Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
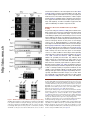
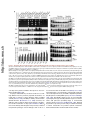
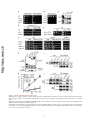
Published in "Molecular Cell 35(5): 563-573, 2009" which should be cited to refer to this work. The Vam6 GEF Controls TORC1 by Activating the EGO Complex Matteo Binda,1 Marie-Pierre Péli-Gulli,1 Grégory Bonfils,1 Nicolas Panchaud,1 Jörg Urban,2 Thomas W. Sturgill,3 Robbie Loewith,2 and Claudio De Virgilio1,* 1Division of Biochemistry, Department of Medicine, University of Fribourg, CH-1700 Fribourg, Switzerland of Molecular Biology, University of Geneva, CH-1211 Geneva, Switzerland 3Department of Pharmacology, University of Virginia Health Sciences Center, Charlottesville, VA 22908, USA *Correspondence: [email protected] 2Department http://doc.rero.ch SUMMARY regulators of TORC1 and play important roles in coupling aminoacid-derived signals to TORC1 in both Drosophila and mammalian cells (Kim et al., 2008; Sancak et al., 2008). RagA and RagB are very similar to each other and orthologous to yeast Gtr1, whereas RagC and RagD are similar and orthologous to yeast Gtr2 (Hirose et al., 1998; Schürmann et al., 1995; Sekiguchi et al., 2001). Rag and Gtr proteins function in heterodimeric complexes, which contain one Gtr1-like GTPase and one Gtr2like GTPase (Nakashima et al., 1999; Sekiguchi et al., 2001). Importantly, amino acid signals are thought to impinge on GTP loading of RagA/B, but it is not known which factors regulate the GTP loading of these GTPases. We previously reported that the EGO complex in yeast, consisting of Ego1/Meh1, Ego3/Slm4, Gtr2, and Gtr1, which was identified subsequently (Gao and Kaiser, 2006; Gao et al., 2005), may function upstream of TORC1 to mediate amino acid signaling (De Virgilio and Loewith, 2006a, 2006b; Dubouloz et al., 2005). In this study, we provide evidence that the EGO complex indeed functions directly upstream of TORC1. TORC1 activity is dictated by the nucleotide-bound state of Gtr1, and this is dependent on the presence of Tco89, a nonessential component of TORC1. Furthermore, we demonstrate that Vam6 colocalizes with the EGO complex/TORC1 at the limiting membrane of the vacuole and functions as a guanine nucleotide exchange factor (GEF) for Gtr1. Thus, in addition to its regulatory role in homotypic vacuolar fusion and vacuole protein sorting as part of the HOPS complex, Vam6 also controls the activity of TORC1 by activating the Gtr1 subunit of the EGO complex. The target of rapamycin complex 1 (TORC1) is a central regulator of eukaryotic cell growth that is activated by a variety of hormones (e.g., insulin) and nutrients (e.g., amino acids) and is deregulated in various cancers. Here, we report that the yeast Rag GTPase homolog Gtr1, a component of the vacuolar-membrane-associated EGO complex (EGOC), interacts with and activates TORC1 in an aminoacid-sensitive manner. Expression of a constitutively active (GTP-bound) Gtr1GTP, which interacted strongly with TORC1, rendered TORC1 partially resistant to leucine deprivation, whereas expression of a growth inhibitory, GDP-bound Gtr1GDP, caused constitutively low TORC1 activity. We also show that the nucleotide-binding status of Gtr1 is regulated by the conserved guanine nucleotide exchange factor (GEF) Vam6. Thus, in addition to its regulatory role in homotypic vacuolar fusion and vacuole protein sorting within the HOPS complex, Vam6 also controls TORC1 function by activating the Gtr1 subunit of the EGO complex. INTRODUCTION The target of rapamycin complex 1 (TORC1) is a structurally and functionally conserved, essential multiprotein complex that posi- RESULTS AND DISCUSSION tively regulates cell growth by promoting anabolic processes (e.g., protein synthesis) and inhibiting catabolic processes (e.g., EGOC Acts Upstream of TORC1 autophagy) in response to diverse signals, including mitogenic Loss of EGO complex (Ego1, Ego3, Gtr1, and Gtr2) or TORC1 growth factors, energy/ATP levels, and amino acids (De Virgilio (Tco89) subunits results in an inability to restart growth following and Loewith, 2006a; Wullschleger et al., 2006). The mechanisms exposure to rapamycin (Figure 1A; Dubouloz et al., 2005). Similar that couple growth factors and energy levels to mammalian to wild-type cells treated with rapamycin or the more recently TORC1 (mTORC1) have been characterized in considerable described TORC1 inhibitor caffeine (Kuranda et al., 2006; Reinke detail and implicate the phosphatidylinositol 3-kinase (PI3K), et al., 2006; Wanke et al., 2008), these mutants also possess Akt, TSC1/TSC2, and Rheb, a small guanosine triphosphate decreased TORC1 activity as assessed by monitoring phosphor(GTP)-binding protein that directly binds to and stimulates ylation of the TORC1 substrate Sch9 (Urban et al., 2007) mTORC1 (Sarbassov et al., 2005). The mechanisms through (Figure 1A). Moreover, the potent increase in TORC1 activity which amino acids signal to TORC1, in contrast, have largely re- observed in wild-type cells treated with cycloheximide, a transla1 mained elusive until very recently. Two complementary studies tion elongation inhibitor that may indirectly boost the levels of have reported that the conserved Rag GTPases act as upstream free intracellular amino acids (Beugnet et al., 2003; Urban 1 http://doc.rero.ch Figure 1. EGOC Acts Upstream of TORC1 (A) Loss of TORC1 (Tco89) or EGOC (Ego1, Ego3, Gtr1, and Gtr2) subunits causes a defect in recovery from rapamycin-induced growth arrest and decreases TORC1 activity. (Top) Wild-type and isogenic mutant cells were grown exponentially in YPD, treated for 6 hr with rapamycin (200 ng ml1), and then washed twice and spotted as 10-fold serial dilutions on YPD plates. (Bottom) Indicated strains expressing a plasmid-based copy of SCH9T570A-HA5 were grown exponentially (in YPD + 0.2% glutamine; EXP) and treated for 30 min with rapamycin (RAP; 200 ng ml1), caffeine (CAF; 20 mM), or cycloheximide (CHX; 25 mg ml1). Protein extracts were prepared and subsequently treated with NTCB to cleave polypeptide chains at cysteine residues (Urban et al., 2007). Aliquots were then analyzed by SDS-PAGE, and immunoblots were probed with anti-HA antibodies (only the migration pattern of the C terminus of Sch9 is shown). The extent of Sch9 phosphorylation (and, by proxy, TORC1 activity) was determined by quantifying the signal of the slowest migrating (most highly phosphorylated) species of Sch9 (*) and dividing this by the total signal observed for all species. For comparison, this ratio was set to 100% for wild-type cells and calculated correspondingly (in percent) for the indicated mutants (numbers below lowest chart; standard deviations were below 5% in each case). (B) Gtr1GTP suppresses neither the defects in recovery following rapamycin treatment nor the intrinsically low TORC1 activity in tco89D cells. Wild-type and double gtr1D tco89D mutant strains were transformed with an empty vector or vectors that express GTR1, GTR1GTP, or GTR1GDP from the endogenous promoter and were assayed as in (A). (C) Growth inhibition following overproduction of Gtr1GDP and growth inhibition on nitrogen poor (proline- and urea-containing) media following overproduction of Gtr1GTP depend on the presence of Tco89. Single gtr1D and double gtr1D tco89D mutant strains were transformed with an empty vector or vectors that express GTR1, GTR1GTP, or GTR1GDP from the doxycycline-inducible TetON promoter; were grown to exponential phase in doxycycline-containing media; and were spotted on plates containing, or not, doxycyline (DOX; 5 mg ml1) and rapamycin (RAP; 10 ng ml1) and either 75 mM NH4+ (top three panels), 10 mM proline, or 10 mM urea as nitrogen source as indicated. (D) Hyperactive TOR1A1957V and TOR1I1954V alleles, unlike wild-type or the hypoactive TOR1W2176R allele, suppress the defect in recovery following rapamycin treatment in gtr1D, but not tco89D cells. Wild-type and single gtr1D and tco89D mutant strains were transformed with an empty vector or vectors that express TOR1, TOR1A1957V, TOR1I1954V, or TOR1W2176R from the endogenous promoter and were assayed as in (A). et al., 2007), was partially dependent on the presence of Ego1, Ego3, Gtr1, or Gtr2, further supporting the idea that the EGO complex may function upstream of TORC1 (Figure 1A). Observations from several genetic experiments are in line with this interpretation. First, expression of Gtr1GTP or Gtr1GDP alleles, which are predicted to be restricted to either a GTP- or GDP-bound conformation (Gao and Kaiser, 2006; Nakashima et al., 1999), 2 did not alter the intrinsically low TORC1 activity or the inability to recover following rapamycin treatment of tco89D cells 2 (Figure 1B). Second, overexpression of Gtr1GDP reduced growth rate on rich media, whereas overexpression of Gtr1GTP reduced growth rate on media containing poor nitrogen sources (proline and urea), and these phenotypes were suppressed in tco89D cells (Figure 1C). Third, hyperactive TOR1A1957V and TOR1I1954V alleles (Reinke et al., 2006), unlike wild-type TOR1 or the hypoactive TOR1W2176R allele, fully suppressed the defect in recovery following rapamycin treatment in gtr1D cells, but not in tco89D cells (Figure 1D). Lastly, TORC1 inhibition results in the activation and nuclear localization of the transcription factor Gln3 (Beck and Hall, 1999), an event that is growth inhibitory for cells. Deletion of GLN3 suppressed the defect in recovery from rapamycin treatment, but not the TORC1 activity defect of EGO complex mutants (Figure S1 available online). Collectively, these observations support the idea that the EGO complex signals, both positively and negatively, to TORC1 and further suggest that this signal is mediated by the TORC1 component Tco89. http://doc.rero.ch EGOC Does Not Control TORC1 Indirectly via Gap1 Sorting In a previous study (Gao and Kaiser, 2006), all four subunits of the EGO complex were shown to be required for intracellular sorting of the general amino acid permease Gap1, a process that is controlled by the quality of the external nitrogen source. Accordingly, in wild-type cells grown on a relatively poor nitrogen source such as urea, Gap1 is sorted to the plasma membrane (where it is active for transport), whereas in cells grown on rich nitrogen sources such as glutamate/glutamine or ammonium (NH4+), active Gap1 is internalized and directed to the vacuole for degradation (Springael and André, 1998), and newly synthesized Gap1 is directly sorted from the trans-Golgi to the vacuole (De Craene et al., 2001). Although we observed that loss of EGOC affected TORC1 activity even when cells were grown on rich nitrogen sources, i.e., under conditions in which Gap1 is transcriptionally repressed and not sorted to the plasma membrane (De Craene et al., 2001; Jauniaux and Grenson, 1990), we decided to address the possibility that the EGOC may indirectly affect TORC1 activity via a potential effect on Gap1 sorting. To our surprise, examination of the subcellular distribution of Gap1GFP, which was transiently expressed from a galactose-inducible GAL1 promoter (Nikko et al., 2003), revealed that sorting of Gap1 from exclusively intracellular compartments (in cells grown on NH4+-containing medium) to the plasma membrane (in cells shifted to urea-containing medium) occurred normally in both wild-type and egoc mutant cells (Figure 2A). Similarly, when assayed by subcellular fractionation analysis, loss of Gtr2, which was suggested to be particularly important for Gap1 sorting (Gao and Kaiser, 2006), had no impact on the cells’ ability to Figure 2. EGOC Does Not Control TORC1 Indirectly via Gap1 Sorting (A) EGO complex subunits (i.e., Gtr1 or Gtr2) are not required for sorting Gap1 to the plasma membrane. Prototrophic wild-type, gtr1D, and gtr2D strains expressing a plasmid-encoded Gap1-GFP under the control of the galactose-inducible GAL1 promoter were pregrown on SD medium (containing 75 mM NH4+ as nitrogen source and 2% raffinose/0.1% sucrose as carbon source). Cells were then incubated for 2 hr in the presence of 2% galactose (to induce Gap1-GFP expression) and analyzed by fluorescence microscopy either directly (NH4+) or following an additional 2 hr incubation on SD media containing 10 mM urea as nitrogen source (urea) and 2% glucose (to repress Gap1-GFP de novo synthesis). As reported previously, overproduction of Gap1-GFP from the GAL1 promoter causes some