Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
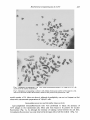
J. gen. Virol. (1981), 56, 421-430. Printed in Great Britain 421 Key words: varicella-zoster virus/thymidine kinase/biochemical transformation Biochemical Transformation of Mouse Cells by Variceila-Zoster Virus By K O I C H I TAKEO YAMANISHI,* YOSHINO MATSUNAGA, O G I N O AND P A T R I C I A L O P E T E G U I Department o f Virology, Research Institute f o r Microbial Diseases, Osaka University, Yamada-kami, Suita, Osaka 565, Japan (Accepted 27 M a y 1981) SUMMARY Mouse L cells lacking the enzyme thymidine kinase (Ltk-) were infected with varicella-zoster virus (VZV). Even though virus did not replicate in Ltk- cells, the presence of virus antigen could be observed by use of an anti-complement immunofluorescent technique at 4 h post-infection and the VZV-specific thymidine kinase could be detected in VZV-infected Ltk- cells. Ltk- cells were converted to a tk + phenotype (Ltk +) by infection with cell-associated VZV. Clones possessing the ability to grow in selective medium were isolated and cultured successfully for more than 20 passages. One of the clones grew very slowly, but other clones showed almost the same growth rate as that of the parental Ltk- cells. The chromosome analyses of Ltk- cells and transformed cells revealed that the isolated clones were of mouse origin. VZV-specific antigen could be detected in the nuclei of Ltk + cell clones by an immunofluorescent test, while tk activity was greatly enhanced in extracts prepared from transformed cells and its activity was neutralized by hyperimmune serum against VZV. INTRODUCTION Several enzymes are induced in cells infected by herpesviruses. Of these, thymidine kinase (tk) has been studied extensively. The tk induced by herpes simplex virus (HSV) has been shown to be encoded by the virus genome (Kit et al., 1975). Littlefield (1964) has shown that mouse L cells (Ltk +) can be selected from an L cell population lacking thymidine kinase (Ltk-) by the method of cell-cell fusion using a tissue culture medium that contains aminopterin, thymidine, hypoxanthine and glycine. Munyon et al. (1971) were the first to show that tk- cells could be stably transformed to a tk + phenotype by infection with u.v.-irradiated HSV. This process is known as biochemical transformation. Recently, it has been reported that fragments of HSV DNA can be introduced into the tkcells; the state of herpes virus DNA sequences in the transformed cells and the stability of these cells have also been reported in detail (Bacchetti & Graham, 1977; Maitland & McDougall, 1977; Wigler et al., 19.77; Minson et al., 1978; Pellicer et al., 1978; Leiden et al., 1980). Two reports have demonstrated that varicella-zoster virus (VZV) is also capable of inducing a virus-specific tk (Dobersen et al., 1976; Ogino et al., 1976). In the present study, we describe the biochemical transformation of Ltk- cells by VZV and some of the properties of biochemically transformed cells. METHODS Cells. An Ltk- cell line was obtained from Dr Okada in our Institute (Murayama & Okada, 1970). These cells were serially cultivated in Roux bottles in growth medium (medium 199 and Eagle's minimum essential medium supplemented with 100 ~tg/ml streptomycin, 100 Downloaded from www.microbiologyresearch.org by 0022-1317/81/0000-4599 $02.00 © 1981 SGM IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 422 K. Y A M A N I S H I AND O T H E R S units/ml penicillin, 10% calf serum and N a H C O 3) containing 50 /tg/ml 5bromodeoxyuridine. Human embryonic fibroblast (HEF) cells were grown in Roux bottles in growth medium only. Cells were subcultured at 6 or 7 day intervals, and cultures from the 10th to the 20th passage were used for the experiments. Viruses. Three strains of VZV (Kawaguchi, Tanaka and Oka strains), originally isolated by Dr Takahashi in our laboratory, were passaged 10, 5 and 11 times respectively in HEF cells. Cell-free virus was obtained by sonication and titrated on HEF cells as previously reported (Yamanishi et al., 1980). The appropriate dilution of VZV (0-2 ml/plate) or infected cells (for infectious centre assay) was plated on confluent monolayers of HEF cells grown in 60 mm glass plates. After an adsorption period of 1 h at 37 °C, cultures were flooded with maintenance medium (this was the growth medium supplemented with 3 % calf serum instead of 10% calf serum). At the fifth day of incubation in a humidified CO2 incubator at 37 °C, cultures were fixed in 5 % formalin and stained with 0 . 