Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
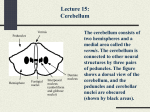
Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 1 of 6 *Note: He used the PDF version of the lecture that Dr. Lester sent, not the PowerPoint version. I. Cerebellum [S1]: Today we are going to talk about the cerebellum and how it is involved in motor systems. II. The Motor System is Organized as a Hierarchy [S2] a. He hopes to convince you that the cerebellum is not involved in direct initiation of movements; rather it functions as a rapid, corrective feedback loop. b. Its purpose is not to initiate movement, but rather to smooth and coordinate movement. The job of actually doing the movement is done by the motor cortex. c. You’ve just heard (referencing the 1:00-2:00 lecture) how basal ganglia can also do similar types of functions. The cerebellum is also involved in that—giving sensory and motor coordination and that feeds onto brainstem centers and those feed toward your skeletal muscles. d. In the cerebellum’s case, the importance is posture. The cerebellum is getting information from every muscle and joint you have to keep you upright and not falling over. e. As you heard, basal ganglion is a structure involved in Parkinson’s disease and Huntingdon’s disease. The cerebellum is similar because it also coordinates movement. f. This picture is from a book by Luther Gulick, MD. He was a big proponent at the beginning of the 19th century of the idea that you need to develop muscle memory through repetitive motions (for example, physical therapy). Repetitive motion would build a muscle memory into the system. His biggest claim to fame is his book about how to do repetitive motions. He also had a student who was asked to develop a sport that people could do during the winter time because it was so cold. This guy was James Naismith, who created basketball! III. What Types of Information Does the Cerebellum Receive? [S3] a. The cerebellum gets input from multiple modalities: 1. Touch (somatosensory) 2. Visual cues; everything you see helps you stand-up right and keep your posture. 3. Auditory signals; if something is loud on one side, you can turn your head and not fall over. 4. Vestibular input (this will be discussed in future lectures, not here.) 5. Proprioceptive feedback allows the cerebellum to know where everything is in the body. People with lesions in the cerebellum don’t know where their hand is or the angle of their elbow. This is problem that will effect everyday movements. IV. Cerebellum: a Fast Feedback Loop for Coordinating Movement [S4] a. How does the cerebellum integrate all of these modalities into refining motor movement? i. The idea is that it is really adept to receiving this feedback from all of these modulatory inputs, sending information up to relay nuclei. The thalamus then sends back information to the cortex and there is some correction being made. ii. We know this because cerebellar lesions cause several different problems. Most of the cerebellar lesions studied were caused by trauma like gunshot wounds, but now with modern technology, we can see that there are also some genetic lesions that occur in the cerebellum. Trauma and genetic lesions will both produce the effects listed below: 1. Nystagmus, which is rapid oscillations of the patient’s eyeballs. 2. Ataxia, which is when the patient walks, they don’t do it with their legs together; rather they will try to widen their gate to make it more stable (going back to the posture idea). 3. Dysdiadochokinesia is something that you see where the patient has pronounced inability to perform rapid alternating movements. You ask them to touch between two different places quickly, but they cannot do it. 4. They also cannot control their range of movement. If you ask them to reach for an object, they will either over-shoot or under-shoot the area they are reaching for. This is called dysmetria. 5. Intention tremors are a little bit different than a regular tremor because you see the oscillatory movement only when you are initiating the movement. Once you have initiated the movement, if you just have a cerebellar lesion, you are good to go. They have trouble beginning their motions. 