accumulation of Gap1GFP in the vacuole (De Craene et al., 2001). TR, transmission. (B) Prototrophic wild-type and gtr2D cells growing on SD medium containing 10 mM urea as nitrogen source were lysed and fractionated by equilibrium density centrifugation on continuous 20%–60% sucrose gradients (Kaiser et al., 2002). The fractions were assayed for the presence of Gap1, the plasma membrane marker protein Pma1, or the endosomal marker protein Pep12 using specific antibodies that recognize the corresponding proteins. (C) Gap1 is not required for recovery of cells following rapamycin treatment. Prototrophic wild-type and gap1D cells were treated as in Figure 1A. (D) Gap1 is neither required for normal TORC1 activity in exponentially growing cells (EXP) nor for TORC1 activation by cycloheximide (CHX). Prototrophic 3 wild-type and gap1D cells were grown on YPD (+ 0.2% glutamine), or on SD media containing 75 mM NH4+ (NH4+) or 10 mM urea (urea) as nitrogen source and were assayed for TORC1 activity as in Figure 1A. 3 http://doc.rero.ch Figure 3. Alternative Nucleotide-Bound States of Gtr1 and Gtr2 Are Required for Amino-Acid-Dependent Control of TORC1 (A) The nucleotide-binding states of Gtr1 and Gtr2 control both the cell’s ability to recover from rapamycin treatment and TORC1 activity. Rapamycin recovery and TORC1 activity assays (as in Figure 1A) were determined in a gtr1D gtr2D double-mutant strain carrying all combinations of plasmids between pGTR1, pGTR1GTP, or pGTR1GDP and pGTR2, pGTR2GTP, or pGTR2GDP. (B) Intracellular pools of free amino acids increase rapidly following CHX treatment. Intracellular, free amino acids were determined in prototrophic wild-type cells subjected to CHX (25 mg ml1) treatment for the times indicated and expressed as fold increase compared to the levels detected in exponentially growing (EXP) cells. Cells were grown on YPD (+ 0.2% glutamine) or (as indicated) on SD without amino acids (NH4+). Experiments were done in triplicate and expressed as mean + SD. As assessed by one-way analysis of variance (ANOVA) followed by posttest analysis, the CHX-induced increases are statistically significant for all amino acids (except for arginine) with P values that are < 0.05 for glutamate and methinonine and < 0.001 for all other amino acids. (C) TORC1 is reversibly inactivated in response to amino acid starvation. Leucine (leu2D; top two panels), lysine (lys2D), and histidine (his3D) auxotrophic wildtype strains were grown to exponential phase (EXP) in medium containing leucine (LEU), lysine (LYS), or histidine (HIS), respectively, and were then transferred to a medium lacking the corresponding amino acid. Leucine (2.8 mM), lysine (0.4 mM), and histidine (0.5 mM) were readded after 90 min of starvation. Samples were taken at the times indicated following the medium changes and were assayed as in Figure 1A. The leucine starvation experiment was also carried out in the presence of cycloheximide (25 mg ml1), which was added at time point 0 (before starvation; second panel from top). (D) TORC1 is partially insensitive to leucine deprivation in cells expressing Gtr1GTP. Leucine auxotrophic wild-type and gtr1D cells carrying the empty vector or vectors that express GTR1, GTR1GTP, or GTR1GDP from the doxycycline-inducible TetON promoter were assayed for TORC1 activity (as in C). sort Gap1 to the plasma membrane when grown on urea-containing medium (Figure 2B). In trying to understand why our above results were so strikingly different from the previously reported observations, we noted that, in contrast to various wild-type strains, including S1278b (Grenson, 1983), KT1960 (see below), and the S288Cderived BY4741/2 (Brachmann et al., 1998) used here, the wild-type strain used by the Kaiser group appears to be defective for NH4+-inactivation of Gap1 (Gao and Kaiser, 2006). In 4 this context, Roberg and colleagues have previously noted that this particular phenotype of their wild-type strain is due to a 4 loss-of-function allele at the PER1 locus (Roberg et al., 1997), which apparently is closely linked to (and possibly within the same transcriptional unit as) the NADP+-dependent glutamate dehydrogenase-encoding GDH1 gene (Courchesne and Magasanik, 1983). Remarkably, defects in per1 are associated with a pleiotropic phenotype, including aberrant responses of various amino acid permeases (including Gap1) to environmental nitrogen signals (Courchesne and Magasanik, 1983). Although beyond the scope of this present study, it would be interesting to further characterize the potential synthetic interactions between EGOC/TORC1 and Per1/Gdh1 in future studies. Regarding the present study, loss of Gap1 did not reduce the cells’ ability to recover from a rapamycin treatment (Figure 2C) and had no impact on TORC1 activity in cells grown on rich (YPD + 0.2% glutamine or SD-NH4+) or on poor (urea) nitrogen sources (Figure 2D). Taken together, these results, which were also reproducible in the completely unrelated KT1960 strain background (Pedruzzi et al., 2003; Stuart et al., 1994; data not shown), show that EGOC does not affect TORC1 via Gap1 sorting. http://doc.rero.ch Alternative Nucleotide-Bound States of Gtr1 and Gtr2 Are Required for Amino-Acid-Dependent Control of TORC1 To further examine the role of Gtr1 and Gtr2 in TORC1 regulation, we tested different combinations of nucleotide-restricted GTR1 and GTR2 alleles (Gao and Kaiser, 2006; Nakashima et al., 1999) for their effect on TORC1 activity and the cells’ ability to recover from a rapamycin treatment. Expression of Gtr1GTP combined with Gtr2 or Gtr2GDP activated TORC1 (when compared to the corresponding heterodimers containing wild-type Gtr1) (Figure 3A). In contrast, expression of Gtr2GTP combined with Gtr1 inhibited TORC1 (when compared to the wild-type heterodimer). Moreover, expression of Gtr1GDP, irrespective of the nucleotide-binding status of Gtr2, was dominant-negative, as it abolished TORC1-controlled Sch9 phosphorylation under all conditions (including cycloheximide treatment) (Figure 3A). Consistent with previous studies (Kim et al., 2008; Sancak et al., 2008), these observations demonstrate that GTP-loaded Gtr1 and GDP-loaded Gtr2 stimulate TORC1, whereas