1 % methylene blue. The plaques were counted using a dissecting microscope at a × 20 magnification. Chromosome analyses. Forty-eight h cultures of HEF, Ltk- cells and the transformed cell lines L(K)cl 1 (passaged 35 times) and L(O)cl 3 (passaged 48 times), were used for the chromosome analysis. Cells were arrested at metaphase by addition of colcemid to a final concentration of 0.1 Ftg/ml. After trypsinization, cell harvesting was standard (Moorhead et al., 1960), with hypotonic pretreatment in 0.075 M-KC1 for 15 min followed by three fixes in methanol:acetic acid (3:1). The final suspension was dropped on to cold, wet slides and flame-dried. Slides were stained with Giemsa 1 : 50 in phosphate buffer pH 6.8. At least 50 metaphases were analysed under a microscope in order to establish the modal chromosome number of each cell type and to look for any marker chromosomes that would allow the characterization of the cells. Immunofluorescence test and immune serum. In order to detect VZV antigen in cells infected with VZV or in biochemically transformed cells, the anti-complement immunofluorescence method was applied. Ltk- cells grown on round glass coverslips and infected with VZV, were washed with phosphate-buffered saline (PBS) three times, air-dried and fixed in a mixture of acetone and methanol (1:1) for 10 min at - 2 0 °C. Biochemically transformed cells grown on the coverslips were also treated as described above. Fixed cells were kept at - 8 0 °C until staining. VZV hyperimmune serum was prepared in green monkeys as described previously (Yamanishi et al., 1980). Fixed cells were covered with about 0.1 ml of a mixture containing equal proportions of antiserum from green monkeys and fresh, human serum diluted 1 : 10 as the complement source (anti-VZV antibody-negative serum), and incubated at 37 °C for 30 min in a humidified chamber. Cells were then washed with PBS three times, covered with anti-human complement C3 goat serum conjugated with fluorescein isothiocyanate (Hyland, Ca., U.S.A.) and incubated at 37 °C for another 30 min. After this, stained coverslip preparations were washed with PBS, mounted in a 1:9 solution of PBS and glycerin, and observed in an Olympus fluorescence microscope. Preparation of cell extracts. H E F cells were infected with VZV (Kawaguchi strain) at an m.o.i, of 0.1 and incubated at 37 °C for 72 h. Ltk- cells were infected as described above and incubated for 18 h at 37 °C. Infected cultures, mock-infected cultures and biochemically transformed Ltk + cell cultures were harvested for the tk activity assay as described previously (Ogino et al., 1976). Monolayers were washed three times with PBS, scraped off with a rubber policeman and pelleted at 200 g for 15 min at 4 °C. They were then suspended in 0.5 ml 0.15 M-KC1 in 0.05 M-tris buffer pH 8, containing 3 mM-2-mercaptoethanol. The cell suspensions were disrupted by ultrasonic vibration. The sonicated preparations were centrifuged at 20 000 rev/min for 60 min in a Hitachi 55-p centrifuge (rotor no. 40). The supernatant fractions were used as the Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 Biochemical transformation by V Z V 423 sources of enzyme extracts. Protein determinations of cell extracts were carried out by the method of Lowry et al. (1951). Thymidine kinase activity assay. The assay of tk activity was performed as described previously (Ogino et al., 1976). The assay mixture contained the following final concentrations of solutions: 0.2/tCi [14C]TdR (56.6 mCi/mmol, Amersham), 5 mM-ATP, 5 mM-MgC12, cell extracts (30 to 100 pg protein) and 0-05 M-tris buffer pH 8 in a final vol. of 0-25 ml. The reaction mixture was incubated at 38 °C for 15 min and the reaction was terminated by immersing the mixtures in a boiling water bath for 2 min. A 0.25 ml vol. of the reaction mixture was spotted on DEAE-cellulose discs. These were washed three times with 1 mM-ammonium formate buffer, once with distilled water and once with 95 % ethanol, and then dried and the radioactivity