trafficking signals ......................... 125
xiii
LIST OF ABBREVIATIONS AND SYMBOLS
A (Ala)
aa
AnK
cDNA
CNG
COP
CT
DDM
DMEM
DNA
DTT
EDTA
EGTA
ER
ERG
ERGIC
F (Phe)
FBS
GAPDH
GC
GFP
HA
HCN
HEK293
I (Ile)
Ih
INL
IPL
IS
ISPM
KATP
kDa
Kir
L (Leu)
LDS
MMR
MUT
N
N (Asn)
NaCl
NKA
NT
ONL
OPL
Alanine
Amino acid
Ankyrin
Complementary DNA
Cyclic nucleotide gated channel
Coat protein complex
C-terminus
n-Dodecyl-beta-D-maltoside
Dulbecco's Modified Eagle's medium
Deoxyribonucleic acid
Dithiothreitol
Ethylenediaminetetraacetic acid
Ethylene glycol tetraacetic acid
Endoplasmic reticulum
Electroretinography
ER-Golgi intermediate compartment
Phenylalanine
Fetal Bovine Serum
Glyceraldehyde 3-phosphate dehydrogenase
Ganglion cells
Green fluorescent protein
Hemagglutinin
Hyperpolarization activated cyclic nucleotide channel
Human embryonic kidney 293
Isoleucine
Hyperpolarization-activated current
Inner nuclear layer
Inner plexiform layer
Inner segment
Inner segment plasma membrane
ATP-sensitive potassium channels
Kilodalton
Inward rectifier potassium channel
Leucine
Lithium dodecyl sulfate
Marc’s Modified Ringer
Mutant
Nuclei
Asparagine
Sodium chloride
Sodium potassium ATPase
N-terminus
Outer nuclear layer
Outer plexiform layer
xiv
OS
P (Pro)
PBS
PCR
PDC
PM
R (Arg)
RNA
RT
S (Ser)
SDS-PAGE
ST
SUR
TMD
TRIP8b
Tris-HCL
V (Val)
WT
X. laevis
X. tropicalis
Outer segment
Proline
Phosphate buffered saline
Polymerase chain reaction
Phosducin
Plasma membrane
Arginine
Ribonucleic acid
Reverse transcriptase
Serine
Sodium dodecylsulfate polyacrylamide gel electrophoresis
Synaptic terminal
Sulfonylurea receptor
Transmembrane domain
Tetratricopeptide repeat-containing Rab8b interacting protein
Tris-Hydrochloride
Valine
Wildtype
Xenopus laevis
Xenopus tropicalis
α
β
γ
δ
ɛ
ζ
η
θ
alpha
beta
gamma
delta
epsilon
zeta
eta
theta
xv
CHAPTER I - INTRODUCTION
Functional Compartmentalization of Vertebrate Photoreceptors
The retina contains multiple types of neurons that are organized in a layer-wise
structure to constitute an efficient vision system. Photoreceptors are the light-sensing
neurons present at the outermost layer of the retina. There are two types of
photoreceptors: rods and cones. They differ in sensitivity, acuity and color detection.
Rods have high sensitivity while cones have higher accuracy and can detect colors.
Because of their unique characteristics, rods function primarily at low light intensities
and cones function at medium to bright light intensities.
Rods and cones share a common cellular organization consisting of four major
compartments: an outer segment (OS), an inner segment (IS), a nucleus and a synaptic
terminal (ST) (Figure 1-1). This specialized compartmentalization separates the cellular
events in a temporal and spatial manner. The OS is a sensory cilium responsible for
photon detection. In the dark, there is a dark current circulating in the cell, resulting in
depolarization of the cell membrane and the tonic release of neurotransmitters from the
ST. The dark current is mainly carried by the Na+ and Ca2+ ions coming from the cyclic
nucleotide gated (CNG) channel in the OS (Figure 1-1A). Upon light stimulation, the
cells are hyperpolarized due to the closure of CNG channels and subsequent inhibition
of the dark current. Consequently, neurotransmitter release from the ST is inhibited
(Figure 1-1B).
The IS is most analogous to a generic cell soma, yet it contains membrane
proteins, especially channels and transporters, critical for shaping the membrane
potential both in dark and light conditions. In the dark, the charge of Na+ influx through
1
CNG channels is balanced by the outflow of K+ carried by potassium channels (e.g. the
intermediate and large conductance calcium-activated potassium channels1) in the IS.
At the same time, the sodium potassium ATPase (NKA) located in the IS continuously
pumps Na+ out of the cell while K+ into the cell to compensate for the change of
intracellular Na+/K+ concentration due to the dark current (Figure 1-1A).
Pharmacological inhibition of NKA functions abolishes typical retinal responses and
causes photoreceptor degeneration2.
Upon light stimulation, there is a transient hyperpolarization of the photoreceptor
membrane (Figure 1-1B). The membrane potential is repolarized rapidly in part by the
activities of the hyperpolarization activated cyclic nucleotide gated channel 1 (HCN1)
located in the IS (Figure 1-1C). HCN1 opens when the cell membrane is hyperpolarized
and carries a feedback current (influx of K+ and Na+ ions) that resets the membrane
potential between dark and light conditions3,4,5. As expected, HCN1 knockout mice
show prolonged light responses and present impaired vision at medium to bright light
intensities4,5. Patients taking ivabradine, a pharmacological inhibitor of HCN channels
for treating heart diseases, occasionally report phosphene as a visual side effect6.
Another potassium channel that complements the function of HCN1 is the heteromeric
channel consisting of Kv2.1 (voltage gated potassium channel 2.1) and KCNV2
(potassium channel, subfamily V, member 2), which have mutations identified in
patients with cone dystrophy with supernormal rod response7. Altogether, it is clear that
maintaining the proper function of those IS membrane proteins is key to supporting the
normal function of photoreceptors in vision.
2
Membrane Protein Trafficking
There are multiple ways to regulate the function of membrane proteins.
Regulating their trafficking has proved to be an important aspect in photoreceptors, as
well as in other cellular systems. In fact, many pathways controlling membrane protein
trafficking are shared by various functionally distinct cells (Figure 1-2). Integral
membrane proteins are generally co-translationally synthesized in the ER, followed by a
folding process assisted by ER chaperones. Subsequently, the protein is sorted to ER
exit sites and forwarded to the ER Golgi intermediate compartment (ERGIC) and the
Golgi apparatus, a process called anterograde transport. In complement to the
anterograde transport, a retrograde transport process takes place in the ERGIC and
Golgi to retrieve ER-resident and poorly assembled proteins back to the ER. These
processes are referred to as the early secretory pathways, where coat protein (COP)
complexes are the key players in forming and shipping the transport vesicles. COPII
and COPI complexes carry cargoes in the anterograde and retrograde transport
pathways respectively.
Components of the COPII complex decorate the ER exit sites and facilitate
formation and budding of those vesicles in the anterograde transport pathway8,9. There
are five major components of the COPII complex and they form three sub-complexes.
They are the inner coat complex Sec24/Sec23 that selects for cargoes, the outer coat
complex Sec13/Sec31 and the GTPase Sar1. COPI shares conceptual similarities with
COPII but differs in the coat structure. The system contains at least seven COPI
subunits (α, β, β’, ɛ, γ, δ and ζ) and requires the function of the small GTPase Arf1.
Those subunits form two major complexes: the B-subcomplex (αβ’ɛ) that functions
3
similar to the outer coat of the COPII complex and the F-subcomplex (βγδζ) that is
implicated in cargo binding10. Two categories of trafficking signals were described for
guiding membrane protein trafficking within the early secretory pathways, namely ER
export signals and ER retention/retrieval signals (hereafter referred to as ER retention
signal since the net effect of the signal is ER retention).
It was proposed that properly folded and assembled membrane proteins leave
the ER via two modes. One is the selective active transport mode and the other is a
slower ‘bulk flow’ mode with less selectivity. The active transport mode applies to
membrane proteins containing ER export signals that recruit COPII components to
achieve fast ER export. Proteins that do not contain specific ER export signals leave the
ER via the bulk flow mode. Conventional ER export signals include di-acidic and
hydrophobic motifs8. For example, the most popular protein model for studying
trafficking, the vesicular stomatitis virus (VSV)-G protein, contains a di-acidic (DxE) ER
export motif that is required for its ER to Golgi transport11. Another class of proteins that
have been studied intensely are the p24 proteins which contain hydrophobic (FF) ER
export motifs12,13. Many ER export signals function via interacting with Sec24 proteins in
the COPII complex.
Properly folded and assembled proteins are forwarded to the ERGIC/Golgi. What
happens to misfolded and unassembled proteins in the ER? It is known that misfolded
proteins undergo re-folding processes with the help of ER-resident chaperons (e.g.
BiP), or are subjected to ER-associated degradation. However the mechanism of how
poorly assembled proteins are retained in the ER is hardly understood, and a second
degree of control is required because some unassembled proteins can escape the ER
4
and enter the ERGIC/Golgi presumably via ‘bulk-flow’. Fortunately, the cell has the
specific retrograde transport system built for such proteins. The KDEL (Lys-Asp-GluLeu) motif is a classic ER retention signal and is present in many lumenal ER-resident
proteins including glucose-regulated protein-78 (GRP78), GRP94 and protein disulphide
isomerase (PDI)14. Proteins with the KDEL motif are recognized by the KDEL receptors
at the ERGIC/Golgi. The loaded KDEL receptor further interacts with the COPI
components, resulting in retrograde transport of the ER-resident proteins15-17.
Another class of ER retention signals are the di-basic (typically a combination of
lysine and arginine amino acids) motifs that were identified in both ER-resident and nonER-resident integral membrane proteins18. Some of these motifs found in multimeric
complexes were proposed to monitor the subunit assembly state. Using the heteromeric
ATP-sensitive potassium (KATP) channels containing Kir6.2 (inward rectifier potassium
channel 6.2) and SUR1 (sulfonylurea receptor 1) as the model, Zerangue et al., showed
that the di-arginine motifs present in both subunits will inhibit their ER export unless they
are assembled into a functional heterooctamer19,20, suggesting that a properly folded
and assembled protein is able to mask the ER retention signals.
Once the protein reaches the Golgi, it is further glycosylated and sorted into
transport vesicles and targeted to various compartments depending mainly on the
trafficking signals present at the cytoplasmic face of the protein. Compared to the
knowledge about carriers in the ER-Golgi transport, little is known about carriers
facilitating Golgi-plasma membrane (PM) transport. A complex named carriers from the
trans Golgi network to the cell surface (CARTS) was discovered by Wakana et al., in
2012. The CARTS vesicles contain myosin II, Rab6a, Rab8a and synaptotagmin II, in
5
addition to a number of secretory and integral membrane proteins. But CARTS is not
the only carrier in this pathway, because the Golgi-PM transport of collagen and VSV-G
is independent of CARTS21. Membrane proteins at the PM are internalized eventually
via the endocytic pathways. Some undergo clathrin-dependent endocytosis. The clathrin
coated vesicles share structural similarities with the COPI coatomer. Many clathrin
independent pathways were also reported, suggesting there is great diversity in
regulating protein endocytosis22. The endocytosed membrane proteins can re-enter the
PM via the recycling pathway or be degraded via the lysosomal pathway.
Regulation of Membrane Protein Trafficking in Photoreceptors
Membrane protein trafficking in the early secretory pathways is generally
consistent across cell types. In contrast, trafficking pathways at later stages vary
dependent on both the protein and the cell type. Distinct cellular compartmentalization
usually correlates with the cell’s specific function. Epithelial cells are compartmentalized
into apical and basolateral domains to ensure polarized transport of materials. Neurons
are compartmentalized into dendrites, soma, and axons to achieve vectorial
communication; photoreceptors are similarly organized because they are sensory
neurons. The OS is anatomically a sensory cilium but it functions similar to a dendrite.
The IS is the cell soma where the protein production organelles (e.g. ER and Golgi) are
located. The IS is followed by the axon and the synaptic terminal that contains a ribbon
synapse for the tonic release of neurotransmitters. Although the synapses in
photoreceptors are atypical, the principle of synaptic communication stays the same as
in other neurons.
6
Just like any other type of neuron, photoreceptors are very sensitive to changes
in protein compartmentalization. Mistargeted membrane proteins in the photoreceptor
can lead to cell malfunction and often degeneration, resulting in vision loss. The
trafficking pathway of the photoreceptor specific protein, rhodopsin, has been the most
extensively studied. Rhodopsin is the light-sensing pigment that functions as a Gprotein coupled receptor in the OS. Light-activated rhodopsin triggers the activation of
the downstream phototransduction effectors including transducin, cGMP
phosphodiesterase and the CNG channels. The C-terminal end of rhodopsin has been
shown to be a hotspot for genetic mutations in patients with autosomal dominant retinitis
pigmentosa, an early-onset, severe form of retinal degeneration.
Rhodopsin is localized exclusively to the OS except for the trace amount of newly
synthesized protein present in the IS. Researchers have found that the C-terminal end
of rhodopsin contains a VxP OS/ciliary motif that is required for concentrating rhodopsin
in the OS. Genetic mutations in rhodopsin resulting in change or loss of the VxP motif
can cause mistrafficking of rhodopsin, demonstrated as a diffused localization through
both the IS and OS, forbidding rhodopsin from fully functioning in the phototransduction
cascade13,23. Photoreceptors expressing the rhodopsin trafficking mutants undergo
degeneration either due to the overloaded protein burden in the IS or malfunction of the
phototransduction pathway. In addition to the VxP OS/ciliary motif, membrane
association via palmitoylation and myristoylation is also required for fully targeting
rhodopsin to the OS24, indicating that membrane protein trafficking is usually achieved
by multiple trafficking signals.
7
The fact that the rhodopsin mutants lacking the OS targeting signals are found in
both the IS and OS implies that specific signals are needed to target any compartment
in the cell. Consistent with this idea, Baker et al., showed that other membrane proteins
lacking their trafficking signals are present in both the IS and OS using Xenopus
photoreceptors as the model25. While a number of OS targeting signals have been
identified in multiple OS-resident proteins26-28, nothing is known about how trafficking to
the IS is regulated. For this reason, we set our goal to understand how trafficking of ISresident membrane proteins is controlled. The IS harbors multiple membranous subcompartments including the ER, Golgi and the inner segment plasma membrane
(ISPM). Identity of targeting signals to any of the IS sub-compartments will be helpful to
understand IS membrane protein trafficking. In this study, we chose two unrelated IS
localized membrane proteins - HCN1 and NKA - as our model to study trafficking to
various IS organelles. Both proteins carry out important functions in photoreceptors as
well as other systems, therefore the mechanisms discovered in the photoreceptor may
be applied to other cells in the future.
Function and Structure of HCN channels
HCN channels belong to the voltage-gated potassium channel superfamily. They
are activated by hyperpolarization and modulated by cyclic nucleotides. Upon activation,
HCN carries a weak selective cation current named as If (funny current) in the heart and
contributes to the cardiac pacemaker activity29. The current is also referred to as Ih
(hyperpolarization-activated current) in neurons and Iq (queer current) in retina; in both
places HCN regulates signal integration and transmission30. There are four HCN
proteins, HCN1, 2, 3 and 4, that share sequence and structure similarities but vary in
8
activation kinetics and cyclic nucleotide sensitivity. For instance, HCN1 is the fastest
activating HCN channel with the least sensitivity to cyclic nucleotides. In addition to the
difference in channel activity, the tissue expression pattern of each HCN channel also
varies. In the heart, HCN4 and HCN2 are the principle carriers of If31. In the brain, HCN1
and HCN2 are the most abundant HCN proteins yet with distinct expression profiles in
different neuron populations. In the retina, all members of the HCN family were found in
the retina although with distinct expression profiles5,32-42. Rods express only HCN1.
Cones express HCN1 and HCN3. HCN1, 2 and 4 are all found in bipolar cells and
ganglion cells but the expression pattern in sub-types of those cells varies.
All HCN channels function as tetramers; they can either form homo- or heterotetramers depending on the cell type and cellular activities43-47. HCN monomers share
the same structure consisting of six transmembrane domains (TMD) that are the
building blocks of the channel (Figure1-3A). The channel’s voltage sensing region lies in
the fourth TMD, and the channel pore which conducts ions lies between the fifth and
sixth TMD. The cytoplasmic regions of HCN channels modulate their gating and
trafficking properties. The C-terminus is the largest cytoplasmic region (~460aa) of HCN
and contains two structured domains: the C-linker and the cyclic nucleotide binding
domain (CNBD). The C-linker regulates gating of HCN channels via the cyclic
nucleotide binding activity of the CNBD. Following the C-linker and the CNBD, there is
an intrinsically disordered region - the post-CNBD region - that contains sites for
protein-protein interactions and is poorly conserved among the different HCN proteins.
Multiple proteins (filamin A, Nedd4-2 and TRIP8b) have been shown to interact
with the post-CNBD region of HCN1 and affect its trafficking in various ways (Figure1-
9
3A). Filamin A is an actin binding protein that plays a scaffolding role in cells48,49. It was
recently found that filamin A inhibits the surface expression of HCN1 in hippocampal
neurons by promoting internalization50. A ubiquitin ligase, Nedd4-2 (neuronal precursor
cell expressed developmentally downregulated), can also suppress HCN1 surface
expression when tested in HEK293 cells; likely by promoting HCN1 ubiquitination and
degradation51. Among all proteins that interact with HCN1, the tetratricopeptide repeatcontaining Rab8b-interacting protein (TRIP8b) is the most multi-functional regulator due
to more than one interaction site in HCN1 and a large number of TRIP8b isoforms.
HCN1 Trafficking and TRIP8b
In photoreceptors, HCN1 is excluded from the OS which functions analogously to
a dendrite, and is concentrated in the IS which is anatomically similar to the cell soma of
a pyramidal neuron (Figure 1-3B). There are multiple reasons why HCN1 may be
excluded from the OS. Because of its specific function in the phototransduction
cascade, the OS is subject to high levels of photons and free radicals. Therefore,
approximately 10% of the OS is renewed each day so that the cytotoxicity induced by
the light stimulation does not build up in the photoreceptor52. Proteomic studies in the
OS suggested that there are only a small number of different membrane proteins
present in the OS53. One possible reason for limiting the types of proteins in the OS is to
limit the number of proteins needed to be renewed constantly, leaving proteins that are
not directly involved in the phototransduction cascade to general mechanisms of protein
turnover. Another possible reason for restricting types of proteins in the OS is that
changes in the phototransduction pathway can easily affect vision and health of
photoreceptors. Expression of ectopic membrane proteins such as HCN1 can very likely
10
interfere with the function of OS resident membrane proteins. Importantly, HCN1 binds
and responds to cyclic nucleotides. HCN1 is primarily modulated by cAMP (K1/2 ~
0.5µM, K1/2 defined as the concentration required to achieve half of the maximal
channel activity)56-58, which is concentrated in the IS59, but HCN1 can also interact with
cGMP (K1/2 ~ 6µm)58 which is an important regulator of CNG (K1/2 ~ 1mM)60 channels in
the OS. If HCN1 is expressed in the OS, the competition between HCN1 and CNG for
cGMP may result in an alteration of the dark current or even degeneration of
photoreceptors.
When reviewing HCN1 localization in the central nervous system, it is intriguing
that distinct HCN1 compartmentalization can be observed5,33,61-63. HCN1 can
preferentially localize to axons, soma or dendrites depending on the cell type. In
photoreceptors, HCN1 localizes to the soma (IS) and synaptic terminal; however, in
hippocampal neurons, HCN1 is concentrated at the distal dendrites64. More surprisingly
in the inner ear cell, HCN1 is found in the stereocilium, an actin-based structure61,62.
The distinct localization pattern of HCN1 suggests a cell-type specific regulation of
HCN1 trafficking.
TRIP8b was identified as an accessory subunit of HCN channels and regulates
both their gating and trafficking. Trafficking of HCN1 in neurons can either be TRIP8bdependent or TRIP8b independent65,66. It was shown that the polarized localization of
HCN1 in the distal dendrites of hippocampal neurons is primarily contributed by
TRIP8b67,68. In the absence of TRIP8b, HCN1 is evenly distributed through the entire
dendritic tree. Additionally, in the medial perforant path, TRIP8b inhibits the axonal
distribution of HCN1. However, trafficking of HCN1 to the synaptic terminals of
11
entorhinal cortical neurons is not affected in the absence of TRIP8b. These previous
studies suggest that TRIP8b contributes to the cell-type specific trafficking of HCN1.
One important factor leading to the cell-type specific regulation of HCN1 via
TRIP8b is the presence of many TRIP8b isoforms. The isoforms come from a
combination of different promoter usage and alternative splicing (Table 1-1). All TRIP8b
isoforms downregulate the gating of HCN1 channels by binding to the CNBD, yet they
regulate HCN1 trafficking diversely via interacting with the last three amino acids (SNL)
of HCN1. Some of the isoforms can both affect the layer-wise compartmentalization of
HCN1 as well as trafficking of HCN1 from the internal compartments to the plasma
membrane (i.e. surface expression). Given the diverse function of TRIP8b proteins,
different neurons may express certain isoforms that together lead to cell-type specific
regulation of HCN1 trafficking. Therefore it will be very helpful to test in photoreceptors
the expression of TRIP8b isoforms and their roles in HCN1 trafficking.
Function and Structure of NKA
Another important IS membrane protein is NKA whose ubiquitous function is to
pump Na+ and K+ ions across the cellular membrane in opposite directions to create an
electrochemical gradient. NKA is also a membrane receptor of cardiotonic steroids and
neuronal agrin, thus is involved in a number of cellular signaling pathways69-71.
Additionally, NKA plays a role in cellular adhesion via interacting with a secreted
protein, retinoschisin that is affected in X-linked juvenile retinoschisis, a progressive
degenerative disease characterized by splitting of the retinal layers and consequently
decreased neurotransmission from photoreceptors to downstream neurons72,73,74.
12
The minimal functional unit of NKA is a heterodimer containing an α subunit and
a β subunit, both of which are integral membrane proteins (Figure1-4A). NKAα is the
catalytic subunit that carries out ATP hydrolysis and active transport of ions. The α
subunit contains ten transmembrane domains and three cytoplasmic domains: the
actuator (A) domain, the nucleotide-binding (N) domain and the phosphorylation (P)
domain75-78. The catalytic mechanism of NKAα starts with ATP binding (at the N
domain) followed by binding to three Na+ ions in the cytoplasm. ATP hydrolysis leads to
phosphorylation of an aspartate residue in the P domain. Subsequent release of ADP
triggers a conformational change that releases Na+ ions to the extracellular space. At
this conformation, two K+ ions bind to the protein and induce dephosphorylation which
changes the protein conformation back to the original state and releases the K+ ions to
the cytoplasm. The A domain is responsible for dephosphorylation of the P domain. The
NKAβ subunit contains a single transmembrane domain and its extracellular N-terminus
is highly glycosylated. The β subunit is involved in regulating the catalytic activity and
ER export of the NKA complex.
There are four and three genes encoding for the α and β subunits respectively
with tissue-specific expression patterns79-88. The NKAα isozymes share the basic
functions but vary in affinity to small molecules such as ions and ouabain, the
pharmacological inhibitor of NKA. NKAα1 and β1 are ubiquitously expressed and are
the predominant NKA in the kidney. NKAα2 is enriched in cardiac and skeletal muscles.
It is also found in adipocytes and the brain. NKAα3 is the most abundant and restricted
isozyme in the nervous system including photoreceptors. NKAα4 is exclusively
expressed in mammalian sperm. NKAβ2 is expressed in skeletal muscle and the
13
nervous system. NKAβ3 is found in multiple tissues including retina, testis, lung and
liver. In photoreceptors, NKAα3β2 is the dominant form. In some tissues, a third
regulatory γ subunit further modulates the activity of the NKA complex.
NKA Trafficking and Ankyrin
Subcellular localization of NKA varies depending on the cell type. In most
epithelial cells NKA is found in the basolateral membrane to ensure vectorial ion
transport (e.g. NKAα1β1 in the tubule cells of the kidney89) but in rare cases such as in
the retinal pigment epithelium, NKAα1β1 is found in the apical domain. In sperm,
NKAα4 is found in the mid-piece of the flagellar tail, a ciliated organelle that controls the
sperm mobility86,87,90,91. Loss of NKAα4 results in low sperm motility and infertility. In
photoreceptors, NKAα3 is restricted to the inner segment plasma membrane (ISPM)
(Figure 1-4B). One interesting localization pattern in photoreceptors is that the
mitochondria is restricted to a region named ellipsoid in the apical portion of the IS and
close to the bottom of the OS. In the photoreceptor, most energy is consumed by NKA
and synaptic activities in the dark while by the phototransduction cascade in the light92.
It was shown that in many neurons, mitochondria localization is determined by local
demands of energy93,94. The reason why mitochondria are localized in the specific
region of the photoreceptor is probably to fulfill the large energy demands from both
NKA and the phototransduction cascade. The close proximity of mitochondria to both
the OS and IS would seemingly limit the amount of ATP available to the synapse, but
this potential problem is minimized by having energy flow from the IS to the synapse in
14
the form of phosphocreatine95. Given the important functions of NKA in all cell types, it is
surprising how little is known about how its sub-cellular localization is regulated.
What is known about NKA trafficking remains mainly in the early secretory
pathway and PM tethering of the pump. The NKAβ subunit plays roles in both steps. For
instance, ER export of NKA requires assembly of the αβ heterodimer in a one to one
ratio96-98. In the absence of NKAβ, ER retention of NKAα is achieved by a di-basic ER
retention motif present at its N-terminus. This ER retention motif functions by binding to
β-COP, a component of the COPI complex10. NKAα with the di-basic motif mutated is
able to traffic from the ER to the PM; however it is not functional without associating
with the β subunit. After the intact NKAαβ heterodimer arrives at the PM, it was shown
that the β subunit stabilizes their localization via intercellular homotypic interactions (i.e.
two NKAβ proteins in adjacent cells interact with each other). It is clear that the β
subunit is critical for exporting NKA from the endomembrane system and securing its
localization at the PM, however, at least one additional signal is needed to guide the
NKA complex to specific regions of the PM.
Ankyrin (Ank) is a family of adaptors between membrane proteins and the
spectrin-based cytoskeleton99-101. Ankyrins are crucial for trafficking of several
membrane proteins like voltage-gated calcium channel Cav1.3, KATP, as well as NKA102107.
There are three ankyrin proteins: AnkR, AnkG, and AnkB. They are thought to
facilitate NKA trafficking in multiple cellular systems101,102,104. In epithelial cells, AnkR
facilitates the ER to Golgi trafficking of NKAα1 via direct binding. The subsequent
trafficking from Golgi to the PM is assisted by AnkG. In cardiomyocytes, AnkB organizes
a signaling complex that includes NKAα1 and NKAα2 at the T-tubule/SR
15
microdomain106,108. In Madin-Darby canine kidney (MDCK) cells, ischemia induced loss
of NKA polarity is likely due to the dissociation between spectrin and AnkG109. In the
retina, AnkB has been proposed to regulate the expression of NKA since a reduced
level of NKA protein was observed in retina of AnkB+/- mice107. Whether any of the
ankyrin proteins can influence NKA trafficking in photoreceptors requires further
investigation.
Focus of the Thesis
The ultimate goal of this research is to understand how trafficking of IS
membrane proteins is regulated at various stages. There may exist a general control
system that is shared by a number of IS membrane proteins and even OS membrane
proteins at some stages such as early secretory pathways. Understanding such
regulation will allow us to dissect the principles underlying membrane protein trafficking
in general and help identify master regulators in the regulatory pathways. At some other
stages, for example the OS targeting pathway, trafficking of each membrane protein
may be regulated differently. Identification of protein specific regulation is also valuable,
because it allows us to understand mechanisms behind specific genetic mutations that
result in trafficking defects.
In this study, we aimed at identifying trafficking regulators (either generic or
specific) that are involved in membrane protein trafficking in the photoreceptors. The
thesis focuses on examining trafficking of two unrelated IS membrane proteins: HCN1
and NKA. On one hand, we studied the influences on their trafficking in vertebrate
photoreceptors by the candidate regulatory proteins (TRIP8b for HCN1 and ankyrins for
NKA); On the other hand, we utilized a specific approach in transgenic Xenopus
16
photoreceptors to identify novel trafficking signals/motifs that control the localization of
HCN1 and NKA in vivo.
