Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Synaptic gating wikipedia , lookup
Neuroanatomy wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Subventricular zone wikipedia , lookup
Development of the nervous system wikipedia , lookup
Optogenetics wikipedia , lookup
Channelrhodopsin wikipedia , lookup
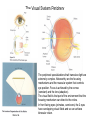
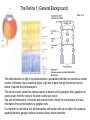
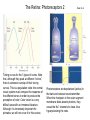
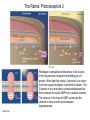
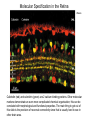
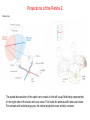
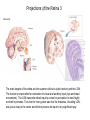
The Visual System Periphery Bear et al. The peripheral specializations that\ transduce light are extremely complex. Noteworthy are the focusing mechanisms and the muscular system that controls eye position. Focus is achieved by the cornea (constant) and the lens (adaptive). The visual field is that part of the environment that the focusing mechanism can direct to the retina. In front facing eyes (primates, carnivores) the 2 eyes have overlapping visual fields and so can achieve binocular vision. The Retina 1 (General Background) Bear et al. The initial detection of light is via photoreceptors- specialized cells that can transduce a small number of photons into an electrical signal. Light has to pass through the retinal neurons before it reaches the photoreceptors. The retinal circuitry leads from photoreceptors to bipolar cells to ganglion cells; ganglion cell axons project from the retina to the brain via the optic nerve. Two sets of interneurons, horizontal and amacrine cells, modify the transmission of visual information from photoreceptors to ganglion cells. It is important to remember that photoreceptors and bipolar cells do not spike- they produce graded potentials; ganglion cells do produce classic action potentials. The Retina: Photoreceptors 1 Bear et al. There are 2 types of photoreceptors: rods (all wavelengths) and cones (3 subtypes: each sensitive to a specific range of wavelengths, for color vision). Cones are present at highest density in the foveal region- this is the region you use for focus. In the foveal region, ganglion cells receive input from single cones (ultra-high acuity vision) while in peripheral retina there is extensive convergence for greater sensitivity and less acuity. The Retina: Photoreceptors 2 Tuning curves for the 3 types of cones. Note that, although they peak at different “colors”, there is extensive overlap of their tuning curves. This is a population code: the central visual system must compare the response of the different cones in order to produce the perception of color. Color vision is a very difficult area with an immense literature. Although it is immensely important for primates, we will not cover it in this course. Bear et al. Photoreceptors are depolarized (active) in the dark and release neurotransmitter. When the rhodopsin in the outer segment membrane disks absorb photons, they cause the Na+ channels to close, thus hyperpolarizing the rods. The Retina: Photoreceptors 3 Rhodopsin constitutively binds retinal; in its inactive form this prevents rhodopsin from binding to a Gprotein. When light hits retinal, it converts to an active form that causes rhodopsin to bind the G-protein. The G-protein in turn activates a phosphodiesterase that then removes the cyclic GMP from a sodium channel. The removal of the bound cGMP causes the Na+ channel to close and the photoreceptor hyperpolarizes. Bear et al. The Retina: Circuitry and Function 1 Photoreceptors make synaptic contacts onto two types of neurons: bipolar cells and horizontal cells. The horizontal cells are GABAergic (inhibitory) interneurons. The horizontal cells contact the bipolar cells at a site near to where they receive input from the photoreceptor; this complicated synaptic region is within invaginations of the photoreceptors and the details are beyond the scope of this course. The key point is that there are two kinds of bipolar cells: On center bipolar cells and Off center bipolar cells. The functional distinction is that On center cells respond to light with depolarization; Off center cells respond to darkness with depolarization. The cellular reason for this distinction is that these bipolar cells have different receptors for the photoreceptor neurotransmitter (likely glutamate). Bear et al. The Retina: Circuitry and Function 2 Bear et al. Cone pedicles (b) have ribbon synapses; On bipolar (blue) and horizontal cell (yellow) dendrites invaginate into the cone pedicle. Off bipolar dendrites (purple) remain on the outside. The Retina: Circuitry and Function 3 Kandel et al. Rods are depolarized in the dark and release glutamate; this excites the Off-center bipolar and inhibits the On-center bipolar. In the light, the rod hyperpolarizes and stops releasing glutamate; the On-center bipolar now depolarizes (disinhibits) and the Off-center bipolar hypeerpolarizes. The bipolar cells then contact ganglion cells selectively, producing On and Off-center ganglion cells; ganglion cells spike. The Retina: Ganglion Cells 1 Bear et al. The center-surround antagonistic organization of ganglion cell RFs reduces the spatial redundancy of the visual signals- regions of constant illumination do not cause discharge and only information about changes in illumination are transmitted to higher centers of the brain. Of course other cells do signal the overall level of illumination- the ones that project to hypothalamus and pretectum- but there are only a small number of such cells. The Retina: Ganglion Cells 2 Bear et al. The major distinction among ganglion cells is On versus Off-center. Within each class there are many additional varieties. One major distinction is between M (magnocellular) and ) P (parvocellular) types. The M type is large and phasic- it responds well to movement over a large receptive field. The P type is small and tonic- it responds well to stationary input over a small receptive field. The projections from M and P types stay partly separate up to cortex and basically define two key visual operations: sensitivity to motion (M type) and to object identification (P type). There are many additional types of ganglion cells senstive to a variety of spatio-temporal patterns. We won’t go into these varieties in this course. The M-type