6. They have several other motor learning difficulties, for example they cannot play the piano. Fine motor control, fine coordination of their bodies and fingers is severely impaired. [SQ:] Will a cerebellar lesion produce contralateral or ipsilateral dysfunction? It really depends because lesions are not usually pinpointed to these little areas, so usually it will produce effects on both areas, as far as I’ve seen. A lot of people call lesions that are in the deep cerebellar nuclei and down in the brainstem— cerebellar lesions—but it is really more towards the output of the cerebellum. What you call the cerebellum depends on what anatomy you are looking at. Some people include deep cerebellar nuclei as part of the cerebellum and some do not. You will see this with some of the genetic disorders, like spinocerebellar ataxia; the area where you lose neurons is kind of all over the place. But if you look at these genetic lesions (not caused by trauma), there are some diseases called Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 2 of 6 spinocerebellar ataxia, where you don’t see a lesion but you do see a remarked atrophy of the cerebellum. It is also in these cases where you have marked cerebellar problems. Dr. Lester draws a picture to further explain the answer to this question: With the cerebellum the connections are to the cortex which is ultimately going to talk with the opposite side of the body. It talks to the opposite cerebellum but loops back, so there is double loop. It interacts with the contralateral cerebellum, so if the lesion is in the cerebellum, it is going to be ipsilateral for the cerebellar damage. Therefore, if the cerebellum is lesioned on one side, the movement problems are going to be ipsilateral. Right motor cortex damage, left motor problems. Left cerebellar damage, left motor problems, but it is still involved in the right because of the dull cross, which you should probably know. For lesions on the output of the cerebellum, you cannot really predict which side the effects will be on and the effects could present on both sides. Dr. Waldwick: For lesions on the output of the cerebellum, you cannot really predict which side the effects will be on and the effects could present on both sides. V. Atrophy of Cerebellum Cause by SCA1 [S5] a. This is a case where there is a genetic abnormality in the cerebellum. b. If you don’t have a lesion that is caused by a trauma, genetic disorders can cause atrophies of the cerebellum. These genetic disorders are called spinocerebellar ataxia. There are 20-30 of these and they are due to repeats of different genes. c. In spinocerebellar ataxia (SCA), there is a CAG repeat in a gene called Ataxin. It actually causes degeneration of the purkinje cells of the cerebellum. VI. Spinocerebellar Ataxia Figures [S6] a. This slide shows the different spinocerebellar ataxia and the orange are the loci where you have gross degeneration of sets of neurons. b. You can see that a lot of them have deep cerebella nuclei deficits. c. Most of them will have deficits in cortex or brainstem, so it is not necessarily that they are just cerebellar but they manifest themselves with the symptoms listed above (dysmetria, dysdiadochokinesia, etc). d. Cell loss from the cerebellar purkinje layer is usually a feature you find with a lot of these spinocerebellar ataxia diseases. VII. Anatomical Regions of the Cerebellum [S7] a. Anatomically, you know the cerebellum has all of these folds just like cortex, and the idea is to increase surface area to increase computational output. b. You can divide the cerebellum into these zones that are perpendicular to the fissures. c. On either side of the vermis (the middle layer), you have regions that are called the nodulus and the floculus. Those are more vestibular anatomical areas. VIII. Functional Regions of Cerebellum [S8] a. People who study the cerebellum really describe regions on a functional level, rather than an anatomical level. b. There are 3 functional regions: 1. Cerebrocerebellum on the left and right of the vermis. It is thought to be part of coordination of voluntary movements and cognitive effects. People with lesions here do show cognitive deficits. 2. Spinocerebellum and 3. Vestibulocerebellum are also associated with the vermis. They both are for posture, balance, and eye movements (ability of eye to stay focused on one subject while your head turns. IX. Peduncles and Deep Cerebellar Nuclei [S9] a. The connection between the cerebellum and the rest of the brain occurs through 3 large pathways known as the cerebellar peduncles: i. The superior cerebral peduncle is almost entirely efferent. Runs from the deep cerebellar nuclei to various areas of the brain. ii. The middle cerebral peduncle is mostly afferent pathway. Neurons in this pathway originate in the pons and come through the middle peduncle into the cerebellum. iii. The inferior cerebral peduncle has both pathways (efferent and afferent) going in and out. b. The cerebral peduncles are just large bundles of axons that course in and out of the cerebellum. X. Inputs TO the Cerebellum [S10] a. The largest input into the cerebellum is from the cerebral cortex through the pons (this input pathway is conveying information about motor