determined in a liquid scintillation counter. For serological neutralization tests, cell extracts were preincubated for 2 h at 4 °C with an equal volume of preimmune or immune serum, which was prepared as described previously (Yamanishi et al., 1980) and dialysed for 2 days against 0.05 M-tris buffer pH 8 and the tk activity was assayed as described above. RESULTS Infection of L t k - cells with V Z V Monolayers of Ltk- cells grown in 60 mm plastic plates were infected with VZV at an m.o.i, of 0.1. At 1, 2 or 3 days after infection, cells were trypsinized and then suspended in 1 ml maintenance medium. Serial dilutions of suspended cells were plated on monolayers of HEF cells grown in 60 mm glass plates for infectious centre assay. No cytopathic effect (c.p.e.) was observed in infected cultures of Ltk- cells and no infectious centres were detected at any time. Monolayers of Ltk- cells on round glass coverslips were infected at an m.o.i, of 0.1. After a 1 h adsorption period at 37 °C, cells were washed with PBS and maintenance medium was added. Cultures were harvested at 4 h and 24 h post-infection and immunofluorescent tests were performed as described above. Nuclear antigen was detected in 5 to 10% of cells infected with VZV at 4 h post-infection (Fig. 1 b), but no specific immunofluorescence was observed at 24 h post-infection (Fig. 1 c). Tk activity of VZV-infected Ltk- cells was determined and compared with the tk activity of mock-infected L t k - cells (Table 1). The activity of the infected cells was approx, threefold greater than that of the mock-infected cells. The anti-VZV serum reduced the activity of the preparation from the infected cells by 71.5 %, Biochemical transformation with VZ V The method of cell-to-cell-infection was applied to infect Ltk- cells with VZV. The H E F cells grown in Roux bottles were infected with the Kawaguchi, Oka or Tanaka strains of VZV at an m.o.i, of approx. 0.01. Two days later, infected cells were trypsinized, suspended in maintenance medium and plated on monolayers of uninfected H E F cells grown in Roux bottles. Three days after infection, when more than 90% of the cell sheet showed the typical c.p.e., cells were trypsinized and suspended in growth medium. Sparse monolayers of Ltkcells (5 x 105 cells/plate) grown in 100 mm plastic plates were inoculated with VZV-infected HEF cells (5 x 105 cells/plate) suspended in 15 ml of growth medium. The inoculated cultures were then incubated for 48 h to allow gene establishment and expression. The medium was removed and replaced with selective medium (growth medium containing 6 × 10 -~ M-aminopterin, 5 x 10 -5 M-guanosine, 5 x 10 -5 M-adenosine, 10 -4 M-glycine, 5 x 10 -3 M-glutamine and 1.6 x 10 -5 M-thymidine). Five days later cells were fed with fresh selective medium and, thereafter, every 7 days. The dishes were incubated for a total of 4 to 20 weeks Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 424 K. YAMANISHI AND OTHERS Fig. I. Immunofluorescence photomicrography of VZV infected Ltk- cells. Ltk cells grown on coverslips were infected with the Kawaguchi strain of VZV at an m.o.i, of 0.1. Cultures were harvested at either 4 h or 24 h post-infection and stained by the anti-complement immunofluorescent method. (a) Non-infected Ltk- cells; (b) Ltk- cells infected and harvested at 4 h post-infection (note the prominent nuclear fluorescence at this time); (c) Ltk cells infected and harvested at 24 h post-infection. Table 1. Serological neutralization of thymidine kinase from VZV-infeeted or bioehemically transformed cells* Tk activity (ct/min) Extract HEF HEF VZV-infected LtkLtk- VZV-infected L(K)cl 1 L(O)cl 3 22 173 301853 1406 4375 26686 25 925 c Residual activity (%) ~" ~ Preimmune serum Immune serum 95.2 117.0 93.9 93.6 95.4 103-0 107.0 8.1 91.2 28.5 1.8 2-9 * Enzyme neutralization tests were carried out as described in Methods. Tk activity was shown as ct/min/mg protein of the extracts at 38 °C for 15 min by calculation. The activity of Ltk- cells was assayed by an incubation period of 60 min because of its low activity. at 37 °C in a humidified 5 % C O 2 - 9 5 o~ air atmosphere. Small colonies were visible under the microscope by 3 weeks, and by 5 to 6 weeks after infection 1 to 4 colonies per dish were observed. These