Chapter II focuses on studying the interaction between HCN1 and TRIP8b in
mouse photoreceptors. We first characterized the expression profile of TRIP8b isoforms
in the entire retina as well as specifically in photoreceptors by RT-PCR and
immunohistochemistry. Later by studying HCN1 compartmentalization and surface
expression in TRIP8b knockout mice, we found that TRIP8b does not affect trafficking of
HCN1 in photoreceptors. However, it is important for maintaining the expression level of
HCN1. In the absence of TRIP8b, there is an approximately 40% reduction of both total
and surface HCN1. Interestingly, the remaining HCN1 at the cell surface is sufficient to
support vision as measured by electroretinography (ERG), suggesting that the
photoreceptor tolerates this down-regulation of HCN1. This study was reported in a
manuscript entitled “TRIP8b is required for maximal expression of HCN1 in the mouse
retina” published in PLOS ONE on January 7, 2014110.
Since HCN1 trafficking in photoreceptors is TRIP8b independent, we sought for
novel trafficking signals located within the protein sequence of HCN1. We used a
transgenic Xenopus approach, which has the advantages of large size and short
generation time, to study which regions of HCN1 are required for its trafficking in
photoreceptors. As a result we identified two novel signals in trafficking HCN1 via the
early secretory pathways. In Chapter III, the trafficking behavior of the C-terminus of
HCN1 was dissected and we uncovered a di-arginine ER retention motif in the postCNBD region. HCN1 with a defective ER retention motif shows higher surface/total ratio
in HEK293 cells and is able to target the ISPM of Xenopus photoreceptors. This
17
localization of HCN1 at the PM of photoreceptor IS is due to a leucine-based ER export
motif we later identified in Chapter IV. When the ER export signal is mutated in HCN1,
the ER retention signal dominates and retains the protein in the ER. These two signals
together make a push-pull control system to regulate the amount of HCN1 being
transported from the ER to the PM.
The contents of Chapter III are adapted from a manuscript published in Cellular
and Molecular Life Sciences on August 21, 2014 entitled “A di-arginine ER retention
signal regulates trafficking of HCN1 channels from the early secretory pathway to the
plasma membrane”111. The findings in Chapter IV are reported in a manuscript named
“An N-terminal ER export signal facilitates the plasma membrane targeting of HCN1
channels in photoreceptors” that was accepted by IOVS on April 21, 2015 (in press).
In Chapter V, we studied trafficking of NKA in photoreceptors by first testing the
co-localization and interaction between AnkB/AnkR and NKA in bovine photoreceptors.
We found that the ankyrin proteins do not co-localize with NKA in photoreceptors
although they interact to a certain extent in other retinal regions. The result suggests
that trafficking of NKA in photoreceptors is ankyrin-independent. We next sought to
begin addressing the question about how the different compartmentalization of NKAα
subunits is controlled. By examining the subcellular localization of NKAα3 and NKAα4
subunits expressed in Xenopus laevis photoreceptors, we found that NKAα3 localizes
properly to the ISPM when overexpressed in transgenic Xenopus photoreceptors while
NKAα4 localizes to the OS. Note that the OS is anatomically a ciliary organelle, as is
the sperm flagella where NKAα4 is normally expressed. Analysis of a series of α3/α4
chimeras and deletion mutants revealed a VxP OS/ciliary targeting motif at the N18
terminus of NKAα4. The VxP motif is necessary for localizing NKAα4 to the OS but
insufficient to counter the mechanisms restricting NKAα3 to the ISPM. This work
expands the repertoire of potential mechanisms contributing to differential subcellular
compartmentalization of NKA isozymes. This study is reported in a manuscript entitled
“Identification of a VxP targeting signal in the flagellar Na+/K+-ATPase” that is currently
under review.
All work described above will be summarized and discussed in Chapter VI. I will
discuss my model for regulating membrane protein trafficking in photoreceptors using
HCN1 and NKA as examples. I will also discuss how the newly discovered trafficking
signals function in their specific pathways. I envision several research directions in the
future including potential regulators of HCN1 trafficking in the early secretory pathways
and searching for additional trafficking signals responsible for the ISPM localization of
NKAα3. The studies conducted in my thesis and the future directions proposed will help
to elucidate the regulatory pathways controlling membrane protein trafficking in
photoreceptors as well as other systems.
19
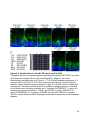
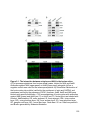
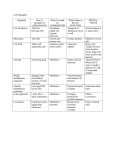
Major TRIP8b isoforms
in the brain
Effect on HCN1 surface
expression112,113
Effect on HCN1
compartmentalization114
1a-2-4
up-regulation
no effect
1a-4
up-regulation
promotes dendritic localization
1a-2
no effect
no effect
1a
up/down-regulation
inhibits axonal localization
1b-2-3-4
not known
no effect
1b-2-4
down-regulation
no effect
1b-2
down-regulation
no effect
Table 1-1. Roles of TRIP8b isoforms in regulating HCN1 trafficking
The major TRIP8b isoforms identified in the brain are listed, along with their role in
HCN1 trafficking as tested either in vivo or in vitro. The isoforms are named based on
their exon compositions (e.g. 1a-2-4 contains exons 1a, 2 and 4).
20
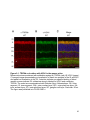
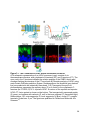
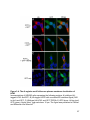
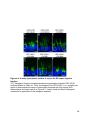
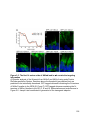
Figure 1-1. Compartmentalization and function of vertebrate photoreceptors
A cartoon of a rod photoreceptor. The outer segment (OS), inner segment (IS), nucleus
(N) and synaptic terminal (ST) are indicated on the left. Some membrane proteins
involved and the current flow during dark (A), light (B) and resetting (C) conditions are
shown. Abbreviations: CNG, cyclic nucleotide gated channel; NKA, sodium potassium
ATPase; HCN1, hyperpolarization activated cyclic nucleotide gated channel 1.
21
Figure 1-2. Major membrane protein trafficking pathways in the cell
A cartoon showing the major trafficking compartments and pathways for integral
membrane proteins. Major carriers involved in these processes are indicated.
Abbreviations: COP, coat protein; PM, plasma membrane; ERGIC, ER Golgi
intermediate compartment.
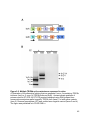
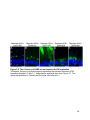
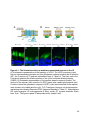
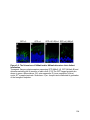
22
Figure 1-3. Structure and retinal localization of HCN1
A) A cartoon of an HCN1 tetramer (top) and an HCN1 monomer (bottom) in the lipid
bilayer. The voltage sensing region in the transmembrane domain 4 is marked with
positive signs. Proteins that interact with the post-CNBD region of HCN1 and the
interaction sites are indicated. Note that TRIP8b also interacts with the CNBD in
addition to the last three amino acids of HCN1. B) Localization of HCN1 (green) in the
mouse retina. The ONL layer contains photoreceptor nuclei and the OPL layer contains
the synaptic terminals from photoreceptors. The asterisk indicates non-specific labeling
of blood vessels. The nuclei are counterstained with Hoechst (blue). Abbreviations: NT,
N-terminus; CT, C-terminus; OS, outer segment; IS, inner segment; ONL, outer nuclear
layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer;
GC, ganglion cell layer. Scale bar: 20µm.
23
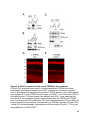
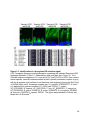
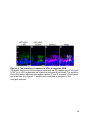
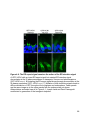
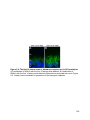
Figure 1-4. Structure and retinal localization of NKAα
A) A cartoon of the minimal functional NKA complex containing the α (red) and the β
(purple) subunit. The cytoplasmic catalytic domains, N-terminus (NT) and C-terminus
(CT) of NKAα are shown. NKAβ contains a highly glycosylated extracellular domain. B)
Staining of endogenous NKAα (red) in the mouse retina. Abbreviations as in Figure 1-3
24
CHAPTER II - ROLE OF TRIP8B IN REGULATING HCN1 TRAFFICKING
IN PHOTORECEPTORS
Abstract
Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels are cationselective channels present in retina, brain and heart. The activity of HCN channels
contributes to signal integration, cell excitability and pacemaker activity. HCN1 channels
expressed in photoreceptors participate in keeping light responses transient and are
required for normal mesopic vision. The subcellular localization of HCN1 varies among
cell types. In photoreceptors HCN1 is concentrated in the inner segments while in other
retinal neurons, HCN1 is evenly distributed though the cell. This is in contrast to
hippocampal neurons where HCN1 is concentrated in a subset of dendrites. A key
regulator of HCN1 trafficking and activity is tetratricopeptide repeat-containing Rab8b
interacting protein (TRIP8b). Multiple splice isoforms of TRIP8b are expressed
throughout the brain and can differentially regulate the surface expression and activity
of HCN1. The purpose of the present study was to determine which isoforms of TRIP8b
are expressed in the retina and to test if loss of TRIP8b alters HCN1 expression or
trafficking. We found that TRIP8b co-localizes with HCN1 in multiple retina neurons and
the retina express all major TRIP8b splice isoforms, three of which are found in
photoreceptors. In TRIP8b knockout mice, the ability of HCN1 to traffic to the surface of
specific compartments in retinal neurons is unaffected. However, there is a large
decrease in the total amount of HCN1. We conclude that TRIP8b in the retina is needed
to achieve maximal expression of HCN1.
25
Introduction
The hyperpolarization activated current (Ih) was discovered in photoreceptors
where absorption of light triggers a signal transduction cascade leading to closure of the
CNG channels and causing the cell to hyperpolarize. The subsequent influx of Ih helps
to rapidly reset the membrane potential3-5,115-117. Ih was later found to be carried out by
the HCN channels that are expressed throughout the nervous system30,67. All members
of the HCN family were found in the retina although with distinct expression profiles32,33.
Rod photoreceptors express HCN1 which is concentrated in the inner segments (IS),
excluded from the outer segment (OS) and to a lesser degree in the plasma membrane
surrounding the nuclei and synaptic terminals5,33-35. Cones express HCN1 in a similar
pattern but also contain HCN3 at the synapse33. HCN1 is also expressed in multiple
inner retina neurons and is found in multiple cellular compartments (dendrites, soma,
axons and presynaptic terminals)5,33-42.
In other neuronal populations HCN1 can be distributed throughout the cell or
restricted to particular subcellular domains. For example, HCN1 is found in stereocilia
and afferent dendrites of cochlear hair cells61,62 and in the soma and nodes of Ranvier
in dorsal root ganglia63. But in hippocampal CA1 and layer V neocortical pyramidal
neurons HCN1 is concentrated in distal dendrites64. Since changes in HCN1 abundance
or subcellular localization are associated with learning and memory, epilepsy and
pain67,118, it is important to understand how the trafficking of HCN1 is regulated in
various cell types. The best understood regulator of HCN1 subcellular localization is
TRIP8b.
26
TRIP8b is a cytoplasmic protein that binds to the C-terminus of HCN channel
subunits via two contact sites64,113,119. TRIP8b interactions with the cyclic nucleotide
binding domain of HCN allows it to modulate the gating and surface expression of HCN
channels, while TRIP8b’s interaction with the last three amino acids of HCN C-terminus
is important for proper trafficking of the channel114,119,120. Multiple splice isoforms of
TRIP8b are expressed in the brain and can have opposing influences on the localization
of HCN1 (Table 1-1)67,114. In the hippocampus, TRIP8b is essential for maintaining the
expression level, surface availability and the concentration of HCN1 channels in distal
dendrites of pyramidal neurons67. Furthermore, TRIP8b can inhibit the axonal
distribution of HCN1 in the medial perforant path65. However, trafficking of HCN1 to
presynaptic terminals in the cortex is independent of TRIP8b66 so the role of TRIP8b
can vary depending on neuronal population and subcellular localization. It is not known
if TRIP8b regulates the trafficking of HCN1 channels in retinal neurons.
In this study we found that all major splice isoforms of TRIP8b are expressed in
the retina and TRIP8b co-localizes with HCN1 in photoreceptors and inner retina
neurons. In the absence of HCN1, TRIP8b is expressed at normal levels in the retina,
but it is not fully recruited to membranes. Conversely, in the absence of all TRIP8b
isoforms, HCN1 levels are reduced by 40% of normal. Despite this, the trafficking of
HCN1 to the surface of retinal neurons is maintained and visual function as measured
with ERG is normal in TRIP8b knockout animals. We conclude that trafficking of HCN1
channels in the retina can take place independent of TRIP8b but this accessory subunit
is necessary to maintain maximal expression of the channel.
27
Materials and Methods
Animals
C57BL/6J pigmented wildtype mice were purchased from the Jackson Laboratory
as were the HCN1-/- mice originally described by Nolan and colleagues121. The TRIP8b-/and TRIP8b 1b/2-/- lines were maintained as previously described65,67,114. All three
knockout lines are congenic on the C57BL/6 strain background. Mice were maintained
on a standard 12/12 hour light/dark cycle; food and water were provided ad libitum. This
study was carried out in strict accordance with the recommendations in the Guide for
the Care and Use of Laboratory Animals of the National Institutes of Health. The
experiments were approved by the Institutional Animal Care and Use Committee at the
University of Iowa, adhered to the ARVO guidelines for animal use in vision research.
Prior to tissue collection, mice were humanely euthanized by CO2 asphyxiation followed
by cervical dislocation and all efforts were made to minimize suffering.
Immunohistochemistry
Individual mouse eyes were enucleated and eyecups were prepared by removing
the cornea, iris and lens. Eyecups were fixed in 4% paraformaldehyde prepared in PBS
at room temperature for 15min, cryoprotected in 30% sucrose at 4°C O/N then frozen in
O.C.T (Tissue-Tek) before collecting 10 µm cryosections. For immunostaining, the
sections were permeablized in PBS containing 0.5% Triton X-100 for 10min, followed by
incubation in 10% goat serum to block nonspecific labeling. Eyecup sections were
incubated with primary antibodies at 4°C O/N, washed and incubated with secondary
antibodies conjugated to either Alexa-488 or Alexa-568 and Hoechst 33342 (Life
Technologies) to label nuclei. The primary antibodies used were: rabbit α-HCN1 raised
28
against a C-terminal epitope, NTNLTKEVRPLSAS, (developed at GenScript), mouse αHCN1 (NeuroMab, 1:500), rabbit α-HCN1 (Alomone, 1:500), rabbit α-TRIP8b-constant
(1:1000)122, mouse α-TRIP8b-exon 4 (NeuroMab, 1:1000). HCN1-/- or TRIP8b-/- retina
were stained in parallel with wildtype retina sections to ensure specificity of antibody
labeling. Images were collected using a Zeiss 710 confocal microscope (Central
Microscopy Research Facility, University of Iowa). Manipulation of images was limited to
adjusting the brightness and contrast levels using Zen Light 2009 (Carl Zeiss) or
Photoshop (Adobe). Experiments were replicated with a minimum of 3 individual mice.
Laser capture micro-dissection
Mouse retinas were dissected as for immunohistochemistry, immediately frozen
in O.C.T., sectioned and slides were stored at -80°C. All equipment was thoroughly
cleaned with 70% ethanol followed by RNaseZap (Ambion) to minimize RNA
degradation. Photoreceptor inner segments were collected with a 7.5 µm diameter laser
beam on an Arcturus PIXCELL II Laser Capture Microscope (Central Microscopy
Research Facility, University of Iowa). The selected regions were then absorbed by the
Arcturus CapSure Macro LCM Caps (Applied Biosystems) and immediately processed
for RNA extraction as described below. The experiments were replicated with a
minimum of 4 individual mice.
RT-PCR
RNA from one mouse retina was extracted with the RNeasy Mini kit (Qiagen), MMLV reverse transcriptase (Life Technologies) with either random hexamers or genespecific primers was used for cDNA synthesis, followed by standard PCR reactions to
29
amplify specific products. After separation on agarose gels, all amplification fragments
were extracted and verified by sequencing (University of Iowa DNA facility). The
experiment was replicated 3 times. Primers used for RT-PCR: TRIP8b-exon 1a-forward,
5’gagcagaatgtaccagggacacat; TRIP8b-exon 1b-forward, 5’ggaaggactcacattccatctctac;
TRIP8b-exon 5-reverse, 5’tggatgtcactggctttgcaatggc.
Fractionation of cytosolic and membrane proteins
Freshly isolated mouse retinas were homogenized in hypotonic buffer (50 mM
Tris-HCl, 10 mM NaCl, 0.32 M sucrose, 5 mM EDTA, 2.5 mM EGTA, pH7.4)
supplemented with protease inhibitor cocktail (Complete mini, Roche), followed by
centrifugation at 1,000 x g for 10min at 4°C to pellet unbroken cells and nuclei. The
supernatant was centrifuged at 240,000 x g for 30min at 4°C. The supernatant,
containing cytosolic proteins, was collected. The pellet, containing membrane and
cytoskeletal proteins, was resuspended in hypotonic buffer plus detergent (1.5% Triton
X-100 and 0.75% DOC). After centrifugation at 240,000 x g for 30min at 4°C, the
supernatant, containing solubilized membrane proteins, was collected. The experiment
was replicated 3 times.
Immunoprecipitation
Twenty retinas from either wildtype or HCN1-/- mice were homogenized in 20 mM
HEPES, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, and 1 mM PMSF containing
complete protease inhibitor cocktail. The homogenate was centrifuged at 800 x g for 10
min at 4oC to remove nuclei and the post-nuclear supernatant was centrifuged at
100,000 x g for 1 h at 4oC to pellet the retinal membranes. The membrane fraction was
solubilized in homogenization buffer supplemented with 1% DDM (n-Dodecyl-beta-D30
Maltoside) for 1 h at 4oC on a rotator and subsequently centrifuged at 100,000 x g for 1
h at 4oC to remove residual unsolubilized material. 250 µg of solubilized membrane
extract was incubated with rabbit α-HCN1 antibody for 2 h at 4oC on a rotator. After
addition of 1.5 mg Dynabeads Protein G (Life Technologies), incubation was continued
for an additional hour. Dynabeads were washed three times in homogenization buffer
supplemented with 1% DDM to remove unbound protein, transferred to a new tube, and
bound protein was eluted with NuPAGE LDS sample buffer (Life Technologies) lacking
reducing agent. The experiment was replicated 3 times.
Biotinylation assay
Freshly isolated mouse retina (two for each experiment) was incubated with 1
mg/mL sulfo-NHS-SS-biotin (Thermo Scientific) for 10min at 4°C to label only the
surface proteins. The reaction was quenched (100 mM glycine, 25 mM Tris-HCl, pH7.4)
for 15min, and then the solution was washed in PBS three times; all procedures were
carried out at 4°C. The biotinylated retinas were homogenized in lysis buffer (50 mM
Tris-HCl, 10 mM NaCl, 0.32 M sucrose, 5 mM EDTA, 2.5 mM EGTA, 1.5% Triton X-100,
0.75% DOC, 0.1% SDS, pH7.4) supplemented with protease inhibitor cocktail
(Complete, mini, Roche). Insoluble material was removed by centrifugation at 10,000 x
g for 10min and the supernatant was incubated with 100 ul NeutrAvidin agarose resin
(Thermo Scientific) with gentle mixing at 4°C O/N. Beads were washed with lysis buffer
three times, and proteins bound to the beads were eluted with a reducing NuPage LDS
sample buffer (Life Technologies). The experiment was replicated with a minimum of 3
individual mice of each genotype.
31
Western blotting
Protein content in samples was measured using the BCA assay (Thermo
Scientific). Proteins were fractionated on 10% Mini-PROTEAN TGX gels and transferred
to PVDF membranes (Bio-Rad). Membranes were blocked in 5% milk and incubated
with the following primary antibodies: rabbit α-HCN1 (3µg/mL), rabbit α-TRIP8b
(1:10,000), mouse α-NKA (M7-PB-E9, Santa Cruz, 1:1000), rabbit α-PDC (gift from Dr.
Maxim Sokolov, 1:3000) and secondary antibodies conjugated to HRP. Blots were
incubated with SuperSignal West Femto Maximum Sensitivity Substrates (Thermo
Scientific) and visualized with a CCD camera (ChemiDoc XR+, Bio-Rad). The software
package Image Studio v3.1 (LI-COR Biosciences) was used for analysis of the images.
Electroretinography
Full field ERG was obtained using the Espion V5 Diagnosys system (Diagnosys
LLC, MA, USA). After overnight dark-adaptation, mice were anesthetized with an
intraperitoneal injection of ketamine (87.5 mg/kg) and xylazine (2.5 mg/kg). Mice were
5-16 weeks of age, and both HCN1-/- and TRIP8b-/- animals were compared to wildtype
littermates obtained from breeding heterozygotes. ERGs were recorded simultaneously
from the corneal surface of each eye after pupil dilation (1% tropicamide) using gold ring
electrodes (Diagnosys) referenced to a needle electrode (Roland/LKC) placed on the
back of the head. Another needle electrode placed in the tail served as the ground. A
drop of methylcellulose (2.5%) was placed on the corneal surface to ensure electrical
contact and to maintain corneal integrity. Body temperature was maintained at a
constant temperature of 38°C using a regulated heating pad. All stimuli were presented
in a ColorDome (Diagnosys) ganzfeld bowl and the mouse head and electrode positions
32
were monitored on the camera attached to the system. Dim red light was used for room
illumination until dark adapted testing was completed. A modified ISCEV protocol 123,124
was used including a dark adapted dim flash of 0.01 cd.s/m2, maximal combined
response (standard combined response or SCR) to bright flash of 3 cd.s/m2, light
adapted bright flash of 3 cd.s/m2,and 5 Hz flicker stimuli at 3 cd.s/m2. An escalating
intensity protocol was utilized with intensities starting at 0.0001 cd.s/m2 and increasing
incrementally to 31.6 cd.s/m2. A dark-adapted increasing flicker frequency protocol was
utilized in which the intensity of flash was held constant at 3.1 cd.s/m2 and the flash
frequency increased from 0.5 Hz incrementally to 30 Hz. The a-wave was measured
from the baseline to the trough of the first negative wave. The b-wave was measured
from the trough of the a-wave to the peak of the first positive wave, or from the baseline
to the peak of the first positive wave if no a-wave was present.
Results
Multiple TRIP8b isoforms are present in the retina
TRIP8b influences HCN1 trafficking and/or activity in various neurons125, but its
expression in the retina had not been investigated. Immunohistochemistry of the mouse
retina revealed that TRIP8b is expressed in multiple regions throughout the retina;
including photoreceptors, bipolar cells and ganglion cells (Figure 2-1A). In
photoreceptors, TRIP8b is present in the IS and synaptic terminals (upper strata of the
outer plexiform layer, OPL) but excluded from the OS – the same distribution as HCN1
(Figure 2-1B, C). In the inner plexiform layer (IPL) that contains synapses between
bipolar and ganglion cells, TRIP8b labeling could be distinguished in all sublamina,
including the sublamina that is prominently stained by anti-HCN1 antibodies due to the
33
expression of HCN1 in type 5 bipolar axon terminals5,33,35. This demonstrates that
TRIP8b is expressed in cells containing HCN1, but not exclusively. This is reasonable
given that other HCN channels (e.g. HCN2 and HCN4) are also expressed in various
sublamina of the IPL and can interact with TRIP8b35,33.
TRIP8b is subject to alternative promoter usage and alternative splicing,
generating 9 unique variants which can differentially regulate the trafficking of
HCN1112,113. To determine which TRIP8b splice isoforms are expressed in the retina, we
performed RT-PCR using RNA isolated from mouse retina and primer pairs designed to
distinguish between the isoforms (Figure 2-2A). We found seven products in the retina
(Figure 2-2B), corresponding to the major isoforms expressed throughout the brain.
Each product was sequenced to verify that it was the expected TRIP8b splice variant.
The retina is a multilayered tissue composed of five principle types of neurons
and several types of glial cells. To determine which of the various TRIP8b isoforms are
expressed in photoreceptors, we used laser capture micro-dissection of the mouse
retina to collect the IS, the layer where the majority of photoreceptor transcripts are
translated. The only other cell type possibly collected in this procedure would be a very
small fraction of the Muller glial cells that span the retina from the ganglion cell layer to
the outer limiting membrane between the IS and ONL layers. Imaging of the retina
before and after micro-dissection was used to confirm the efficiency of IS isolation
(Figure 2-3A, B). RT-PCR performed using RNA isolated from the IS revealed three
isoforms: TRIP8b 1a (IsoA5), TRIP8b 1a-4 (IsoA4) and TRIP8b 1a-2-4 (IsoA2) (Figure
2-3C). This observation is consistent with the abundance of these transcripts in other
neuronal populations, where TRIP8b 1a and 1a-4 together comprise ~60% of all
34
TRIP8b isoforms expressed in the brain with 1b containing isoforms concentrated in
glial cells112,114.
The expression of exon 4 containing isoforms in photoreceptors can be
confirmed by immunohistochemistry using an anti-TRIP8b antibody specific to an
epitope in this exon. This antibody should label photoreceptors in wildtype retina
(isoforms 1a-4 and 1a-2-4) and in TRIP8b 1b/2-/- (a strain that does not express TRIP8b
isoforms containing exon 1b or exon 2) retina (isoform 1a-4). As predicted, labeling of IS
and the OPL is similar to that seen with the anti-TRIP8b antibody that detects all
isoforms (Figure 2-3D, E, compare to Figure 2-1A).
Interaction of HCN1 and TRIP8b in the retina
HCN1 and TRIP8b interact in the retina as confirmed by immunoprecipitation
from retinal membranes (Figure 2-4A). Membranes prepared from the retinas of HCN1-/mice were used for the negative control and we noted a reduction of TRIP8b in this
preparation. When total retina extracts were probed with anti-TRIP8b antibodies
(constant or exon-4 specific), the expression level of TRIP8b was found to be the same
as that in wildtype retinas (Figure 2-4B). Due to the similarity in molecular weight, the
various TRIP8b isoforms are detected as just 2 or 3 bands on Western blots. To further
explore the difference in TRIP8b levels observed in Figures 2-4A and 2-4B, we
compared cytosolic and membrane fractions of the retina side by side; the efficiency of
this separation was monitored by probing for a soluble protein, phosducin (PDC), and
the membrane bound sodium potassium ATPase (NKA). There was a reduction in the
amount of TRIP8b in the isolated membrane fraction and a corresponding increase in
the cytosolic fraction from the HCN1-/- mouse compared to the wildtype (Figure 2-4C).
35
This demonstrates that HCN1 is needed to fully recruit TRIP8b to retinal membranes.
The remaining pool of membrane-associated TRIP8b likely reflects the recruitment of
TRIP8b by HCN2, 3, or 4. Note, as expected because the size of the photoreceptors
does not provide the resolution needed to distinguish between plasma membrane and
cytoplasm, there is no change in the pattern of TRIP8b immunostaining in the absence
of HCN1 (Figure 2-4D, E).
Reciprocally, TRIP8b can control the subcellular localization of HCN1,
particularly in neurons where HCN1 is restricted to a sub-domain of dendrites64.
However, HCN1 does not display this restricted expression pattern in the retina, so we
tested if there is any alteration in HCN1 due to the loss of TRIP8b. In the TRIP8b 1b/2-/retina which lacks 5 of the 7 TRIP8b isoforms65,67,114, but still expresses 2 of the 3
isoforms present in photoreceptors (1a and 1a-4), immunolabeling with anti-HCN1
antibodies revealed no change in the compartmentalization of HCN1 (compare Figure
2-5A and B). In the total TRIP8b-/- retina, HCN1 labeling was reduced in signal intensity
but present in the same compartments as in wildtype retinas (compare Figure 2-5A and
C). The signal intensity of HCN1 in the IPL was least affected by the loss of all TRIP8b
isoforms. Western blotting demonstrated that the levels of total HCN1 protein was not
changed in the TRIP8b 1b/2-/- retina but reduced by ~40% in TRIP8b-/- retinas (Figure 25D). Interestingly, the reduction in HCN1 levels due to either haploinsufficiency of HCN1
or loss of TRIP8b is indistinguishable (Figure 2-5E).