GDPloaded Gtr1 and GTP-loaded Gtr2 inhibit TORC1. Activation of TORC1 by expression of Gtr1GTP and by cycloheximide treatment was not additive, suggesting that aminoacid-dependent control of TORC1 may be controlled, at least in part, by the nucleotide-binding status of Gtr1 (Figure 3A). To explore this possibility further, we first sought to verify that cycloheximide treatment causes, as speculated, an increase in the pools of free intracellular amino acids. In prototrophic wildtype cells, we observed a significant accumulation of various amino acids within 5–15 min following cycloheximide treatment (Figure 3B). This effect (which was comparable in gtr1D cells; data not shown) and the corresponding activation of TORC1 were both observed to a similar extent in cells growing on YPD (+ 0.2% glutamine) and in cells growing on SD without amino acids (Figures 3B and S2), indicating that the accumulation of free amino acids following cycloheximide treatment does not depend on the uptake of extracellular amino acids. Interestingly, among the various amino acids, leucine appeared to accumulate most strongly and, hence, may play a particular role in TORC1 Figure 4. Gtr1, Preferentially in Its GTP-Bound Form, Physically Interacts with TORC1 (A and B) TORC1 subunits Tco89 and Kog1 preferentially interact with GTP-bound Gtr1GTP. Wild-type control cells (A and B, lane 1) or wild-type cells expressing Tco89-TAP (A, lanes 2–4) or Kog1-TAP (B, lanes 2–4) were transformed with plasmids expressing GST-Gtr1 (A and B, lanes 1 and 2; WT), GST-Gtr1GTP (A and B, lane 3; GTP), or GST-Gtr1GDP (A and B, lane 4; GDP). Cell lysates (Input) and TAP pull-down fractions were subjected to SDS-PAGE, and immunoblots were probed with anti-GST or anti-protein A (anti-TAP) antibodies as indicated. (C) Gtr1 and Gtr1GTP, but not Gtr1GDP, specifically interact with Ego1 and Tco89 in two-hybrid assays. Possible interactions between Gtr1 variants (expressed from the pPR3-N prey vector) and Ego1 or Tco89 (both expressed from the pCabWT bait vector) were evaluated using the membrane-based split ubiquitin yeast two-hybrid system. Numbers represent mean b-galactosidase activities ± SD (in Miller units) from at least three different transformants for each pair of plasmids. Values that were at least 15-fold higher than the corresponding control (empty bait vector) are shown in bold. (D and E) Gtr1-TORC1 interaction, but not Gtr1GTP-TORC1 interaction, is sensitive to leucine starvation. Tco89-TAP (D) or Kog1-TAP (E) were precipitated from extracts prepared from wild-type cells coexpressing GST-Gtr1 or GST-Gtr1GTP. Cells were either harvested in exponential growth phase prior 5 to (+) or following a 60 min period of leucine starvation (). Cell lysates (Input) and TAP pull-down fractions were subjected to SDS-PAGE, and immunoblots were probed with anti-GST or anti-protein A (anti-TAP) antibodies as indicated. 5 http://doc.rero.ch Figure 5. Vam6 Is a Gtr1 Nucleotide Exchange Factor (A) Overproduction of Gtr1GDP results in a synthetic growth defect when combined with gtr1D or vam6D, but not when combined with ypt7D. Indicated strains expressing GTR1 or GTR1GDP from the galactose-inducible GAL1 promoter were grown overnight to exponential phase and spotted on galactose-containing plates. (B and C) Loss of Vam6, like loss of the EGOC subunits Gtr1 and Gtr2, causes a defect in recovery from a rapamycin-induced growth arrest (B) and decreases TORC1 activity (C). For experimental details, see Figure 1A. wild-type cells, but not gtr1D or tco89D strains, resistant to low rapamycin (D) Overproduction of Vam6 from a doxycycline-inducible TetON promoter renders 6 concentrations. Strains carrying the indicated plasmids were grown overnight to exponential phase and spotted (in serial 10-fold dilutions) on plates containing 5 mg ml1 doxycyline and 3 ng ml1 rapamycin. 6 dictating the nucleotide-binding status of Gtr1. Accurate experimental determination of the Gtr1 nucleotide-binding status in vivo is not straightforward because Gtr1 and Gtr2 appear to be asymmetrically loaded with guanine nucleotides within heterodimeric complexes. To corroborate our model by alternative means, we therefore sought to isolate the Gtr1 GEF, which would allow us to more specifically modulate the nucleotide-binding status of Gtr1 within cells. Based on our finding that overproduction of Gtr1GDP was semidominant with respect to growth inhibition, we reasoned that loss of the Gtr1 GEF, like loss of Gtr1 (Figure 5A), should yield a strong synthetic growth defect when combined with a construct that allows accumulation of Gtr1GDP. Consequently, we carried out a systematic, genome-wide synthetic dosage lethal (SDL) screen (Measday et al., 2005) in which we overproduced Gtr1GDP and tracked down corresponding synthetic growth defects. The two strongest positive hits in this screen included gtr1D—confirming the validity of our screening procedure—and vam6D (Figure 5A). The conserved Vam6 protein is a subunit of the homotypic fusion and vacuole protein sorting (HOPS/class C-Vps) complex in yeast (Caplan et al., 2001; Ostrowicz et al., 2008), which has recently also been suggested to control, via an unknown mechanism, TORC1 signaling in response to amino acids (Zurita-Martinez et al., 2007). Intriguingly, the HOPS complex is thought to facilitate the transition from tethering to trans-SNARE pairing during fusion at the vacuole, in part by Vam6-mediated nucleotide exchange on Ypt7, which is homologous to the mammalian Rab-7 GTPase (Wurmser et al., 2000). Combined with our data above (and the fact that Vam6, but not Ypt7, is required for cells to grow in the presence of elevated Gtr1GDP levels; Figure 5A), this led us to speculate that Vam6 may have a dual role in activation of both Ypt7 and Gtr1. In accordance with a model in which Vam6 activates Gtr1, we found that loss of Vam6, like loss of Gtr1 (or other EGOC subunits), resulted in a defect in recovery from rapamycininduced growth arrest, constitutively reduced TORC1 activity, and reduced cycloheximide-induced TORC1 hyperactivation (Figures 5B and 5C). Moreover, overproduction of Vam6 http://doc.rero.ch activation in yeast (Figure 3B). In line with this interpretation, we