coordination). b. Sensory information comes from vestibular nuclei and from the spinal cord. In the spinal cord, most of the input comes from the dorsal nucleus of Clarke. c. The entire cerebellum receives very important modulatory input from the inferior olive of the brainstem. This is the source of a set of axons that make up the climbing fibers in the cerebellum. d. The inferior olive is extremely important in motor learning and memory for the cerebellum. XI. Outputs FROM the Cerebellum [S11] Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 3 of 6 a. The upper motor neurons in the cortex get input through the thalamus. b. The inferior olive, vestibular nuclei, and red nucleus all get input back from the cerebellum so they are giving input to the cerebellum and there is a feedback right back to them. That is all happening through the deep cerebellar nuclei. c. The processing power of the cerebellum is happening in the cerebellar cortex, and all the output neurons (the purkinje neurons) are synapsing onto the deep cerebellar nuclei. d. There are several different types of deep cerebellar nuclei, each of which send input back to cortex, the olive, the nuclei, and the red nucleus. You see how these are feedback mechanisms—all these systems that get information also give information to the cerebellar. e. Remember that the information being sent out goes through the deep cerebellar nuclei. XII. Cellular Anatomy [S12] a. At the cellular level within the cerebellar cortex, the anatomy is repetitive (same as what you should see in the olfactory bulb and the retina). There is a system that repeats itself within lobes of the cerebellum. b. There are 3 cell layers: i. Granular cell layer ii. Purkinje cell layer iii. Molecular layer c. The granular cell layer got its name because of small excitatory neurons that have their cell bodies in this layer. They are getting input from the mossy fibers coming from the pons, synapcing onto the granule cells. The granule cells send out their axons up through the purkinje cell layer and bifurcate in the molecular layer of the cerebellum. d. The importance of these granular cells is that the estimate is 5 x 1010 granule cells. e. People who study the cerebellum and its function try to postulate why there are so many granule cells. At one time, it was thought that there are as many granule cells in the cerebellum as they are neurons in the rest of your brain. That number is actually quite close! The current thought is that there are so many of them because these cells are getting input from all of these different modalities and they have to somehow make sense of that input. So there is a representation of the auditory, somatosensory, vestibular, all within those granule cells. Granule cells send out axons that synapse onto the purkinje cells. f. Coursing up through this pathway and climbing up to the purkinje cell is the input from the inferior olive. That makes up the climbing fibers of the purkinje fibers as you can see labeled. g. These two excitatory inputs have very different properties but both synapse onto the purkinje cell. h. The reason the purkinje cell is so important is because all of the computation done by excitatory and modulatory inhibitory cells within the cerebellar cortex is being made at the purkinje cell. The purkinje cell has to figure out what is coming in from the parallel fibers in the purkinjes and the climbing fibers as far as excitatory. i. How it is being modulated by the interneurons within the cerebellar cortex and then send the information back out. This is the sole output of the cerebellar cortex down to deep cerebellar nuclei. j. So in spinocerebellar ataxia, these patients have large deficits and it is these cells that you see a lot of degeneration in. If you don’t have the purkinje cells, basically your cerebellum is not working well. XIII. Purkinje Cells: Cerebellar Integrators [S13] a. Purkinje cells are really great cellular integrators because they are taking all of this information and trying to figure out what to do with it. XIV. Mossy Fiber / Granule Cells [S14] a. This is a figure from a transgenic mouse that shows how rick the information coming from the mossy fibers to the granule cells really is. Each color represents one axon coming from the pons, synapsing onto cerebellar granule cells. All the sensory information comes through this pathway, and then the granule cells send their information to the purkinje cells. XV. Cerebellar Cytoarchitecture [S15] a. When you put it together and take a sagittal view, this is what you have. You have all of the granule cells residing in this area. b. The purkinje cells and their dendrites are in the molecular layer. c. And the vast majority of