To determine if the reduced levels of HCN1 in the retinas of TRIP8b-/- animals
compromise retina function we recorded electroretinograms (ERG). One of the major
phenotypes in ERG responses from HCN1-/- animals is the failure to resolve high
36
frequency flickering light stimuli; this suppression of cone responses is hypothesized to
be driven by abnormally prolonged signaling in rods 4,5,36,126. Similar defects in ERG
responses are expected in TRIP8b-/- mice if the TRIP8b-dependent 40% decrease in
HCN1 affects vision. Responses to a single flash under dark or light adapted conditions
were similar between TRIP8b-/- and wildtype animals (data not shown). In response to a
dark-adapted flicker frequency series, HCN1-/- mice failed to respond to cone-isolating
frequencies as expected, but responses from TRIP8b-/- animals were not significantly
different from that of wildtype (p > 0.2 by ANOVA; Figure 2-6). This demonstrates that
the decreased HCN1 levels in TRIP8b-/- animals are sufficient for ERG responses.
HCN1 surface expression in the absence of TRIP8b
TRIP8b can regulate the trafficking of HCN1 to the plasma membrane. To
investigate HCN1 surface levels in retinal neurons we used an ex vivo biotinylation
assay. Freshly isolated retinas were incubated with a non-membrane permeable biotin
reagent to cross-link surface proteins to biotin. Labeling sectioned retinas with
fluorescently conjugated streptavidin verified that the biotinylation reagent penetrated all
regions of the retina (Figure 2-7A). After labeling with biotin, surface proteins were
isolated using NeutrAvidin-agarose beads and analyzed by Western blotting. The
labeling of NKA was monitored as a positive labeling control and PDC as a negative
labeling control to confirm that only membrane proteins were biotinylated in the assay.
HCN1 was readily detected on the surface of retina neurons using this approach and we
observed a decreased amount of biotinylated HCN1 in the TRIP8b-/- retinas (Figure 27B). To take into account the different abundance of HCN1 in these two mice we
compared the ratio of surface to total HCN1 and found that it was unchanged in the
37
absence of TRIP8b. This ratio is similarly preserved in HCN+/- animals which have a
TRIP8b independent reduction in HCN1 (Figure 2-7C).
Discussion
In this study we characterized the expression of TRIP8b in the retina. TRIP8b is
an accessory subunit of HCN channels and can modulate the function and location of
HCN channels in various neuronal populations. We found that TRIP8b is expressed in
multiple neurons of the retina and co-localizes with HCN1 in photoreceptors and in the
inner retina. The interaction between these two proteins was confirmed by coimmunoprecipitation and supported by the dependence on HCN1 for membrane
recruitment of TRIP8b. TRIP8b undergoes alternative splicing, generating distinct
isoforms that can regulate HCN channels in opposing ways (Table 1-1). All the major
splice isoforms are expressed in the retina and three different forms were found in
photoreceptors alone (1a, 1a-4, 1a-2-4). TRIP8b 1a was reported to inhibit the axonal
localization of HCN1, and may up- or down-regulate HCN1 surface expression
depending on the cell type in which it is expressed. Isoforms 1a-4 has also been shown
to regulate HCN1 compartmentalization114. Both 1a4 and 1a-2-4 can up-regulate HCN1
surface expression65,112,114.
This observation led us to test three predictions arising from the hypothesis that
TRIP8b regulates trafficking of HCN1 – HCN1 expression levels, compartmentalization,
or surface expression should be altered with the loss of TRIP8b. We used two different
lines of TRIP8b knockout mice, TRIP8b 1b/2-/- which lacks just the minor splice isoforms
(retaining expression of the more abundant 1a and 1a-4 isoforms), and TRIP8b-/- lacking
all isoforms. We did not observe any changes in HCN1 as long as the 1a and 1a-4
38
isoforms were present. In the TRIP8b-/- retina, the most striking observation was a 40%
down regulation of HCN1 protein levels. Similar TRIP8b dependent decreases in HCN1
have been observed in other neuronal populations and this is likely due to excessive
routing of HCN1 to lysosomes in the absence of TRIP8b67. The staining intensity of
HCN1 in the inner retina was similar in wildtype and TRIP8b-/- mice but notably
decreased in the photoreceptor layers. This indicates that TRIP8b has the largest
influence on HCN1 expression in photoreceptors.
In photoreceptors, HCN1 is most abundant in the IS and found to a lesser degree
in the synaptic terminals. Since the TRIP8b 1a isoform can inhibit axonal localization in
both hippocampal CA1 pyramidal and medial perforant path neurons65,114, we
considered that it might also limit the amount of HCN1 that accumulates in the
photoreceptor synaptic terminal. This is not the case as the compartmentalization of
HCN1 was unaltered with the loss of all TRIP8b isoforms. We also tested if trafficking to
the plasma membrane of retinal neurons was altered in the absence of TRIP8b using
surface biotinylation assays. Interestingly, the ratio of surface to total HCN1 in retinal
neurons was maintained even with the reduction of available HCN1. This indicates that
the trafficking of HCN1 in retina neurons occurs independently of TRIP8b.
Our findings differ from the situation in hippocampal CA1 and neocortical layer V
pyramidal neurons in which TRIP8b is required for the trafficking of HCN1 in distal
dendrites114 . This difference is not completely unexpected as in the retina HCN1 is
most abundantly expressed in photoreceptors which lack canonical dendrites. The OS
is the subcellular compartment functionally analogous to a dendrite and HCN1 is
excluded from this compartment. Interestingly, a recent study has demonstrated that
39
TRIP8b is not needed for the pre-synaptic localization of HCN1 in entorhinal cortex66.
Our results further extend the concept that the trafficking of HCN1 outside of dendrites
can be maintained independently of TRIP8b.
In the visual system, HCN1 is activated by light induced hyperpolarization of
photoreceptors and carries an inward current that is thought to be required to prevent
prolonged rod signaling. Consequently patients taking ivabradine, an HCN channel
blocker used to control heart rate, occasionally report the occurrence of phosphenes, a
condition of seeing a ring or spot of light, as a side effect127,128. Despite the striking
decease in the amount of HCN1 in the absence of TRIP8b, we did not see a loss of
retinal function as measured with ERGs. While the ERG provides a rapid assessment of
the summed output of the retina, this approach lacks sensitivity. An in depth
electrophysiological study of individual TRIP8b-/- retina neurons would be helpful to
determine how this HCN accessory subunit influences the biophysical properties of
retinal HCNs and contributes to visual signaling.
HCN channels play multiple roles in the nervous and cardiac systems and their
dysregulation can lead to epilepsy, chronic pain, and heart disease30,31,118.
Pharmacological manipulation of HCN expression and function therefore holds promise
for the management of numerous diseases. But to effectively target specific diseases it
is important to continue dissecting the modes of regulation that ensure differential
regulation of HCN trafficking and function in various subcellular compartments, cell, and
tissues.
40
Figure 2-1. TRIP8b co-localizes with HCN1 in the mouse retina
Mouse retina immunostained with antibodies against A) TRIP8b (red), B) HCN1 (green),
C) merged image demonstrating co-localization of these two proteins in the IS and OPL
and partial co-localization in the IPL. Asterisks indicate non-specific labeling of blood
vessels; arrows indicate IPL sublamina strongly labeled for HCN1 and containing
TRIP8b. The nuclei are counterstained with Hoechst (blue). Abbreviations: OS, outer
segment; IS, inner segment; ONL, outer nuclear layer; OPL, outer plexiform layer; INL,
inner nuclear layer; IPL, inner plexiform layer; GC, ganglion cell layer. Scale bar: 20µm.
The figure was published in in PLOS ONE110.
41
Figure 2-2. Multiple TRIP8b splice variants are expressed in retina
A) Illustration of the alternative splicing that can generate 6, exon 1a containing TRIP8b
isoforms (IsoA) or 3, exon 1b TRIP8b isoforms (IsoB). Arrows indicate positions of
primers used for RT-PCR. B) RT-PCR products obtained using RNA isolated from
mouse retina and primer pairs to amplify TRIP8b IsoA (lane 1) or IsoB splice variants
(lane 2). Reverse transcriptase (RT) was omitted as a negative control (lanes 3 and 4).
The figure was published in in PLOS ONE110.
42
Figure 2-3. Multiple TRIP8b splice variants are expressed in photoreceptors
A) Mouse retina section before laser capture micro-dissection, dotted white lines
indicate region subsequently collected and shown in (B). C) RT-PCR products obtained
using RNA isolated from tissue collected as in (B) with the isoform specific primers
depicted in Figure 2-2A. Reverse transcriptase was omitted as a negative control.
Immunostaining with an antibody recognizing exon 4 in TRIP8b labels photoreceptors of
wildtype (D), TRIP8b 1b/2-/- (E), but not TRIP8b-/- (F) mouse retina. Asterisks indicate
non-specific labeling of blood vessels. Abbreviations and scale bars as Figure 2-1. The
figure was published in in PLOS ONE110.
43
Figure 2-4. HCN1 is required to fully recruit TRIP8b to the membrane
A) Anti-HCN1 antibodies were used to co-immunoprecipitate TRIP8b from retinal
membranes. Membranes prepared from HCN1-/- retinas were used as the negative
control. B) Western blot comparing the amount of TRIP8b present in total retina lysates
from wildtype (WT), both TRIP8b knockout lines, and HCN1-/- mice. Phosducin (PDC) is
the loading control. C) Retina lysates from wildtype and HCN1-/- mice separated into
cytosolic (sol) and membrane fractions (mem) probed with anti-TRIP8b, anti-TRIP8b
exon 4 (ex4) and anti-HCN1 antibodies. PDC and sodium/potassium ATPase (NKA) are
loading controls for each fraction. Immunostaining of TRIP8b in wildtype (D) and HCN1/- retina (E) is indistinguishable. Abbreviations and scale bars as Figure 2-1. The figure
was published in in PLOS ONE110.
44
Figure 2-5. HCN1 protein levels are reduced in the absence of TRIP8b
Anti-HCN1 antibodies were used to immunostain retina from wildtype (A), TRIP8b 1b/2-/(B), or TRIP8b-/- (C) mice. Abbreviations and scale bars as Figure 2-1. D) Western blots
of total retina lysates probed with anti-HCN1, with phosducin (PDC) used as a loading
control. E) Relative amount of HCN1 expressed in the retina of wildtype (WT), HCN1
heterozygous (HCN1+/-), and TRIP8b-/- mice as quantified by densitometry. The figure
was published in in PLOS ONE110.
45
Figure 2-6. Flicker frequency responses are normal in the absence of TRIP8b
A) Representative waveforms from flicker ERGs of wildtype (WT), TRIP8b-/-, and HCN1/- (n=3 mice for each genotype). Dark-adapted mice were stimulated with a flash
intensity of 3.1 cd.s/m2 at frequencies of 0.5, 1, 2, 3, 5, 7, 10, 12, 15, 18, 20, and 30 Hx.
B) ERG amplitudes plotted as a function of flicker frequency. The figure was published
in in PLOS ONE110.
46
Figure 2-7. The surface expression of HCN1 is maintained in TRIP8b-/- mice
A) Streptavidin staining of control (left) and biotinylated retina (right). B) After
biotinylation, surface proteins from either control wildtype (WT) or TRIP8b-/- mice were
pulled down using NeutrAvidin beads. The level of HCN1 in the total (input) and surface
(eluted from NeutrAvidin beads) pools was detected by Western blotting. Phosducin
(PDC) and sodium potassium ATPase (NKA) were used as negative and positive
controls, respectively. C) Densitometry of Western blots represented in (B) were used to
calculate the surface to total ratio of HCN1 after normalization to the loading control
NKA. Abbreviations and scale bars as Figure 2-1. The figure was published in in PLOS
ONE110.
47
CHAPTER III - IDENTIFICATION OF A DI-ARGININE ER RETENTION
SIGNAL IN THE C-TERMINUS OF HCN1
Abstract
Hyperpolarization-activated cyclic nucleotide-gated 1 (HCN1) channels carry the
hyperpolarization-activated current (Ih) which contributes to neuronal excitability and
signal transmission in the nervous system. Controlling the trafficking of HCN1 is an
important aspect of its regulation, yet the details of this process are poorly understood.
Here we investigated how the C-terminus of HCN1 regulates trafficking by testing for its
ability to redirect the localization of a non-targeted reporter in transgenic X. laevis
photoreceptors. We found that HCN1 contains an ER localization signal and through a
series of deletion constructs, identified the responsible di-arginine ER retention signal.
This signal is located in the intrinsically disordered region of the C-terminus of HCN1.
To test the function of the ER retention signal in intact channels we expressed wildtype
and mutant HCN1 in HEK293 cells and found this signal negatively regulates surface
expression of HCN1. In summary, we report a new mode of regulating HCN1 trafficking
through the use of a di-arginine ER retention signal that monitors processing of the
channel in the early secretory pathway.
Introduction
HCN channels are expressed in the brain and heart where they modulate signal
integration, synaptic transmission, or rhythmic changes in membrane excitability30. Not
surprisingly, disruption of their activity influences behavior and is pathological. For
48
instance, alterations in HCN2 and HCN4 lead to cardiac arrhythmia129. HCN1 is more
broadly expressed and influences behavior in diverse ways; it contributes to vision at
medium and bright light intensities3-5, memory formation121, and sleep130. Perturbation of
HCN1 underlies some forms of epilepsy and chronic pain67,118,131. The ongoing efforts to
elucidate the mechanisms of HCN channel regulation are critical to making progress in
understanding their roles in disease progression.
HCN channels are regulated by several convergent mechanisms. HCN open
relatively slowly in response to membrane hyperpolarization and their gating is
modulated by factors including cAMP, phosphatidylinositol 4,5-bisphosphate (PIP2), and
the accessory subunit, TRIP8b30. Controlling subcellular trafficking is another important
aspect of channel regulation but is not well understood. This is in part because there are
likely overlapping mechanisms to account for the range of HCN localization found
across cells. HCN1 can be found broadly distributed throughout the plasma membrane
of a neuron or be restricted to a particular compartment (for details, see introduction in
Chapter II). In photoreceptors, HCN1 is most abundant in inner segments (IS) and
excluded from outer segments (OS)5,33-35. TRIP8b is the major regulator of HCN1
trafficking as it is essential for reducing axonal, but enhancing, dendritic localization of
the channel65,67,114,119,120. However, our study (Chapter II) together with others showed
that TRIP8b is not required to localize HCN1 in retinal neurons66,110. Determining how
the localization of HCN1 in various cells and compartments is controlled is a current
challenge for the field.
The cytoplasmic C-terminus (CT) of HCN1 and related channels is important for
trafficking. Their C-termini contain two conserved structured regions, the C-linker which
49
contributes to tetramerization and the cyclic nucleotide binding domain (CNBD) which
allows for modulation by cAMP, followed by a non-conserved post-CNBD region57. The
CT of HCN1 contains multiple binding sites to proteins that can affect HCN1 trafficking
(Figure 1-3A). The CT of HCN2 binds KCNE2, a promiscuous potassium channel
accessory subunit that increases surface expression of some binding partners132-135.
HCN3 surface expression is enhanced by its own accessory subunit, K+ channel
tetramerization domain-containing protein 3 (KCTD3), which interacts with the portion of
the CT containing the C-linker and CNBD domains136. In a more distantly related
Shaker-like K+ channel, Arabidopsis thaliana 2, specific residues in the C-linker are
necessary for promoting surface expression of the channel137. It is not known how these
factors influence HCN1 trafficking in different types of specialized cells such as
photoreceptors.
Our purpose in this study is to identify additional regulatory elements controlling
the trafficking of HCN1. We used X. laevis photoreceptors, a powerful model system for
dissecting neuronal and ciliary trafficking pathways138. Fusing the CT of HCN1 to a
reporter that alone accumulates in the OS resulted in redirecting the reporter to the ER
within the IS. By analyzing the localization of a series of mutants of this reporter-HCN1CT construct we uncovered a di-arginine ER retention motif. Di-arginine motifs have
been found in a select subset of channels and receptors and have been proposed to
function as signals for improperly assembled channels. This predicts that disruption of
the di-arginine motif in otherwise intact HCN1 channels would increase the amount of
channel that exits the ER and localizes to the cell surface. Immunocytochemistry and
biotinylation assays of HCN1 channels expressed in HEK293 cells verified that mutation
50
of the di-arginine motif caused an increase in surface expression. In conclusion, a diarginine ER retention signal that influences the trafficking of HCN1 from the ER to the
plasma membrane (PM) has been discovered. We propose that this signal works in
combination with as yet to be determined forward trafficking signals to ensure the
proper delivery of functional HCN1 channels to the PM of neurons.
Materials and Methods
Animals
Xenopus laevis were purchased from Nasco (Fort Atkinson) and maintained by
the Office of Animal Research at the University of Iowa. All experiments were approved
by the Institutional Animal Care and Use Committee and adhered to the ARVO
guidelines for animal use in vision research. Transgenic Xenopus tadpoles were
generated using restriction enzyme mediated integration as previously described139,140.
Briefly, linearized and purified plasmid DNA was integrated into sperm nuclei in the
presence of a Ca2+ activated egg cytoplasmic extract. Treated nuclei were transplanted
into unfertilized eggs obtained by inducing mature animals with human chorionic
gonadotropin (Prospec). Embryos were housed in 0.3 x Marc’s Modified Ringer (30 mM
NaCl, 0.6 mM KCL, 0.3 mM MgCl2, 0.6 mM CaCl2 and 1.5 mM HEPES, pH
7.4).Transgenic tadpoles were identified at St 42 by screening for GFP expression in
the eye and humanely euthanized between St 45 and 55 by immersion in 0.2% tricaine
(Sigma-Aldrich), prior to processing for immunohistochemistry.
51
Molecular cloning
All constructs used for transgenesis were subcloned into the XOP5.5 vector
containing the Xenopus opsin promoter to ensure rod photoreceptor specific
expression141. All inserts were generated by standard PCR protocols and verified by
Sanger sequencing (Iowa Institute of Human Genetics). The membrane reporter
consists of the transmembrane domain from mouse activin receptor type 2A (aa 117165), followed by EGFP, followed by a palmitoylated peptide from Xenopus rhodopsin
(aa 311-349). The template for the X. tropicalis HCN1 inserts was obtained from retina
cDNA; amino acid numbers thereby correspond to accession #XP002933077. All
constructs used for transfection of HEK293 cells were subcloned into the pEGFPN1
vector containing the CMV promoter. The mouse HCN1 insert was a generous gift from
Dr. Yoav Noam113,142, the GFP-TRIP8b (1a-4) insert was a generous gift from Dr. Dane
Chetkovich113.
Immunostaining
Transgenic tadpoles were fixed in 4% paraformaldehyde (Electron Microscopy
Sciences), cryoprotected in 30% sucrose, and frozen in Tissue-Tek O.C.T (Electron
Microscopy Sciences). Sections collected on charged glass slides were permeablilized
in 0.5% Triton X-100, blocked with 5% goat serum and 0.5% Triton X-100 in PBS,
incubated in mouse α−GFP antibodies (diluted 1:500; Clontech Laboratories) or rabbit
α-calnexin antibodies (diluted 1:100; Enzo Life Sciences), followed by secondary goat
α-rabbit or goat α-mouse antibodies conjugated to Alexa 488 or 568 (Life
Technologies,) mixed with 2 μg/mL Hoechst 33342 (Life Technologies) to label the
52
nuclei. Images were collected using a Zeiss 710 confocal microscope (Central
Microscopy Research Facility, University of Iowa). Manipulation of images was limited to
adjusting the brightness and contrast levels using Zen Light 2009 (Carl Zeiss
Microscopy) or Photoshop (Adobe). A minimum of four individual transgenic tadpoles
were studied for every DNA construct.
Cell culture
HEK293 cells (ATCC) were maintained in DMEM supplemented with 10% FBS,
250 μg/mL fungizone and 1% penicillin/streptomycin (Life Technologies). For
immunocytochemistry, cells were seeded at a density of 0.3 x 106 cells/mL on
chambered glass coverslips (Thermo Scientific). A total of 0.4 μg plasmid DNA (0.2 μg
for the HA-HCN1 constructs and 0.2 μg for the GFP or GFP-TRIP8b) was transfected
using Lipofectamine 2000 (Life Technologies). 24 hrs post-transfection, cells were fixed
in 4% paraformaldehyde and stained as described above with α-HA antibodies (diluted
1:500; Thermo Scientific). Each experiment was replicated a minimum of three times.
Biotinylation assays
HEK293 cells were seeded at a density of 0.75 x 106 cells/mL in 6-well plates
and transfected with a total of 2 μg plasmid DNA (1 μg for the HA-HCN1 constructs and
1 μg for the GFP or GFP-TRIP8b) using Lipofectamine 2000 (Life Technologies). 24h
post-transfection, cells were incubated with 1mg/mL sulfo-NHS-SS-biotin (Thermo
Scientific) for 15min at 4°C to label only the surface proteins. The reaction was
quenched in TBS (50 mM Tris-Cl and 150 mM NaCl, pH7.5) for 15min at 4°C, and
washed in PBS. The biotinylated cells were homogenized in 200 µl lysis buffer (50 mM
53
Tris-HCl, 10 mM NaCl, 0.32 M sucrose, 5 mM EDTA, 2.5 mM EGTA, 1.5% Triton X-100,
0.75% DOC and 0.1% SDS, pH7.4) supplemented with protease inhibitor cocktail
(Complete mini, Roche). Insoluble material was removed by centrifugation at 16,000 x g
for 10min. 100 µl of the supernatant was collected and incubated with 50 µl NeutrAvidin
agarose resin (Thermo Scientific), with rotation at 4°C overnight. Proteins bound to the
beads were washed with PBS three times and eluted with 50 µl reducing NuPage LDS
sample buffer (Life Technologies) followed by Western blotting with the following
antibodies: rabbit α-HCN1110, rabbit α-TRIP8b110, mouse α-NKA (M7-PB-E9, diluted
1:1000; Santa Cruz Biotechnology), rabbit α-GAPDH (diluted 1:500; Abcam), goat αrabbit HRP and goat α-mouse HRP (Sigma-Aldrich). The software package Image
Studio v3.1 (LI-COR Biosciences) was used for analysis of the blots. Each experiment
was replicated a minimum of three times.
Results
The C-terminus of HCN1 redirects an integral membrane reporter to the ER in
Xenopus photoreceptors
Photoreceptors express homotetrameric HCN1 channels. The major features of
each monomer are an intrinsically disordered cytoplasmic N-terminus, followed by six
transmembrane domains, and a cytoplasmic C-terminus. The membrane proximal
portion of the C-terminus consists of the C-linker and CNBD followed by a long
intrinsically disordered region (i.e. the post-CNBD region) (Figure 3-1A,B)30,143. To
determine the contribution of the HCN1 C-terminus to trafficking of the channel in
photoreceptors, we fused this sequence to a membrane reporter. This reporter consists
54
of a single transmembrane domain anchoring GFP which is followed by a peptide
sequence that can be dually palmitoylated (Figure 3-1C)24. The advantage of this
reporter is that it accumulates in the OS when expressed in transgenic Xenopus
photoreceptors, but relocalizes when fused to a protein sequence containing targeting
information. Furthermore, as an integral membrane protein it will be co-translationally
inserted into the ER, mimicking the initial steps of HCN1 trafficking. The membrane
reporter fused to the C-terminus of X. tropicalis HCN1 (Figure 3-1D) was found in
internal compartments within the IS. This pattern is reminiscent of the distribution of ER
in frog rod photoreceptors25. Immunostaining the same section with antibodies against
calnexin (an ER-resident protein) verified co-localization of the exogenous proteins with
the ER (Figure 3-1D-F). This result suggests that the C-terminus of HCN1 contains an
ER-targeting signal; a possibility tested with a series of deletion mutants described
below. The results are summarized in Table 3-1.
The ER targeting signal is located within the intrinsically disordered region
We first tested if the C-linker or/and CNBD are responsible for the ER localization
of the reporter-HCN1 CT construct. Note, the expression of fusion proteins containing
the CNBD resulted in large numbers of intracellular aggregates and sometimes cell
death in transgenic photoreceptors (data not shown). Incorporation of a single point
mutation, R524E, previously shown to disrupt cyclic nucleotide binding but not the other
functions of HCN1144, significantly reduced this problem. Therefore, the five constructs
tested in Xenopus photoreceptors that contain the CNBD include the R524E
modification. Using either the individual C-linker or CNBD domains fused to the reporter,
resulted in proteins that were not localized to either the OS or ER.
55
Instead, they were found accumulating in large amorphous structures in the
apical portion of the inner segments and/or synaptic terminals (Figure 3-2A,B). These
structures may be extremely large aggregates or the isolation of these two structural
domains may have exposed a cryptic association with mitochondria. Regardless, the
problem was abrogated when the C-linker and CNBD were used in tandem. This
caused the reporter to localize in OS (Figure 3-2C). Appending the disordered region to
the CNBD or using just the disordered region restored ER localization of the reporter
(Figure 3-2D,E). Together these data indicate that the ER targeting signal is located
downstream of the structured domains.
We next tested if any readily recognizable motifs in the disordered region of
HCN1 were responsible for the ER localization. Two regions drew our attention. There
is a stretch of amino acids enriched in glutamines and prolines, that we call the QP
region. The function of this region is currently unknown. Also, the last three amino acids
(SNL) are highly conserved in HCN channels and serve as the binding site for the
accessory subunit, TRIP8b119,120,145. Deleting the SNL signal did not prevent the ER
localization (Figure 3-3A), consistent with our previous finding that TRIP8b is not
required for regulating HCN1 trafficking in photoreceptors110. Similarly, eliminating the
QP region did not alter the ER localization pattern (Figure 3-3B).
Comparison of all the constructs exhibiting ER localization tested thus far allowed
us to constrain the region of interest to a shared 60 amino acid long region in the
beginning of the disordered portion of the C-terminus (Table 3-1, gray box). Deletion of
this region resulted in localization of the reporter to OS (Figure 3-3C, also 3-2C).
Considering that motifs involved in protein targeting are often conserved across
56
species, we examined an alignment of this region which drew our attention to a putative
di-arginine ER retention signal146. This motif is conserved among amphibians, reptiles,
birds, and mammals (Figure 3-3E). The sequence of HCN1 in X. tropicalis varies slightly
from the other species in having two overlapping di-arginine motifs (RMRTR). Three
point mutations (R631A, R633A, R635A) were made to test if either motif was
functional. Indeed, this resulted in localization of the reporter to the OS (Figure 3-3D,
compare with 3-2E). In conclusion, a conserved di-arginine ER retention signal in the
middle of the HCN1 C-terminus was responsible for the redistribution of the membrane
reporter to the internal membranes of the IS.
The ER retention signal regulates the trafficking of HCN1 to the cell surface
Di-arginine motifs found in a number of channels and receptors are thought to
mediate constitutive ER retention of partially folded or unassembled oligomers. Proper
assembly of the protein complex likely causes a physical masking of the di-arginine
motif thereby allowing exit from the ER18. The robust ER localization we observed in
Xenopus photoreceptors is consistent with this idea as the reporter constructs carrying
only the C-terminus of HCN1 cannot assemble into a properly organized channel. To
substantiate this, the requirement for the di-arginine motif should be tested in the
context of the full length HCN1. This is problematic in transgenic photoreceptors
because the experimental versions of HCN1 would likely oligomerize with endogenous
channels and override any mutations we incorporate. Therefore, we turned to a simpler
model system for assessing the efficiency of ER export for HCN1 channels.
We expressed HA-tagged mouse HCN1WT or HCN1MUT (R648A and R650A, thus
mutating the RxR motif to AxA) in HEK293 cells. Insertion of the HA-tag in the second
57
extracellular loop of mammalian HCN1 has been demonstrated to result in functional
channels with normal biophysical properties when overexpressed in HEK293 cells113,142.