found TORC1 to be reversibly inactivated in response to leucine (and less pronouncedly in response to lysine or histidine) deprivation (Figure 3C). Notably, inhibition of TORC1 activity following leucine starvation could be partially suppressed by either treatment of cells with cycloheximide or overproduction of Gtr1GTP (Figures 3C and 3D); reactivation of TORC1 by readdition of leucine depended on Gtr1 but was abolished in the presence of Gtr1GDP (Figure 3D). Because overproduction of Gtr1GTP was unable to reverse TORC1 inactivation following NH4+ starvation (Figure S3), we infer from this data set that Gtr1, within an intact EGO complex, likely mediates at least part of the amino acid signaling to TORC1. Gtr1, Preferentially in Its GTP-Bound Form, Physically Interacts with TORC1 To examine whether Gtr1 may directly interact with TORC1, we coexpressed C terminally TAP-tagged versions of the TORC1 subunits Tco89 or Kog1 (the yeast homolog of mammalian raptor; Loewith et al., 2002) and different GST-tagged Gtr1 variants. Using coimmunoprecipitation assays, we were able to demonstrate that both TORC1 subunits specifically and preferentially interacted with GTP-bound Gtr1, whereas the corresponding interactions with Gtr1GDP were close to background levels (Figures 4A and 4B). Employing a membrane-based twohybrid system, we confirmed that Gtr1 and, more efficiently, Gtr1GTP, but not Gtr1GDP, specifically associated with Tco89 as well as with the EGO complex subunit Ego1 (Figure 4C). Given the role of Gtr1 in mediating an amino acid signal to TORC1, we also evaluated whether amino acids regulate the interaction between Gtr1 and TORC1. We found that the Gtr1-TORC1 interaction was sensitive to leucine starvation, whereas Gtr1GTP remained associated with TORC1 under the same conditions (Figures 4D and 4E). Vam6 Is a Gtr1 Nucleotide Exchange Factor Our findings above indicate that Gtr1 controls TORC1 function and that amino acid signals may impinge on this process by (E) Overproduction of Vam6 suppresses the semidominant growth defect conferred by overproduction of Gtr1GDP. Wild-type cells expressing GTR1 or GTR1GDP under the control of the galactose-inducible GAL1 promoter were transformed with either an empty vector or a vector that expresses VAM6 from the doxycyclineinducible TetON promoter, grown overnight to exponential phase, and spotted (in serial 10-fold dilutions) on galactose- and doxycyline-containing plates. (F) Expression of hyperactive TOR1A1957V or TOR1I1954V alleles renders wild-type and vam6D cells resistant to low rapamycin concentrations and enables vam6D cells to recover from a rapamycin-induced growth arrest. Wild-type and vam6D strains were transformed with an empty vector or vectors that express TOR1, TOR1A1957V, or TOR1I1954V from their endogenous promoter and were spotted on plates containing, or not, 10 ng ml1 rapamycin and/or assayed for their ability to recover from a rapamycin-induced growth arrest as in Figure 1A. (G) Gtr1 interacts with Vam6. Equal amounts of recombinant GST-Gtr1, GST-Ypt7, GST-Ras2, and GST alone, preloaded on glutathione-sepharose 4B resin, were incubated with extracts prepared from wild-type cells expressing HIS6-HA3-Vam6. Cell lysate (Input) and GST pull-down fractions were then subjected to SDS-PAGE, and immunoblots were probed with anti-GST (top panel) or anti-HA (two lower panels) antibodies. (H) Vam6 is a nucleotide exchange factor for Gtr1 and Ypt7, but not for Ras2. Recombinant GST-tagged GTP-binding proteins (i.e., 20 pMol of Gtr1 (C, B), Ypt7 (-, ,), or Ras2 (:, 6) were preloaded with [3H]GDP and incubated in the presence of HIS6-HA3-Vam6 (Vam6, filled symbols) or HIS6-HA3 (control [c], open symbols) that were purified from exponentially growing yeast. The time course of [3H]GDP release is shown (in percent). Prior heat inactivation of Vam6 (10 min at 95 ) reduced its GDP release activity toward Gtr1 to background levels (data not shown). Experiments were done in triplicate and expressed as mean + SD. (I) Gtr1 and Gtr1GTP interact strongly with Ego1. GST-Ego1 was precipitated from extracts prepared from wild-type cells coexpressing Gtr1-TAP, Gtr1GTP-TAP, or Gtr1GDP-TAP. Cell lysates (Input) and GST pull-down fractions were subjected to SDS-PAGE, and immunoblots were probed with anti-GST or anti-protein A (anti-TAP) antibodies as indicated. (J) Loss of Vam6, but not loss of Ypt7, abolishes the interaction between Gtr1 and 7 Ego1. GST-Ego1 was precipitated from extracts prepared from wild-type, vam6D, or ypt7D cells coexpressing Gtr1-TAP. Cells were harvested in exponential growth phase prior to ( CHX) or after a 30 min cycloheximide (25 mg ml1) treatment (+ CHX). For details, see Figures 4A and 4B. 7 http://doc.rero.ch Figure 6. TORC1 and EGO Complex Subunits Colocalize with Vam68at Vacuolar and Endosomal Membranes (A) GFP-Vam6, Gtr1-GFP, Ego1-GFP, Tor1-GFP, Tco89-GFP, and GFP-Sch9 all localize to the limiting membrane of the vacuole. Leucine (leu2D) auxotrophic wild-type cells expressing functional versions of GFP-fusion proteins either from a plasmid (GFP-Sch9) or from the endogenous chromosomal locus (Gtr1-GFP, 8 http://doc.rero.ch rendered wild-type cells, but not gtr1D or tco89D cells, resistant to low rapamycin concentrations (Figure 5D) and suppressed the semidominant growth defect resulting from Gtr1GDP overproduction (Figure 5E). Finally, expression of hyperactive TOR1A1957V and TOR1I1954V rendered rapamycin-sensitive vam6D cells as resistant to low rapamycin concentrations as wild-type cells and enabled vam6D cells to recover from a rapamycin-induced growth arrest (Figure 5F). Because these genetic experiments place Vam6 upstream of Gtr1, we determined whether Vam6 could interact with Gtr1. We found that Vam6 coprecipitated well with both GST-Gtr1 and the positive control GST-Ypt7; only very weakly with GST-Ras2, which is very closely related to Gtr1; and not at all with GST alone (Figure 5G). Further, Vam6 stimulated GDP release both from its known target Ypt7 and from Gtr1, but not from Ras2 (Figure 5H), indicating that Vam6 acts as a GEF for Gtr1 in vitro. Addition of nucleotide-free GST-Gtr2 did not significantly alter the Vam6mediated GDP release on Gtr1 (data