colonies were 2 mm or more in diam. (Table 2). Dense foci, which consisted of epithelioid cells and giant cells, were observed. Neither the control cells (Ltk monolayers not inoculated with infected cells, but cultured in selective medium) nor Ltk- cells infected with cell-free virus at an m.o.i, of 0.1, showed the formation of Ltk + cell colonies when assayed according to the method described above. Some properties of the transformed cells About 50 colonies were isolated and have been grown in selective medium for more than 6 months and over 20 passages. The cell growth rate is shown in Fig. 2. Almost all clones had about the same growth rate as that of the parental Ltk- cells. However, one clone which was derived from the cultures infected with the Kawaguchi strain, L(K)cl 1, grew more slowly than the parental cells. Another clone, which was derived from the cultures infected with Oka strain, L(O)cl 3, was chosen as representative of the clones having the same growth rate as that of the parental cells (Fig. 2). Although clones from cultures infected with the Tanaka Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 Biochemical transformation by V Z V 425 T a b l e 2. Ltk- to Ltk + transformation by V Z V infection* Infection Mock-infected Kawaguchi Tanaka Oka No. of foci/100 mm plate ~ ~" ~ Average 0, 0, 0, 0, 0, 0 0 4, 3, 2, 3 3.0 1, 1, 2, 1 1.3 2, 3, 4, 2 2.8 * Data are results of a single experiment representative of a total of three experiments for each virus. Ltk- cells (5 × 105 cells) were infected with cell-associated VZV. Numbers of foci were counted 6 weeks after infection. 10 i i 2 Time of incubation (days) 4 Z z d 5 ¸ .2 e¢ ! 0 Fig. 2. Growth curves of Ltk- cells and VZV-transformed L(K)cl 1 and L(O)cl 3 cells. Cells were seeded in 60 mm plastic dishes at a concentration of 2 x 105 cells/dish, and incubated in a CO 2 incubator. No, Cell number at time 0; N 1, cell number at given time. Ltk- cells (A), L(K)cl 1 (0), I40)cl 3 (m). Fig. 3. Ltk + cells biochemically transformed with VZV. Transformed cells were subcultured 20 times in selective medium and stained with haematoxylin-eosin. (a) Control (Ltk-) cells; (b) L(K)cl 1 cells; (c) L(O)cl 3 cells. strain were also o b t a i n e d (Table 2), they were not used for the i m m u n o f l u o r e s c e n c e and t h y m i d i n e kinase activity tests. L ( K ) c l 1 cells stained with h a e m a t o x y l i n - e o s i n exhibited a variety o f cell types t h a t consisted o f m u l t i n u c l e a r cells, and small and large cells (Fig. 3 b). H o w e v e r , L ( O ) c l 3 cells had a similar m o r p h o l o g y to the p a r e n t a l cells (Fig. 3 a, c). Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 426 K. Y A M A N I S H I AND O T H E R S ? Fig. 4. Metaphase corresponding to the HEF cells used throughout the present study. Modal chromosomenumber is 46 (46 XX). An attempt was made to isolate virus from the transformed cells, but no virus could be isolated by the method of co-cultivation with H E F cells. Transformed cells were superinfected with the Kawaguchi strain, but no c.p.e, was observed which implied that the virus could not replicate in these ceils. Chromosome analyses The chromosome analysis of the HEF cells revealed that these cells were of female origin with a normal modal chromosome number of 46 (46 XX), as shown in Fig. 4. The modal chromosome number of Ltk- cells was 54 (range 46 to 56). These cells presented multiple structural rearrangements, and both hyperdiploid and hypodiploid cells were frequently observed. This cell line differs from normal mouse L cells in that besides the normal acrocentric chromosomes (Levan et al., 1962), many metacentric chromosomes are also present. Fig. 5 shows a metaphase of Ltk- cells. Although this cell has only 50 chromosomes, it was chosen because marker chromosomes are clearly seen. In spite of the multiple rearrangements that take place in Ltk- cells, two large dicentric chromosomes were always present. One acrocentric chromosome with large satellites, a large submetacentric chromosome and a medium-sized submetacentric chromosome with a secondary constriction are markers which are not present in all cells. The biochemically transformed cells were shown to be of mouse origin by cytogenetic studies. Fig. 