Immunostaining of transfected cells revealed that HCN1WT was largely localized to the
ER. However, PM localization of HCN1MUT was prominent compared to that of the
wildtype channel (Figure 3-4A,B).
As a positive control, HCN1WT was co-expressed with GFP tagged TRIP8b (1a4), because this specific splice isoform of TRIP8b is known to increase surface
expression of HCN165,112,114. Indeed the PM localization of HCN1WT was dramatically
increased (Figure 3-4C). Note, in experiments lacking GFP-TRIP8b (1a-4), cotransfection of GFP was used to balance the amount of DNA transfected. In the
negative control, only GFP was expressed to demonstrate specific HA immunostaining.
The amount of PM HCN1 appeared greatest for HCN1WT + TRIP8b, intermediate for
HCN1MUT, and lowest for HCN1WT alone.
To quantify the surface expression of HCN1 we used biotinylation assays. Briefly,
all proteins presented on the cell surface were labeled with a membrane-impermeable
biotin reagent. The biotinylated surface proteins were subsequently isolated by an
affinity column and Western blotting was used to compare the amount of HCN1 in the
pool of surface proteins versus total cellular proteins. Co-transfection of GFP-TRIP8b
(1a-4) with HCN1WT increased the surface expression of HCN1WT by four-fold.
Consistent with the qualitative imaging analysis, HCN1MUT also increased surface
expression over HCN1WT but only by two-fold (Figure 3-5). In conclusion, these data
demonstrate that the di-arginine motif functions in the intact channel by limiting the
amount of channel allowed to traffic from the ER to the PM.
58
Discussion
The central finding of this study is that HCN1 contains a di-arginine ER retention
signal, conserved across species, that influences the amount of channel trafficked to the
PM of both undifferentiated (HEK293) and polarized (photoreceptors) cells. As one of
the carriers of Ih, HCN1 plays essential roles in neuronal cells by modulating membrane
excitability and signal integration thereby influencing a diverse set of processes.
Controlling the amount of HCN1 available in the PM can affect the amount of current
that can be conducted in response to various stimuli.
To date a number of missense mutations reported in human HCN1 demonstrate
that disrupting this channel leads to various forms of epilepsy147,148. Most of these
mutations occur near the pore thus causing altered activity and concomitantly reducing
expression of the channel. In places such as the retina, it seems that excess HCN1 is
available as the 40% reduction in HCN1 heterozygous mice or TRIP8b knockout mice
does not grossly affect visual function110. This implies that an additional insult that
further reduces the amount of surface expressed channel or a mutation in the diarginine motif that would enhance the surface expression of a channel with altered
activity could tip the balance and cause even more severe disorders.
The di-arginine ER retention signal is found in unrelated channels and receptors
including NMDA receptors, GABAB receptors and ATP sensitive potassium channels
(KATP)19,149-152. It has been proposed that di-arginine motifs are only functional if found
within approximately 16-46Å from the membrane153. The di-arginine motif in HCN1 is
separated from the membrane by the C-linker and CNBD that together occupy 52Å in
the crystal structure of HCN257. This difference in distance is small and we predict that
59
as more structures for membrane proteins become available, other di-arginine motifs
will be found within this range.
One model of the function of di-arginine motifs was proposed for the KATP
channels which require four inward rectifier potassium (Kir6.2) channel subunits and four
sulfonylurea receptor (SUR) subunits for proper assembly. Both Kir6.2 and SUR contain
the di-arginine ER retention signal and neither subunit is expressed at the PM alone.
Assembly into a heterooctamer likely sterically hinders the di-arginine motifs, allowing
trafficking to the PM19. Could the di-arginine motif in HCN1 function by a similar
mechanism? We observed that fragments of HCN1 containing the di-arginine motif but
incapable of forming a channel show robust ER localization. However, in intact channels
a mixture of PM and ER localized channels was observed with an intact di-arginine
motif shifting the balance more towards the ER. This supports the idea that the diarginine motif in HCN1 could function similar to the KATP channels. The only known
subunit that hetero-oligomerizes with HCN1 in photoreceptors is TRIP8b, which we’ve
shown is dispensable for surface expression of HCN1 in the retina (Chapter II)110.
Therefore, the di-arginine ER retention signal present in HCN1 is more likely monitoring
homo-oligomerization in photoreceptors rather than hetero-oligomerization.
Specific HCN channels are enriched in particular brain regions although most
show some degree of overlap such as hippocampal neurons which express both HCN1
and HCN243-47. It is speculated that in these neurons HCN1 and HCN2 are forming
heteromeric channels which in model expression systems have distinct biophysical
properties. Since HCN2 and HCN4 lack a di-arginine motif, hetero-oligomerization with
HCN1 would provide an additional level of control to the trafficking of these HCN hetero60
oligomers. On the other hand, HCN3 contains a putative di-arginine motif (RKR)
sequence following the CNBD and interestingly, HCN3 has been shown to localize
primarily to ER-like structures when transfected into opossum kidney cells154. Clearly
the di-arginine motif in HCN1 regulates trafficking out of the endomembrane system, but
the role of this motif in the trafficking of related channels remains to be dissected.
Another outstanding question is how regulating trafficking in the early secretory
pathway by the di-arginine motif is coordinated with other modulators of HCN1
trafficking. In this study, we focused on the role of the C-terminus exactly because this
region has been found to contain the binding sites for proteins proposed to regulate
HCN trafficking in various cells30. We had previously been surprised to find that loss of
TRIP8b did not decrease the surface expression of HCN1 in the retina (beyond that
dictated by the reduction in total HCN1 protein available) as it does in cortical
neurons110. Using photoreceptors as one of our model systems in this study, we were
able to reveal a previously undiscovered mode of regulation. When we mutated the diarginine motif in our reporter-based assay we observed that the reporter filled the outer
segments but was not specifically accumulating in the PM of the IS, where the
endogenous channel is found. This result suggests one of two scenarios; either
additional signals operating through the C-terminus of HCN1 can only function in the
context of a full length molecule and/or properly folded channel, or regions of HCN1
outside of the C-terminus carry signals that influence its trafficking. Future studies
designed to distinguish among these possibilities would be valuable in mapping out the
trafficking pathway of HCN1 from ER to PM which in turn should pinpoint targets that
61
can be manipulated to modify the amounts of HCN1 that are trafficked to the surface in
either healthy or diseased cells.
In summary, we identified a novel di-arginine ER retention signal in the Cterminus of HCN1. The signal functions to limit the amount of HCN1 that traffics from
ER to the PM. We propose this is part of the tight control imposed on HCN1 trafficking
given that altered surface expression of HCN1 is often observed in diseases such as
epilepsy45.
62
Table 3-1. A summary of the design and targeting behaviors of all constructs
expressed in transgenic Xenopus photoreceptors
The linear arrangement of sequence motifs in the HCN1 C-terminus are displayed on
top. The range of HCN1 amino acids attached to the reporter in each construct is listed
in the first column. The gray shaded area is required for ER localization. The red cross
represents mutation to the di-arginine motif. Abbreviations: CNBD, cyclic nucleotide
binding domain; QP, glutamine and proline rich region; OS, outer segment; ER,
endoplasmic reticulum. The table was published in Cellular and Molecular Life
Sciences111.
63
Figure 3-1. The C-terminus of HCN1 directs localization to the ER
A) A schematic representation of an HCN1 monomer (cyan) consists of six
transmembrane domains, with cytoplasmic N-terminus (NT) and C-terminus (CT). The
cyan oval in the C-terminus indicates the relative position of the CNBD. Amino acid
numbers flanking the domains of the CT are shown B) Disorder analysis of HCN1 using
meta Protein DisOrder prediction System. Residues above the threshold (grey dashed
line) are predicted to be intrinsically disordered. C-D) Transgenic Xenopus rod
photoreceptors expressing the reporter alone (C) or in fusion to the cytoplasmic Cterminus (aa 373-839, D) of X. tropicalis HCN1. A cartoon of the reporter and reporterHCN1-CT fusion protein is shown on the bottom. The transgenically expressed protein
(D, green) co-localizes with calnexin (E, red) in the inner segment (F, transmitted light).
Abbreviations: OS, outer segments; IS, inner segments; N (blue), nuclei; ST, synaptic
terminals. Scale bars: 5 μm. The figure was published in Cellular and Molecular Life
Sciences111.
64
Figure 3-2. The C-linker and CNBD are not required for ER localization
Transgenic Xenopus rod photoreceptors expressing the indicated Reporter-HCN1
fragments detailed in Table 3-1. Abbreviations and scale bars as in Figure 3-1. The
figure was published in Cellular and Molecular Life Sciences111.
65
Figure 3-3. Identification of a di-arginine ER retention signal
A-D) Transgenic Xenopus rod photoreceptors expressing the indicated Reporter-HCN1
fragments detailed in Table 3-1. Abbreviations and scale bars as in Figure 3-1. E) A
ClustalW sequence alignment of X. tropicalis HCN1 (aa 576-635) to HCN1 from various
animal species. Identical residues shaded in black, partially conserved residues in gray,
with the di-arginine motif outlined in red (asterisks mark arginines forming the RxR (red)
or RxRxR (blue) motif). Accession numbers are: X. tropicalis, XP002933077; X. laevis
a, b deduced from genomic scaffold v7.1 52441 and 337825. C. mydas,
XP_007052900; A. sinensis, XP_006017356; C. livia, XP_005500951; F. peregrinus,
XP_005242028; G. gallus, XP429145; B. mutus, ELR46479; R. norvegicus, W9JKB0;
M. musculus, O88704; H. sapiens, O60741. The figure was published in Cellular and
Molecular Life Sciences111.
66
Figure 3-4. The di-arginine motif influences plasma membrane localization of
HCN1
Immunostaining of HEK293 cells expressing the following proteins: A) wildtype HAtagged HCN1 and GFP; B) HA-tagged HCN1 with a mutated ER retention signal (RxR
to AxA) and GFP; C) Wildtype HA-HCN1 and GFP-TRIP8b; D) GFP alone. HA tag (red),
GFP (green), Nuclei (blue), and scale bars: 10 μm. The figure was published in Cellular
and Molecular Life Sciences111.
67
Figure 3-5. Mutating the di-arginine motif enhances surface expression
Biotinylation assays of HEK293 cells transfected as described in Figure 3-4A. A) After
biotinylation of transfected cells, surface proteins were pulled down using NeutrAvidin
beads. The level of HCN1 and TRIP8b in the total and surface pools was detected by
Western blotting. NKA and GAPDH were used as controls for membrane and cytosolic
proteins, respectively. B) Densitometry of Western blots represented in (A) was used to
calculate the surface to total ratio of HCN1 after normalization to the loading control
NKA. Asterisks indicate statistical significance with p < 0.01 (t-test). The figure was
published in Cellular and Molecular Life Sciences111.
68
CHAPTER IV - AN N-TERMINAL ER EXPORT SIGNAL FACILITATES
THE PLASMA MEMBRANE TARGETING OF HCN1 CHANNELS IN
PHOTORECEPTORS
Abstract
Hyperpolarization-activated cyclic nucleotide-gated 1 (HCN1) channels are
widely expressed in the central nervous system to modulate properties of the cell
membrane. Subcellular localization of HCN1 varies dramatically among cell types but
the mechanisms are poorly understood. We previously identified a di-arginine ER
retention motif that negatively regulates the surface targeting of HCN1. Using transgenic
X. laevis photoreceptors as the model system, we sought to identify a forward trafficking
signal that could counter the function of the ER retention signal. We found that the
HCN1 N-terminus can redirect a membrane reporter from outer segments to the plasma
membrane of the inner segment. The sequence necessary for this behavior was
mapped to a 20 amino acid region containing a leucine-based ER export motif. The ER
export signal is necessary for forward trafficking but not channel oligomerization as
demonstrated using velocity sedimentation. Moreover, this ER export signal alone
sufficiently counteracted the di-arginine ER retention signal. This expands our
understanding of how HCN1 trafficking is shifted from the secretory pathway to the
plasma membrane.
69
Introduction
Trafficking is one of the key regulatory mechanisms for HCN1 function,
exemplified by the altered HCN1 trafficking observed in CA1 pyramidal neurons after
temporal lobe epilepsy122. We have previously identified a di-arginine ER retention
signal at the C-terminus of HCN1 using X. laevis rod photoreceptors (Chapter III). This
ER retention signal negatively regulates the surface expression of HCN1. However, the
process of overcoming the retention signal under physiological conditions remains
unclear. An ER retention signal is usually counteracted by a forward trafficking signal
that promotes the movement of the protein from the endomembrane system to the
surface (i.e. plasma membrane). However, the overall effect of fusing the HCN1 Cterminus on a reporter membrane protein is to retain the reporter in the ER. This result
suggests that the forward trafficking signal is present in other regions of HCN1. One
candidate for mediating the forward trafficking signal is protocadherin 15 which interacts
with the N-terminus of HCN1 in inner ear cells155. However, HCN1 and protocadherin 15
are unlikely to interact in photoreceptors because they do not co-localize in
photoreceptors156,157, raising the question as to which other regions in HCN1 can
regulate its trafficking.
The goal of this study was to seek novel forward trafficking signals using our
established transgenic X. laevis approach for understanding membrane protein
trafficking138. We found that the intracellular N-terminus of HCN1 is necessary for the
protein to target the inner segment plasma membrane (ISPM). Through investigating a
series of truncation mutants, we identified a leucine-based ER export signal that can
override the di-arginine ER retention signal. This finding of combinatorial trafficking
70
signals controlling HCN1 localization provides insight into how the amount of HCN1
functioning at the cell surface is regulated under normal and disease conditions.
Materials and Methods
Molecular cloning, animals and immunohistochemistry were performed as
described in Chapter III.
Velocity sedimentation
HEK293 cells (ATCC, Manassas, VA USA) were maintained in DMEM
supplemented with 10% FBS and 1% penicillin/streptomycin (Life Technologies). Cells
grown in T75 flasks to confluence were transfected with 9 μg plasmid DNA using
Lipofectamine 2000 (Life Technologies). 48 hrs post-transfection, the cells were lysed in
PBS containing 1% CHAPS and 2x protease inhibitor cocktail (Roche). After clarification
by centrifugation at 10,000 x g for 15 min, the lysates were layered on a 10mL 5%-20%
continuous sucrose gradient and centrifuged at 209,500 x g for 16h at 4oC. 0.5mL
fractions were collected and analyzed by Western blotting. The antibodies used were
rabbit α-HCN1 (diluted 1:3000)110 and mouse α-NKA (diluted 1:100; M7-PB-E9, Santa
Cruz).
Results
The N-terminus is required for HCN1 to target the ISPM
In order for HCN1 to reach the PM, the di-arginine ER retention motif that we
previously identified should be balanced by the action of an as yet unknown forward
trafficking signal. The most probable locations for trafficking signals within HCN1 are the
71
intracellular domains, as they are spatially available to the majority of trafficking
regulators. HCN1 has four intracellular domains; the N-terminus (NT), the segment
connecting transmembrane domains 2-3 (L1), the segment connecting transmembrane
domains 4-5 (L2), and the C-terminus (CT) (Figure 4-1A). We fused each of these
HCN1 domains to a palmitoylated GFP reporter and examined their subcellular
localization when expressed in rods of transgenic X. laevis.
The membrane reporter consists of an EGFP followed by a palmitoylated peptide
from Xenopus rhodopsin (aa 311-349, Figure 4-1B). When expressed alone, the
reporter localizes mainly to the outer segment (OS) (Figure 4-1C)24,158. Consistent with
our previous finding using a similar integral membrane reporter, fusing the CT of HCN1
to the reporter redirects the protein to the ER (Figure 4-1D)111. Fusion of L1 did not
affect the default localization of the reporter (Figure 4-1E). Fusion of L2 drove the
reporter to the ellipsoid, a sub-compartment of the inner segment (IS) highly
concentrated with mitochondria (Figure 4-1F). It is possible that expression of this
isolated loop resulted in unmasking of a cryptic mitochondrial targeting signal. We did
not pursue this observation further because full-length HCN1 is not associated with
mitochondria. Fusion of the HCN1-NT to the reporter prevented it from localizing to OS.
Instead it was found in the IS with variable localization to both internal membranes with
morphology consistent with the ER and Golgi, and to the ISPM (Figure 4-1G). This
indicates that the NT of HCN1 contains trafficking information.
Since the NT only displays partial ISPM targeting when fused to the reporter, we
brought back other elements in HCN1 to test if the NT is required for the targeting of full
length HCN1 to the ISPM. When the full-length GFP-tagged Xenopus tropicalis HCN1
72
(aa 1-839) was expressed in transgenic Xenopus photoreceptors, the majority of the
protein was found at the ISPM (Figure 4-2A). HCN1 lacking the NT (GFP-HCN1; aa
122-839) was retained in the ER, evidenced by co-localization with calnexin, an ER
marker (Figure 4-2B-D). In total, these data show that the NT of HCN1 contains an
ISPM targeting signal.
Identification of an ER export motif in HCN1
We next set out to map the NT targeting signal. A series of N-terminal truncation
mutants of GFP-HCN1 were made to analyze their localization in transgenic Xenopus
rods (Figure 4-3). All HCN1 constructs generated in this study are summarized in Table
4-1. GFP-HCN1 still localized efficiently to the ISPM with the first 34, 54, or 74 amino
acids deleted (Figure 4-3A-C). However, removal of the first 94 amino acids changed
the localization to the ER (Figure 4-3D). Deletion of only aa 75-94 similarly resulted in
ER localization (Figure 4-3E). We conclude that these 20 amino acids are required for
HCN1 to exit from the ER and target to the ISPM.
Trafficking motifs are usually well conserved so we examined an alignment of the
critical 20 amino acid region in HCN1 from multiple species and found that it is almost
identical among amphibians, birds, and mammals (Figure 4-3F). Therefore any or all of
these residues could participate in trafficking. The first serine residue (Figure 4-3F,
arrow) in this region was found to be mutated to phenylalanine (S100F) in an early
infantile epileptic encephalopathy patient147. Human HCN1S100F has different
physiological characteristics from the wildtype HCN1. Compared to the wildtype HCN1,
the S100F mutant displays a half-activation voltage shifted to a more depolarized
potential, a faster activation and a slower deactivation. Interestingly, there is reduced
73
current density and decreased surface expression of this mutant compared to its
wildtype counterpart when heterologously expressed in CHO cells, indicating that the
serine residue may be crucial for trafficking HCN1 to the PM. To test this hypothesis, we
expressed GFP-HCN1S75F (X. tropicalis HCN1S75F is the equivalent mutant to human
HCN1S100F) in transgenic Xenopus photoreceptors. We found that HCN1S75F is able to
target to the ISPM with no more striking localization to internal membranes than the
wildtype channel (Figure 4-3G). We conclude that this serine is not essential for
trafficking and the alterations to the biophysical properties of HCN1 are the most likely
explanation for the development of epilepsy.
We next focused on investigating the –VNKFSL– sequence in the center of the
trafficking motif because it is similar to a VxxSL ER export motif identified in another
member of the voltage-gated potassium channel superfamily, Kv1.4159. Mutating any
one of the three key residues decreased the amount of Kv1.4 present at the cell surface
when tested in HEK293 cells. To determine if the VNKFSL sequence in HCN1 functions
similarly, we mutated three of the residues to alanines (V81A, S85A, L86A; GFPHCN1∆74:3A) and examined the localization of this mutant in photoreceptors. In support
of the hypothesis, GFP-HCN1∆74:3A lost the ability to target the ISPM and instead was
retained in the ER (Figure 4-3H).
We tested the requirement for each the three defining amino acids in the putative
VxxxSL motif by mutating each to alanine one at a time (Figure 4-4). Mutating the valine
gave rise to a variable phenotype. In two out of the six transgenic animals expressing
GFP-HCN1∆74:V81A the GFP-tagged mutant HCN1 localized primarily to the ISPM. But in
the remaining four animals GFP-HCN1∆74:V81A localized to the ER. This variability likely
74
reflects different expression levels in the individual animals and indicates that V81 is
part of the trafficking motif but is not essential (Figure 4-4A, B). Unlike the case with
Kv1.4, the serine in the HCN1 trafficking motif is not required as GFP-HCN1∆74:S85A
localized to the ISPM (Figure 4-4C). Mutating the leucine abolished the protein’s ability
to target the ISPM as GFP-HCN1∆74:L86A invariantly localized to the ER (Figure 4-4D).
We conclude that L86 is key to the function of this trafficking motif and it functions more
reliably with a valine several residues upstream.
The mutation of L86 to an alanine maintains a hydrophobic residue at this
position albeit a much smaller one. To gain further insight into the properties needed for
a functional ER export signal, we made two additional mutants. One mutant maintains
the hydrophobicity and size of the residue by converting it to an isoleucine (GFPHCN1∆74:L86I). The other changes the valine to an asparagine thereby maintaining the
size but not the hydrophobicity (GFP-HCN1∆74:L86N). GFP-HCN1∆74:L86I localized to the
ISPM while GFP-HCN1∆74:L86N was retained in the ER like GFP-HCN1∆74:L86A, verifying
that a relatively large hydrophobic residue is required at this position in the ER export
motif (Figure 4-4E, F).
It is possible that the trafficking defect we observe with the L86 mutations is
secondary to a defect in overall protein folding and assembly of a channel qualified for
post-ER processing through the endomembrane system. One biochemical approach to
assay membrane protein folding and assembly is to study their behavior in a density
gradient using velocity sedimentation. Mis-folded membrane proteins tend to aggregate
therefore having a higher density than the properly folded counterparts. On the other
hand, unassembled or partially assembled subunits usually sediment at lower density
75
fractions than the fully assembled complex. Lysates from HEK293 cells transfected with
either wildtype (HA-HCN1WT) or mutant mouse HCN1 (HA-HCN1VSLmut) were used to
provide an abundant source of HCN1 channels.
The sedimentation of HCN1 was determined by Western blotting for HCN1 in
fractions collected from 5-20% continuous sucrose gradients subjected to overnight high
speed centrifugation. The distribution of HA-HCN1WT and HA-HCN1VSLmut were not
significantly different (Figure 4-5). There is a slight shift of the peak of HA-HCN1VSLmut
fractions towards fraction 5, which may indicate a small change in the shape of the
channel. We used the sodium/potassium ATPase (NKA) as our internal size standard
because the NKAαβ heterodimer is about 130 kDa in size, similar to an HCN1
monomer. The distribution of NKA peaked much higher in the gradient than HCN1 at
fraction 7-8 vs fraction 4. We conclude that under these conditions HCN1 is more likely
an intact homotetramer than an unassembled monomer. We also generated the mutant
with only the leucine in the VSL motif mutated. As expected, HA-HCN1L100A behaved
the same as HA-HCN1VSLmut (data not shown). This leads us to conclude that the
mutation in this leucine (L)-based motif is primarily a trafficking defect.
The ER export motif overrides the function of the ER retention motif
In the next set of experiments we investigated the interaction between trafficking
signals carried out by the L-based ER export motif and the di-arginine ER retention
motif that we previously described. The PM localization of both wildtype HCN1 and
HCN1 with the di-arginine motif mutated (GFP-HCN1RxRmut) (Figure 4-2A and 4-6A)
indicates that the L-based ER export signal dominates over the ER retention signal.
76
Alternatively, there could be additional as yet unknown motifs promoting ER exit. To test
this possibility, we used triple mutant of the L-based ER export motif described above
(VNKFSL to ANKFAA) in the context of full-length HCN1, with an intact di-arginine ER
retention motif (GFP-HCN1VSLmut, Figure 4-6B). This mutant localized to the ER in
Xenopus photoreceptors, leading us to conclude that if there are any additional ER
export motifs in this channel they are not strong enough on their own to counteract the
information conveyed by the di-arginine ER retention motif. When we incorporated both
mutations (GFP-HCN1RxR + VSL) the GFP signal was found diffusely throughout the
photoreceptor OS and IS, including within the nucleus (Figure 4-6C). This pattern is
characteristic of soluble GFP and indicates degradation of the mutant HCN channel. It is
not surprising that crippling two important trafficking signals could trigger rapid
degradation. Altogether, our results point to the importance of both ER transport signals
in maintaining proper expression levels of HCN1 with the L-based ER export motif
dominating over the di-arginine ER retention motif.
Discussion
In response to membrane hyperpolarization, HCN1 channels mediate inward
cationic currents that modulate multiple aspects of neuronal function. Thus HCN1 is
subject to multiple levels of regulation. One level of cellular regulation involves
controlling the amount of HCN1 at the PM by integrating information from multiple
trafficking signals. We previously reported the discovery of a di-arginine ER retention
signal, which is a negative regulator of HCN1 surface expression. The aim of this study
was to identify a forward trafficking signal that could counter the action of the negative
77
regulator thereby increasing surface expression. By expressing a series of HCN1
mutants in transgenic Xenopus photoreceptors, we identified a leucine-based ER export
signal that positively regulates the PM targeting of HCN1.
The ER export signal in HCN1 is located in the cytoplasmic N-terminal domain
near the first transmembrane domain. This region is highly conserved, both across
species and even among the related HCN2, HCN3, and HCN4 channels, indicating a
shared mechanism of HCN channel trafficking. The leucine residue at position 100 is
essential to the function of the ER export signal, as mutation of this residue to either
alanine or asparagine resulted in ER retention of the channel. The critical leucine in
HCN1 is preceded by a valine and serine (VxxxSL) in a pattern similar to the ER export
signal found in Kv1.4 (VxxSL)160. However, the order of importance of the three
residues is different; in Kv1.4 it is L > S > V, but in HCN1 the order is L > V with S being
dispensable. Our observation, and the study on Kv1.4, point out the importance of the
bulky hydrophobic leucine residue in facilitating ER export. It is likely that the ER export
signals in HCN1 and Kv1.4 function in the same manner but the importance of individual
residues varies based on their microenvironment.
ER export signals function by recruiting COPII coat components which then
polymerize and deform the membrane so that cargo-containing transport vesicles bud
off from the ER161. The primary cargo receptor that binds to ER export signals in cargo
proteins is Sec24. Sec24 has the remarkable ability to select a diverse array of cargo
proteins in part because it has multiple cargo binding sites and modes of binding162-165.
A very intriguing recent study discovered that many multi-pass integral membrane
proteins can bind directly to Sec24 and indirectly via the Erv14 receptor 166. The authors
78
propose that cargo selection by coincidence detection is likely to enhance the efficiency
of selectively sorting proteins destined for the PM. The ER export signal we have
identified in HCN1 is most similar to hydrophobic motifs (e.g. the IRFTL motif in
Erv14167. Elucidation of the mechanisms of HCN1 and COPII recruitment would be
helpful in further delineating the trafficking pathway of HCN1.
Trafficking out of the ER is a major quality control checkpoint. Therefore, it
seems properly folded and, in the case of multi-subunit complexes, assembled cargo
proteins should be the most efficient at recruiting Sec24. It is not completely understood
how this occurs. In some cases oligomerization in and of itself may be the signal for
export168. In the case of GPI-anchored luminal cargoes, maturation of the glycan moiety
is required for binding the p24 receptor, which then recruits Lst1p (a Sec24
homologue)169. The sedimentation profile of HCN1 was not changed when the leucinebased ER export motif was mutated therefore this motif is not needed for
oligomerization of the channel. Perhaps either the glycosylation of HCN1 and/or
oligomerization promotes binding to a sorting receptor similar to p24 or Erv14.
The necessity of properly regulating HCN1 is illustrated by de novo HCN1
mutations found in patients with early infantile epileptic encephalopathy147. The
HCN1S100F mutation displays lower current density as well as decreased surface/total
ratio when expressed in CHO cells, implying a possible trafficking defect. The S100
residue is in close proximity to the ER export signal (the critical leucine residue is
located at position 111 of human HCN1), so the S100F mutation may affect function of
the ER export signal via mechanisms such as phosphorylation. Alternatively, the S100F
mutation may alter the stability of HCN1 because the mutant also displays higher
79
susceptibility to proteasomal degradation than its wildtype counterpart. We tested the
effect of the S100F mutation in HCN1 trafficking/stability in vivo in Xenopus
photoreceptors but the mutant does not show obvious trafficking or stability defects in
this system. It should be noted that photoreceptors seem to have a greater tolerance for
HCN1 mutation/degradation than cortical and hippocampal neurons given that TRIP8b
knockout mice with approximately 40% loss of functional HCN1 retain normal vision but
have defects in motor learning and greater resistance to depressive behaviors67.