not shown). To explore whether Vam6 functions as a GEF for Gtr1 in vivo, we made use of the fact that Gtr1GTP-TAP, but not Gtr1GDP-TAP, specifically coprecipitates with GST-Ego1 (Figure 5I); hence, the level of Ego1-associated Gtr1 can be used to estimate the relative amount of Gtr1GTP within cells. Using this assay, we noted that loss of Vam6, but not loss of Ypt7, severely reduced the interaction between Gtr1 and Ego1 (both prior to and following cycloheximide treatment), indicating that Vam6 also functions as a GEF for Gtr1 in vivo (Figure 5J). Taken together, our results show that Vam6 regulates the nucleotide-binding status of Gtr1 both in vitro and in vivo and suggest that Vam6 may integrate amino acid signals to coordinate the control of TORC1 activity and vacuolar fusion events. GFP, or Ego1-GFP, however, RFP-Vam6 (and similarly GFPVam6) was found specifically enriched at the contact sites between two adjacent vacuoles (Figures 6A and 6B), which underscores the previously described role of Vam6 in vacuolar fusion. To exclude the possibility that the reduced TORC1 activity observed in vam6D cells is simply a result of their abnormal, fragmented vacuolar morphology (Figure 6C), we also measured TORC1 activity in ypt7D cells, which exhibit similarly fragmented vacuoles as vam6D cells (Figure 6C). The corresponding experiments clearly showed that only loss of Vam6, but not loss of Ypt7, seriously reduced TORC1 activity and prevented CHX-induced TORC1 activation, indicating that vacuolar fragmentation per se does not have an impact on TORC1 activity control (Figure 6D). Thus, together with the observation that the EGO complex is not required for vacuolar fusion (Dubouloz et al., 2005), it appears that Vam6 regulates two effector branches: one to control vacuolar fusion (via Ypt7) and one to control TORC1 (via the EGO complex subunit Gtr1). Conclusions In conclusion, we have demonstrated that Vam6 activates Gtr1 and that Gtr1 and, by extension, the EGO complex act upstream of TORC1 to regulate its activity. Thus, our present work extends and confirms the evolutionarily conserved importance of Rag family GTPases as upstream regulators of TORC1. However, we were surprised that, in contrast to the situation in mammalian cells in which GTP-locked RagB apparently completely uncouples mTORC1 from signals derived from leucine availability (Sancak et al., 2008), GTP-locked Gtr1 had only a partial effect. Perhaps in yeast, there are other signaling routes by which amino acid availability is communicated to TORC1. Alternatively, it is possible that GTP loading must be followed by hydrolysis for Gtr1 to fully activate TORC1 in yeast; this later reaction would not be possible with the GTP-locked Gtr1 protein. In this context, it is also worth noting that mammalian cells express, due to alternative mRNA splicing, two RagB isoforms (RagB/RagBs and RagBl), which are structurally significantly different from each other (Schürmann et al., 1995). It would, therefore, be of interest to also study the role of RagBl in mTORC1 regulation. Finally, our finding that leucine deprivation did not appreciably affect the localization of GFP-Vam6, Gtr1-GFP, Ego1-GFP, Tor1GFP, Tco89-GFP, and GFP-Sch9 in yeast again contrasts with observations made in mammalian cells in which the Rag GTPases were proposed to mediate amino-acid-induced relocalization of mTOR within the endomembrane system to a compartment that contains the TORC1 activator Rheb (Sancak et al., 2008). Though this aspect of TORC1 regulation may have diverged during evolution—the budding yeast homolog of TORC1 and EGO Complex Subunits Largely Colocalize with Vam6 at the Vacuolar Membrane In line with previous reports (Araki et al., 2005; Gao and Kaiser, 2006; Jorgensen et al., 2004; Nakamura et al., 1997; Reinke et al., 2004; Sturgill et al., 2008; Urban et al., 2007), we found that GFP-Vam6, Gtr1-GFP, Ego1-GFP, Tor1-GFP, Tco89-GFP, and GFP-Sch9 all localized predominantly at the vacuolar membrane in both exponentially growing and leucine-starved cells (Figure 6A). In addition, all GFP fusion proteins (except for GFP-Sch9) occasionally localized to punctate structures adjacent to the vacuole, which in the case of Vam6, Gtr1, and Ego1 may correspond to late endosomes (Cabrera et al., 2009; Gao and Kaiser, 2006). Notably, RFP-Vam6 colocalized with TORC1 and EGO complex subunits (i.e., with Tor1-GFP, Tco89-GFP, and Ego1-GFP) at the vacuolar and the presumed endosomal membranes (Figure 6B). Unlike Tor1-GFP, Tco89- Ego1-GFP, Tor1-GFP, and Tco89-GFP) or corresponding leu2D vam6D cells expressing functional GFP-Vam6 from a plasmid were labeled with the vacuolar membrane fluorescent dye FM4-64, and the localization of GFP fusion proteins was compared to FM4-64 staining by fluorescence microscopy. Cells were analyzed during exponential growth in leucine-containing SC medium (+ LEU) or following a 60 min leucine starvation period on SC medium lacking leucine ( LEU). TR, transmission. (B) RFP-Vam6 colocalizes with Tor1-GFP, Tco89-GFP, and Ego1-GFP at vacuolar and presumed endosomal (green/red dots) membranes. In addition, RFP-Vam6 is specifically enriched at vacuolar fusion sites (arrows; see also GFP-Vam6 in A). Functional RFP-Vam6 was expressed from a plasmid. For details, see (A). (C) Loss of Vam6 or Ypt7 results in fragmentation of vacuoles. Exponentially growing wild-type, vam6D, and ypt7D strains were stained with FM4-64 and analyzed as in (A). TR, transmission. 