Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
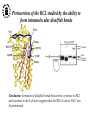
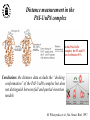
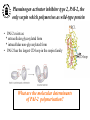
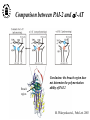
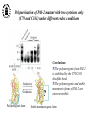
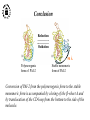
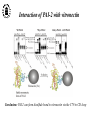
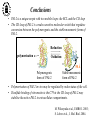
What are the serpins? • It is a family of proteins characterised by a common molecular architecture • Most of the serpins are serine protease inhibitors, but some of them have other functions • Today, more than 500 serpins have been identified in animals, plants, bacteria and viruses • Serpin structure • Inhibitory mechanism of serpins • Serpin polymerisation PAI-1 (plasminogen activator inhibitor type 1), the only serpin which spontaneously converts to the latent form • inhibits the plasminogen activators, uPA and tPA • regulates fibrinolysis (dissolving of blood clots) and cell migration • the specificity and stability of PAI-1 is regulated by cofactors such as heparin and vitronectin • lacks cysteine residues (from Sharp et al. 1999) Why PAI-1 spontaneously converts to latent form? Distance measurement using donor-donor energy migration (DDEM) Time Localisation of the RCL in PAI-1 by intramolecular distance measurements Distances measured P1’ - 313 P3 - 313 Distances in stable PAI-1 mutant (X-ray ) (Å) 69 68 Distances determined by the DDEM method (Å) 55 2 55 2 Preinsertion of the RCL studied by the ability to form intramolecular disulfide bonds Complex Intact Cleaved Oxidized Conclusion: formation of disulfide bonds between the cysteines in RCL and cysteines in the A--sheet suggests that the RCL in active PAI-1 can be preinserted. Conclusion In contrast to other serpins, active PAI-1 has RCL located close to the core and preinserted. This may be a reason why PAI-1 spontaneously converts to latent form. P. Hägglöf et al., J. Mol. Biol. 2003. Inhibitory mechanism of serpins What was known: •serpins form very stable/irreversible complexes with their target proteases • when the complexes were analysed by SDS-PAGE or amino acid sequencing, the serpins were cleaved Major questions: • Are serpins cleaved in the native complexes or the cleavage is an artifact of the analyses? • How look the serpin/protease complex? Quantification of free N-terminals in native serpin/protease complexes PCF Result: in native serpin-protease complexes the N-terminus of PCF is blocked to the same extent as the other N-termini Conclusion: in the native serpin/protease complex the reactive centre of serpin is cleaved and the protease covalently bound to the serpin M. Wilczynska, et al., J. Biol. Chem. 1995. What is the conformation of serpin/protease complex? Hypothetical conformations of stable serpin/protease complex Distance measurement in the PAI-1/uPA complex Conclusion: the distance data exclude the “docking conformation” of the PAI-1/uPA complex but does not distinguish between full and partial-insertion models X M. Wilczynska, et al., Nat. Struct. Biol. 1997. Structural analysis of PAI-1/uPA complex by distance measurement and triangulation Model of the complex P3-266 Distances (Å) P3-185 P3-P1’ P3-313 43,6 34,2 60,3 39,2 49,8 52,1 60,3 8,6 52 52 60 Partial-insertion model Full-insertion model Distances measured by DDEM <30 Conclusion: the distances measured are compatible with full-insertion model M. Fa, et al., Struct. Fold. and Des. 2000. Serpin inhibitory mechanism is driven by serpin metastability Serpin inhibition involves reactive center cleavage and full loop insertion, so the covalently linked protease is translocated from the initial docking site to distal end of serpin. Loop-sheet polymerisation of serpins Wild-type serpins polymerise only under mild