the processing power of the purkinje cells is being done out in the dendrites. d. Then the purkinje cells send their axons back out down to deep cerebellar nuclei. XVI. Circuits in Cerebellum [S16] a. The excitatory synapses (shown in blue teal color) previously described are just the first pass. Here are the mossy fibers making granule cells, bifurcating onto the parallel fibers, and the purkinje cells receiving the climbing fiber input from the olive. b. In red color are all the inhibitory cells. The climbing fibers make its only synapse onto the inhibitory purkinje cell. But the mossy fiber contacts granule cells and directly contacts deep cerebellar nuclei. The granule cells not only contact purkinje cells, but also synapse on stellate cells, basket cells, and golgi cells (go back to figure on Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 4 of 6 slide #12). Basket cells, stellate cells, and golgi cells are interneurons within the cerebellar cortex and their function is thought to be to shape how the purkinje cell is integrating the excitatory input by the mossy fiber (through the granular cell) and the climbing fiber. c. On the third panel, we’ve added the deep cerebellar nuclei to show that inputs can be indirect through the purkinje cell processing power or directly through the deep cerebellar nuclei which will feed to the brainstem and thalamus and ultimately effect motor function. XVII. How Does Purkinje Neuron Firing Affect Movement?[S17] a. The purkinje cell is an inhibitory neuron. b. So how does it affect firing and movement? The idea is that the inhibitory spikes that the purkinje cells produces…so if you have a purkinje cell in your brain, it is normally just firing at some frequency (maybe around 1-2 hertz every ½-1 second). That keeps a break on the deep cerebellar nuclei because they are excitatory neurons and that goes towards motor neurons to give voluntary movements or any reflexes you might have. c. The purkinje cell keeps, so these little red lines are the purkinje cells firing their action potentials and that keeps the motor neurons from firing their action potentials and that is because the purkinje cell is releasing GABA onto the deep cerebellar nuclei. But as soon as you have some kind of an input that stops purkinje cells from firing, the deep cerebellar nuclei are disinhibited and allowed to fire to get voluntary movement or reflexes. As soon as that ceases, the purkinje cell starts firing once again. So disinhibition is key in cerebellar processing and its effect to deep cerebellar nuclei. XVIII. Purkinje Cell Death is Common in Genetic Forms of Ataxia [S18] a. Again, you can see why the degeneration of purkinje cells leads to motor dysfunction. b. In this slide, you see a normal patient with their purkinje cells stained with calbindin (a calcium-binding protein that is found in great numbers in purkinje cells). You can see how the dendrites of the purkinje cells spread through the molecular layer. c. But in a spinocerebellar ataxia patient (the bottom picture), you can see that the purkinje cells don’t look healthy. Therefore the processing power of the cerebellum is going to be altered. XIX. Purkinje Cells Show Two Firing Patterns [S19] a. How does the purkinje cell really influence the cerebellar nuclei? It seems like the coding of information, as far as frequency, is very important. The purkinje cell fires two types of spikes: i. Simple spikes are action potentials that most neurons will give off. They are fast to arise and fast to decay; they may have an after hyper polarization. This is what happens in purkinje cells as a result of granular cell input. These spikes happen probably around 1-2 hertz. ii. Atypical action potentials are called complex spikes. This is how the purkinje cells responds when there is climbing fiber input coming from the inferior olive. This type of input is extremely important in motor learning. If you have a lesion in the olive, you cannot really learn. iii. Remember, the climbing cell has a 1:1 ratio with the purkinje cell, meaning that one climbing cell will synapse onto one purkinje cell. Whereas, granule cells are thousands of granule cells to each purkinje cell. iv. When the climbing fiber goes off, it is not trying to represent what is happening with modality, but rather telling the system that something is wrong. For example, you are walking alone and you don’t raise your foot properly and you almost fall over. Next time you come to that curb, the climbing fiber which had told your system that your foot needed to be higher will now make your system learn that your foot needs to be higher so that you don’t trip. So the purkinje cell is the way for the inferior olive to tell your system— LEARN