9 (D) Ypt7 is not required for TORC1 activation by cycloheximide (CHX). For experimental details, see Figure 1A. 9 Rheb is unlikely to perform a similar function in TORC1 activation (De Virgilio and Loewith, 2006a)—it will be most interesting to study whether Vam6 homologs are implicated in the control of TORC1 function by mediating nucleotide exchange on Rag GTPases in higher eukaryotes. Supplemental Data include two tables and three figures ACKNOWLEDGMENTS EXPERIMENTAL PROCEDURES http://doc.rero.ch SUPPLEMENTAL DATA Strains, Growth Conditions, and Plasmids The S. cerevisiae strains used in this study are listed in Table S1. Unless stated otherwise, prototrophic strains were pregrown overnight in synthetic medium without amino acids (SD; 0.17% yeast nitrogen base, 0.5% ammonium sulfate, and 2% glucose). Before each experiment, cells were diluted to an OD600 of 0.2 in YPD medium supplemented with 0.2% of glutamine and grown until they reached an OD600 of 0.8. For amino acid deprivation experiments, strains that were specifically auxotrophic for one amino acid were grown to an OD600 of 0.8 on complete synthetic medium (SC; i.e., SD plus all amino acids), washed twice, and resuspended in starvation medium (SC-Leu, SC-Lys, or SC-His lacking specifically one relevant amino acid). For nitrogen starvation, prototrophic cells were grown to an OD600 of 1.0 on SD (without amino acids), washed twice, and resuspended in the same medium without ammonium sulfate. Low-quality nitrogen media were SD media, which contained 10 mM urea or proline instead of ammonium sulfate. The plasmids used in this study are listed in Table S2. Sch9 Phosphorylation Analyses . To analyze Sch9T570A-HA5 C-terminal phosphorylation, we used the chemical fragmentation analysis as described previously (Urban et al., 2007; Wanke et al., 2008). For quantifications of Sch9 phosphorylation, NTCB-cleaved extracts were separated by 7.5% SDS-PAGE followed by immunoblotting with anti-HA antibody 12CA5. The anti-HA antibody was detected with farred fluorescent Alexa Fluor 680 dye-labeled secondary anti-mouse antibody (Invitrogen, A21057), and fluorescence intensity was measured using the Odyssey Infrared Imaging System (LI-COR). GDP Release Assay To assay GDP release, 20 pmol of bacterially expressed GST-tagged G-proteins (i.e., GST-Gtr1, GST-Ypt7, and GST-Ras2) were preloaded by incubating with 40 pmol 50 ,80 -[3H]GDP (32.8 Ci mmol1; NEN) in preload buffer (20 mM HEPES [pH 7.2], 20 mM KOAc, 1 mM DTT, 5 mM EDTA, 1 mg ml1 BSA) for 15 min at 30 C as described (Jones et al., 2000). At the end of the incubation, samples were placed on ice, and MgCl2 was added to 10 mM. Reactions were carried out in 50 ml containing 20 mM HEPES (pH 7.2), 5 mM Mg(OAc)2, 0.5 mM GTP, 0.5 mM GDP, 1 mM DTT, 0.4 mg ml1 BSA, and HIS6-HA3-Vam6 or HIS6-HA3, both purified from exponentially growing yeast. Exchange reactions were initiated by the addition of 10 pmol of the preloaded G proteins. Incubations were carried out at 30 C for varying periods of time, as noted. At intervals, 5 ml samples were removed, added to 3 ml of ice-cold wash buffer (20 mM Tris-HCl [pH 7.5], 20 mM NaCl, 5 mM MgCl2, 1 mM DTT), and filtered through nitrocellulose filters, which were then washed twice with 3 ml of ice cold wash buffer. Radioactivity bound to filters was quantified by liquid scintillation spectrometry using Filtron-X (National Diagnostics, LS-201) scintillation fluid. In all experiments, initial values were 2–4 3 103 dpm ml1. Miscellaneous Coimmunoprecipitation experiments were essentially done as described (Dubouloz et al., 2005; Loewith et al., 2002). Tco89-TAP and Kog1-TAP were purified from lysates with magnetic beads (Invitrogen) that were precoated overnight in PBS with 1 mg ml1 IgG (Sigma) in the presence of 1 M (NH4)2SO4 to facilitate the binding. For quantifications of free amino acids, cells (about 10 OD600) were harvested by filtration, washed three times, and resuspended in distilled water. Following boiling (15 min) and centrifugation (10 min at 13,000 rpm), free amino acids were quantified in the supernatant by pulsed electrochemical detection after separation by anion exchange chromatog-10 raphy with an AAA-direct Dionex Amino Acid Analyzer, using a sodium acetate gradient to increase the ionic strength. 10 We thank Christian Ungermann, Bruno André, and Ted Powers for strains, plasmids, and/or antibodies and Malika Jaquenoud and Patricia Matthey for technical assistance. This research was supported by the Swiss National Science Foundation (R.L. and C.D.V.), the Leenaards Foundation, and the Cantons of Geneva and Fribourg. REFERENCES Araki, T., Uesono, Y., Oguchi, T., and Toh, E.A. (2005). LAS24/KOG1, a component of the TOR complex 1 (TORC1), is needed for resistance to local anesthetic tetracaine and normal distribution of actin cytoskeleton in yeast. Genes Genet. Syst. 80, 325–343. Beck, T., and Hall, M.N. (1999). The TOR signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 402, 689–692. Beugnet, A., Tee, A.R., Taylor, P.M., and Proud, C.G. (2003). Regulation of targets of mTOR (mammalian target of rapamycin) signalling by intracellular amino acid availability. Biochem. J. 372, 555–566. Brachmann, C.B., Davies, A., Cost, G.J., Caputo, E., Li, J., Hieter, P., and Boeke, J.D. (1998). Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14, 115–132. Cabrera, M., Ostrowicz, C.W., Mari, M., LaGrassa, T.J., Reggiori, F., and Ungermann, C. (2009). Vps41 phosphorylation and the Rab Ypt7 control the targeting of the HOPS complex to endosome-vacuole fusion sites. Mol. Biol. Cell 20, 1937–1948. Caplan, S., Hartnell, L.M., Aguilar, R.C., Naslavsky, N., and Bonifacino, J.S. (2001). Human Vam6p promotes lysosome clustering and fusion in vivo. J. Cell Biol. 154, 109–122. Courchesne, W.E., and Magasanik, B. (1983). Ammonia regulation of amino acid permeases in Saccharomyces cerevisiae. Mol. Cell. Biol. 3, 672–683. De Craene, J.O., Soetens, O., and André, B. (2001). The Npr1 kinase controls biosynthetic and endocytic sorting of the yeast Gap1 permease. J. Biol. Chem. 276, 43939–43948. De Virgilio, C., and Loewith, R. (2006a). Cell growth control: little eukaryotes make big contributions. Oncogene 25, 6392–6415. De Virgilio, C., and Loewith, R. (2006b). The TOR signalling network from yeast to man. Int. J. Biochem. Cell Biol. 38, 1476–1481. Dubouloz, F., Deloche, O., Wanke, V., Cameroni, E., and De Virgilio, C. (2005). The TOR and EGO protein complexes orchestrate microautophagy in yeast. Mol. Cell 19, 15–26. Gao, M., and Kaiser, C.A. (2006). A conserved GTPase-containing complex is required for intracellular sorting of the general amino-acid permease in yeast. Nat. Cell Biol. 8, 657–667. Gao, X.D., Wang, J., Keppler-Ross, S., and Dean, N. (2005). ERS1 encodes a functional homologue of the human lysosomal cystine transporter. FEBS J. 272, 2497–2511. Grenson, M. (1983). Inactivation-reactivation process and repression of permease formation regulate several ammonia-sensitive permeases in the yeast Saccharomyces cerevisiae. Eur. J. Biochem. 