denaturing conditions. Some of natural serpin mutants spontaneously polymerise in vivo. This results in diseases like cirrhosis and emphysemia (polymerisation of 1-antitrypsin), angioedema (polymerisation of C1-inhibitor), and dementia (polymerisation of neuroserpin). The polymerisation is accompanied by loss of inhibitory activity. Plasminogen activator inhibitor type 2, PAI-2, the only serpin which polymerises as wild-type protein • PAI-2 exists as: * extracellular glycosylated form * intracellular non-glycosylated form • PAI-2 has the largest CD-loop in the serpin family What are the molecular determinants of PAI-2 polymerisation? Comparison between PAI-2 and 1-AT Breach region Conclusion: the breach region does not determine the polymerisation ability of PAI-2 M. Wilczynska et al., Febs Lett. 2003 Polymerisation of native and DTT-reduced PAI-2 Non-denaturing PAGE Native PAI-2 Reduced PAI-2 Conclusion: reduction of PAI-2 makes the protein resistant to polymerisation. Identification of a cysteine which is important for polymerisation ability of PAI-2 Non-denaturing PAGE Conclusion: Substitution of C79 or C161 to serine makes PAI-2 resistant to polymerisation. Analysis of trypsin-degraded wt PAI-2 by Maldi-tof mass spectrometry Cysteines 79 and 161 form disulfide bond Polymerisation of PAI-2 mutant with two cysteines only (C79 and C161) under different redox conditions 2 Oxidation Polymerogenic form ? Conclusions: The polymerogenic form PAI-2 is stabilised by the C79/C161 disulfide bond. The polymerogenic and stable monomeric forms of PAI-2 are interconvertible. Stable monomerogenic form Triangulation of the C79 in stable monomeric PAI-2 by intramolecular distance measurements using DDEM Conclusion: Stable monomeric form of PAI-2 has the CD-loop folded on a side of the molecule Is the translocation of CD-loop in PAI-2 linked to conformational changes in the A-β-sheet of the inhibitor? + Non-annealed + uPA RCL peptide PAI-2 Annealed *SDS/PAGE Complex *Western blot *Quantification of cleaved PAI-2 by phosphorimager Annealing [%] Annealing of synthetic RCL-peptide into wt PAI-2 and its mutants to compare the opening of the A--sheet Wt PAI-2 C5S PAI-2 C145S PAI-2 C79S PAI-2 C161S PAI-2 Nonsecific peptide 30 20 10 Cleaved = annealed 0 0 25 50 75 100 Peptide excess Conclusion: the A--sheet of PAI-2 is more open in the polymerogenic form than in the stable monomeric form of the inhibitor. Conclusion Reduction Oxidation 50 Å Polymerogenic form of PAI-2 Stable monomeric form of PAI-2 Conversion of PAI-2 from the polymerogenic form to the stable monomeric form is accompanied by closing of the -sheet A and by translocation of the CD-loop from the bottom to the side of the molecule. Do the polymerogenic and stable monomeric forms of PAI-2 exist in nature? Polymerisation of PAI-2 from the cytosol (wt PAI-2) and from the secretory pathway (SP-PAI-2) of CHO cells Conclusions: • in the cytosol, PAI-2 exists mainly in the stable monomeric form • in secretory pathway, PAI-2 is in the polymerogenic form. Interaction of PAI-2 with vitronectin Conclusion: PAI-2 can form disulfide-bond to vitronectin via the C79 in CD-loop Conclusions • PAI-2 is a unique serpin with two mobile loops: the RCL and the CD-loop • The CD-loop of PAI-2 is a redox-sensitive molecular switch that regulates conversion between the polymerogenic and the stable monomeric forms of PAI-2. Reduction polymerisation Oxidation Polymerogenic form of PAI-2 50 Å Stable monomeric form of PAI-2 • Polymerisation of PAI-2 in vivo may be regulated by redox status of the cell. • Disulfide-binding of vitronectin to the C79 in the CD-loop of PAI-2 may stabilise the active PAI-2 in extracellular compartments. M. Wilczynska et al., EMBO J. 2003; S. Lobov et al., J. Mol. Biol. 2004.