because something wrong has happened. XX. Complex Spikes Indicate Errors [S20] a. This is what is represented in this simple diagram where motor performance is plotted as a function of trials. This is a motor learning paradigm. When your performance is doing good, your simple spikes from your granular cells are firing around 50 hertz. Then, when there is a novel task that gets your attention like stubbing your toe on the curb, your complex spikes, your inferior olive fires off and your motor task decreases. Within a certain number of trials, the frequency with which the simple spikes fire off decreases, so there is a depression of the simple spike activity. Once that simple spike activity has been learned, the complex spike can return back to its behavior and with that, the motor performance becomes good again. This is why cerebellar people say that the fibers act as teachers and adapt the shape of cerebellar functions to mistakes. XXI. Cerebellum Dependent Motor Learning[S21] a. There are two forms of motor learning that allow us to gage what the cerebellum is doing and try to figure out how it is working in diseases where purkinje cells or interneurons are affected, how it is being altered. b. We are only going to talk about the classical condition—the eyeblink reflex. This is something that is cerebellar in origin. c. Later, when you talk about the vestibular systems, you will discuss the vestibule-ocular reflex (VOR). XXII. 1. Classical or Pavlovian Conditioning [S22] a. This first form of learning is really analogous to classical or Pavlovian conditioning. Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 5 of 6 b. When you present the dog with the conditioned stimulus (the bell), the dog does not do anything. However, when you pair the bell with a cheeseburger, you start getting the response (salivation). After a number of trials, you can just present the conditioned stimulus and the dog will begin to salivate. This is associative conditioning. c. In the cerebellum, you can get rabbits (the best mammalian system to study lesions in that are equivalent to humans) to blink when you have an air puff and you can pair that air puff with a tone and you can have the same sorts of learning. XXIII. 1. Eyelid Movements During a Classical Conditioning Experiment [S23] a. You can do the same kind of classical conditioning in the cerebellum as Pavlov did with the dogs. b. You can do the experiment with air puff and the tone. i. Before training, the tone does not do anything on its own, so this is the conditioned stimulus. ii. If you pair it, you see the upward deflection representing the eyes having closed. As you train the rabbit, you start seeing the tone giving the response on its own. Then if you look at the pairing stimuli, the conditioned and the unconditioned, you see that the eye blink is happening a little bit earlier. iii. After many trials, you get a response from the tone alone and you see a dual hump response when you give the conditioned stimulus with the unconditioned stimulus. iv. This is different from the results of the dog experiment because you are not just getting a response with the conditioned stimulus alone, but now the timing of the conditioned stimulus (the tone alone) is happening before the air puff. The cerebellum is saying that the tone is coming so you should close your eyes before the tone is coming. So the timing is really important for these conditioned cerebellar experiments. This is at the heart of what the cerebellum does. It sees a mistake, and then, it is not just going to correct that mistake, it has to know that mistake happened some time ago. That window of time is about 100 milliseconds. If something happens so that there is no association between the tone and the air puff, then the system does not learn that that has to be a respond you need to learn. If the tone had come much earlier, you would not get this learning because it would be outside the window of learning. The cerebellum is teaching that there is a timing between the tone and the air puff. Remember, if the tone occurred more than 100ms before or after the air puff, the rabbit probably would not have associated the two, so you would not get a conditioned response. In real life, you are probably learning much quicker than having to repeat the trials as many times to get the conditioned response. The air puff is represented by the climbing fibers, and the tone is the modality (the granular cells). XXIV. Temporal relationship of CS to US [S24] a. This slide basically tells you that timing is really important. If you move the tone and the air puff, you can change when the rabbit has the eye blink reflex occur. XXV. Eyeblink Conditioning Circuitry [S25] a. This