Transgenic mice expressing disease-specific HCN1 mutants at a physiological level will
help to elucidate the cell-type specific requirements for HCN1 trafficking in the
progression of disease.
A question raised in the discussion of Chapter III that remains to be answered is
what are the signals involved in targeting HCN1 to the ISPM in photoreceptors. Both
endogenous and exogenously expressed HCN1 localize to the ISPM and are excluded
from the OS. It was shown previously in Xenopus photoreceptors that exogenously
expressed membrane proteins require some type of IS targeting information to escape
the bulk flow of membrane to the continuously renewing OS25. The IS
compartmentalization signal of HCN1 is unlikely to be present in the C-terminus
because this portion of the channel can only redirect reporters to the ER111.
Furthermore, reporter constructs with mutations in the ER retention signal are released
to the OS, not to the ISPM. The N-terminus of HCN1 may contain at least part of the IS
targeting information because this portion of the channel has the ability to redirect the
reporter to the ISPM, though some of these proteins are also found to variable degrees
within internal compartments. Our truncation mutants of GFP-HCN1 suggests that such
80
a signal may reside in the essential 20 amino acid region we mapped in this study (aa
75-94 in Xenopus tropicalis HCN1) because GFP-HCN175-839 was able to target the
ISPM. However, further truncation mutants are not favorable for identification of the IS
compartmentalization signal because any mutation that cripples the ER export signal
allowed the ER retention signal to predominate such that the channels were trapped at
the earliest stage of trafficking through the endomembrane system. Site-specific
mutations across aa 75-121 of HCN1 without affecting the ER export signal will be
required to test this idea.
In conclusion, we identified a novel L-based hydrophobic ER export signal in the
N-terminus of HCN1. The signal serves as a positive regulator in controlling the surface
amount of HCN1 by promoting trafficking of HCN1 from the ER to the surface. There
are two major trafficking signals controlling the equilibrium between internal and surface
pools of HCN1 in photoreceptors. Under normal conditions, the ER export signal
dominates and the majority of the channels are found in the ISPM. Crippling the diarginine ER retention signal releases more HCN1 so that the surface levels increase.
Crippling the ER export signal allows the ER retention signal to dominate. When both
signals are disrupted the protein is subject to degradation. Overall, this work reveals a
hierarchy of trafficking signals regulating the surface availability of HCN1 channels.
81
Table 4-1. A summary of the design and targeting behaviors of all constructs
expressed in transgenic Xenopus photoreceptors
The reporter-HCN1 fusion constructs are listed on the top. The range of HCN1 amino
acids attached to the reporter in each construct is listed in the first column. The GFPHCN1 constructs are listed on the bottom. Regions of HCN1 are indicated above the
brackets. The gray shaded area is required for ISPM localization. The primary
localization and corresponding figure number for each construct are listed in the last two
columns. Abbreviations: TMD, transmembrane domain; CT, C-terminus; NT, Nterminus; L1/L2, intracellular loop 1/loop 2; OS, outer segment; IS, inner segment;
ISPM, inner segment plasma membrane; ER, endoplasmic reticulum; CYT, cytoplasm;
N, nucleus.
82
Figure 4-1. The N-terminus drives a membrane-associated reporter to the IS
A) A schematic representation of the topology of HCN1. The HCN1 monomer (cyan)
has six transmembrane domains and four cytoplasmic regions including the N-terminus
(NT), the C-terminus (CT) and two intracellular loops (L1 and L2). The cyan oval in the
C-terminus indicates the relative position of the cyclic nucleotide binding domain
(CNBD). B) Schematic demonstration of the reporter-based constructs studied. The
cartoon of the reporter (left) consists of a GFP (green) and a palmitoyled peptide (red).
Positions where the cytoplasmic regions of HCN1 (cyan) were attached to the reporter
were shown in the middle and the right. C-G) Transgenic Xenopus rod photoreceptors
expressing the indicated Reporter-HCN1 fragments detailed in Table 4-1. Abbreviations:
OS, outer segments; IS, inner segments; N (blue), nuclei; ST, synaptic terminals. Scale
bars: 5 μm. The figure in panel C was produced by Joseph Laird.
83
Figure 4-2. The N-terminus is required for HCN1 to target the ISPM
Transgenic Xenopus rod photoreceptors expressing the GFP tagged (green) full length
HCN1 (A) or HCN1 mutant with the N-terminus truncated (B) are shown. The section in
B was also labeled with antibodies against calnexin (C and D, magenta). Abbreviations
and scale bars as in Figure 4-1. Joseph Laird contributed to generation of the
transgenic tadpoles.
84
Figure 4-3. Identification of a VxxxSL ER export motif in HCN1
Transgenic Xenopus rod photoreceptors expressing the indicated GFP-HCN1 truncation
(A-E) and point mutation (G and H) mutants detailed in Table 4-1 are shown.
Abbreviations and scale bars as in Figure 4-1. F) A ClustalW sequence alignment of X.
tropicalis HCN1 (aa 75-94) to HCN1 from various animal species. Identical residues
shaded in black, partially conserved residues in gray. The VxxxSL motif is marked with
red asterisks. The serine residue found to be mutated in epilepsy patients is indicated
by the black arrow. Accession numbers are: X. tropicalis, XP002933077; X. laevis a, b
deduced from genomic scaffold v7.1 52441 and 337825; G. gallus, XP429145; B.
mutus, XP005909291; R. norvegicus, W9JKB0; M. musculus, O88704; H. sapiens,
O60741. Joseph Laird and David Yamaguchi contributed to generation of the transgenic
tadpoles.
85
Figure 4-4. A bulky hydrophobic residue is key for the ER export signal to
function
A-F) Transgenic Xenopus rod photoreceptors expressing the indicated GFP-HCN1
mutants detailed in Table 4-1. Note, two images of the GFP-HCN1∆74:V81A mutation are
shown to demonstrate the range of phenotypes observed with this mutant (A,B).
Abbreviations and scale bars as in Figure 4-1. Joseph Laird and David Yamaguchi
contributed to generation of the transgenic tadpoles.
86
Figure 4-5. Disrupting the ER export motif does not affect the assembly of HCN1
A) Lysates from the HEK293 cells expressing either HA-tagged wildtype (HCN1WT)
HCN1 or mutant HCN1 with the ER export motif mutated (HCN1VSLmut) were subjected
to velocity sedimentation through 5-20% sucrose gradients. Fractions collected from the
sucrose gradient were analyzed via Western blotting. The endogenous sodium
potassium ATPase (NKA) was used as the standard for protein size and fraction
alignment. B) Quantification of the chemiluminescent signals from (A) and no
statistically significant difference was detected between HA-HCN1WT and HAHCN1VSLmut.
87
Figure 4-6. The ER export signal counters the action of the ER retention signal
A) GFP-HCN1 with an intact ER export signal but mutated ER retention signal
accumulates at the IS plasma membrane in transgenic Xenopus rod photoreceptors
(GFP-HCN1RxRmut). B) mutating the ER export signal alone prevents accumulation at the
IS plasma membrane (GFP- HCN1VSLmut). C) Mutation of both trafficking signals leads to
diffuse distribution of GFP throughout the cytoplasm and nucleoplasm. Lower panels
are the same image as in the upper panels with the nuclear stain not shown.
Abbreviations and scale bars as in Figure 4-1. Joseph Laird and David Yamaguchi
contributed to generation of the transgenic tadpoles.
88
CHAPTER V – A VxP CILIARY TARGETING MOTIF IS REQUIRED FOR
THE DIFFERENTIAL TRAFFICKING OF NKAα
α3 AND NKAα
α4
Abstract
Sodium potassium ATPases (NKA) are responsible for several cellular activities
including setting electrochemical gradients, cardiotonic steroid signaling and cellular
adhesion. NKA isozymes display distinct subcellular localization patterns. For instance,
NKAα4 localizes to the ciliary structure in the sperm where it is endogenously
expressed, as well as in the cilia (outer segments or OS) of photoreceptors when
exogenously expressed; In contrast, the localization of endogenous NKAα3 is restricted
to the inner segment plasma membrane (ISPM) of photoreceptors. The mechanism
controlling this distinct localization of NKAα isozymes is not understood. Ankyrin, a
family of scaffold proteins linking membrane proteins and spectrin, can regulate the
trafficking of NKA in some tissues. Thus it may contribute to the localization of NKA in
photoreceptors. In this study, we analyzed the interaction between ankyrin and NKA but
found that they are localized to different compartments in photoreceptors. To gain new
insights into NKA trafficking, we next analyzed the localization of a series of
chimera/truncation mutants of NKAα3/α4 expressed in transgenic X. laevis
photoreceptors. We uncovered that OS localization of NKAα4 is dependent upon a VxP
motif that was also identified in other cilia-localizing proteins (e.g. rhodopsin). Given the
structural similarities between the sperm flagellum and the photoreceptor OS, our
findings shed light on how NKAα4 is targeted to sperm flagella to support fertility.
89
Introduction
The sodium potassium ATPase (NKA) creates an electrochemical gradient by
actively shuttling Na+ ions outside and K+ ions inside cells. Since NKA utilizes ATP
hydrolysis for transporting ions, it is one of the major consumers of energy in the cell.
Additionally, NKA carries out cellular functions including cell-cell adhesion and signaling
transduction69-73,170,171. NKA functions as a heterodimer containing a large catalytic α
subunit and a small but heavily glycosylated β subunit (for details, see Chapter I). There
are four genes (ATP1A1-4) encoding for the four NKAα isozymes (NKAα1-4), and their
expression pattern is tissue-specific.
The functions of NKAα isozymes are critical for the specific-tissues they are
expressed in. NKAα1 is ubiquitously expressed and it is not surprising that genetic
ablation of ATP1A1 in mouse is embryonic lethal172. Mutations in NKAα1 can lead to
aldosterone-producing adenomas and hypertension173-175. NKAα2 can be found in
muscles and the brain. Its mutations are linked to migraine83. NKAα3 is enriched in the
nervous system, and its mutations can cause movement disorders, rapid-onset dystonia
parkinsonism and alternating hemiplegia of childhood, type 2176,177. NKAα4 is
specifically expressed in mammalian sperm85-88. Genetic deletion of ATP1A4 in mice
results in low sperm motility and infertility.
Distinct sub-cellular localization of NKAα isozymes adds another level of control
of this ion pump. In hippocampal neurons, NKAα1 shows diffused localization in the
plasma membrane (PM) while NKAα3 shows clustered localization at the contacting
points between the PM and the ER178. In photoreceptors, the localization of NKAα3 is
90
restricted to the ISPM. This restricted localization of NKAα3 ensures that its large
energy expenditure does not interfere with the energy flow in other compartments. In
the sperm, NKAα4 is located in sperm flagella, the motile cilia supporting sperm
mobility. The distinct sub-cellular compartmentalization of NKAα isozymes supports
their function in the specific cell. To date, the players involved in the differential
trafficking remain unclear.
It is known that NKAβ plays important roles in the ER export and PM anchoring
of NKAα but does not seem to determine the sub-cellular compartmentalization.
Ankyrins (Ank) are adaptors for many membrane proteins to spectrin-based
cytoskeleton99-101. Three groups of ankyrins were previously identified, namely AnkR,
AnkB, and AnkG. A three-step model was proposed for trafficking NKAα1 in epithelial
cells. The first step is to traffic NKAα1 from the ER to the Golgi via direct binding to
AnkR. The second step is to traffic NKAα1 from the Golgi to the PM via the help from
AnkG104. The third step is to tether NKAα1 to the lateral side of the PM, a process likely
dependent on the binding between AnkB and spectrin. It is not known if this model
established in epithelial cells can be applied to other systems such as the
photoreceptors, where ankyrin was proposed to regulate the expression and possibly
the trafficking of NKA107. Here in this study, we evaluated the possibility by testing AnkB
and AnkR as candidates for regulating NKA compartmentalization in photoreceptors but
did not find a proof of co-localization.
To obtain more insight into how trafficking of NKAα isozymes is regulated using
photoreceptors as a model, we investigated trafficking of NKAα3 and NKAα4 in
91
transgenic Xenopus photoreceptors. We found that when overexpressed in
photoreceptors, NKAα3 is found in the ISPM similar to the endogenous NKAα3. In
contrast, NKAα4 localized to the OS, in agreement with its ciliary localization in the
sperm. By analyzing localization of a series of mutants using confocal microscopy, a
VxP motif was found to be responsible for the OS targeting of NKAα4. However, the
VxP motif is insufficient to redirect NKAα3 from the ISPM to the OS. Our study serves
as one of the initial steps for understanding how distinct compartmentalization of NKAα
isozymes can be achieved.
Materials and Methods
Molecular cloning
All transgenic constructs were subcloned into the XOP5.5 vector to drive rod
photoreceptor expression. Constructs were created using standard PCR protocols and
verified with Sanger sequencing (Iowa Institute of Human Genetics, University of Iowa).
Sequences of plasmid inserts and primers are available on request. The cDNAs used
as templates for cloning the various constructs included Xenopus laevis ATP1A3
(BC043743), Xenopus laevis ATP1A1 (NP_989407.1), and Homo sapiens ATP1A4
(BC094801) were all purchased from Open Biosystems.
Animals
Mice (C57/Bl6J) and adult Xenopus laevis were obtained and maintained as
described in Chapter II and Chapter III respectively. Transgenic Xenopus tadpoles were
generated using restriction enzyme mediated integration as described in Chapter III.
92
Immunohistochemistry
Transgenic tadpoles were processed as described in Chapter III with the
following antibodies: mouse α-NKA (1:1000, M7-PB-E9, Santa Cruz), rabbit α-Ankyrin B
(1:500, H-300, Santa Cruz), mouse α-Ankyrin B (1:100, clone N105/13, NeuroMab),
rabbit α-Ankyrin R (1:100, H-133, Santa Cruz), mouse α-Ankyrin R (1:100, H-4, Santa
Cruz).Images were collected using a Zeiss 710 confocal microscope (Central
Microscopy Research Facility, University of Iowa). Manipulation of images was limited to
adjusting the brightness and contrast levels using Zen Light 2009 (Carl Zeiss
Microscopy) or Photoshop (Adobe). A minimum of three individual transgenic tadpoles
were studied for every DNA construct.
Subcellular fractionation
Frozen bovine retinas (Lawson) were homogenized in IP buffer (50 mM Tris, pH
7.3, 10 mM NaCl, 0.32 M sucrose, 5 mM EDTA, 2.5 M EGTA and 1 mM PMSF)
supplemented with protease inhibitor cocktail (Complete, Roche) and clarify by
centrifugation at 1,000 x g for 10min. Immunoprecipitations were carried out with 1 mg
of lysate and 5 µg of either mouse α-NKA (M7-PB-E9, Santa Cruz), mouse α-NKA α3
(XVIF9-G10, Thermo Scientific), mouse α-Ankyrin B (clone N105/13, NeuroMab), or
mouse IgG (Sigma) incubated with 1.5 mg Protein G Dynabeads (Life Technologies) for
2h. The beads were washed in PBS and eluted in LDS sample buffer. IP with each
precipitating antibody were replicated at least twice. For retina fractionation experiments
the retinas were depleted of photoreceptor outer segments by gentle shaking in Buffer A
(50% sucrose, 10 mM HEPES, pH7.4, 1 mM EDTA and 5 mM MgCl2) supplemented
93
with protease inhibitor cocktail. After centrifugation at 13,000 x g, 1hr the pelleted
retinas were resuspended in Buffer A without sucrose. A centrifugation step at 750 x g
for 10min was used to clarify the lysate. The “total” lysate was then centrifuged at
100,000 x g for 1h to separate soluble proteins from membranes. Antibodies for
Western blotting were used as follows: mouse α-NKA (1:1000, M7-PB-E9, Santa Cruz),
rabbit α-HCN1 (1:2500)110, mouse α-actin (1:1000, AC-74, Sigma), rabbit α-GAPDH
(1:250, Abcam), mouse α-Ankyrin B (1:1000, clone N105/13, NeuroMab), mouse αAnkyrin R (1:200, H-4, Santa Cruz).
Results
Distinct compartmentalization of ankyrins and NKAα in photoreceptors
Ankyrins are important for NKA trafficking at various stages in epithelial cells. To
test if ankyrin proteins can also regulate NKA trafficking in photoreceptors, we started
by testing for an interaction between NKA and AnkB using co-immunoprecipitation
(Figure 5-1A). The result shows that AnkB fails to co-immunoprecipitate with NKA from
bovine retina although the reciprocal experiment using anti-AnkB antibodies did pull
down a small percentage of NKA. Next, we separated the retina lysate into soluble and
membrane protein fractions (Figure 5-1B). Actin and GAPDH are soluble proteins
therefore serve as markers for the soluble fraction. The hyperpolarization activated
cyclic nucleotide gated channel 1 (HCN1), a voltage-gated channel, is used as the
marker for the membrane faction. As expected, NKA is only present in the membrane
fraction, however AnkB is greatly enriched in the soluble fraction.
94
The co-immunoprecipitation and subcellular fractionation assays suggest that
only a small fraction of AnkB is interacting with NKA in the retina. This prompted us to
revisit the localization of AnkB and NKA in the bovine retina. In the photoreceptor layer,
NKA is only detected in the IS, where the AnkB signal is not found (Figure 5-1C).
Instead, AnkB is found concentrating at the synaptic layers of the retina (OPL and IPL).
We tested AnkR using the same assays and found similar results (Figure 5-1B,D).
AnkG was not tested because it localizes to the OS of photoreceptors172. The lack of colocalization between ankyrins and NKA in photoreceptors led us to search for alternative
mechanisms controlling the localization of NKA in these cells.
The N-terminus of NKAα contains information for their differential trafficking
In order to search for determinants that control NKA trafficking in photoreceptors,
we used the transgenic Xenopus approach described in Chapter III and IV for
identification of novel trafficking signals. We expressed GFP-α3 in Xenopus
photoreceptors and the protein localizes to the ISPM as expected (Figure 5-2A). In
contrast when GFP-α4, the sperm-specific isozyme, was expressed in Xenopus
photoreceptors, it localizes to the OS (Figure 5-2B). The localization of NKAα4 in the
photoreceptor is consistent with its localization in the sperm because it is found in the
ciliary compartments of both cells. The distinct localization between NKAα3 and NKAα4
in photoreceptors suggests that targeting of these two isozymes is controlled by
different mechanisms. The sequences among NKAα subunits are highly conserved.
Although NKAα4 is the most divergent isozyme, it preserves 76% sequence identity to
95
NKAα3. Within the entire protein, the N-terminus of NKAα is the least conserved region
and is most likely to participate in the differential regulation of NKAα trafficking.
We hypothesized that the N-terminus of NKAα3 and/or NKAα4 contains targeting
signals responsible for their distinct localization in photoreceptors. To test the
hypothesis, we generated α3/α4 chimeras whose N-termini are switched. All constructs
analyzed in this study are summarized in Table 5-1. By studying the chimeras, we found
that the N-terminus of NKAα3 redirects NKAα4 to the IS (Figure 5-2C). Similarly, the Nterminus of NKAα4 redirects NKAα3 to the OS (Figure 5-2D). The results suggest that
the targeting signals are located in the N-terminal regions of NKAα3 and/or NKAα4.
In order to locate the targeting signals within the N-terminus of NKAα3 and/or
NKAα4, we analyzed the disorder tendency of their N-termini because trafficking signals
are often present in disordered/unstructured regions146,179. The structure of NKA has
been resolved however it lacks the N-terminus, suggesting the flexible nature of the Nterminus76,78. Therefore we used the meta Protein DisOrder Prediction System143 to
analyze the disorder tendency of the N-termini of NKAα3 and NKAα4. The disorder
tendency plot shows that the first ~40aa of these two proteins are intrinsically
disordered (Figure 5-3A).
To test if the disordered regions contain trafficking signals, we generated another
set of chimera mutants containing shorter N-terminal chimeric region (~40aa from the
start of the N-terminus). The result shows that GFP-α3(1-38)/α4 localizes to the ISPM
(Figure 5-3B) and GFP-α4(1-44)/α3 localizes to the OS (Figure 5-3C), recapitulating the
phenotype of chimeras containing the entire N-terminus and supporting the idea that
96
targeting signals are within the intrinsically disordered region of NKAα3 and/or NKAα4.
We subsequently generated chimeras with shorter N-terminus to restrict the regions
with potential trafficking signals in aa 1-14 of both NKAα3 and NKAα4 (Figure 5-3D-G).
A VxP OS/ciliary targeting motif is required for NKAα4 to target the OS
To decide which of the isozymes contains the specific trafficking signal, we
deleted aa 1-14 in NKAα3 and NKAα4 respectively and tested if the regions are
required for their own localization. Deleting aa 1-14 from NKAα3 does not alter its ISPM
localization (Figure 5-4A), but deleting the same region in NKAα4 prohibits its OS
localization, and the mutant is targeted to the ISPM instead (Figure 5-4B). Our results
suggest that NKAα4 has an OS targeting signal within aa 1-14. Alignment of NKAα4
sequence in comparison to the one of NKAα3 reveals a region reminiscent of a VxP
ciliary/OS targeting motif180,181 (Figure 5-5A). To test whether both of the valine and
proline residues are important for the OS targeting of NKAα4, we engineered alanine
mutations to these two residues.
In contrast to the OS localization of GFP-α4 (Figure 5-2B), the mutant GFPα4V10A, P12A localizes to the ISPM instead (Figure 5-5B), likely through the same
mechanism controlling NKAα3 targeting. Similar to GFP-α4V10A, P12A, GFP-α4V10A and
GFP-α4P12A mutants are found in the ISPM, indicating that both the valine and proline
are necessary for the OS targeting of NKAα4 (Figure 5-5C,D). Interestingly, NKAα3
contains a valine but not a proline in the same relative position as the VxP motif in
NKAα4 (Figure 5-5A). We tested if the ISPM targeting of NKAα3 can be overridden by
an artificial VxP motif by generating a NKAα3 mutant GFP-NKAα3T13P (Figure 5-5E).
97
The mutant does not lose its ISPM localization, suggesting that the VxP motif is
necessary but not sufficient for the OS targeting.
In summary, our results indicate that a VxP motif present in NKAα4 but absent in
NKAα3 is responsible for the differential trafficking we observed for these two NKA
isozymes in photoreceptors. NKAα4 is a sperm-specific isozyme whose localization is
restricted to the ciliary flagellum of the sperm. We found that the VxP motif directs
trafficking of NKAα4 to the photoreceptor OS, which is also a ciliary organelle. This
finding provides valuable information on how NKAα4, the sperm-specific isoform, is
localized to the ciliary compartment in its host cell.
Discussion
Our study provides two major findings regarding the trafficking of NKA. The first
is that contrary to its role in epithelial cells, ankyrin is unlikely to play a role in the
trafficking of NKA in photoreceptors. Second, a VxP motif directs NKAα4 to the OS
when NKAα4 is heterologously expressed in photoreceptors. We propose that this
mechanism controls the localization of NKAα4 in the sperm flagellum since both sperm
flagella and photoreceptor OS are ciliary organelles. Altogether, this work emphasizes
the concept that there are distinct mechanisms controlling the cell-specific trafficking of
NKA isozymes.
In some cell types ankyrin does play a more prominent role in the localization of
NKA than it does in photoreceptors. In addition to the three-step regulation of NKA
trafficking in epithelial cells, AnkB is important for organizing NKA into a specific microdomain in the T-tubule of ventricular cardiomyocytes. This microdomain contains NKA,
98
the Ca2+ pump SERCA2, and the Na+/Ca2+ exchanger NCX1. The micro-domain allows
efficient Ca2+ clearance from the intracellular space. Both NCX1 and NKA are absent
from the T-tubule with reduced expression of AnkB182 In the brain, a similar role of AnkB
in organizing NKA and NCX1 at the ER - PM junctional membranes has been
discussed106,183. In photoreceptors, AnkB was implicated to regulate the localization of
NKA107,184. However, we did not detect AnkB in the ISPM but in the synaptic layers of
the retina. The expression of AnkR in the retina had not been previously investigated
and we found that it has a similar though not identical distribution to AnkB. Even though
the majorities of AnkB and AnkR are present in the soluble fraction that does not include
NKA, some NKA can be co-immunoprecipitated with AnkB. This likely reflects
interactions coming from the synaptic layers in the retina.
In order to identify new mechanisms controlling the NKA trafficking in
photoreceptors, we turned to the transgenic Xenopus model system that has been
particularly fruitful in elucidating novel trafficking signals. We first confirmed that
overexpressed GFP-tagged NKAα3 localizes correctly to the ISPM. Since trafficking of
NKAα to the PM is known to be dependent on the formation of the NKAαβ heterodimer,
this indicates that the GFP-NKAα3 is able to associate with the endogenous β subunit.
In contrast, GFP-NKAα4 exogenously expressed in photoreceptors localizes to the
ciliary OS. This distinct localization led us to search for trafficking signals by analyzing
the localization of chimeric and truncated NKAα mutants. We determined that a VxP
motif in the intrinsically disordered N-terminus of NKAα4 is necessary for its ciliary
localization in photoreceptors.
99
Cilia are specialized cellular compartments that serve multiple important
functions like signal transduction (sensory cilia) and cellular motility (motile cilia).
Transporting membrane components into and out of cilia is under tight controls. It is
generally believed that membrane proteins are packed into transport vesicles in the
Golgi, docked near basal bodies and enter cilia with the help from the intraflagellar
transport complex185,186. These processes are guided by specific ciliary targeting
signals, one of which is the VxP motif first discovered in rhodopsin11, the light-sensitive
pigment present in the OS of photoreceptors. The VxP motif was later found in the Nterminus of polycystin-2 (PC2), a protein affected in the autosomal dominant polycystic
kidney disease, and is necessary for targeting PC2 to the cilia187.
The VxP motif is possibly the determinant for NKAα4 to enter and stay in sperm
flagella. An ideal experiment to test this hypothesis is to express NKAα4V10A, P12A in the
sperm followed by examining its localization and sperm motility. Unfortunately, sperms
are transcriptionally and translationally silent so it is technically challenging to
exogenously introduce the NKAα4 mutant188. As an alternative, we examined
exogenous NKAα4 localization in inner medullary collecting duct (IMCD) and MadinDarby canine kidney (MDCK) cells that were often used to study ciliary trafficking.
Unlike photoreceptors or sperms, no NKAα4 was observed in the cilia of IMCD and
MDCK cells (data not shown), likely due to insufficiency of certain accessory proteins.
The accessory proteins needed for the VxP motif to function are not fully
resolved. One model based on rhodopsin trafficking suggests that the VxP motif
interacts with Arf4, a small GTPase within a ciliary targeting complex in the Golgi, to
generate rhodopsin-containing vesicles181. The later budding and trafficking of those
100
vesicles are regulated via a series of protein-protein interactions. In addition, the VxP
containing region in rhodopsin has been shown to interact with t complex testis
expressed-1 (Tctex-1), a ubiquitously expressed dynein light chain, indicating a role of
dynein in transporting rhodopsin-containing vesicles along microtubules189. Whether the
VxP motif in NKAα4 can interact with Arf4 and/or Txtex-1 in the sperm requires future
investigation both in vivo and in vitro.
In conclusion, we identified a VxP OS/ciliary trafficking signal in the spermspecific NKAα4 isozyme and predict that NKAα4 is targeted by the same signal to the
sperm flagella where it is normally present. Our findings serve as a valuable first step to
unravel the mechanisms leading to differential trafficking of NKA proteins.
101
Table 5-1. A summary of NKA constructs expressed in transgenic Xenopus
photoreceptors
The major structural elements of GFP-NKAα are shown on the top. GFP is represented
with a green oval. NKAα3 is displayed in orange and NKAα4 in black. The organization
of each construct used in this study is shown, along with a summary of the primary
localization and the corresponding figure number. Abbreviations: TM, transmembrane;
TMD, transmembrane domains; ISPM, inner segment plasma membrane; OS, outer
segment.