6 shows a metaphase spread from an L(K)cl 1 cell. These cloned cells presented a great variation in the number of chromosomes (modal chromosome number of 50, range 42 to 54) when compared with Ltk- cells and L(O)cl 3 cells. The markers described for Ltkcells were also observed in the transformed cells, both L(K)cl 1 and L(O)cl 3 cells, and their occurrence varied in the same way as in the original cells, i.e. at least the two dicentric chromosomes were always present, but the other marker chromosomes were observed less frequently. Chromosome studies on L(O)cl 3 cells also show that they were derived from the Ltkparent, i.e. chromosomes correspond with those of Ltk- cells, presence of markers and a Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 Biochemical transformation by V Z V 427 Fig. 5. Metaphase corresponding to Ltk cells. Modal chromosome number is 54 (range 46 to 56). The arrows show the marker chromosomes. Fig. 6. Metaphase corresponding to L(K)cl 1 cells. Modal chromosome number is 50 (range 42 to 54). The arrows show the marker chromosomes, which are also present in the parental Ltk cells. m o d a l number of 54 (data not shown), although hypodiploidy was not so frequent as that observed in chromosome preparations of L(K)cl 1 cells. Immunofluorescent test and thymidine kinase activity Anti-complement immunofluorescent tests were performed to detect the presence o f V Z V antigen in the transformed cells. M o s t cells were found to be positive for nuclear fluorescence (Fig. 7 a, b), although the intensity of staining varied between the cell lines. C y t o p l a s m i c fluorescence was only observed in the large cells of cell line L(K)cl I (Fig. 7 a). Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 428 K. Y A M A N I S H I AND OTHERS Fig. 7. Immunofluorescence photomicrography of VZV-transformed Ltk + cells (20th passage). Stained by the anti-complement immunofluorescent method. (a) L(K)cl 1 cells; (b) L(O)cl 3 cells. Note only nuclear fluorescence in (b) and both nuclear and cytoplasmic fluorescence in (a). The two transformed cell lines, L(K)cl 1 and L(O)cl 3, were tested for tk activity, and were found to have about 25 times more tk activity than the Ltk cells. Thymidine kinase neutralization tests were carried out on VZV-infected HEF cells and the biochemically transformed cells using immune serum (Table 1). The anti-VZV serum reduced the tk activity from the preparations in H E F cells infected with VZV by 91-9%. The same serum also reduced the tk activity in extracts from the transformed cell lines L(K)cl 1 and L(O)cl 3, 98.2% and 97.1% respectively. DISCUSSION The present study showed that the thymidine kinase gene of VZV could be introduced into Ltk- cells. When mouse tk- cells are infected with HSV, the frequency of transformation of tk- into the tk + phenotype varies from 10 -3 to 10-6 depending on the method of virus inactivation or virus strains employed (Rapp & Turner, 1978). In the experiments described here, the frequency of transformation induced by cell-associated VZV was about 10-6 (Table 2). It would be hard to compare the frequency of transformation mediated by HSV with the one mediated by VZV, because cell-free virus was used for the transformation tests by HSV, and cell-associated virus was used in our experiment with VZV. So far, we have not found any colonies when cell-free VZV has been used for the transformation assay. This might be due to the low titre of VZV. At present, experiments are under way in order to obtain VZV of a high titre to repeat the transformation assay. Since HEF cells infected with VZV were inoculated on to Ltk- mouse cells in the transformation tests, H E F cells, which were not infected with VZV, might survive in selective medium as well as Ltk + mouse cells. Thus, karyotype analyses of the isolated clones were performed in order to determine if they were really derived from Ltk- cells. The results of these analyses showed no differences between the cloned cells and those of the parental Ltk- cells except for the modal number of L(K)cl 1 cells. Although the chromosome analyses clearly show that the transformed cells are of mouse origin, chromosomes of human origin in mouse-human hybrid cells are lost on serial passage (Weiss & Green, 1967), and so the presence of metacentric and submetacentric chromosomes does not rule out the possibility that these cloned cells are hybrids. To clarify this point chromosome preparations were stained with a combined quinacrine and 33258 Hoechst technique according to a method described by Yoshida et al. (1975). This demonstrated that all chromosomes in the cloned cells were of mouse origin (data not shown). Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 Biochemical transformation by V Z V 429 Transformed cells were also superinfected with VZV to further confirm that clones were not of HEF cell origin. No c.p.e, appeared and there was no evidence of virus growth. From these data, it can be stated that cell clones picked up from the colonies are undoubtedly of Ltkmouse origin. Although clones of human origin were picked up very often, it was relatively easy to distinguish Ltk + mouse clones from HEF cells by morphological criteria. The clones that arose in selective medium differed from parental Ltk- cells in the following properties: (i) one of the clones grew more slowly than parental cells; (ii) differences in size and shape were recorded (Fig. 3); (iii) they contained nuclear antigens which reacted with hyperimmune serum against VZV (Fig. 7); (iv) tk activity in the transformed cells was neutralized by the immune serum (Table 1). For the cell lines examined the growth rate of L(O)cl 3 cells was almost the same as that of the parental cells, while L(K)cl 1 cells grew more slowly. Furthermore, nuclear antigens were detected in all transformed cell lines, but cytoplasmic antigens were only observed in large cells of L(K)cl 1 cell line (Fig. 7). As shown in Fig. 7, almost all cells had nuclear antigens reacting to hyperimmune serum and they have remained positive for more than 6 months after transformation. Cytoplasmic antigens have been observed in morphologically transformed cells (Duff & Rapp, 1971, 1973; Takahashi & Yamanishi, 1974) and, cytoplasmic or nuclear antigens were detected in biochemically transformed cells (Chadha & Munyon, 1975; Kurchak et al., 1977; Rapp & Turner, 1978). The different location of antigens in HSV- and VZV-transformed cells is interesting, but the significance at present is unclear. However, when we infected Ltk- cells with cell-free virus, only nuclear antigen was detected at 4 h post-infection (Fig. l b) and no virus was rescued from the cultures as mentioned above. Richards et al. (1979) have analysed electrophoretically the DNAs from VZV isolates including the Oka and Kawaguchi strains and reported minor differences between different VZV DNAs. In the case of tk transformation by HSV, it has been shown that a wide variety of non-tk gene virus DNA sequences can be stably incorporated into the transformed cells (Bastow et al., 1980, Leiden et al., 1980). It therefore might be possible to postulate that the VZV DNA fragments may exist in the transformed cells and the antigens expressed in these cells would correspond to the early antigens formed in Ltk- cells at 4 h post-infection with VZV, and that these antigens would include the tk of VZV. Even though these clones have the VZV-specific thymidine kinase in common, it is probable that other segments of VZV DNA were also introduced into the cells, and that they would be different in the various clones. Experiments for the analysis of these antigens are now in progress. We wish to thank Professor M. Takahashi for his support throughout these investigations and Dr J. Furuyama of Hyogo College of Medicine for helpful discussion concerning chromosomal analyses. REFERENCES BACCHETTI, S. & GRAHAM, F. L. (1977). Transfer of the gene for thymidine kinase to thymidine kinase-deficient human cells by purified herpes simplex viral DNA. Proceedings of the National Academy of Sciences of the United States of A merica 74, 1590-1594. BASTOW, K. V., DARBY, G., WILDY, P. & MINSON, A. C. (1980). Properties of cells carrying the herpes simplex virus type 2 thymidine kinase-negative phenotype. Journal of Virology 36, 746-755. CHADHA, K. C. & MUNYON, W. (1975). Presence of herpes simplex virus related antigens in transformed L cells. Journal of Virology 15, 1475-1486. DOBERSEN, M., JERKOFSKY, M. & GREER, S. (1976). Enzymatic basis for the selective inhibition of varicella-zoster virus by 5-halogenated analogues of deoxycytidine. Journal of Virology 20, 467-486. DUFF, a. & RAPF, F. (1971). Properties of hamster embryo fibroblasts transformed in vitro after exposure to ultra-violet irradiated herpes simplex virus type 2. Journal of Virology 8, 469-477. DUFF, R. & RAPF, F. (1973). Oncogenic transformation of hamster embryo cells after exposure to inactivated herpes simplex virus type 1. Journal of Virology 12, 209-217. KIT, S., LEUNG, W., JORGENSEN, G. N., TRKULA, D. & DUBBS, D. R. (1975). Viral-induced thymidine kinase isozymes. Progress in Medical Virology 21, 13-34. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55 430 K. YAMANISHI AND OTHERS KURCHAK, M., DUBBS, D. R. & KIT, S. (1977). Detection of herpes simplex virus-related antigens in the nuclei and cytoplasm of biochemically transformed cells with peroxidase antiperoxidase immunological staining and indirect immunofluorescence. International Journal of Cancer 20, 371-380. LEIDEN, J. M., FRENKEL, N. & RAPP, F. (1980). Identification of the herpes simplex virus D N A sequences present in six herpes simplex virus thymidine kinase-transformed mouse cell lines. Journal of Virology 33, 272-285. LEVAN, a., HSU, T. C. & STICH, n. F. (1962). The idiogram of the mouse. Hereditas 48, 676-687. LITTLEFIELD, J. W. (1964). Selection of hybrids from mating fibroblasts in vitro and their presumed recombinants. Science 145, 709-710. LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193, 265-275. MAITLAND, N. J. & McDOUGALL,J. K. (1977). Biochemical transformation of mouse cells by fragments of herpes simplex virus DNA. Cell 11,233-241. MINSON, A. C., WILDY, P., BUCHAN, A. & DARBY, G. (1978). Introduction of the herpes simplex virus thymidine kinase gene into mouse cells using virus D N A or transformed cell D N A . Cell 13, 581-587, MOORHEAD, P. S., NOWELL, P. C., MELLMAN, W. J., BATTIPS, D. M. & HUNGERFORD, D. A. (1960). Chromosome preparation of leukocytes cultured from human peripheral blood. Experimental Cell Research 20, 613-616. MUNYON, W. M., KRAISELBURD, E., DAVIS, O. & MANN, J. (1971). Transfer of thymidine kinase to thymidine kinaseless L cells by infection with ultraviolet irradiated herpes simplex virus. Journal of Virology 7, 813-820. MURAYAMA,F. & OKADA, V. (1970). Efficiency of hybrid cell formation from heterokaryons fused by HVJ. Biken Journal 13, 1-9. OGINO, T., OTSUKA, T. & TAKAHASHI,M. (1976). Induction of deoxypyrimidine kinase activity in human embryonic lung cells infected with varicella-zoster virus. Journal of Virology 2 l, 1232-1235. PELLICER, A., W1GLER, M., AXEL, a. & S1LVERSTEIN, S. (1978). The transfer and stable integration of the HSV thymidine kinase gene into mouse cells. Celt 14, 133-141. RAm', V. & TURNER, N. (1978). Biochemical transformation of different methods by herpes simplex virus type 1 and 2: comparison of different methods for inactivation of viruses. Archives of Virology 56, 77-87. RICHARDS, J. C., HYMAN, R. W. & RAPP, ~. (1979). Analysis of the D N A s from seven varicella-zoster virus isolates. Journal of Virology 32, 812-821. TAI~AHASm, M. & YAMANISHI, K. (1974). Transformation of hamster embryo and human embryo cells by temperature sensitive mutants of herpes simplex virus type 2. Virology 61, 306-311. WEISS, M. C. & GREEN, H. (1967). Human-mouse hybrid cell lines containing partial complements of human chromosomes and functioning h u m a n genes. Proceedings of the National Academy of Sciences of the United States of America 58, 1104-1111. WI(3LER, M., SILVERSTEIN, S., LEE, L., PELLICER, A., CHENG, Y. & AXEL, R. (1977). Transfer of purified herpes virus thymidine kinase gene to cultured mouse cells. Cell 11, 223 232. YAMANISHI, K., MATSUNAGA, Y., OGINO, T., TAKAHASHI, M. & TAKAMIZAWA, A. (1980). Virus replication and localization of varicella-zoster virus antigens in human embryonic fibroblast cells infected with cell-free virus. Infection and Immunity 28, 536-541. YOSHIDA, M. C., IKEUCHI, T. & SASAKI, M. (1975). Differential staining of parental chromosomes in interspecific cell hybrids with a combined quanacrine and 33258 Hoechst technique. Proceedings of the Japan Academy 51, 184-187. (Received 4 February 1981) Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 04 Aug 2017 00:00:55