is essentially the circuit that we were talking about. b. The tone is being represented by the mossy fibers and the granule cells. c. The climbing fibers are giving us the air puff correction. The output of that goes back to the eyelid muscles to close the eyes (the conditioned response). d. The interneurons act to shape the purkinje cell response, so if you have a problem with your interneurons of the cerebellum, the timing is also messed up. XXVI. Inactivation of Cerebellar Cortex Affect Timing of Eyeblink Response [S26] a. On a gross level, you can really see this by lesions. If you lesion the cerebellum, you don’t extinguish the response, but you really impair the timing of the response. b. In black is your closing of the eyelid. The tone is given in red. c. With the lesion, you get this mis-timed tone to response. d. Without the lesion, this (upper graph) is the normal timing of events. e. You can do this also pharmacologically by inhibiting the GABA receptors on the deep cerebellar nuclei. Remember that the purkinje cells release GABA. If you inhibit those postsynaptic neurons of the deep cerebellar nuclei that respond to the GABA, then you can really dull the response in the deep cerebellar nuclei. f. The bottom graph shows sequential trials of the animal closing its eyes. The blue is the control when you’ve inject picrotoxin which inhibits GABA receptors. You see that after that injection, you barely get any eye blink reflex response. That is a nice experiment that tells us that what is happening is happening in the deep cerebellar nuclei, out of the cerebellum. XXVII. Which Synapses are Modified During Eyeblink Conditioning? [S27] a. Imagine two different synapses. Somehow you have to stop the purkinje cell from firing. You can do this in two ways: i. You can decrease the granule cell to purkinje cell input; that is an excitatory input that drives the purkinje cell to release more GABA. Therefore, if you decrease this input, you would cause the purkinje cell to fire less. Neuro: 2:00 - 3:00 Scribe: Caitlin Cox Wednesday, February 18, 2009 Proof: Marjorie Hannon Dr. Wadiche Cerebellum Page 6 of 6 ii. You could excite the deep cerebellar nuclei directly by bypassing the integral function that the purkinje cells would provide. b. Either way, whether you produce LTD or long-term depression of the granule cell synapse, you would cause more firing of the deep cerebellar nuclei. Or if you produce LTP or long-term potentiation of these fibers, you would cause an increase in the firing of the deep cerebellar nuclei and closing of the eye. XXVIII. Evidence for the Anatomical Substrates of CS and US [S28] a. This is just a list of evidence of why we think the conditioned stimulus and the unconditioned stimulus are represented by where I told you there are. b. Lesions of the mossy fibers prevent learning. c. You can actually mechanically stimulate these areas and produce the same kind of effect as you would get with the tone or the air puff. d. The mossy fibers can be the tone and the olive can be the puff, and lesions of the olive can prevent learning. XXIX. Skip [S29-33] XXX. Cellular Mechanisms of Cerebellar LTD [S34] a. At the cellular level, the input of granule cells when there is coincident activation of the inferior olive, the climbing fiber and the parallel fiber. What you get is the output. This is monitoring the output of the purkinje cells, which depresses overtime. This depression of the output of the purkinje cells overtime is what lets the deep cerebellar nuclei fire more often to cause the reflex. XXXI. LTD of PF Synapses [S35] a. This is called long-term depression. b. What people know about this is that the climbing fiber leads to a huge atypical action potential and causes a big rise in calcium in the purkinje cell. c. At the same time, the parallel fiber activation will lead to the production of diacylglyceride (DAG) through metabotropic glutamate receptors (mGluRs). d. All these signals lead to internalization of AMPA receptors (glutamatergic receptors on purkinje cells). e. The same activity of granule cells leads to less receptor activation and so a smaller effect of the purkinje cells firing down to the deep cerebral nuclei. XXXII. Summary [S36] a. Cerebellum doesn’t just refine movement, it is almost predictive, and it is really corrective. As you are making mistakes, it is really trying to refine motor movements. This is happening at least at two sites: i. Within the cerebellar cortex, that will use all the purkinje cells and the power of integration. ii. You can also bypass the cerebellar cortex and go right to the deep cerebellar nuclei and excite those synapses. b. Think about why one might be better than the other? [end 49:23 min]