102
Figure 5-1. The interaction between ankyrins and NKA in the bovine retina
A) Co-immunoprecipitations from bovine retinal lysates, blotted for NKA and AnkB.
Antibodies against NKA (upper panel) or AnkB (lower panel) alongside IgG as a
negative control were used for the immunoprecipitation. B) Subcellular fractionation of
the bovine retina into soluble (verified by the enrichment of actin and GAPDH) and
membrane (verified by the presence of HCN1) fractions. AnkB, AnkR and NKA were
blotted using specific antibodies. C-D) Immunostaining of bovine retina with antibodies
against NKA (green) and AnkB (red, C) or AnkR (red, D). Nuclei (blue) are labeled with
Hoechst. Abbreviations: OS, outer segment; IS, inner segment; ONL, outer nuclear
layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer;
GC, ganglion cell layer; NFL, nerve fiber layer. Scale bars: 20 µm. Data from panels A
and B were generated by Modestos Modestou.
103
Figure 5-2. The N-terminus of NKAα
α3 and/or NKAα
α4 determines their distinct
localization
Transgenic Xenopus photoreceptors expressing GFP-NKAα3 (A), GFP-NKAα4 (B) and
chimeras switching the N-terminus of each other (C-D).The GFP tagged proteins are
shown in green. Abbreviations: OS, outer segments; IS, inner segments; N (blue),
nuclei; ST, synaptic terminals. Scale bars: 5 μm. Joseph Laird contributed to generation
of the transgenic tadpoles.
104
Figure 5-3. The first 14 amino acids of NKAα
α3 and/or α4 contain the targeting
information
A) Disorder analysis of the N-termini from NKAα3 and NKAα4 using meta Protein
DisOrder prediction System. Residues above the threshold (grey dashed line) are
predicted to be intrinsically disordered. GFP-tagged chimeras containing the N-terminus
of NKAα3 localize to the ISPM (B, D and F). GFP-tagged chimeras containing the Nterminus of NKAα4 localize to the OS (C, E and G). Abbreviations and scale bars as in
Figure 5-2. Joseph Laird contributed to generation of the transgenic tadpoles.
105
Figure 5-4. The first 14 amino acids of NKAα
α4 are required for its OS localization
A) Localization of NKAα3 with the first 14 amino acids deleted. B) Localization of
NKAα4 with the first 14 amino acids deleted. Abbreviations and scale bars as in Figure
5-2. Joseph Laird contributed to generation of the transgenic tadpoles.
106
α4
Figure 5-5. Identification of a VxP OS targeting motif in NKAα
A) An alignment of the N-terminal amino acids from Xenopus NKAα3 and human
NKAα4. The dashed line indicates aa 1-14 required for OS localization of NKAα4.
Triangles indicate residues mutated in following panels (B-E). Abbreviations and scale
bars as in Figure 5-2. Joseph Laird contributed to generation of the transgenic tadpoles.
107
CHAPTER VI – DISCUSSION AND FUTURE DIRECTIONS
Summary of the Thesis
The highly compartmentalized photoreceptor cell is among the most popular
models for studying protein trafficking. The outer segment (OS) is the sensory cilium,
housing proteins that are directly involved in the phototransduction cascade. In order to
support the massive activities from the signaling cascade, membrane components are
constantly imported from the bottom of the OS and shed on the top of the OS; therefore
early studies of photoreceptor membrane protein trafficking were focused on the OSresident proteins. Since OS-targeting is an active transport process, it was thought that
proteins without an OS-targeting signal would be retained in the cell soma (i.e. inner
segment, or IS) by default; however, Baker et al. later found that membrane proteins
lacking OS targeting signals can enter the OS, likely by being randomly incorporated
into the OS-destined transport vesicles25. The finding points out that trafficking
membrane proteins to the IS and the OS are equally important for the cell to function
properly. For this reason we started this study to identify regulators involved in
trafficking membrane proteins in photoreceptors with a focus in the IS.
There are a large number of IS-resident membrane proteins190. A majority of
them are channels and transporters that shape the electrical property of the plasma
membrane (PM). We selected two of these proteins (HCN1 and NKA) that are
indispensable for the proper function of photoreceptors as our model to study
membrane protein trafficking in the IS. There are certain regulators identified for
108
trafficking HCN1 and NKA in other cellular systems50,51,68,102-107. Some of them are likely
to play similar roles in photoreceptors; therefore we studied the role of TRIP8b in
photoreceptor trafficking of HCN1 in Chapter II, and studied the potential of ankyrins to
regulate NKA trafficking in photoreceptors in Chapter V.
Besides investigating the functions of known regulators of HCN1 and NKA, we
also set our goal to identify novel signals responsible for their trafficking in
photoreceptors. Among the commonly used laboratory animals, X. laevis is the most
favorable model for researching protein trafficking because of the large size of their
photoreceptors (about three times larger than mouse photoreceptors in diameter) that
enables easy identification of intracellular compartments vs the PM. Moreover, in vivo
study using transgenic Xenopus can be accomplished within a month given the rapid
growth of the embryos. Taking advantage of this model system, we were able to
successfully identify several trafficking signals in HCN1 and NKA including a di-arginine
ER retention signal (Chapter III), a leucine (L)-based ER export signal (Chapter IV) and
a VxP OS/ciliary targeting signal (Chapter V). The major findings in my thesis are
summarized in Figure 6-1.
Regulation of HCN1 by TRIP8b in Photoreceptors
In Chapter II, we investigated the expression profile of TRIP8b in the retina and
investigated its role in HCN1 trafficking. We concluded that TRIP8b is not required for
regulating HCN1 trafficking in photoreceptors because neither the retinal localization nor
surface expression of HCN1 changes in the absence of TRIP8b. We did observe an
interaction between TRIP8b and HCN1 in the mouse retina evidenced by co-
109
immunoprecipitation, subcellular fractionation and co-localization. Then what is the role
of TRIP8b? We found out that the protein level of HCN1 is reduced to ~40% of normal
in the TRIP8b knockout retina compared to the wildtype counterpart; therefore, TRIP8b
functions in the retina to maintain the maximal expression level of HCN1.
The amount of HCN1 protein can be regulated via multiple pathways including
transcription, translation and degradation. TRIP8b does not contain any trait of a
transcriptional or translational regulator, nor was it reported to interact with proteins in
those pathways. Therefore it is unlikely that TRIP8b regulates the production of HCN1
proteins. We favor the hypothesis that TRIP8b maintains the protein level of HCN1 via
preventing its degradation. Supporting this idea, Lewis et al., showed that the transcript
level of HCN1 (as well as other HCN proteins) in the hippocampus does not change
with or without the presence of TRIP8b68. Similar to our observation in the retina, those
authors found that the protein levels of both HCN1 and HCN2 in the hippocampus is
reduced to ~60% of normal, suggesting that the system controlling HCN1 expression via
TRIP8b is likely shared by the two tissues. With further investigation, the authors
showed that TRIP8b inhibits the lysosomal degradation of HCN168. Moreover, knocking
out TRIP8b in mice does not trigger ER stress or affect HCN1 exit from the Golgi68,
suggesting that the lysosomal degradation of HCN1 is not due to a global trafficking
defect. It requires future studies to reveal if TRIP8b maintains the maximal expression
of HCN1 in photoreceptors via the lysosomal pathway. Short-term culture of isolated
retina and treatment with pharmacological inhibitors in the protein degradation pathways
will be useful to test the hypothesis.
110
Other Candidates for Regulating HCN1 Trafficking in Photoreceptors
We showed that TRIP8b is not required for the proper trafficking of HCN1 in
photoreceptors. A question raised from the observation is what are the regulatory
proteins for HCN1 trafficking in these cells. Other HCN1-interacting proteins that could
participate in its trafficking include protocadherin-15, Nedd4-2, filamin A and caveolin-1.
HCN1 interacts with protocadherin-15 in cochlear hair cell stereocilia, but in
photoreceptors they are found in different subcellular compartments156,157.
Protocadherin-15 is concentrated at the OS and the synaptic terminal; therefore, is
unlikely to regulate HCN1 trafficking in the IS. Other than protocadherin-15, both
Nedd4-2 and filamin A can serve as negative regulators of HCN1 surface
expression50,51; however, studies of their expression and localization in photoreceptors
are needed in the future to explore this possibility.
Caveolin-1 is also a candidate to regulate HCN1 trafficking in photoreceptors.
Caveolin-1 functions to traffic and organize cholesterol, signaling molecules, and ion
channels (e.g. HCN4) in specialized lipid rafts on the cell surface called caveolae191,192.
Recently, a stretch of aromatic residues found in the N-terminus of all HCN channels
was shown to be required for the interaction between HCN4 and caveolin-1192,193,194.
Disrupting this interaction decreases the surface expression of HCN4173,174. The
localization of caveolin-1 in the retina has been studied, but the results are
controversial. Berta et al., showed that caveolin-1 is present in the IS of photoreceptors,
while Gu et al., showed that caveolin-1 is absent from photoreceptors and is only found
in Müller cells, a type of retinal glial cells195. Since different caveolin-1 antibodies were
used in those studies, verification of their specificities will be necessary in the future.
111
Trafficking of HCN1 via the Early Secretory Pathways
In Chapter III and IV, we identified two trafficking signals in HCN1 that both
function in the early secretory pathway. The journey of HCN1 starts in the ER where it
needs to be properly folded and assembled (Figure 6-2A, Step 1); therefore, the ER
trafficking motifs in HCN1 have the potential to facilitate one or both of these processes.
HCN1 forms homo-tetramers in the absence of other HCN proteins. We have tested the
behavior of an HCN1 ER export mutant (HCN1VSLmut) in sucrose density gradients and
found no difference in its sedimentation behavior from the wildtype HCN1. This result
suggests that the L-based ER export motif is not involved in either folding or assembly
of HCN1 channels but the subsequent processes which will be discussed later. We
have not tested the role of the ER retention signal in HCN1 folding or assembly using
velocity sedimentation, but it will be a very intriguing experiment to conduct in the future
because the di-arginine ER retention motif was suggested to monitor the subunit
assembly status of complex proteins (e.g. Kir6.2/SUR1).
Mature HCN1 is actively sorted to the ER exit sites and carried by the COPII
complex in the anterograde transport pathway (Figure 6-2A, Step 2). Sec24 proteins in
the COPII complex are responsible for cargo recognition. In human, there are four
paralogs of Sec24 (Sec24A-D) with different cargo selectivity196-198. A transcriptomic
analysis of human retina suggests that Sec24B is the most abundant Sec24 transcript in
the retina199. The amount of Sec24A and Sec24D is less than one third of Sec24B and
Sec24C transcript was not detected. Generally speaking, each Sec24 protein contains
multiple sites for interacting with distinct cargo molecules. So far, four cargo binding
sites (A-, B-, C- and D-site) have been identified in yeast Sec24p.
112
It is intriguing that cargoes with similar ER export motifs respond differently to
disruption of a single binding pocket in Sec24p. For example, Erv41p (IL), Erv46p (FY),
Emp47p (Y…LL) and Erp1p (IL) all contain hydrophobic ER export motifs (the key
residues are indicated in the parentheses) and interact with wildtype Sec24p. When
their interactions with Sec24pL616W (a mutant that inhibits cargo binding to the B-site)
were tested, the binding is not changed for Erv41p/Erv46p, yet decreased for
Emp47p/Erp1p to different degrees162,163. These findings suggest that proteins with
similar ER export motifs can interact with Sec24 at different sites and possibly with
different affinities. Most integral membrane proteins directly recruit Sec24 but some of
them recruit additional adaptors. It was recently shown that coincidental binding of both
the cargo and its receptor to Sec24 increases the efficiency of cargo packing. Although
the L-based motif in HCN1 can be categorized as a hydrophobic ER export motif,
whether and how it interacts with Sec24 is hard to predict without experimental
investigation.
Immature HCN1 channels are either directly retained in the ER or retrieved from
the ERGIC/Golgi compartments (Figure 6-2, Step 3). Interactions between β-COP, a
component of the COPI complex, and di-basic (KK, RR or KR) ER retention motifs have
been reported in several unrelated membrane proteins including Kir6.2, proteinaseactivated receptors 4 (PAR4), neuronal nicotinic acetylcholine receptor (nAChR),
potassium channel subfamily K member 3 (KCNK3), KCNK9 and the lip35 major
histocompatibility antigen class II-associated invariant chain (Table 6-1). A direct
interaction between the di-arginine motif and β-COP was suggested in Kir6.2 using in
vitro binding assays20. Since binding to COPI complexes seems to be shared by many
113
multimeric membrane proteins containing di-basic motifs, it is likely that immature HCN1
is retained, or more specifically retrieved in the ER via the retrograde transport pathway
carried out by COPI complexes.
The subsequent release of those di-basic motif containing proteins from the
ERGIC/Golgi compartment requires exclusion from the COPI coated vesicles (Figure 62, Step 4). A mutually exclusive mechanism via binding to 14-3-3 proteins has been
proposed. 14-3-3 proteins (seven members in mammals) have been reported to
facilitate the forward trafficking of many multimeric membrane proteins (Table 6-1)20,200202.
Several proteins bearing the di-basic ER retention motifs also have binding sites to
14-3-3. Those proteins tend to interact with different 14-3-3 proteins, suggesting a way
to achieve selective protein trafficking. The 14-3-3 binding motif can be either close to
(e.g. in lip35) or distal from (e.g. in KCNK3) the ER retention motif in the protein
sequence. It was shown in multiple cases that the recruitment of 14-3-3 excludes the
previous binding between the di-basic motif and COPI, leading to protein export from
the ERGIC/Golgi. Steric interference is a possible explanation for the exclusive
interaction to COPI or 14-3-3. More structural analysis is needed to test this possibility.
Interestingly, multiple putative 14-3-3 binding sites are present in HCN1 (Figure
6-2B)110. One of them even overlaps with the di-arginine motif, similar to the situation in
KCNK3 and KCNK9 (Table 6-1). Therefore 14-3-3 proteins are potential helpers of
exporting HCN1 to the PM, and this direction will be explored in the future. The
transcriptomic analysis of human retina shows that there are six 14-3-3 proteins (β, ε, γ,
η, θ and ζ) expressed in the retina199. Specifying which 14-3-3 proteins are expressed in
photoreceptors using the laser capture microdissection technique described in Chapter
114
II will be helpful. The most straightforward way to test if 14-3-3 regulates the trafficking
of HCN1 in photoreceptors is to generate 14-3-3 knockout mice followed by analyzing
the localization of HCN1 in the cell; however, multiple 14-3-3 isoforms may be
expressed in photoreceptors and their functions could be redundant. In addition, 14-3-3
proteins are critical for multiple cellular pathways therefore other cellular defects in the
knockout mice may interfere with our study in trafficking. Alternatively, we will explore
this direction by testing the binding between HCN1 and 14-3-3 both in vivo and in vitro,
followed by studying trafficking of specifically designed HCN1 mutants that cannot
interact with 14-3-3.
The mechanism of how 14-3-3 proteins are recruited to the properly assembled
proteins is unknown. Phosphorylation was implicated to be the molecular switch
because the binding between 14-3-3 and its partners requires phosphorylation at one
serine residue within the 14-3-3 binding motif (Table 6-1, KCNK3/9, lip35 and nAChR).
But note that in the case of Kir6.2, phosphorylation is not necessary for binding to 14-33, likely due to the amphipathic nature of the ligand binding groove in 14-3-3203,204.
While the requirement of phosphorylation in recruiting 14-3-3 to membrane proteins is
still under debate, the role of phosphorylation and other post-translational modifications
in HCN1 trafficking needs to be considered.
Potential Roles of Post-translational Modifications in HCN1 Trafficking
Other than interacting with adaptors and coat protein complexes, posttranslational modifications (e.g. N-glycosylation and phosphorylation) in HCN1 itself
may serve important roles in its trafficking. HCN1 is glycosylated at an asparagine (N)
115
residue located in the extracellular loop between transmembrane domains 5 and 6. This
modification is seen as a ~10-20 kDa shift in the mobility of HCN1 in Western blots43,45.
The amount of HCN1 glycosylation can possibly serve as an indicator of disease
progression because it was reported that seizure-like activities promote HCN1
glycosylation in hippocampal neurons45. Whether the extent of HCN1 glycosylation
correlates with its surface expression remains unclear.
The glycosylation site in HCN1 is shared by all HCN channels205,206. It was found
that a HCN2 mutant that cannot be glycosylated fails to target the PM of HEK293
cells43. Is glycosylation also necessary for the surface expression of HCN1? We have
obtained contradictory results in two different systems. The result of our biotinylation
assay in the mouse retina shows an increase in the amount of glycosylated HCN1 in the
surface pool, from roughly 10% of the total HCN1 to nearly half (Figure 2-7). In contrast,
when overexpressed in HEK293 cells, the glycosylated HCN1 is hardly observed at the
cell surface using the same surface biotinylation assay (Figure 3-6). Our results
implicate that the importance of glycosylation in HCN1 trafficking may vary dependent
on the cell system studied.
Besides glycosylation, phosphorylation plays important roles in protein trafficking.
Several kinases and phosphatases were reported to modulate the channel properties of
HCN channels. PKA (tested in HCN4)207 and Src (HCN2 and HCN4)208-210 are
considered positive regulators of HCN activity, and the identified phosphorylation sites
are conserved in HCN1. PKC negatively regulates HCN1 activity, though if there is an
interaction between PKC and HCN1 is unclear146,211-213. When expressed in Xenopus
photoreceptors, the C-terminal region of HCN1 containing the putative PKA
116
phosphorylation site (S796 in X. tropicalis HCN1) is not required for its trafficking
(Figure 3-4C), neither is the region containing the putative Src phosphorylation sites
(Y382 and/or Y386) (Figure 3-3C). Yet a role of PKA and Src in HCN1 trafficking cannot
be completely ruled out because the observations were based on the reporter-fused
constructs. A careful investigation of HCN1 phosphorylation both in vivo and in vitro,
followed by testing the influence of phosphorylation on HCN1 trafficking is encouraged
in the future.
Opposite to PKA and Src kinases, two phosphatases were reported to be
negative regulators of HCN1 gating. Calcineurin was shown to contribute to epilepsydependent downregulation of HCN gating, but whether this phosphatase functions
directly on HCN channels has not been explored214. Receptor-like protein tyrosine
phosphatase-α (RPTPα) was shown to reduce the tyrosine phosphorylation and surface
expression of HCN2 when both proteins were both expressed in HEK293 cells215.
Further investigation on the interaction between RPTPα and HCN1 is required, as well
as the role of RPTPα in photoreceptors.
Role of Ankyrin in the Retinal Localization of NKA
In Chapter V, we studied how NKA trafficking is regulated in photoreceptors. We
studied localization of ankyrin proteins that were shown to regulate NKA trafficking in
some other cell types. However, in photoreceptors we only detected AnkB and AnkR in
retinal synaptic terminals from which NKA is excluded. We did observe some interaction
between AnkB and NKA in the whole retina lysate. This observation likely reflects an
interaction coming from the synaptic compartments of other retinal neurons. In fact,
117
multiple studies have implicated roles for ankyrin in synaptic biology. AnkB is required
for the assembly, excitability and morphology of the neuromuscular junction in
Drosophila216-218. AnkG has been found to have a role in shaping dendritic spines, which
are protrusions along the dendrite forming synapses with other neurons219,220.
In the retina, the two ankyrin proteins we investigated (AnkB and AnkR) label
both synaptic layers (i.e. the OPL and IPL). The staining intensity is greater in the OPL
which contains the photoreceptor to bipolar cell synapses. NKA is excluded from the
OPL upper strata which contains presynaptic terminals from photoreceptors. In the OPL
lower strata containing the dendrites from bipolar cells, AnkB and NKA co-localization
can be observed. AnkR and NKA do not co-localize in the OPL as AnkR is only seen on
the photoreceptor side of this synaptic layer. Future studies will hopefully elucidate the
contributions synaptic ankyrins make to vision.
Trafficking of NKAα to the ISPM of Photoreceptors
We investigated trafficking of two NKAα isozymes, the neuron-specific NKAα3
and the sperm-specific NKAα4 in Chapter V. When expressed in Xenopus
photoreceptors, NKAα3 and NKAα4 were targeted to the ISPM and OS respectively.
We successfully identified a VxP OS/ciliary targeting motif in the N-terminus of NKAα4.
This motif is the reason why NKAα4 is targeted to the OS, because mutating either or
both of the critical residues (V and P) inside the motif relocates NKAα4 from the OS to
the ISPM. This work expands the repertoire of potential mechanisms contributing to
differential sub-cellular compartmentalization of NKA isozymes.
118
One important remaining question is what controls the targeting of NKAα3 to the
ISPM. NKAβ was once thought to be required for the sub-cellular compartmentalization
of NKAα based on the observation that exogenous NKAβ2 localizes to the apical
surface, in contrast to the basolateral localization of endogenous NKAβ1 in MDCK
cells221. However, later research showed that the apical localization of NKAβ2 is due to
the presence of butyrate, a supplemental chemical in the growth media222. An additional
role of NKAβ in the photoreceptor is to interact with retinoschisin, a secreted protein
important for maintaining retinal organization73. However in the extracellular space
surrounding photoreceptors, retinoschisin is also found in the synaptic layer where NKA
is excluded, suggesting that the ISPM localization of NKA is unlikely due to the binding
between the β subunit and retinoschisin.
The fact that the NKAα4 mutant with a non-functional OS trafficking motif
localizes to the ISPM implicates that there is a secondary ISPM targeting signal present
in NKAα4. Moreover, exogenously expressed NKAα1 also localizes to the ISPM similar
to NKAα3 (data not shown). It is possible that the ISPM targeting signal is located in
conserved regions across these NKAα proteins. The NKAα isozymes share a high
degree of sequence similarity; therefore other clues are needed to predict regions
containing the ISPM targeting signal. From our experience as well as others’, most
trafficking signals are located in regions with high disorder tendency. We thus analyzed
the conservation and disorder tendency of NKAα1, α3 and α4, based on the crystal
structure of NKAα177,78,223. The analysis suggests to us three regions that may contain
119
the ISPM targeting signal of NKAα3 (Figure 6-3). We will generate specific mutations in
those regions and study NKAα3 localization in Xenopus photoreceptors in the future.
Conclusions
In summary, we investigated the trafficking of HCN1 and NKA in photoreceptors
using mouse and Xenopus as the animal models. So far, this work provides the only
experimental evidence of trafficking mechanisms regulating the localization of IS
membrane proteins. It is anticipated that our findings will stimulate additional work so
that a comprehensive picture can eventually be built to describe how this process
contributes to the health of photoreceptors. By studying HCN1 localization in TRIP8b
knockout mice, we found that TRIP8b is not necessary for supporting HCN1 trafficking
in this specific cell type. Later, by studying localization of HCN1 mutants in transgenic
Xenopus photoreceptors, we discovered two novel trafficking signals (a di-arginine ER
retention signal and a L-based ER export signal) in HCN1. These two counteracting
signals function together to control HCN1 trafficking along the early secretory pathways
and maintain a balance of HCN1 amounts between the intracellular space and the
plasma membrane.
To resolve what controls the specific localization of NKAα subunits, we first
tested ankyrins as candidates but found no co-localization between ankyrins and NKA
in photoreceptors. Subsequently, we investigated why targeting of NKAα3 and NKAα4
is different in photoreceptors. We found that their trafficking is differently regulated
partially due to a VxP ciliary targeting motif present in NKAα4. It is a motif that drives
120
NKAα4 to the ciliary compartment of the photoreceptor and we propose that this motif is
also the determinant of NKAα4 localization in the sperm.
Since the 1980s, studies in membrane protein trafficking in photoreceptors have
been focused on the OS exclusively. The IS resident membrane proteins were
overlooked for decades yet they carry out fundamental functions to maintain the
activities of photoreceptors. Since specific trafficking signals are required to support the
functions of IS resident membrane proteins, our study served as the first step to
uncover the underlining mechanisms. Our results revealed a shared mechanism
controlling membrane protein trafficking in early secretory pathways. The di-basic ER
retention motif we identified in HCN1, previously identified in NKA85, as well as many
other membrane proteins (Table 6-1) suggest the role of di-basic ER retention motifs in
retaining immature membrane proteins in the ER. This finding expands our current
knowledge about the trafficking control system that retains immature proteins in the ER.
Unlike the early secretory pathways where most mechanisms are shared by distinct
membrane proteins, the mechanism that guides mature membrane protein complexes
out of the early secretory pathway varies. For example, the VxP motif in NKAα4 guides
its post-Golgi trafficking to the cilium. In the case of HCN1, ER exit is guided by a Lbased motif we characterized; however, regulators involved in post-Golgi trafficking of
HCN1 are not clear. Identification of specific trafficking regulators will help develop
therapeutic tools that target genetic mutations identified in the future in HCN1 that
results in trafficking defects. We proposed a role of 14-3-3 proteins in guiding HCN1
from the Golgi to the ISPM in photoreceptors. This direction will be explored in the
future.
121
Protein
di-basic ER
retention motif
COPI
interaction
14-3-3 binding
motif
14-3-3
binding
exclusive
binding?
Ref
#
PAR4
HPLRARAL (L)
yes
not known
14-3-3ζ
yes
224
KCNK3
MKRQNVAC
(NT)
yes
LMKRRSSPV
(CT)
14-3-3β
yes
225
KCNK9
MKRQNVAC
(NT)
yes
LMKRRKSPV
(CT)
14-3-3β
yes
225
lip35
MHRRRSRSC
(NT)
yes
RRRSRSPCRE
(NT)
14-3-3β
yes
160,
nAChR
TWVRRVFLDI
(L)
yes
KARSLSPVQH
(L)
14-3-3β/η
not known
26
Kir6.2
PLRKRSVA
(CT)
yes*
PLRKRSVA
(CT)
14-3-3ζ/ε
yes
20
161
225,2
* direct interaction between the di-basic motif and β-COP reported
Table 6-1. A summary of interactions between di-basic ER retention motifs and
COPI or 14-3-3 proteins
Previous reports on interactions between the di-basic ER retention motif and COPI or
14-3-3 proteins are summarized. Residues required for the interaction are underlined.
Whether the residues are located in the N-terminus (NT), intracellular loop (L), or Cterminus (CT) is indicated. Exclusive binding indicates whether binding to COPI and 143-3 is exclusive for each di-basic motif containing protein. Abbreviations: PAR4,
proteinase-activated receptors 4; nAChR, neuronal nicotinic acetylcholine receptor; Ref
#, reference number.
122
Figure 6-1. A model for photoreceptor trafficking of HCN1 and NKA
An enlarged view of the photoreceptor (left) inner segments (IS) containing the
endoplasmic reticulum (ER) and the ER Golgi intermediate compartment (ERGIC)/
Golgi is shown (right). Processes of HCN1 trafficking from the ER to the ERGIC/Golgi,
NKAα3 trafficking to the IS plasma membrane (ISPM) and NKAα4 targeting to the outer
segments (OS) are indicated. Trafficking regulators and signals studied in the thesis are
indicated for those trafficking events. Black boxes outline the proteins that are unlikely
to regulate trafficking of HCN1 or NKAα3. Green boxes outline positive trafficking
signals and the red box outlines the negative trafficking signal.
123
Figure 6-2. A proposed model for HCN1 trafficking via early secretory pathways
A) A summary of the four major steps of membrane protein trafficking in the early
secretory pathways using HCN1 as an example. Each cube represents one monomer of
HCN1 that contains an ER export (green) and an ER retention (red) signals. The
putative role of 14-3-3 (blue) in counteracting the COPI mediated retrograde transport is
indicated in Step 4. B) The sequence and position of the two ER trafficking signals in
the mouse HCN1 are shown (highlighted and underlined, the key residues are bolded).
The putative 14-3-3 binding motifs which are conserved across species are colored
blue. Note that one 14-3-3 binding site partially overlaps with the ER retention signal.
The 14-3-3 binding sites were predicted by PROSITE (ELM) based on the motif pattern
R..[^P]([ST])[IVLM] and [RHK][STALV].([ST]).[PESRDIFTQ]. Abbreviations: PM, plasma
membrane; ERGIC, ER Golgi intermediate compartment; ER, endoplasmic reticulum;
COP, coat protein; NT, N-terminus; CT, C-terminus.