133, 135–139. Hirose, E., Nakashima, N., Sekiguchi, T., and Nishimoto, T. (1998). RagA is a functional homologue of S. cerevisiae Gtr1p involved in the Ran/Gsp1GTPase pathway. J. Cell Sci. 111, 11–21. Jauniaux, J.C., and Grenson, M. (1990). GAP1, the general amino acid permease gene of Saccharomyces cerevisiae. Nucleotide sequence, protein similarity with the other bakers yeast amino acid permeases, and nitrogen catabolite repression. Eur. J. Biochem. 190, 39–44. Jones, S., Newman, C., Liu, F., and Segev, N. (2000). The TRAPP complex is a nucleotide exchanger for Ypt1 and Ypt31/32. Mol. Biol. Cell 11, 4403–4411. Jorgensen, P., Rupes, I., Sharom, J.R., Schneper, L., Broach, J.R., and Tyers, M. (2004). A dynamic transcriptional network communicates growth potential to ribosome synthesis and critical cell size. Genes Dev. 18, 2491–2505. Kaiser, C.A., Chen, E.J., and Losko, S. (2002). Subcellular fractionation of secretory organelles. Methods Enzymol. 351, 325–338. Kim, E., Goraksha-Hicks, P., Li, L., Neufeld, T.P., and Guan, K.L. (2008). Regulation of TORC1 by Rag GTPases in nutrient response. Nat. Cell Biol. 10, 935–945. http://doc.rero.ch Kuranda, K., Leberre, V., Sokol, S., Palamarczyk, G., and François, J. (2006). Investigating the caffeine effects in the yeast Saccharomyces cerevisiae brings new insights into the connection between TOR, PKC and Ras/cAMP signalling pathways. Mol. Microbiol. 61, 1147–1166. Reinke, A., Anderson, S., McCaffery, J.M., Yates, J., III, Aronova, S., Chu, S., Fairclough, S., Iverson, C., Wedaman, K.P., and Powers, T. (2004). TOR complex 1 includes a novel component, Tco89p (YPL180w), and cooperates with Ssd1p to maintain cellular integrity in Saccharomyces cerevisiae. J. Biol. Chem. 279, 14752–14762. Reinke, A., Chen, J.C., Aronova, S., and Powers, T. (2006). Caffeine targets TOR complex I and provides evidence for a regulatory link between the FRB and kinase domains of Tor1p. J. Biol. Chem. 281, 31616–31626. Roberg, K.J., Bickel, S., Rowley, N., and Kaiser, C.A. (1997). Control of amino acid permease sorting in the late secretory pathway of Saccharomyces cerevisiae by SEC13, LST4, LST7 and LST8. Genetics 147, 1569–1584. Sancak, Y., Peterson, T.R., Shaul, Y.D., Lindquist, R.A., Thoreen, C.C., Bar-Peled, L., and Sabatini, D.M. (2008). The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320, 1496–1501. Sarbassov, D.D., Ali, S.M., and Sabatini, D.M. (2005). Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 17, 596–603. Schürmann, A., Brauers, A., Massmann, S., Becker, W., and Joost, H.G. (1995). Cloning of a novel family of mammalian GTP-binding proteins (RagA, RagBs, RagB1) with remote similarity to the Ras-related GTPases. J. Biol. Chem. 270, 28982–28988. Sekiguchi, T., Hirose, E., Nakashima, N., Ii, M., and Nishimoto, T. (2001). Novel G proteins, Rag C and Rag D, interact with GTP-binding proteins, Rag A and Rag B. J. Biol. Chem. 276, 7246–7257. Loewith, R., Jacinto, E., Wullschleger, S., Lorberg, A., Crespo, J.L., Bonenfant, D., Oppliger, W., Jenoe, P., and Hall, M.N. (2002). Two TOR complexes, only one of which is rapamycin sensitive, have distinct roles in cell growth control. Mol. Cell 10, 457–468. Springael, J.Y., and André, B. (1998). Nitrogen-regulated ubiquitination of the Gap1 permease of Saccharomyces cerevisiae. Mol. Biol. Cell 9, 1253–1263. Measday, V., Baetz, K., Guzzo, J., Yuen, K., Kwok, T., Sheikh, B., Ding, H., Ueta, R., Hoac, T., Cheng, B., et al. (2005). Systematic yeast synthetic lethal and synthetic dosage lethal screens identify genes required for chromosome segregation. Proc. Natl. Acad. Sci. USA 102, 13956–13961. Stuart, J.S., Frederick, D.L., Varner, C.M., and Tatchell, K. (1994). The mutant type 1 protein phosphatase encoded by glc7-1 from Saccharomyces cerevisiae fails to interact productively with the GAC1-encoded regulatory subunit. Mol. Cell. Biol. 14, 896–905. Nakamura, N., Hirata, A., Ohsumi, Y., and Wada, Y. (1997). Vam2/Vps41p and Vam6/Vps39p are components of a protein complex on the vacuolar membranes and involved in the vacuolar assembly in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 272, 11344–11349. Sturgill, T.W., Cohen, A., Diefenbacher, M., Trautwein, M., Martin, D.E., and Hall, M.N. (2008). TOR1 and TOR2 have distinct locations in live cells. Eukaryot. Cell 7, 1819–1830. Nakashima, N., Noguchi, E., and Nishimoto, T. (1999). Saccharomyces cerevisiae putative G protein, Gtr1p, which forms complexes with itself and a novel protein designated as Gtr2p, negatively regulates the Ran/Gsp1p G protein cycle through Gtr2p. Genetics 152, 853–867. Nikko, E., Marini, A.M., and André, B. (2003). Permease recycling and ubiquitination status reveal a particular role for Bro1 in the multivesicular body pathway. J. Biol. Chem. 278, 50732–50743. Urban, J., Soulard, A., Huber, A., Lippman, S., Mukhopadhyay, D., Deloche, O., Wanke, V., Anrather, D., Ammerer, G., Riezman, H., et al. (2007). Sch9 is a major target of TORC1 in Saccharomyces cerevisiae. Mol. Cell 26, 663–674. Wanke, V., Cameroni, E., Uotila, A., Piccolis, M., Urban, J., Loewith, R., and De Virgilio, C. (2008). Caffeine extends yeast lifespan by targeting TORC1. Mol. Microbiol. 69, 277–285. Wullschleger, S., Loewith, R., and Hall, M.N. (2006). TOR signaling in growth and metabolism. Cell 124, 471–484. Ostrowicz, C.W., Meiringer, C.T., and Ungermann, C. (2008). Yeast vacuole fusion: a model system for eukaryotic endomembrane dynamics. Autophagy 4, 5–19. Wurmser, A.E., Sato, T.K., and Emr, S.D. (2000). New component of the vacuolar class C-Vps complex couples nucleotide exchange on the Ypt7 GTPase to SNARE-dependent docking and fusion. J. Cell Biol. 151, 551–562. Pedruzzi, I., Dubouloz, F., Cameroni, E., Wanke, V., Roosen, J., Winderickx, J., and De Virgilio, C. (2003). TOR and PKA signaling pathways converge on the protein kinase Rim15 to control entry into G0. Mol. Cell 12, 1607–1613. Zurita-Martinez, S.A., Puria, R., Pan, X., Boeke, J.D., and Cardenas, M.E. (2007). Efficient Tor signaling requires a functional class C Vps protein complex in Saccharomyces cerevisiae. Genetics 176, 2139–2150. 11 11