cells can be considered as reducing temporal redundancy by only responding to changes in illumination over time. Molecular Specification in the Retina Wassle Calbindin (red) and calretinin (green) are 2 calcium binding proteins. Other molecular markers demonstrate an even more complicated chemical organization; this can be correlated with morphological and functional properties. The main thing to get out of this slide is the precision of neuronal connectivity since that is usually hard to see in other brain area. The Retina carries out more complicated computations than Center-Surround Berry, 1999 The upper panel shows the effect of a light flash in the RF center of a ganglion cell. The cell fires with a latency of ~100 ms- this delay is too long. A moving object causes the same cell to begin firing ~300 ms before the object reaches the RF center. The cell anticipates (predicts) the location of the moving object. Schwartz, 2007 Ganglion cells anticipate (predict) the timing of a light pulse. First their response to the repeated,predictable pulses is to stop firing- they cancel the expected signal to reduce redundancy. When the signal stops, they then respond strongly at exactly the time when the signal should have appeared. Projections of the Retina 1 The suprachiasmatic nucleus is a diencephalic (hypothalamus) structure. The superior colliculus is a mesencephalic (midbrain) structure. The lateral geniculate is a diencephalic (thalamus) structure. The Superior Colliculus: The Interface between Sense Input and Directed Movement Fish, Frog Mammals Retina Tectum Retina Pallium Memory Brainstem Spinal Cord: Eye/Body Movement Oriented towards Visual Input Superior Colliculus Cortex: Reward, Memory, Intentions Brainstem Spinal Cord: Eye/Head/Body Movement Oriented towards Visual Input Primates: Saccadic Eye Movements The tectum receives retinal input and can direct the eyes (or head/body) towards the stimulus (e.g. food). This is the orienting response. The tectum (at least frogs, rodents) can also direct body away from a stimulus (predator). Other senses also use tectal circuitry for orienting responses: audition, touch, electroreception. In mammals cortical input can prepare the superior colliculus for intended movements and for movements that will produce rewards. The Superior Colliculus 2 Torres, 2005 In a fish the tectum is responsible for orienting movements towards visual stimuli. Boehnke, 2008 In primates the superior colliculus is still responsible for orienting but receives massive cortical input that is the main determinant of where to direct the eyes. Fecteau and Munoz (2006) have hypothesized that the superior colliculus generates a priority map that determines what to orient towards. The priority map is computed from a salience map (sensory input- how big, bright etc is the input)- and a relevance map (cortex)- how important is the input. The relevance map must be learned. In this way the phylogenetically ancient tectum is co-opted to serve the complex sensory systems of mammals. Projections of the Retina 2 Bear et al. The partial decussation of the optic nerve resuls in the left visual field being represented on the right side of the brain and vice versa. This holds for animals with binocular vision. For animals with side-facing eyes, the retinal projections are entirely crossed. Projections of the Retina 3 Bear et al. The main targets of the retina are the superior colliculus (optic tectum) and the LGN. The tectum is responsible for orientation to visual and auditory input (eye and head movements). The LGN transmits retinal input to cortex for perception is most highly evolved in primates. The view for many years was that the thalamus, inlcuding LGN, was just a relay to the cortex and did not process its input in any significant way. The Lateral Geniculate Nucleus 1 Retinal input to the LGN keeps the eyes separate and keeps the P and M pathways separate. LGN neurons have the same receptive fields as retinal ganglion cells: On or Off centers + surround. So is the LGN merely a relay nucleus that does not process retinal input? Emphatically no. Bear et al. The Lateral Geniculate Nucleus 2 Krahe and Gabbiani Thalamic relay cells, including LGN, express a T-type Ca channel that is inactivated at rest. When the cell is hyperpolarized (by an IPSP) the T channel inactivation is removed and the next excitatory input activates a long lasting Ca2+ spike that triggers bursts of action potentials. So LGN cells can produce isolated spikes or spike bursts. Certain types of neurons have biophysical mechanisms that cause bursting, while others don’t. Is bursting a special code? The Lateral Geniculate Nucleus 3 Krahe and Gabbiani The LGN is the main source of visual input to cortex. Yet it receives less than 20% of its input from cortex. About 80% of LGN input is feedback from visual cortex and it is topographically organized: an LGN projecting to a region of cortex receives feedback from the same region. One function of the cortical feedback is to control LGN bursting. It has been suggested that LGN bursts are a “wake up call” to cortex: bursts tell cortex that some visual input is very important. The Lateral Geniculate Nucleus 4 Lesica and Stanley Modern studies of sensory processing typically use both natural stimuli and noise stimuli designed to mimic some but not all of the characteristics of natural stimuli. Natural stimuli are of course most relevant to the animal; noise stimulli are easy to work with mathematically; so using both is often the most informative. The Lateral Geniculate Nucleus 5 Lesica and Stanley LGN bursts are triggered by objects; they presumably signal something different from isolated spikes. LGN is not merely a relay station but shapes information flowing to the cortex. The Lateral Geniculate Nucleus 6 Feature-linked synchronization of LGN relay cells caused by feedback from visual cortex. Feedback Present + visual input: 2 LGN cells show correlated firing. The neural code in LGN presumably includes correlated firing of relay neurons. This code is controlled by feedback. 1. The thalamus is not merely a feedback center. 2. Correlated activity across a neural population can be an important code. 3. Spike bursts can be an important code. Visual input No Feedback: No correlations are seen. Sillito et al.