124
Figure 6-3. Regions of NKAα
α3 with putative ISPM trafficking signals
A predicted structure of mouse NKAα3 based on the crystal structure from Squalus
acanthias NKA78. The three cytoplasmic catalytic domains (A, N, and P) are indicated.
Regions that may contain ISPM trafficking signals are shown in green along with the
corresponding amino acids. Abbreviations: PM, plasma membrane.
125
REFERENCES
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Pelucchi, B., Grimaldi, A. & Moriondo, A. Vertebrate rod photoreceptors express
both BK and IK calcium-activated potassium channels, but only BK channels are
involved in receptor potential regulation. Journal of neuroscience research 86,
194-201, doi:10.1002/jnr.21467 (2008).
Murphy, M. J. & Crewther, S. G. Ouabain inhibition of Na/K-ATPase across the
retina prevents signed refractive compensation to lens-induced defocus, but not
default ocular growth in young chicks. F1000Research 2, 97,
doi:10.12688/f1000research.2-97.v1 (2013).
Barrow, A. J. & Wu, S. M. Low-conductance HCN1 ion channels augment the
frequency response of rod and cone photoreceptors. J Neurosci 29, 5841-5853,
doi:10.1523/JNEUROSCI.5746-08.2009 (2009).
Seeliger, M. W. et al. Modulation of rod photoreceptor output by HCN1 channels
is essential for regular mesopic cone vision. Nat Commun 2, 532,
doi:10.1038/ncomms1540 (2011).
Knop, G. C. et al. Light responses in the mouse retina are prolonged upon
targeted deletion of the HCN1 channel gene. Eur J Neurosci 28, 2221-2230,
doi:10.1111/j.1460-9568.2008.06512.x (2008).
Cervetto, L., Demontis, G. C. & Gargini, C. Cellular mechanisms underlying the
pharmacological induction of phosphenes. Br J Pharmacol 150, 383-390,
doi:10.1038/sj.bjp.0706998 (2007).
Wissinger, B. et al. Cone dystrophy with supernormal rod response is strictly
associated with mutations in KCNV2. Invest Ophthalmol Vis Sci 49, 751-757,
doi:10.1167/iovs.07-0471 (2008).
Barlowe, C. Signals for COPII-dependent export from the ER: what's the ticket
out? Trends in cell biology 13, 295-300 (2003).
Venditti, R., Wilson, C. & De Matteis, M. A. Exiting the ER: what we know and
what we don't. Trends in cell biology 24, 9-18, doi:10.1016/j.tcb.2013.08.005
(2014).
Jackson, L. P. Structure and mechanism of COPI vesicle biogenesis. Current
opinion in cell biology 29, 67-73, doi:10.1016/j.ceb.2014.04.009 (2014).
Nishimura, N. & Balch, W. E. A di-acidic signal required for selective export from
the endoplasmic reticulum. Science 277, 556-558 (1997).
Dominguez, M. et al. gp25L/emp24/p24 protein family members of the cis-Golgi
network bind both COP I and II coatomer. J Cell Biol 140, 751-765 (1998).
Fiedler, K., Veit, M., Stamnes, M. A. & Rothman, J. E. Bimodal interaction of
coatomer with the p24 family of putative cargo receptors. Science 273, 13961399 (1996).
Munro, S. & Pelham, H. R. A C-terminal signal prevents secretion of luminal ER
proteins. Cell 48, 899-907 (1987).
Capitani, M. & Sallese, M. The KDEL receptor: new functions for an old protein.
FEBS letters 583, 3863-3871, doi:10.1016/j.febslet.2009.10.053 (2009).
126
16
17
18
19
20
21
22
23
24
25
26
27
28
Beck, R., Rawet, M., Wieland, F. T. & Cassel, D. The COPI system: molecular
mechanisms and function. FEBS letters 583, 2701-2709,
doi:10.1016/j.febslet.2009.07.032 (2009).
Teasdale, R. D. & Jackson, M. R. Signal-mediated sorting of membrane proteins
between the endoplasmic reticulum and the golgi apparatus. Annual review of
cell and developmental biology 12, 27-54, doi:10.1146/annurev.cellbio.12.1.27
(1996).
Michelsen, K., Yuan, H. & Schwappach, B. Hide and run. Arginine-based
endoplasmic-reticulum-sorting motifs in the assembly of heteromultimeric
membrane proteins. EMBO reports 6, 717-722, doi:10.1038/sj.embor.7400480
(2005).
Zerangue, N., Schwappach, B., Jan, Y. N. & Jan, L. Y. A new ER trafficking
signal regulates the subunit stoichiometry of plasma membrane K-ATP channels.
Neuron 22, 537-548, doi:10.1016/S0896-6273(00)80708-4 (1999).
Yuan, H., Michelsen, K. & Schwappach, B. 14-3-3 dimers probe the assembly
status of multimeric membrane proteins. Current biology : CB 13, 638-646
(2003).
Wakana, Y. et al. A new class of carriers that transport selective cargo from the
trans Golgi network to the cell surface. The EMBO journal 31, 3976-3990,
doi:10.1038/emboj.2012.235 (2012).
Grant, B. D. & Donaldson, J. G. Pathways and mechanisms of endocytic
recycling. Nature reviews. Molecular cell biology 10, 597-608,
doi:10.1038/nrm2755 (2009).
Lee, E. S. & Flannery, J. G. Transport of truncated rhodopsin and its effects on
rod function and degeneration. Invest Ophthalmol Vis Sci 48, 2868-2876,
doi:10.1167/iovs.06-0035 (2007).
Tam, B. M., Moritz, O. L., Hurd, L. B. & Papermaster, D. S. Identification of an
outer segment targeting signal in the COOH terminus of rhodopsin using
transgenic Xenopus laevis. J Cell Biol 151, 1369-1380,
doi:10.1083/jcb.151.7.1369 (2000).
Baker, S. A. et al. The outer segment serves as a default destination for the
trafficking of membrane proteins in photoreceptors. J Cell Biol 183, 485-498,
doi:10.1083/jcb.200806009 (2008).
Luo, W., Marsh-Armstrong, N., Rattner, A. & Nathans, J. An outer segment
localization signal at the C terminus of the photoreceptor-specific retinol
dehydrogenase. The Journal of neuroscience : the official journal of the Society
for Neuroscience 24, 2623-2632, doi:10.1523/JNEUROSCI.5302-03.2004
(2004).
Pearring, J. N., Lieu, E. C., Winter, J. R., Baker, S. A. & Arshavsky, V. Y. R9AP
targeting to rod outer segments is independent of rhodopsin and is guided by the
SNARE homology domain. Molecular biology of the cell 25, 2644-2649,
doi:10.1091/mbc.E14-02-0747 (2014).
Pearring, J. N., Salinas, R. Y., Baker, S. A. & Arshavsky, V. Y. Protein sorting,
targeting and trafficking in photoreceptor cells. Progress in retinal and eye
research 36, 24-51, doi:10.1016/j.preteyeres.2013.03.002 (2013).
127
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
Pape, H. C. Queer current and pacemaker: the hyperpolarization-activated cation
current in neurons. Annu Rev Physiol 58, 299-327,
doi:10.1146/annurev.ph.58.030196.001503 (1996).
Wahl-Schott, C. & Biel, M. HCN channels: structure, cellular regulation and
physiological function. Cellular and molecular life sciences : CMLS 66, 470-494,
doi:10.1007/s00018-008-8525-0 (2009).
Bucchi, A., Barbuti, A., Difrancesco, D. & Baruscotti, M. Funny Current and
Cardiac Rhythm: Insights from HCN Knockout and Transgenic Mouse Models.
Frontiers in physiology 3, 240, doi:10.3389/fphys.2012.00240 (2012).
Santoro, B. & Tibbs, G. R. The HCN gene family: molecular basis of the
hyperpolarization-activated pacemaker channels. Ann N Y Acad Sci 868, 741764 (1999).
Muller, F. et al. HCN channels are expressed differentially in retinal bipolar cells
and concentrated at synaptic terminals. Eur J Neurosci 17, 2084-2096 (2003).
Demontis, G. C. et al. Functional characterisation and subcellular localisation of
HCN1 channels in rabbit retinal rod photoreceptors. J Physiol 542, 89-97 (2002).
Fyk-Kolodziej, B. & Pourcho, R. G. Differential distribution of hyperpolarizationactivated and cyclic nucleotide-gated channels in cone bipolar cells of the rat
retina. The Journal of comparative neurology 501, 891-903,
doi:10.1002/cne.21287 (2007).
Della Santina, L. et al. Processing of retinal signals in normal and HCN deficient
mice. PloS one 7, e29812, doi:10.1371/journal.pone.0029812 (2012).
Oi, H., Partida, G. J., Lee, S. C. & Ishida, A. T. HCN4-like immunoreactivity in rat
retinal ganglion cells. Vis Neurosci 25, 95-102,
doi:10.1017/S095252380808005X (2008).
Partida, G. J., Stradleigh, T. W., Ogata, G., Godzdanker, I. & Ishida, A. T. Thy1
associates with the cation channel subunit HCN4 in adult rat retina. Investigative
ophthalmology & visual science 53, 1696-1703, doi:10.1167/iovs.11-9307 (2012).
Stradleigh, T. W. et al. Colocalization of hyperpolarization-activated, cyclic
nucleotide-gated channel subunits in rat retinal ganglion cells. The Journal of
comparative neurology 519, 2546-2573, doi:10.1002/cne.22638 (2011).
Koizumi, A., Jakobs, T. C. & Masland, R. H. Inward rectifying currents stabilize
the membrane potential in dendrites of mouse amacrine cells: patch-clamp
recordings and single-cell RT-PCR. Mol Vis 10, 328-340 (2004).
Ivanova, E. & Muller, F. Retinal bipolar cell types differ in their inventory of ion
channels. Vis Neurosci 23, 143-154, doi:10.1017/S0952523806232048 (2006).
Kim, I. B., Lee, E. J., Kang, T. H., Chung, J. W. & Chun, M. H. Morphological
analysis of the hyperpolarization-activated cyclic nucleotide-gated cation channel
1 (HCN1) immunoreactive bipolar cells in the rabbit retina. The Journal of
comparative neurology 467, 389-402, doi:10.1002/cne.10957 (2003).
Much, B. et al. Role of subunit heteromerization and N-linked glycosylation in the
formation of functional hyperpolarization-activated cyclic nucleotide-gated
channels. J Biol Chem 278, 43781-43786, doi:10.1074/jbc.M306958200 (2003).
Brewster, A. L., Bernard, J. A., Gall, C. M. & Baram, T. Z. Formation of
heteromeric hyperpolarization-activated cyclic nucleotide-gated (HCN) channels
128
45
46
47
48
49
50
51
52
53
54
55
56
57
in the hippocampus is regulated by developmental seizures. Neurobiology of
disease 19, 200-207, doi:10.1016/j.nbd.2004.12.015 (2005).
Zha, Q., Brewster, A. L., Richichi, C., Bender, R. A. & Baram, T. Z. Activitydependent heteromerization of the hyperpolarization-activated, cyclic-nucleotide
gated (HCN) channels: role of N-linked glycosylation. Journal of neurochemistry
105, 68-77, doi:10.1111/j.1471-4159.2007.05110.x (2008).
Moosmang, S. et al. Cellular expression and functional characterization of four
hyperpolarization-activated pacemaker channels in cardiac and neuronal tissues.
Eur J Biochem 268, 1646-1652 (2001).
Santoro, B. et al. Molecular and functional heterogeneity of hyperpolarizationactivated pacemaker channels in the mouse CNS. J Neurosci 20, 5264-5275
(2000).
Parton, R. G. & del Pozo, M. A. Caveolae as plasma membrane sensors,
protectors and organizers. Nature reviews. Molecular cell biology 14, 98-112,
doi:10.1038/nrm3512 (2013).
Nakamura, F., Stossel, T. P. & Hartwig, J. H. The filamins: organizers of cell
structure and function. Cell adhesion & migration 5, 160-169 (2011).
Noam, Y. et al. Filamin A promotes dynamin-dependent internalization of
hyperpolarization-activated cyclic nucleotide-gated type 1 (HCN1) channels and
restricts Ih in hippocampal neurons. The Journal of biological chemistry 289,
5889-5903, doi:10.1074/jbc.M113.522060 (2014).
Wilkars, W. et al. Nedd4-2 regulates surface expression and may affect Nglycosylation of hyperpolarization-activated cyclic nucleotide-gated (HCN)-1
channels. FASEB journal : official publication of the Federation of American
Societies for Experimental Biology 28, 2177-2190, doi:10.1096/fj.13-242032
(2014).
Sung, C. H. & Chuang, J. Z. The cell biology of vision. J Cell Biol 190, 953-963,
doi:10.1083/jcb.201006020 (2010).
Skiba, N. P. et al. Proteomic identification of unique photoreceptor disc
components reveals the presence of PRCD, a protein linked to retinal
degeneration. Journal of proteome research 12, 3010-3018,
doi:10.1021/pr4003678 (2013).
Harosi, F. I. Absorption spectra and linear dichroism of some amphibian
photoreceptors. The Journal of general physiology 66, 357-382 (1975).
Harosi, F. I. Cynomolgus and rhesus monkey visual pigments. Application of
Fourier transform smoothing and statistical techniques to the determination of
spectral parameters. The Journal of general physiology 89, 717-743 (1987).
Scott, S. P., Shea, P. W. & Dryer, S. E. Mapping ligand interactions with the
hyperpolarization activated cyclic nucleotide modulated (HCN) ion channel
binding domain using a soluble construct. Biochemistry 46, 9417-9431,
doi:10.1021/bi6026049 (2007).
Zagotta, W. N. et al. Structural basis for modulation and agonist specificity of
HCN pacemaker channels. Nature 425, 200-205, doi:10.1038/nature01922
(2003).
129
58
59
60
61
62
63
64
65
66
67
68
69
70
Ludwig, A., Zong, X., Jeglitsch, M., Hofmann, F. & Biel, M. A family of
hyperpolarization-activated mammalian cation channels. Nature 393, 587-591,
doi:10.1038/31255 (1998).
Traverso, V., Bush, R. A., Sieving, P. A. & Deretic, D. Retinal cAMP levels during
the progression of retinal degeneration in rhodopsin P23H and S334ter
transgenic rats. Invest Ophthalmol Vis Sci 43, 1655-1661 (2002).
Gordon, S. E. & Zagotta, W. N. Localization of regions affecting an allosteric
transition in cyclic nucleotide-activated channels. Neuron 14, 857-864 (1995).
Ramakrishnan, N. A., Drescher, M. J., Khan, K. M., Hatfield, J. S. & Drescher, D.
G. HCN1 and HCN2 proteins are expressed in cochlear hair cells: HCN1 can
form a ternary complex with protocadherin 15 CD3 and F-actin-binding filamin A
or can interact with HCN2. J Biol Chem 287, 37628-37646,
doi:10.1074/jbc.M112.375832 (2012).
Yi, E., Roux, I. & Glowatzki, E. Dendritic HCN channels shape excitatory
postsynaptic potentials at the inner hair cell afferent synapse in the mammalian
cochlea. J Neurophysiol 103, 2532-2543, doi:10.1152/jn.00506.2009 (2010).
Acosta, C. et al. HCN1 and HCN2 in Rat DRG neurons: levels in nociceptors and
non-nociceptors, NT3-dependence and influence of CFA-induced skin
inflammation on HCN2 and NT3 expression. PloS one 7, e50442,
doi:10.1371/journal.pone.0050442 (2012).
Santoro, B., Wainger, B. J. & Siegelbaum, S. A. Regulation of HCN channel
surface expression by a novel C-terminal protein-protein interaction. J Neurosci
24, 10750-10762, doi:10.1523/JNEUROSCI.3300-04.2004 (2004).
Wilkars, W. et al. Regulation of axonal HCN1 trafficking in perforant path involves
expression of specific TRIP8b isoforms. PloS one 7, e32181,
doi:10.1371/journal.pone.0032181 (2012).
Huang, Z. et al. TRIP8b-independent trafficking and plasticity of adult cortical
presynaptic HCN1 channels. The Journal of neuroscience : the official journal of
the Society for Neuroscience 32, 14835-14848, doi:10.1523/JNEUROSCI.154412.2012 (2012).
Lewis, A. S. et al. Deletion of the hyperpolarization-activated cyclic nucleotidegated channel auxiliary subunit TRIP8b impairs hippocampal Ih localization and
function and promotes antidepressant behavior in mice. J Neurosci 31, 74247440, doi:10.1523/JNEUROSCI.0936-11.2011 (2011).
Lewis, A. S. & Chetkovich, D. M. HCN channels in behavior and neurological
disease: too hyper or not active enough? Molecular and cellular neurosciences
46, 357-367, doi:10.1016/j.mcn.2010.11.007 (2011).
Reinhard, L., Tidow, H., Clausen, M. J. & Nissen, P. Na(+),K (+)-ATPase as a
docking station: protein-protein complexes of the Na(+),K (+)-ATPase. Cellular
and molecular life sciences : CMLS 70, 205-222, doi:10.1007/s00018-012-10399 (2013).
Xie, J. X., Shapiro, A. P. & Shapiro, J. I. The Trade-Off between Dietary Salt and
Cardiovascular Disease; A Role for Na/K-ATPase Signaling? Frontiers in
endocrinology 5, 97, doi:10.3389/fendo.2014.00097 (2014).
130
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
Hilgenberg, L. G., Su, H., Gu, H., O'Dowd, D. K. & Smith, M. A. Alpha3Na+/K+ATPase is a neuronal receptor for agrin. Cell 125, 359-369,
doi:10.1016/j.cell.2006.01.052 (2006).
Friedrich, U. et al. The Na/K-ATPase is obligatory for membrane anchorage of
retinoschisin, the protein involved in the pathogenesis of X-linked juvenile
retinoschisis. Human molecular genetics 20, 1132-1142,
doi:10.1093/hmg/ddq557 (2011).
Molday, L. L., Wu, W. W. & Molday, R. S. Retinoschisin (RS1), the protein
encoded by the X-linked retinoschisis gene, is anchored to the surface of retinal
photoreceptor and bipolar cells through its interactions with a Na/K ATPaseSARM1 complex. The Journal of biological chemistry 282, 32792-32801,
doi:10.1074/jbc.M706321200 (2007).
Molday, R. S., Kellner, U. & Weber, B. H. X-linked juvenile retinoschisis: clinical
diagnosis, genetic analysis, and molecular mechanisms. Progress in retinal and
eye research 31, 195-212, doi:10.1016/j.preteyeres.2011.12.002 (2012).
Jorgensen, P. L., Hakansson, K. O. & Karlish, S. J. Structure and mechanism of
Na,K-ATPase: functional sites and their interactions. Annu Rev Physiol 65, 817849, doi:10.1146/annurev.physiol.65.092101.142558 (2003).
Morth, J. P. et al. Crystal structure of the sodium-potassium pump. Nature 450,
1043-1049, doi:10.1038/nature06419 (2007).
Toyoshima, C. & Cornelius, F. New crystal structures of PII-type ATPases:
excitement continues. Current opinion in structural biology 23, 507-514,
doi:10.1016/j.sbi.2013.06.005 (2013).
Toyoshima, C., Kanai, R. & Cornelius, F. First crystal structures of Na+,K+ATPase: new light on the oldest ion pump. Structure (London, England : 1993)
19, 1732-1738, doi:10.1016/j.str.2011.10.016 (2011).
Herrera, V. L., Emanuel, J. R., Ruiz-Opazo, N., Levenson, R. & Nadal-Ginard, B.
Three differentially expressed Na,K-ATPase alpha subunit isoforms: structural
and functional implications. J Cell Biol 105, 1855-1865 (1987).
Blanco, G. Na,K-ATPase subunit heterogeneity as a mechanism for tissuespecific ion regulation. Seminars in nephrology 25, 292-303,
doi:10.1016/j.semnephrol.2005.03.004 (2005).
Orlowski, J. & Lingrel, J. B. Tissue-specific and developmental regulation of rat
Na,K-ATPase catalytic alpha isoform and beta subunit mRNAs. The Journal of
biological chemistry 263, 10436-10442 (1988).
Sweadner, K. J. Isozymes of the Na+/K+-ATPase. Biochim Biophys Acta 988,
185-220 (1989).
Gritz, S. M. & Radcliffe, R. A. Genetic effects of ATP1A2 in familial hemiplegic
migraine type II and animal models. Human genomics 7, 8, doi:10.1186/14797364-7-8 (2013).
Dobretsov, M. & Stimers, J. R. Neuronal function and alpha3 isoform of the Na/KATPase. Frontiers in bioscience : a journal and virtual library 10, 2373-2396
(2005).
Jimenez, T., Sanchez, G., Wertheimer, E. & Blanco, G. Activity of the Na,KATPase alpha4 isoform is important for membrane potential, intracellular Ca2+,
131
86
87
88
89
90
91
92
93
94
95
96
97
98
and pH to maintain motility in rat spermatozoa. Reproduction 139, 835-845,
doi:10.1530/rep-09-0495 (2010).
Woo, A. L., James, P. F. & Lingrel, J. B. Roles of the Na,K-ATPase alpha4
isoform and the Na+/H+ exchanger in sperm motility. Molecular reproduction and
development 62, 348-356, doi:10.1002/mrd.90002 (2002).
Wagoner, K., Sanchez, G., Nguyen, A. N., Enders, G. C. & Blanco, G. Different
expression and activity of the alpha1 and alpha4 isoforms of the Na,K-ATPase
during rat male germ cell ontogeny. Reproduction 130, 627-641,
doi:10.1530/rep.1.00806 (2005).
Jimenez, T., McDermott, J. P., Sanchez, G. & Blanco, G. Na,K-ATPase alpha4
isoform is essential for sperm fertility. Proc Natl Acad Sci U S A 108, 644-649,
doi:10.1073/pnas.1016902108 (2011).
Dunbar, L. A. & Caplan, M. J. The cell biology of ion pumps: sorting and
regulation. European journal of cell biology 79, 557-563, doi:10.1078/0171-933500079 (2000).
Hlivko, J. T., Chakraborty, S., Hlivko, T. J., Sengupta, A. & James, P. F. The
human Na,K-ATPase alpha 4 isoform is a ouabain-sensitive alpha isoform that is
expressed in sperm. Molecular reproduction and development 73, 101-115,
doi:10.1002/mrd.20383 (2006).
Miller, M. R., Mansell, S. A., Meyers, S. A. & Lishko, P. V. Flagellar ion channels
of sperm: similarities and differences between species. Cell calcium,
doi:10.1016/j.ceca.2014.10.009 (2014).
Okawa, H., Sampath, A. P., Laughlin, S. B. & Fain, G. L. ATP consumption by
mammalian rod photoreceptors in darkness and in light. Current biology : CB 18,
1917-1921, doi:10.1016/j.cub.2008.10.029 (2008).
Chen, H. & Chan, D. C. Mitochondrial dynamics--fusion, fission, movement, and
mitophagy--in neurodegenerative diseases. Human molecular genetics 18, R169176, doi:10.1093/hmg/ddp326 (2009).
Schwarz, T. L. Mitochondrial trafficking in neurons. Cold Spring Harbor
perspectives in biology 5, doi:10.1101/cshperspect.a011304 (2013).
Linton, J. D. et al. Flow of energy in the outer retina in darkness and in light.
Proceedings of the National Academy of Sciences of the United States of
America 107, 8599-8604, doi:10.1073/pnas.1002471107 (2010).
Tokhtaeva, E., Sachs, G. & Vagin, O. Assembly with the Na,K-ATPase alpha(1)
subunit is required for export of beta(1) and beta(2) subunits from the
endoplasmic reticulum. Biochemistry 48, 11421-11431, doi:10.1021/bi901438z
(2009).
Ackermann, U. & Geering, K. Mutual dependence of Na,K-ATPase alpha- and
beta-subunits for correct posttranslational processing and intracellular transport.
FEBS letters 269, 105-108 (1990).
Morton, M. J. et al. Association with {beta}-COP regulates the trafficking of the
newly synthesized Na,K-ATPase. The Journal of biological chemistry 285,
33737-33746, doi:10.1074/jbc.M110.141119 (2010).
132
99
100
101
102
103
104
105
106
107
108
109
110
111
112
Devarajan, P., Scaramuzzino, D. A. & Morrow, J. S. Ankyrin binds to two distinct
cytoplasmic domains of Na,K-ATPase alpha subunit. Proc Natl Acad Sci U S A
91, 2965-2969 (1994).
Jordan, C., Puschel, B., Koob, R. & Drenckhahn, D. Identification of a binding
motif for ankyrin on the alpha-subunit of Na+,K(+)-ATPase. The Journal of
biological chemistry 270, 29971-29975 (1995).
Nelson, W. J. & Veshnock, P. J. Ankyrin binding to (Na+ + K+)ATPase and
implications for the organization of membrane domains in polarized cells. Nature
328, 533-536, doi:10.1038/328533a0 (1987).
Cunha, S. R. et al. Defects in Ankyrin-Based Membrane Protein Targeting
Pathways Underlie Atrial Fibrillation. Circulation,
doi:10.1161/CIRCULATIONAHA.111.023986 (2011).
Li, J., Kline, C. F., Hund, T. J., Anderson, M. E. & Mohler, P. J. Ankyrin-B
regulates Kir6.2 membrane expression and function in heart. J Biol Chem 285,
28723-28730, doi:10.1074/jbc.M110.147868 (2010).
Stabach, P. R., Devarajan, P., Stankewich, M. C., Bannykh, S. & Morrow, J. S.
Ankyrin facilitates intracellular trafficking of alpha1-Na+-K+-ATPase in polarized
cells. American journal of physiology. Cell physiology 295, C1202-1214,
doi:10.1152/ajpcell.00273.2008 (2008).
Kline, C. F., Scott, J., Curran, J., Hund, T. J. & Mohler, P. J. Ankyrin-B regulates
Cav2.1 and Cav2.2 channel expression and targeting. The Journal of biological
chemistry 289, 5285-5295, doi:10.1074/jbc.M113.523639 (2014).
Mohler, P. J., Davis, J. Q. & Bennett, V. Ankyrin-B coordinates the Na/K ATPase,
Na/Ca exchanger, and InsP3 receptor in a cardiac T-tubule/SR microdomain.
PLoS biology 3, e423, doi:10.1371/journal.pbio.0030423 (2005).
Kizhatil, K., Sandhu, N. K., Peachey, N. S. & Bennett, V. Ankyrin-B is required for
coordinated expression of beta-2-spectrin, the Na/K-ATPase and the Na/Ca
exchanger in the inner segment of rod photoreceptors. Experimental eye
research 88, 57-64, doi:10.1016/j.exer.2008.09.022 (2009).
Liu, X. et al. Ankyrin B modulates the function of Na,K-ATPase/inositol 1,4,5trisphosphate receptor signaling microdomain. The Journal of biological
chemistry 283, 11461-11468, doi:10.1074/jbc.M706942200 (2008).
Woroniecki, R., Ferdinand, J. R., Morrow, J. S. & Devarajan, P. Dissociation of
spectrin-ankyrin complex as a basis for loss of Na-K-ATPase polarity after
ischemia. American journal of physiology. Renal physiology 284, F358-364,
doi:10.1152/ajprenal.00100.2002 (2003).
Pan, Y. et al. TRIP8b Is Required for Maximal Expression of HCN1 in the Mouse
Retina. PloS one 9, e85850, doi:10.1371/journal.pone.0085850 (2014).
Pan, Y., Laird, J. G., Yamaguchi, D. M. & Baker, S. A. A di-arginine ER retention
signal regulates trafficking of HCN1 channels from the early secretory pathway to
the plasma membrane. Cellular and molecular life sciences : CMLS 72, 833-843,
doi:10.1007/s00018-014-1705-1 (2015).
Santoro, B. et al. TRIP8b splice variants form a family of auxiliary subunits that
regulate gating and trafficking of HCN channels in the brain. Neuron 62, 802-813,
doi:10.1016/j.neuron.2009.05.009 (2009).
133
113
114
115
116
117
118
119
120
121
122
123
124
125
126
Lewis, A. S. et al. Alternatively spliced isoforms of TRIP8b differentially control h
channel trafficking and function. J Neurosci 29, 6250-6265,
doi:10.1523/JNEUROSCI.0856-09.2009 (2009).
Piskorowski, R., Santoro, B. & Siegelbaum, S. A. TRIP8b splice forms act in
concert to regulate the localization and expression of HCN1 channels in CA1
pyramidal neurons. Neuron 70, 495-509, doi:10.1016/j.neuron.2011.03.023
(2011).
Fain, G. L., Quandt, F. N., Bastian, B. L. & Gerschenfeld, H. M. Contribution of a
caesium-sensitive conductance increase to the rod photoresponse. Nature 272,
466-469 (1978).
Baylor, D. A., Matthews, G. & Nunn, B. J. Location and function of voltagesensitive conductances in retinal rods of the salamander, Ambystoma tigrinum. J
Physiol 354, 203-223 (1984).
Hestrin, S. The properties and function of inward rectification in rod
photoreceptors of the tiger salamander. J Physiol 390, 319-333 (1987).
Noam, Y., Bernard, C. & Baram, T. Z. Towards an integrated view of HCN
channel role in epilepsy. Curr Opin Neurobiol, doi:10.1016/j.conb.2011.06.013
(2011).
Santoro, B. et al. TRIP8b regulates HCN1 channel trafficking and gating through
two distinct C-terminal interaction sites. J Neurosci 31, 4074-4086,
doi:10.1523/JNEUROSCI.5707-10.2011 (2011).
Han, Y. et al. Trafficking and gating of hyperpolarization-activated cyclic
nucleotide-gated channels are regulated by interaction with tetratricopeptide
repeat-containing Rab8b-interacting protein (TRIP8b) and cyclic AMP at distinct
sites. J Biol Chem 286, 20823-20834, doi:10.1074/jbc.M111.236125 (2011).
Nolan, M. F. et al. The hyperpolarization-activated HCN1 channel is important for
motor learning and neuronal integration by cerebellar Purkinje cells. Cell 115,
551-564 (2003).
Shin, M., Brager, D., Jaramillo, T. C., Johnston, D. & Chetkovich, D. M.
Mislocalization of h channel subunits underlies h channelopathy in temporal lobe
epilepsy. Neurobiology of disease 32, 26-36, doi:10.1016/j.nbd.2008.06.013
(2008).
Drack, A. V. et al. TUDCA slows retinal degeneration in two different mouse
models of retinitis pigmentosa and prevents obesity in Bardet-Biedl syndrome
type 1 mice. Invest Ophthalmol Vis Sci 53, 100-106, doi:10.1167/iovs.11-8544
(2012).
Marmor, M. F. et al. ISCEV Standard for full-field clinical electroretinography
(2008 update). Documenta ophthalmologica. Advances in ophthalmology 118,
69-77, doi:10.1007/s10633-008-9155-4 (2009).
Braun, A. P. Alternative splicing of TRIP8b diversifies its actions as an accessory
subunit of neuronal HCN channels. Channels 3, 217-218 (2009).
Tanimoto, N., Brombas, A., Muller, F. & Seeliger, M. W. HCN1 channels
significantly shape retinal photoresponses. Advances in experimental medicine
and biology 723, 807-812, doi:10.1007/978-1-4614-0631-0_103 (2012).
134
127
128
129
130
131
132
133
134
135
136
137
138
139
140
Demontis, G. C., Gargini, C., Paoli, T. G. & Cervetto, L. Selective Hcn1 channels
inhibition by ivabradine in mouse rod photoreceptors. Investigative
ophthalmology & visual science 50, 1948-1955, doi:10.1167/iovs.08-2659 (2009).
Savelieva, I. & Camm, A. J. I f inhibition with ivabradine : electrophysiological
effects and safety. Drug safety : an international journal of medical toxicology and
drug experience 31, 95-107 (2008).
Roubille, F. & Tardif, J. C. New therapeutic targets in cardiology: heart failure and
arrhythmia: HCN channels. Circulation 127, 1986-1996,
doi:10.1161/CIRCULATIONAHA.112.000145 (2013).
Kanyshkova, T. et al. Postnatal expression pattern of HCN channel isoforms in
thalamic neurons: relationship to maturation of thalamocortical oscillations. The
Journal of neuroscience : the official journal of the Society for Neuroscience 29,
8847-8857, doi:10.1523/JNEUROSCI.0689-09.2009 (2009).
Lewis, A. S., Estep, C. M. & Chetkovich, D. M. The fast and slow ups and downs
of HCN channel regulation. Channels 4, 215-231, doi:10.4161/chan.4.3.11630
(2010).
Eldstrom, J. & Fedida, D. The voltage-gated channel accessory protein KCNE2:
multiple ion channel partners, multiple ways to long QT syndrome. Expert
reviews in molecular medicine 13, e38, doi:10.1017/S1462399411002092
(2011).
Qu, J. et al. MiRP1 modulates HCN2 channel expression and gating in cardiac
myocytes. J Biol Chem 279, 43497-43502, doi:10.1074/jbc.M405018200 (2004).
Yu, H. et al. MinK-related peptide 1: A beta subunit for the HCN ion channel
subunit family enhances expression and speeds activation. Circ Res 88, E84-87,
doi:10.1161/hh1201.093511 (2001).
Ying, S. W. et al. Targeted deletion of Kcne2 impairs HCN channel function in
mouse thalamocortical circuits. PloS one 7, e42756,
doi:10.1371/journal.pone.0042756 (2012).
Cao-Ehlker, X. et al. Up-regulation of hyperpolarization-activated cyclic
nucleotide-gated channel 3 (HCN3) by specific interaction with K+ channel
tetramerization domain-containing protein 3 (KCTD3). The Journal of biological
chemistry 288, 7580-7589, doi:10.1074/jbc.M112.434803 (2013).
Nieves-Cordones, M. et al. Distinct Amino Acids in the C-Linker Domain of the
Arabidopsis K+ Channel KAT2 Determine Its Subcellular Localization and Activity
at the Plasma Membrane. Plant physiology 164, 1415-1429,
doi:10.1104/pp.113.229757 (2014).
Burns, M. E. & Arshavsky, V. Y. Beyond counting photons: trials and trends in
vertebrate visual transduction. Neuron 48, 387-401,
doi:10.1016/j.neuron.2005.10.014 (2005).
Amaya, E. & Kroll, K. L. A method for generating transgenic frog embryos.
Methods Mol Biol 97, 393-414, doi:1-59259-270-8-393 [pii] 10.1385/1-59259270-8:393 (1999).
Kroll, K. L. & Amaya, E. Transgenic Xenopus embryos from sperm nuclear
transplantations reveal FGF signaling requirements during gastrulation.
Development 122, 3173-3183 (1996).
135
141
142
143
144
145
146
147
148
149
150
151
152
153
Knox, B. E., Schlueter, C., Sanger, B. M., Green, C. B. & Besharse, J. C.
Transgene expression in Xenopus rods. FEBS letters 423, 117-121,
doi:10.1016/S0014-5793(98)00018-0 (1998).
Noam, Y. et al. Trafficking and surface expression of hyperpolarization-activated
cyclic nucleotide-gated channels in hippocampal neurons. J Biol Chem 285,
14724-14736, doi:10.1074/jbc.M109.070391 (2010).
Ishida, T. & Kinoshita, K. Prediction of disordered regions in proteins based on
the meta approach. Bioinformatics 24, 1344-1348,
doi:10.1093/bioinformatics/btn195 (2008).
Chen, S., Wang, J. & Siegelbaum, S. A. Properties of hyperpolarization-activated
pacemaker current defined by coassembly of HCN1 and HCN2 subunits and
basal modulation by cyclic nucleotide. The Journal of general physiology 117,
491-504 (2001).
Bankston, J. R. et al. Structure and stoichiometry of an accessory subunit
TRIP8b interaction with hyperpolarization-activated cyclic nucleotide-gated
channels. Proceedings of the National Academy of Sciences of the United States
of America 109, 7899-7904, doi:10.1073/pnas.1201997109 (2012).
Dinkel, H. et al. The eukaryotic linear motif resource ELM: 10 years and counting.
Nucleic acids research 42, D259-266, doi:10.1093/nar/gkt1047 (2014).
Nava, C. et al. De novo mutations in HCN1 cause early infantile epileptic
encephalopathy. Nature genetics 46, 640-645, doi:10.1038/ng.2952 (2014).
Tang, B., Sander, T., Craven, K. B., Hempelmann, A. & Escayg, A. Mutation
analysis of the hyperpolarization-activated cyclic nucleotide-gated channels
HCN1 and HCN2 in idiopathic generalized epilepsy. Neurobiology of disease 29,
59-70, doi:10.1016/j.nbd.2007.08.006 (2008).
Wang, H. X. & Gao, W. J. Prolonged exposure to NMDAR antagonist induces
cell-type specific changes of glutamatergic receptors in rat prefrontal cortex.
Neuropharmacology 62, 1808-1822, doi:10.1016/j.neuropharm.2011.11.024
(2012).
Gassmann, M. et al. The RXR-type endoplasmic reticulum-retention/retrieval
signal of GABAB1 requires distant spacing from the membrane to function.
Molecular pharmacology 68, 137-144, doi:10.1124/mol.104.010256 (2005).
Scott, D. B., Blanpied, T. A., Swanson, G. T., Zhang, C. & Ehlers, M. D. An
NMDA receptor ER retention signal regulated by phosphorylation and alternative
splicing. The Journal of neuroscience : the official journal of the Society for
Neuroscience 21, 3063-3072 (2001).
Wenthold, R. J., Sans, N., Standley, S., Prybylowski, K. & Petralia, R. S. Early
events in the trafficking of N-methyl-D-aspartate (NMDA) receptors. Biochemical
Society transactions 31, 885-888, doi:10.1042/ (2003).
Shikano, S. & Li, M. Membrane receptor trafficking: evidence of proximal and
distal zones conferred by two independent endoplasmic reticulum localization
signals. Proc Natl Acad Sci U S A 100, 5783-5788,
doi:10.1073/pnas.1031748100 (2003).
136
154
155
156
157
158
159
160
161
162
163
164
165
166
167
Hardel, N., Harmel, N., Zolles, G., Fakler, B. & Klocker, N. Recycling endosomes
supply cardiac pacemaker channels for regulated surface expression.
Cardiovascular research 79, 52-60, doi:10.1093/cvr/cvn062 (2008).
Ramakrishnan, N. A., Drescher, M. J., Khan, K. M., Hatfield, J. S. & Drescher, D.
G. HCN1 and HCN2 are expressed in cochlear hair cells: HCN1 can form a
ternary complex with protocadherin 15 CD3 and F-actin-binding filamin A or can
interact with HCN2. J Biol Chem, doi:10.1074/jbc.M112.375832 (2012).
Reiners, J., Marker, T., Jurgens, K., Reidel, B. & Wolfrum, U. Photoreceptor
expression of the Usher syndrome type 1 protein protocadherin 15 (USH1F) and
its interaction with the scaffold protein harmonin (USH1C). Mol Vis 11, 347-355
(2005).
Sahly, I. et al. Localization of Usher 1 proteins to the photoreceptor calyceal
processes, which are absent from mice. J Cell Biol 199, 381-399,
doi:10.1083/jcb.201202012 (2012).
Salinas, R. Y., Baker, S. A., Gospe, S. M., 3rd & Arshavsky, V. Y. A single valine
residue plays an essential role in peripherin/rds targeting to photoreceptor outer
segments. PloS one 8, e54292, doi:10.1371/journal.pone.0054292 (2013).
Li, D., Takimoto, K. & Levitan, E. S. Surface expression of Kv1 channels is
governed by a C-terminal motif. The Journal of biological chemistry 275, 1159711602 (2000).
Zhu, J., Watanabe, I., Gomez, B. & Thornhill, W. B. Trafficking of Kv1.4
potassium channels: interdependence of a pore region determinant and a
cytoplasmic C-terminal VXXSL determinant in regulating cell-surface trafficking.
Biochem J 375, 761-768, doi:10.1042/BJ20030885 (2003).
D'Arcangelo, J. G., Stahmer, K. R. & Miller, E. A. Vesicle-mediated export from
the ER: COPII coat function and regulation. Biochim Biophys Acta 1833, 24642472, doi:10.1016/j.bbamcr.2013.02.003 (2013).
Miller, E., Antonny, B., Hamamoto, S. & Schekman, R. Cargo selection into
COPII vesicles is driven by the Sec24p subunit. The EMBO journal 21, 61056113 (2002).
Miller, E. A. et al. Multiple cargo binding sites on the COPII subunit Sec24p
ensure capture of diverse membrane proteins into transport vesicles. Cell 114,
497-509 (2003).
Mossessova, E., Bickford, L. C. & Goldberg, J. SNARE selectivity of the COPII
coat. Cell 114, 483-495 (2003).
Mancias, J. D. & Goldberg, J. The transport signal on Sec22 for packaging into
COPII-coated vesicles is a conformational epitope. Molecular cell 26, 403-414,
doi:10.1016/j.molcel.2007.03.017 (2007).
Pagant, S., Wu, A., Edwards, S., Diehl, F. & Miller, E. A. Sec24 Is a Coincidence
Detector that Simultaneously Binds Two Signals to Drive ER Export. Current
biology : CB 25, 403-412, doi:10.1016/j.cub.2014.11.070 (2015).
Powers, J. & Barlowe, C. Erv14p directs a transmembrane secretory protein into
COPII-coated transport vesicles. Molecular biology of the cell 13, 880-891,
doi:10.1091/mbc.01-10-0499 (2002).
137
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
Springer, S. et al. Regulated oligomerization induces uptake of a membrane
protein into COPII vesicles independent of its cytosolic tail. Traffic 15, 531-545,
doi:10.1111/tra.12157 (2014).
Manzano-Lopez, J. et al. COPII Coat Composition Is Actively Regulated by
Luminal Cargo Maturation. Current biology : CB 25, 152-162,
doi:10.1016/j.cub.2014.11.039 (2015).
Vagin, O., Dada, L. A., Tokhtaeva, E. & Sachs, G. The Na-K-ATPase
alpha(1)beta(1) heterodimer as a cell adhesion molecule in epithelia. American
journal of physiology. Cell physiology 302, C1271-1281,
doi:10.1152/ajpcell.00456.2011 (2012).
Cereijido, M., Contreras, R. G., Shoshani, L. & Larre, I. The Na+-K+-ATPase as
self-adhesion molecule and hormone receptor. American journal of physiology.
Cell physiology 302, C473-481, doi:10.1152/ajpcell.00083.2011 (2012).
James, P. F. et al. Identification of a specific role for the Na,K-ATPase alpha 2
isoform as a regulator of calcium in the heart. Molecular cell 3, 555-563 (1999).
Beuschlein, F. et al. Somatic mutations in ATP1A1 and ATP2B3 lead to
aldosterone-producing adenomas and secondary hypertension. Nature genetics
45, 440-444, 444e441-442, doi:10.1038/ng.2550 (2013).
Williams, T. A. et al. Somatic ATP1A1, ATP2B3, and KCNJ5 mutations in
aldosterone-producing adenomas. Hypertension 63, 188-195,
doi:10.1161/hypertensionaha.113.01733 (2014).
Azizan, E. A. et al. Somatic mutations in ATP1A1 and CACNA1D underlie a
common subtype of adrenal hypertension. Nature genetics 45, 1055-1060,
doi:10.1038/ng.2716 (2013).
Heinzen, E. L. et al. Distinct neurological disorders with ATP1A3 mutations. The
Lancet. Neurology 13, 503-514, doi:10.1016/s1474-4422(14)70011-0 (2014).
Sweney, M. T., Newcomb, T. M. & Swoboda, K. J. The Expanding Spectrum of
Neurological Phenotypes in Children With ATP1A3 Mutations, Alternating
Hemiplegia of Childhood, Rapid-onset Dystonia-Parkinsonism, CAPOS and
Beyond. Pediatric neurology, doi:10.1016/j.pediatrneurol.2014.09.015 (2014).
Juhaszova, M. & Blaustein, M. P. Na+ pump low and high ouabain affinity alpha
subunit isoforms are differently distributed in cells. Proc Natl Acad Sci U S A 94,
1800-1805 (1997).
Stoops, E. H. & Caplan, M. J. Trafficking to the apical and basolateral
membranes in polarized epithelial cells. Journal of the American Society of
Nephrology : JASN 25, 1375-1386, doi:10.1681/asn.2013080883 (2014).
Deretic, D., Schmerl, S., Hargrave, P. A., Arendt, A. & McDowell, J. H.
Regulation of sorting and post-Golgi trafficking of rhodopsin by its C-terminal
sequence QVS(A)PA. Proc Natl Acad Sci U S A 95, 10620-10625 (1998).
Mazelova, J. et al. Ciliary targeting motif VxPx directs assembly of a trafficking
module through Arf4. The EMBO journal 28, 183-192,
doi:10.1038/emboj.2008.267 (2009).
Kline, C. F. & Mohler, P. J. Evolving form to fit function: cardiomyocyte
intercalated disc and transverse-tubule membranes. Current topics in
membranes 72, 121-158, doi:10.1016/B978-0-12-417027-8.00004-0 (2013).
138
183
184
185
186
187
188
189
190
191
192
193
194
195
196
Lencesova, L., O'Neill, A., Resneck, W. G., Bloch, R. J. & Blaustein, M. P.
Plasma membrane-cytoskeleton-endoplasmic reticulum complexes in neurons
and astrocytes. The Journal of biological chemistry 279, 2885-2893,
doi:10.1074/jbc.M310365200 (2004).
Kizhatil, K., Baker, S. A., Arshavsky, V. Y. & Bennett, V. Ankyrin-G promotes
cyclic nucleotide-gated channel transport to rod photoreceptor sensory cilia.
Science 323, 1614-1617, doi:10.1126/science.1169789 (2009).
Rosenbaum, J. L. & Witman, G. B. Intraflagellar transport. Nature reviews.
Molecular cell biology 3, 813-825, doi:10.1038/nrm952 (2002).
Pazour, G. J. & Bloodgood, R. A. Targeting proteins to the ciliary membrane.
Current topics in developmental biology 85, 115-149, doi:10.1016/S00702153(08)00805-3 (2008).
Geng, L. et al. Polycystin-2 traffics to cilia independently of polycystin-1 by using
an N-terminal RVxP motif. J Cell Sci 119, 1383-1395, doi:10.1242/jcs.02818
(2006).
Miller, D. & Ostermeier, G. C. Spermatozoal RNA: Why is it there and what does
it do? Gynecologie, obstetrique & fertilite 34, 840-846,
doi:10.1016/j.gyobfe.2006.07.013 (2006).
Tai, A. W., Chuang, J. Z., Bode, C., Wolfrum, U. & Sung, C. H. Rhodopsin's
carboxy-terminal cytoplasmic tail acts as a membrane receptor for cytoplasmic
dynein by binding to the dynein light chain Tctex-1. Cell 97, 877-887 (1999).
Baker, S. A. & Kerov, V. Photoreceptor inner and outer segments. Current topics
in membranes 72, 231-265, doi:10.1016/b978-0-12-417027-8.00007-6 (2013).
Barbuti, A. et al. Localization of pacemaker channels in lipid rafts regulates
channel kinetics. Circ Res 94, 1325-1331,
doi:10.1161/01.RES.0000127621.54132.AE (2004).
Barbuti, A. et al. A caveolin-binding domain in the HCN4 channels mediates
functional interaction with caveolin proteins. J Mol Cell Cardiol 53, 187-195,
doi:10.1016/j.yjmcc.2012.05.013 (2012).
Bosman, A. et al. Molecular and Functional Evidence of HCN4 and Caveolin-3
Interaction During Cardiomyocyte Differentiation from Human Embryonic Stem
Cells. Stem cells and development 22, 1717-1727, doi:10.1089/scd.2012.0247
(2013).
Kikuchi, T. et al. Behavior of caveolae and caveolin-3 during the development of
myocyte hypertrophy. Journal of cardiovascular pharmacology 45, 204-210
(2005).
Berta, A. I., Boesze-Battaglia, K., Magyar, A., Szel, A. & Kiss, A. L. Localization
of caveolin-1 and c-src in mature and differentiating photoreceptors: raft proteins
co-distribute with rhodopsin during development. Journal of molecular histology
42, 523-533, doi:10.1007/s10735-011-9360-4 (2011).
Sucic, S. et al. The serotonin transporter is an exclusive client of the coat protein
complex II (COPII) component SEC24C. The Journal of biological chemistry 286,
16482-16490, doi:10.1074/jbc.M111.230037 (2011).
139
197
198
199
200
201
202
203
204
205
206
207
208
209
210
Wendeler, M. W., Paccaud, J. P. & Hauri, H. P. Role of Sec24 isoforms in
selective export of membrane proteins from the endoplasmic reticulum. EMBO
reports 8, 258-264, doi:10.1038/sj.embor.7400893 (2007).
Mancias, J. D. & Goldberg, J. Structural basis of cargo membrane protein
discrimination by the human COPII coat machinery. The EMBO journal 27, 29182928, doi:10.1038/emboj.2008.208 (2008).
Whitmore, S. S. et al. Transcriptomic analysis across nasal, temporal, and
macular regions of human neural retina and RPE/choroid by RNA-Seq.
Experimental eye research 129, 93-106, doi:10.1016/j.exer.2014.11.001 (2014).
Fu, H., Subramanian, R. R. & Masters, S. C. 14-3-3 proteins: structure, function,
and regulation. Annual review of pharmacology and toxicology 40, 617-647,
doi:10.1146/annurev.pharmtox.40.1.617 (2000).
Nufer, O. & Hauri, H. P. ER export: call 14-3-3. Current biology : CB 13, R391393 (2003).
Tzivion, G. & Avruch, J. 14-3-3 proteins: active cofactors in cellular regulation by
serine/threonine phosphorylation. The Journal of biological chemistry 277, 30613064, doi:10.1074/jbc.R100059200 (2002).
Liu, D. et al. Crystal structure of the zeta isoform of the 14-3-3 protein. Nature
376, 191-194, doi:10.1038/376191a0 (1995).
Petosa, C. et al. 14-3-3zeta binds a phosphorylated Raf peptide and an
unphosphorylated peptide via its conserved amphipathic groove. The Journal of
biological chemistry 273, 16305-16310 (1998).
Hegle, A. P., Nazzari, H., Roth, A., Angoli, D. & Accili, E. A. Evolutionary
emergence of N-glycosylation as a variable promoter of HCN channel surface
expression. American journal of physiology. Cell physiology 298, C1066-1076,
doi:10.1152/ajpcell.00389.2009 (2010).
Gong, Q., Anderson, C. L., January, C. T. & Zhou, Z. Role of glycosylation in cell
surface expression and stability of HERG potassium channels. American journal
of physiology. Heart and circulatory physiology 283, H77-84,
doi:10.1152/ajpheart.00008.2002 (2002).
Liao, Z., Lockhead, D., Larson, E. D. & Proenza, C. Phosphorylation and
modulation of hyperpolarization-activated HCN4 channels by protein kinase A in
the mouse sinoatrial node. The Journal of general physiology 136, 247-258,
doi:10.1085/jgp.201010488 (2010).
Yu, H. G., Lu, Z., Pan, Z. & Cohen, I. S. Tyrosine kinase inhibition differentially
regulates heterologously expressed HCN channels. Pflugers Archiv : European
journal of physiology 447, 392-400, doi:10.1007/s00424-003-1204-y (2004).
Zong, X. et al. A novel mechanism of modulation of hyperpolarization-activated
cyclic nucleotide-gated channels by Src kinase. The Journal of biological
chemistry 280, 34224-34232, doi:10.1074/jbc.M506544200 (2005).
Arinsburg, S. S., Cohen, I. S. & Yu, H. G. Constitutively active Src tyrosine
kinase changes gating of HCN4 channels through direct binding to the channel
proteins. Journal of cardiovascular pharmacology 47, 578-586,
doi:10.1097/01.fjc.0000211740.47960.8b (2006).
140
211
212
213
214
215
216
217
218
219
220
221
222
223
224
Iakoucheva, L. M. et al. The importance of intrinsic disorder for protein
phosphorylation. Nucleic acids research 32, 1037-1049, doi:10.1093/nar/gkh253
(2004).
Wong, Y. H. et al. KinasePhos 2.0: a web server for identifying protein kinasespecific phosphorylation sites based on sequences and coupling patterns.
Nucleic acids research 35, W588-594, doi:10.1093/nar/gkm322 (2007).
Reetz, O. & Strauss, U. Protein kinase C activation inhibits rat and human
hyperpolarization activated cyclic nucleotide gated channel (HCN)1--mediated
current in mammalian cells. Cellular physiology and biochemistry : international
journal of experimental cellular physiology, biochemistry, and pharmacology 31,
532-541, doi:10.1159/000350074 (2013).
Jung, S. et al. Downregulation of dendritic HCN channel gating in epilepsy is
mediated by altered phosphorylation signaling. J Neurosci 30, 6678-6688,
doi:10.1523/JNEUROSCI.1290-10.2010 (2010).
Huang, J., Huang, A., Zhang, Q., Lin, Y. C. & Yu, H. G. Novel mechanism for
suppression of hyperpolarization-activated cyclic nucleotide-gated pacemaker
channels by receptor-like tyrosine phosphatase-alpha. The Journal of biological
chemistry 283, 29912-29919, doi:10.1074/jbc.M804205200 (2008).
Pielage, J. et al. A presynaptic giant ankyrin stabilizes the NMJ through
regulation of presynaptic microtubules and transsynaptic cell adhesion. Neuron
58, 195-209, doi:10.1016/j.neuron.2008.02.017 (2008).
Koch, I. et al. Drosophila ankyrin 2 is required for synaptic stability. Neuron 58,
210-222, doi:10.1016/j.neuron.2008.03.019 (2008).
Luchtenborg, A. M. et al. Heterotrimeric Go protein links Wnt-Frizzled signaling
with ankyrins to regulate the neuronal microtubule cytoskeleton. Development
141, 3399-3409, doi:10.1242/dev.106773 (2014).
Smith, K. R. et al. Psychiatric risk factor ANK3/ankyrin-G nanodomains regulate
the structure and function of glutamatergic synapses. Neuron 84, 399-415,
doi:10.1016/j.neuron.2014.10.010 (2014).
Bennett, V. & Healy, J. Membrane domains based on ankyrin and spectrin
associated with cell-cell interactions. Cold Spring Harbor perspectives in biology
1, a003012, doi:10.1101/cshperspect.a003012 (2009).
Wilson, P. D. et al. Apical plasma membrane mispolarization of NaK-ATPase in
polycystic kidney disease epithelia is associated with aberrant expression of the
beta2 isoform. The American journal of pathology 156, 253-268 (2000).
Laughery, M. D., Clifford, R. J., Chi, Y. & Kaplan, J. H. Selective basolateral
localization of overexpressed Na-K-ATPase beta1- and beta2- subunits is
disrupted by butryate treatment of MDCK cells. American journal of physiology.
Renal physiology 292, F1718-1725, doi:10.1152/ajprenal.00360.2006 (2007).
Thogersen, L. & Nissen, P. Flexible P-type ATPases interacting with the
membrane. Current opinion in structural biology 22, 491-499,
doi:10.1016/j.sbi.2012.05.009 (2012).
Cunningham, M. R. et al. Novel role for proteinase-activated receptor 2 (PAR2) in
membrane trafficking of proteinase-activated receptor 4 (PAR4). The Journal of
biological chemistry 287, 16656-16669, doi:10.1074/jbc.M111.315911 (2012).
141
225
226
O'Kelly, I., Butler, M. H., Zilberberg, N. & Goldstein, S. A. Forward transport. 143-3 binding overcomes retention in endoplasmic reticulum by dibasic signals. Cell
111, 577-588 (2002).
Jeanclos, E. M. et al. The chaperone protein 14-3-3eta interacts with the nicotinic
acetylcholine receptor alpha 4 subunit. Evidence for a dynamic role in subunit
stabilization. The Journal of biological chemistry 276, 28281-28290,
doi:10.1074/jbc.M011549200 (2001).
142